
Abstract
We studied the impacts of site, stand and tree variables on the diameter growth of beech trees (Fagus sylvatica L.) on carbonate bedrock and examined to what extent the regional diameter growth model can be used at the forest type level. Based on 12,193 permanent sampling plots (500 m2 each) with 94,770 beech trees, we first developed a linear mixed-effect model of the periodic diameter increment at the regional level (Dinaric region, Slovenia, 1.7 thousand km2). Subsequently, we parametrized the model for five forest types within the region (submontane, thermophilous, montane, upper montane and subalpine) and used a homogeneity-of-slopes model to test whether the covariates have different effects in the five forest types. The regional model suggested the positive impact of tree diameter (nonlinear), tree diameter diversity, mean diurnal temperature range and mean annual temperature and the negative impact of basal area, proportion of beech, inclination, rockiness and annual amount of precipitation. Stand basal area and the proportion of beech contributed > 50% of the total explained diameter increment variability, followed by tree diameter (44%), topographic (3%) and climatic variables (< 2%). The regional model was well transferable to forest types; the only variable with a significantly different effect in forest types was tree diameter. However, models at the forest type level differed with respect to the slopes and significance of several predictors, wherein coefficients for some predictors were even of opposite sign. Not all predictors from the regional model were included in the forest type models if predictor selection and model parameterization were performed independently for each forest type. Our study suggests that some growth characteristics of beech can be detected at the regional level only, while analyses at the forest type level can reveal significant differences in beech growth response to tree, stand and environmental variables.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
European beech (Fagus sylvatica; hereafter beech) is a widely distributed tree species in Europe with high economic and ecological value for Central European (CE) forestry (Ellenberg and Leuschner 2010). Due to its broad ecological amplitude, it dominates in late successional forests. It is a strongly competitive tree species on various types of bedrock and soil conditions and in a broad elevation range. In the northern part of its distribution, beech predominantly grows in colline, submontane or lower montane vegetation belts (Böhn et al. 2004; Bolte et al. 2007), while in the southern part of the distribution, it may also dominate in the upper-mountain and subalpine vegetation belt, even at the upper timber line (e.g. Mayer 1974; Dakskobler 2008; Bončina 2012). During past centuries, the proportion of beech in CE forests was significantly reduced, mainly because Norway spruce was promoted due to its higher profitability, and past management regimes and land use (e.g. forest pasture) favoured early successional tree species. Recently, the share of beech has increased due to the broad acceptance of nature-based forestry and the recognition that it is a more appropriate tree species for adapting forests to climate change due to its stability and resilience (Hanewinkel et al. 2010; Pretzsch et al. 2020). The number of studies on beech growth has increased for the same reason.
The diameter growth of individual trees is influenced by tree, stand and site factors. Among tree variables, diameter at breast height (D) or its transformations (e.g. Tenzin et al. 2017; Hu et al. 2021) and other tree characteristics indicating competitive status, such as crown size, vigour, social status and distance to neighbouring trees (Wykoff et al. 1982; Pretzsch 2009), have been included in analyses of the diameter growth of individual trees.
Stand factors associated with the diameter growth of trees usually encompass variables related to stand density, structural diversity and tree species mixture (Forrester 2019). Stand density is most often described by stand basal area, the number of trees or stand density index. They indicate competition conditions for a single observed tree. Stand basal area or the basal area of larger trees than the observed tree has often been recognized as predictors in individual tree growth models (e.g. Bueno and Bevilacqua 2010; Lhotka and Loewenstein 2011).
Structural diversity may affect the growth of the individual trees in a stand (Pretzsch 1997; Liang et al. 2007; Lei et al. 2009). Structural diversity describes how the number of trees, stand volume or basal area is distributed between tree size classes, which is often measured by structural indices such as the Gini coefficient.
Tree species mixture can noticeably modify the growth pattern of individual trees. The effect of tree species mixture includes competition between species, facilitation and competitive reduction (Forrester and Bauhus 2016). Several studies have indicated the better growth of beech in mixed forests (e.g. Pretzsch et al. 2021). Beech may benefit from spatial niche complementarity and light interception when mixed with other species such as Scots pine (Pinus sylvestris) (Gonzales de Andres et al. 2017), Norway spruce (Picea abies) (Pretzsch et al. 2014), Douglas fir (Pseudotsuga menziesii) (Thurm and Pretzsch 2016) or silver fir (Abies alba) (Bosela et al. 2015b). The admixture of beech can contribute to the reduction of drought effects (Forrester 2015; Jourdan et al. 2019). Goisser et al. (2016) pointed out the importance of the temporal complementarity of beech with conifers. It seems that the complementary effect increases with stand age (Zeller and Pretzsch 2019).
Site factors include climate, topography and soil. The number of studies examining the influence of climatic variables on beech growth has been increasing. The diameter growth of beech at high elevations is controlled by temperature, while at low elevations it is predominantly controlled by summer rainfall and negatively affected by high summer temperatures (e.g. di Filippo et al. 2007; Babst et al. 2013). Beech growth is negatively influenced even by high summer temperature in the previous year (Scharnweber et al. 2011; Zimmermann et al. 2015). Beech has been found to be highly sensitive to warm and dry periods (Friedrichs et al. 2009; Betsch et al. 2011), especially in the colline and partly also in the submontane belt. Drought stress is less likely at higher elevations (Trotsiuk et al. 2020).
Studies of the long-term dynamics of beech growth have yielded different results. Some studies have reported a decline in the growth of beech stands since the 1980s (e.g. Charru et al. 2010; Zimmermann et al. 2015; del Castillo et al. 2022), mainly due to drought effects caused by climate change (Jump et al. 2006), and predicted a severe decline in beech growth in the next decades (del Castillo et al. 2022), especially on the southern edge of its distribution. Other studies have found no evidence of a recent growth decline, while others have even found a recent increase (e.g. Friedrichs et al. 2009; Tegel et al. 2014) or an increase since the late nineteenth century (Pretzsch et al. 2014) or since 1960 (Bosela et al. 2016). However, it is evident that in areas with repeated drought years, the growth of beech decreases (Bosela et al. 2015a, b).
The impact of topographic and soil variables on beech growth has been much less studied. Soil influences the growth of beech (Vospernik 2021); it seems that beech growth depends mainly on soil water holding capacity (Kirchen et al. 2017) and nutrient deposition (Etzold et al. 2020). Among topographic variables, elevation seems to be the most frequently used in beech growth studies (e.g. Di Filippo et al. 2007; Dulamsuren et al. 2016), while aspect, inclination, rockiness and others are less frequently considered. In temperate forests, elevation is an important indicator of climatic conditions (e.g. Bosela et al. 2015a, b) since it highly correlates with mean annual temperature and annual amount of precipitation. Areas with a strong elevational gradient are therefore suitable for analysing global change processes generated by climate change.
Findings on the growth pattern of beech in specific parts of CE tend to be generalized to broader forestry regions such as CE forests (Mina et al. 2018), or mountain forests (Hilmers et al. 2019) or the area at the edge of beech’s distribution range (Pretzsch et al. 2021). Therefore, questions arise as to what extent the findings can be generalized across a wide set of diverse beech forest types within broader regions (Wykoff et al. 1982; Forrester 2019). Beech forest types differ in growth factors. Although some factors are included in tree growth modelling on a regional spatial scale, the question remains whether the impacts of other predictors of the growth model are forest type dependent. Several studies have shown an elevation-dependent growth response of tree species to climatic variables (Hartl-Meier et al. 2014). Similarly, the growth response of a tree species under various environmental factors may be specific for a soil class (Seltmann et al. 2021). The forest site type represents a spatial framework with specific ecological conditions (Roberts 2015) and with probably specific relationships between variables influencing tree and stand growth (Trouillier et al. 2020). Forest type can serve as a surrogate for a set of site and vegetation variables and their interrelations, for which spatially explicit data are not available. Forest type classification is usually done at the landscape or regional spatial scale. Different forest site classifications have been used in European countries (Cajander 1949; Zlatník 1976; Ray 2001); in Central Europe, the Braun–Blanquet method of forest community classification based on potential natural vegetation has been broadly integrated into forest science and forest management. It is assumed that stand dynamics, the natural disturbance regime, the growth of trees and complementarity between tree species are specific for a forest type (Bončina et al. 2021). Despite the known importance of forest types in forest management and planning, they are mainly ignored in modelling tree and stand growth. In growth modelling, forest types can be considered in different ways: indirectly through site and stand variables, which differ between forest types, or directly by including forest types in growth models as a dummy variable. The latter may improve the growth model at a broader spatial scale, but the parameter estimates remain the same for all forest types. An alternative option is to model growth separately for single forest types, which can be done either with the same set of variables so that the comparison of the parameter estimates for the same variable is possible, or with a broader set of potential explanatory variables, likely leading to the inclusion of different variables.
The main objectives of our study were (i) to determine which site, stand and tree variables are the main predictors of the diameter growth of beech on carbonate at the regional level; (ii) to examine to what extent the regional growth model can be used at the forest type level ranging from the submontane to subalpine forests, and whether there are differences in the effect of variables between forest types; and (iii) to parametrize diameter growth models separately for each forest type in the region and compare the differences to the regional model. We hypothesized that the relationship between explanatory variables and diameter growth is forest type specific and that this is reflected in significantly different effects of predictors in the models.
Materials and methods
Study area
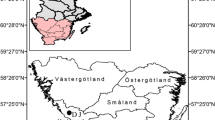
A forest region in the southwestern part of Slovenia covering around 1.7 thousand km2 was used as the study area (Appendix 1). The climate is interferential, i.e. characterized by a combination of temperate oceanic and continental climate with geographical variations mainly due to diverse topographic conditions. The mean annual temperature in the region amounts to 7.6 °C, and the mean annual precipitation amounts to 1860 mm. The prevalent parent materials are limestone and dolomite on which Cambisols and Leptosols of different depths evolved. Close-to-nature forestry based on natural regeneration has been practiced for decades. Planting has been applied only exceptionally, mainly for the conversion of shrub vegetation into more productive forests. The mean stand basal area amounts to 28.2 m2 ha−1, with the main tree species being beech (40%), Norway spruce (25%), silver fir (20%) and valuable broadleaves (7%). The irregular shelterwood system and (in a minor part of the area) selection system are applied.
Five main beech forest types along an elevational gradient from the submontane to subalpine vegetation zone were selected for the analyses. Their basic characteristics are described in Appendix 1; hereafter, they are referred to as submontane, thermophilous, montane, upper montane and subalpine beech forests. Submontane beech forests are characterized by a high mixture of other broadleaves, such as sessile oak (Quercus petraea), while Norway spruce was favoured by planting. The irregular and regular shelterwood silvicultural systems are mainly applied. Thermophilous forests are characterized by warm aspects, steeper terrain, shallow soil and the presence of thermophilous broadleaves such as hop hornbeam (Ostrya carpinifolia), manna ash (Fraxinus ornus), field maple (Acer campestre) and common whitebeam (Sorbus aria). Production potential is lower than that in submontane forests. Montane beech forests are characterized by a relatively high proportion of silver fir. Norway spruce is a natural tree species in this forest type; however, its proportion was increased by forest management. The irregular shelterwood system and in some areas the selection system are mainly applied. Upper montane forests are characterized by the domination of beech in the tree species composition and a relatively high proportion of sycamore (Acer pseudoplatanus) in forest stands. Stand productivity is lower than that in montane forests. The irregular and regular shelterwood systems are mainly applied. Subalpine beech forests cover smaller parts of the whole area, usually at an elevation of between 1300 and 1600 m. There is no active management; the stands are left to natural development. This type is characterized by extreme site conditions, small tree heights (10–20 m), S-shaped stems due to snow pressure along the slope, vegetative regeneration and clump horizontal structure (Bončina et al. 2021).
Data
Forest inventory data (SFS 2014) were used as the primary data source on forest growth. Trees with a diameter at breast height (D) ≥ 10 cm are consecutively measured every 10 years on permanent sampling plots (n = 21,280; area = 500 m2) distributed on sampling grids of 250 m × 250 m and 250 m × 500 m. Beech was present on 12,193 sampling plots, and in total, 94,770 beech trees were used for the analyses (Appendix 2). Maps of vegetation units produced at a spatial scale of 1:10,000 were used for classifying the forest area into forest types. Topographic variables were derived from a digital elevation model (12.5 m resolution) (GURS 2014), while climatic variables were derived from long-term climate records in the period 1971–2000 (SEA 2021) and downscaled from the original 1 km2 resolution to the grid of sampling plots using the nearest-neighbour method.
Explanatory variables and their selection
Periodic diameter increment (DI), computed as the difference between two consecutive measurements of diameter at breast height in a 10-year period, was the dependant variable. Tree, stand and site variables were included in the analyses of beech diameter growth (Table 1). Among the tree explanatory variables, the initial diameter of a tree at the first measurement (D) was used as a proxy for tree size. Additionally, its square (D2) was used to account for the possible nonlinear relationship between DI and D.
Stand basal area (BA), the number of trees per hectare (N) and quadratic mean diameter (QMD) were calculated using the data from the first measurement of trees on the permanent sampling plots. Basal area and tree number per hectare were measures of stand density. QMD indicates the developmental stage or stand age of even-aged beech stands and has occasionally been included in the growth modelling of individual trees (e.g. Lhotka and Loewenstein 2011; Weiskittel et al. 2016). The structural diversity of forest stands was described by the Gini coefficient (GINI) (Weiner and Solbrig 1984), which was calculated at the plot level considering all trees from the first measurement considering the number and basal area of single trees with dbh ≥ 10 cm (Spellerberg 2008). A higher value of GINI, which ranges from 0 to 1, indicates uneven-sized stand structure, while values near 0 indicate even-sized stand structure. Tree species mixture was estimated by the Shannon index (SHAN) and by the proportion of beech in the total stand basal area (PBEECH). SHAN was calculated based on the proportion of single tree species in the total stand basal area for each plot. PBEECH was included in the analyses to test for differences in beech growth between stands with a different beech abundance.
Site productivity measured by the volume of a tree with a reference diameter of 45 cm (K) was included in the analyses. K ranged from 1.1 to 2.8 m3, indicating differences in tree heights of trees with the same diameter (Bončina et al. 2021). Five topographic variables were included as candidate variables in the analyses. Elevation (ELEV), inclination (INCL) and rockiness (ROCK) describe the variability of topographic conditions in the study area and indicate the severity or extremity of the site conditions. ROCK was taken from the forest inventory database (SFS 2014); it was visually assessed as the proportion of the area covered by stones and rocks. On carbonate substrate, rocks and stones are more frequent than on silicate bedrock; therefore, rockiness is often applied to describe growth conditions and protection forest functions and has also been used for forest classification from the perspective of forest vulnerability (Bončina et al. 2021). Eastness (EAST) and northness (NORTH) indices describe the aspect. Three soil variables [pH value (PH), the sum of organic matter (ORG) and soil depth (S_DEPTH)] derived from the digital soil map (MAFF 2021), and the following nine climatic variables representing the climatic averages in the period 1971–2000 (SEA 2021) were included in the analyses: annual amount of precipitation (MAP), mean annual temperature (MAT), minimum temperature (TMIN), maximum temperature (TMAX), mean diurnal range (BIO2 = TMAX-TMIN), maximum temperature of the warmest month (BIO5), minimum temperature of the coldest month (BIO6), annual temperature range (BIO7 = BIO5–BIO6) and isothermality (BIO3 = 100 × BIO2/BIO7).
Pearson’s correlation coefficients were calculated between continuous independent variables to verify collinearity; in pairs of variables with correlation coefficient r ≥ 0.65, one of the variables was not included in the modelling procedure. Among stand variables, N and SHAN were excluded from the procedure due to high correlation with BA and PBEECH, respectively. ELEV was excluded from the analyses due to high correlation with climatic variables. For instance, the Pearson correlation coefficient between ELEV and MAT, and ELEV and MAP, amounted to r = − 0.83 and r = 0.79, respectively. Most of climatic variables highly correlated with MAT and where therefore excluded (Table 1). To account for the possible interactions between precipitation and temperature, we included MAT × MAP in the model.
Mean DI at the regional level amounted to 2.75 cm (Fig. 1). It was highest in the submontane and montane forest types (2.86 cm), followed by the thermophilous (2.65 cm), upper montane (1.99 cm) and subalpine forest types (2.02 cm).

Periodic diameter increment of beech in five forest types (I, submontane; II, thermophilous; III, montane; IV, upper montane; and V, subalpine) presented by the mean value and 95% confidence interval. The vertical line denotes the overall mean in the region
Modelling approach
For modelling the DI of beech at the regional level, we used a linear mixed-effect model (Wykoff 1990; Uzoh and Oliver 2008) in the lmer function in the lme4 R package (Bates and Maechler 2010). Since trees were nested within sample plots, we included sample plots as a random effect. This allowed us to vary the intercepts in the regression equations by plots while keeping the variability of the slopes constant. The general form of a linear mixed-effect model is as follows (Bates and Maechler 2010; Bates et al. 2015):
where \(y_{ij}\) is the value of the predicted DI for a particular \(ij\) tree, \(\beta_{1}\) through \(\beta_{n}\) are the fixed effect coefficients (like regression coefficients), \(x_{{1_{ij} }}\) through \(x_{{n_{ij} }}\) are the fixed effect variables (predictors) for tree \(j\) on plot \(i\) (usually the first is reserved for the intercept/constant; \(x_{{1_{ij} }} = 1\)), \(b_{i1}\) through \(b_{in}\) are the random effect coefficients which are assumed to be multivariate normally distributed, \(z_{1ij}\) through \(z_{nij}\) are the random effect variables (predictors) and \(\varepsilon_{ij}\) is the error for tree \(j\) on plot \(i\) where each plot’s error is assumed to be multivariate normally distributed. The model parameters were estimated with maximum likelihood estimation (MLE) (Myung 2003).
To test the hypothesis about the equality of regression slopes among forest types, we separately parametrized the regional linear mixed-effect model (Eq. 1) for each forest type using the enter method. The differences in the slopes of the predictors between forest types were pairwise tested based on analysing the ANOVA p-value from the interaction of each variable by forest types (sensu ANCOVA, cf. Zar 2010: 364). The multiple comparison testing was performed using the lsmeans and lstrends functions in the emmeans R package (Lenth 2021).
To determine which site, stand and tree variables are the most important predictors for a specific forest type, the DI model (Eq. 1) was parametrized for each forest type with a stepwise procedure using all 17 independent variables as candidate variables (Table 1). The relative importance of individual predictors in the model was estimated by the relative decrease in the marginal R2 [R2m (%)] if the predictor was included in the model versus the model without the predictor. The fit of all models was evaluated with the marginal R2, conditional R2, root mean squared error (RMSE), intraclass correlation coefficient (ICC), random intercept variance (\(\tau_{00}\)), Akaike information criterion (AIC), Bayesian information criterion (BIC) and residual standard deviation (sigma) (Lüdecke et al. 2021). The predictive performance of a fitted model was evaluated with the performance R package (Lüdecke et al. 2021).
Results
The diameter growth of beech at the regional level
Ten of 17 variables remained in the DI model at the regional level (Table 2), which explained 32% of the total DI variability (Table 4). The model showed a nonlinear relationship between D and DI (Fig. 2); tree diameter contributed more than 40% to the explained DI variability. Stand variables contributed the largest portion to the explained variability of DI, with BA and PBEECH being the strongest individual stand predictors of DI. The growth of trees decreases when stand density (BA) increases. DI is lower in pure beech stands than in stands where beech is admixed. DI is higher in stands with uneven-sized structure and in stands with smaller QMD. Two topographic variables which indicate the extremity of the site conditions were included in the model. DI is lower on steeper slopes and in areas with higher rockiness. Three climatic variables contributed a minor part to the explained DI variability; DI is higher in sites with higher MAT and larger BIO2 and lower in sites with higher precipitation. The RMSE value was 1.3 cm (Table 4). Residuals were normally distributed, and the graph of residuals against predicted values showed no evidence of heteroscedasticity.

Testing the differences in the effect of site, stand and tree variables in forest types
The parameterizations of the regional model in each forest type (Table 3) showed that some explanatory variables, which were predictors in the regional model, were no longer significant. The models explained 25–34% of the DI variability at the forest type level, and the main relationships between tree, stand, topographic and climatic variables in explaining the DI variability remained the same as in the regional model. However, the contributions of some variables (e.g. PBEECH, BA) differed noticeably between forest types.
Comparison of parameter estimates between the regional and forest type models revealed significant differences in the effect of tree diameter (D and D2) only (Appendix 3). Parameter estimates for both terms in the model were significantly higher in the submontane forest type than at the regional level, and in the models for the upper montane and subalpine forest type, the effects of D and D2 were significantly lower than in the regional model.
The root mean square error (RMSE) ranged from 1.04 to 1.48 cm and was lower in the upper montane and subalpine forest types than in the forest types at lower elevations (Table 4). Marginal R2 values were in the interval 0.25–0.33. The random effect was significant in all forest types (albeit small), with the exception of subalpine forests, where the number of plots was relatively small and the random intercept variance (\(\tau_{00}\)) indicates negligible differences among plots.
Several model parameters significantly differed between forest types (Fig. 3; Appendix 3). No significant differences were found for GINI and BIO2. Most pairs of forest types significantly differ in the slopes of D and D2, four pairs in the slopes of QMD, three pairs in the slopes of PBEECH and ROCK, two pairs in the slopes of MAP and one pair in the slope of MAT. The least differences in the slopes of predictors were found between the submontane and thermophilous forest type, between the thermophilous and montane forest type and between the upper montane and the subalpine forest type.

Means and the corresponding 95% confidence intervals for parameter estimates in the DI models at the regional (R) and forest type level (I, submontane; II, thermophilous; III, montane; IV, upper montane; V, subalpine). The vertical lines denote no effect
The predictors with contrasting effects in some forest types were QMD, ROCK, MAP and BIO2 (Table 3); however, the differences in parameter estimates were mostly non-significant (Appendix 3). In the submontane forest type, the effect of MAP was positive and significantly different to that in the montane forest type.
DI models for forest types
The marginal R2 of models at the forest type level parametrized by the stepwise method using 17 candidate variables ranged from 26 to 33% (Table 5). The models differed in the variables included. Tree and stand variables contributed the majority of the explained DI variability in forest types (75–97%). Seven site variables were included in one model only, and all contributed negligible amounts to the explanation of DI variability. Two soil properties (PH and S_DEPTH) had a significant effect on beech growth in the upper montane forest types, while the third (ORG) had a significant positive impact on DI in thermophilous forests only. Climatic variables were included mainly in the models for montane and upper montane forests, and site productivity (K) only in the model for the upper montane forest type. All coefficients in the forest type models for the same predictors were of the same direction.
Discussion
Diameter increment of beech
In the studied region, the mean 10-year periodic diameter increment of beech amounted to 2.75 cm. Noticeable differences were found between forest types, with the diameter increment being highest in the submontane and montane forest types (2.9 cm), followed by the thermophilous, subalpine and upper montane forest types, with 7%, 29% and 30% lower diameter increment, respectively. In various growth and yield tables, data on the quadratic mean diameter of trees rather than the mean diameter of trees are available. In the Swiss growth and yield tables (Badoux 1983) for beech sites of the same productivity as the submontane and montane forests observed in our study, the 10-year increment of mean diameter amounted to 3.1 cm, while sites with lower production, such as the thermophilous and upper montane forests in our study, amounted to 2.7 cm and 2.5 cm, respectively. Pretzsch et al. (2021) reported a 10-year diameter increment of 3.2 cm for dominant beech trees across European mountain regions; in our study area, the average diameter increment of dominant beech trees only amounted to 3.7 cm (results not shown). The average mean diameter increment of beech trees across European countries amounted to 3.6 mm year−1, the same as that for Norway spruce (Schelhaas et al. 2018), which is surprisingly high compared to our results.
Predictors of diameter growth at the regional level
Ten variables were included in the regional diameter increment model for beech trees. The model explained 32% of the variation in diameter growth, which is more than that in similar studies on diverse sites (e.g. Schelhaas et al. 2018).
Tree diameter was found to be a crucial predictor of tree diameter increment. Tree diameter indicates the competition status of a tree in a stand and has frequently been included in growth modelling (e.g. Schelhaas et al. 2018), often as a linear effect (e.g. Bueno and Bevilacqua 2010; Hu et al. 2021). A similar result was reported for the diameter growth of dominant beech forests in mountain forests (Pretzsch et al. 2021). Our study showed a nonlinear response of diameter growth in regard to the diameter of trees. The same was found in a study of beech diameter growth in Europe (Schelhaas et al. 2018), and a similar result was reported for several tree species of mixed forests in the USA, including American beech (Weiskittel et al. 2016).
Stand variables explained the majority of the variability in beech diameter growth. Among them, stand basal area was the strongest individual predictor. A similar finding was reported by Schelhaas et al. (2018), who found that basal area explained most of the variance in the diameter growth of tree species in Europe. The relationship between stand basal area as a measure of competition and individual tree growth has been established in several studies (e.g. Wykoff 1990; Monserud and Sterba 1996; Hökkä et al. 1997; Uzoh and Oliver 2008).
The proportion of beech was the second most important stand variable, with a highly negative effect (p < 0.001). The correlations between the DI of beech and the proportions of other tree species at the regional scale (results not presented) showed the highest positive correlation between DI and the proportion of Norway spruce, while correlations with other species were lower but still significant and positive. Many studies on the productivity of mixed stands, especially those composed of Norway spruce and beech, have found mixed stands to be more productive (e.g. Pretzsch and Schütze 2021) than pure stands. Mina et al. (2018) reported that the complementarity of beech and other species strongly varies with stand, site and climatic conditions; however, their analyses did not explicitly consider forest types. Similarly, Huber et al. (2014) found a contrasting positive and negative mixing effect for silver fir and spruce depending on site quality and climate conditions. We did not study the impact of mixture type, but only the impact of the abundance of tree species other than beech on beech growth. Variables describing tree species mixture type were excluded based on preliminary analyses to avoid multicollinearity in the final model.
The impact of vertical structural diversity on the diameter growth of beech was quite weak, but still significant. The diameter increment was larger in structurally diverse stands. This is probably related to the competition status of trees and resource use efficiency. If trees in a given stand are diverse in size, then the available growth space might be more efficiently used than in a stand of the same basal area but with homogenous size structure. A similar impact was reported by Danescu et al. (2016), while many studies found the opposite impact. The impact of structural heterogeneity on stand productivity (e.g. Lei et al. 2009) has been more frequently studied than its impact on individual trees; several studies have confirmed the positive impact of structural heterogeneity on stand growth and have suggested niche complementary, especially in light use, as the underlying reason. According to Pretzsch (2014), this might not occur in all types of forest stands. Several studies (e.g. Liang et al. 2016; Wang et al. 2019) have even found that stand heterogeneity negatively impacts stand growth due to the reduced total light interception in the stand, mainly due to the low interception of smaller trees.
The severity of site conditions also had a significant effect on diameter growth. As expected, the effect was negative. Rockiness explained only a minor part of the variability, while inclination explained > 2% of the variability in the diameter increment, which is more than that explained by single climatic variables. Inclination and other topographic attributes may strongly influence soil attributes (e.g. soil moisture, soil organic carbon and nitrogen content) and thus the growth conditions for trees (Wu 2015).
Climatic conditions were also a source of variation in tree growth at the regional level. The impact of temperature and mean diurnal range was positive, while the diameter increment of beech weakly decreased with precipitation. According to the findings on the sensitivity of beech to warm and dry conditions (e.g. di Filippo et al. 2007; Betsch et al. 2011; Babst et al. 2013), which are especially evident at the southern edge of beech’s distribution (Schelhaas et al. 2018), to which our study area also belongs, we assumed a positive growth response of beech to higher precipitation in warm areas. However, the regional model does not support this hypothesis; the effect of precipitation was negative and the interaction between mean temperature and precipitation was not significant. Mean diurnal range is often used for modelling the current and the potential distribution of plant species due to climate change and is less frequent in tree growth models. In our study, the growth response to mean diurnal range was positive, possibly indicating better thermal conditions for growth at sites with greater means of all the monthly diurnal temperature ranges. The effect of mean diurnal range was significant for submontane and montane forests, but not for forests at higher vegetation belts, where other factors probably outweighed the effect of the monthly diurnal ranges.
The predictors which are recognized as highly influential for growth of single beech trees should be given higher priority in forest management decision-making (Wang et al. 2019), especially if they are easily regulated by silvicultural interventions. This is true for stand basal area and the proportion of beech, which together contributed 50% to the total explained diameter increment variability of beech trees at the regional level. Stand basal area can be regulated by the thinning regime, while the proportion of beech can be controlled by the type of regeneration and tending measures in young stands. All of this is valid on a single tree level, but not necessarily on a stand level.
Regional model versus forest type models
The regional model estimated an average effect of predictors on the diameter increment of beech. Since montane and submontane forest types prevail in the studied region, the fit of the regional model in these two forest types is better than that in the other three forest types (Appendix 3). For predicting the growth of beech at the forest type level, four questions appear to be relevant: (i) are the effects of predictors at the regional level different from the effects of these predictors at the forest type level, (ii) which variables that are predictors in the regional model are not predictors at the forest type level, (iii) are the effects of variables between forest types significantly different, and (iv) do the set of predictors significantly differ between forest types.
Regarding the transferability of the regional model to forest types (question number 1), we can conclude that the regional model was also applicable at the forest type level. The only variable whose effect at the forest type level statistically differed from that at the regional level was tree diameter. That was true for submontane, upper montane and subalpine forests. The differences may be a consequence of the different productivity of certain forest types in comparison to the regional average.
As to the second question about the significance of the predictors at the regional and forest type level, the regional and forest type models differ in the set of predictors. Only the model for the montane forest type included all variables from the regional level, which is probably related to the prevailing proportion of montane forests in the study area. In all other forest types, at least one variable had a non-significant impact on diameter increment. This is evident mainly for climatic variables, which can be explained by lower variability within single forest types. However, the same set of variables (i.e. tree size, stand basal area, heterogeneity and inclination) were significant for all forest types; therefore, their impact can justifiably be generalized across the whole region.
As to the third question about differences in the effect of growth predictors between forest types (Fig. 3; Appendix 3), we found significant differences in the effect of tree diameter between most of forest types. Differences in the effect of stand and climatic variables are of particular interest. The proportion of beech and stand basal area were by far the most important stand predictors of beech diameter growth. We found differences in the slopes of the proportion of beech for three pairs of forest types: upper montane versus submontane, thermophilous and montane. Several studies confirmed that the effect of tree species mixture varies in regard to stand density, stand development phase, and topographic, climatic and soil conditions (e.g. Huber et al. 2014; Mina et al. 2018); at least the last three are conditioned by forest type. Some studies (e.g. Forrester and Bauhus 2016) have reported that the mixing effect tends to increase along a temperature gradient. This appears to be indicated in our study; the slope of the beech proportion in the upper montane forest type was significantly lower than slopes in the submontane, thermophilous and montane forest types. The opposite pattern was observed for basal area; the slopes of basal area decreased from the submontane and thermophilous forest type to the montane and upper montane forest type. The effect of annual precipitation contributed negligibly to the explained variability of diameter increment, and it was not even significant in the single forest type models (Table 5). However, we revealed a significant difference in slopes for the amount of precipitation, which are even of different direction, between the submontane and montane forest types. This indicates that in the lower and warmer vegetation belt, beech grows better in sites with higher precipitation.
As to the fourth question about the best set of growth predictors at the forest type level (Table 5), we found great differences among types in the predictors and their effects, and some specific effects that were not relevant in the regional model (e.g. soil depth and pH). However, the same set of variables (i.e. tree diameter, basal area, proportion of beech) explained the majority of DI variability in all forest types, with the subalpine forest type being an exception. This is probably due to small sample in this type. The area of subalpine forests is minor in comparison with that of submontane or montane beech forests. Despite the small sample size, we included this type in the analyses to provide a complete picture of growth of beech from submontane forests to the upper timber line, which might be particularly relevant under climate change and the possible shift of beech towards higher elevations. The proportion of explained variability in models at the forest type level is virtually the same as if the regional model was parametrized at the forest type level. If the AIC is considered, then the regional model parametrized at the forest type levels shows even better performance for some forest types than separate models. This means that new variables did not noticeably improve the models at the forest type level; rather, they indicate growth specificities or perhaps larger gradients of these variables in a certain forest type.
Conclusions
This study highlights an area of tree diameter growth model development and the application of models to different spatial scales. By comparing a regional growth model to models with local differentiation, the following conclusions can be drawn:
First, the regional model can be transferred to a single forest type quite well because most of the variability in diameter growth is explained by the same set of stand parameters. However, significant differences in beech growth for some basic variables such as diameter at breast height may exist in forest types, which warrants caution when using predictions from the regional model at the forest type level. Including site productivity in the regional model may partially account for the forest type-specific responses, but this should not serve as a replacement for forest type.
Second, the climate–growth response is more prominent at the regional level than at the forest type level. Classification of forest types considers climate and topographic conditions; therefore, reducing the relevance of climatic variables in the models at forest type level was expected. An additional reason could be related to the rather coarse data on climate–growth relationships at the level of the single forest type, which do not enable the detection of climatic impacts.
Third, the best set of predictors and parameter estimates for the same predictors may differ significantly between forest types, with some effects even being of the opposite direction. However, the total explained variability in both the regional model and the forest type-specific models do not markedly differ.
The significant differences in some model parameters between forest types, especially those which contributed the largest portion to the explained variability of diameter increment, and some specific predictors for single forest type models, albeit with small relevance in the models, emphasize the importance of considering forest types in tree growth modelling.
References
Badoux E (1983) Ertragstafeln. Birmensdorf, Zürich
Babst F, Poulter B, Trouet V et al (2013) Site-and species-specific responses of forest growth to climate across the European continent. Glob Ecol Biogeogr 22:706–717. https://doi.org/10.1111/GEB.12023/SUPPINFO
Bates D, Mächler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67:1–48. https://doi.org/10.18637/jss.v067.i01
Bates D, Maechler M (2010) Package lme4: Reference manual for the package. R package v (1.1–15) (2010). http://cran.r-project.org/web/packages/lme4/lme4.pdf
Betsch P, Bonal D, Breda N et al (2011) Drought effects on water relations in beech: the contribution of exchangeable water reservoirs. Agric For Meteorol 151:531–543. https://doi.org/10.1016/J.AGRFORMET.2010.12.008
Böhn U, Gollub G, Hettwer C, Neuhäuslová Z, Raus T, Schlüter H (2004) Karte der natürlichen Vegetation Europas/Map of the natural vegetation of Europe. BfN, Bonn-Bad Godesberg, Germany
Bončina A. (ed.) (2012) Bukovi gozdovi v Sloveniji: ekologija in gospodarjenje (Beech forests in Slovenia: ecology and management). Biotehniška fakulteta, Oddelek za gozdarstvo in obnovljive gozdne vire. Ljubljana, Slovenia
Bolte A, Czajkowski T, Kompa T (2007) The north-eastern distribution range of European beech—a review. For an Int J for Res 80:413–429. https://doi.org/10.1093/FORESTRY/CPM028
Bončina A et al (2021) Gozdni rastiščni tipi Slovenije: vegetacijske, sestojne in upravljavske značilnosti (Forest Site Types in Slovenia: Vegetation, Stand and Management Characteristics). Oddelek za gozdarstvo in obnovljive gozdne vire Biotehniška fakulteta, Ljubljana, Slovenia
Bosela M, Štefančík I, Petráš R, Vacek S (2015a) The effects of climate warming on the growth of European beech forests depend critically on thinning strategy and site productivity. Agric For Meteorol 222:21–31. https://doi.org/10.1016/J.AGRFORMET.2016.03.005
Bosela M, Tobin B, Šebeň V et al (2015b) Different mixtures of Norway spruce, silver fir, and European beech modify competitive interactions in central European mature mixed forests. Can J For Res 45:1577–1586. https://doi.org/10.1139/cjfr-2015-0219
Bueno S, Bevilacqua E (2010) Modeling stem diameter increment in individual Pinus occidentalis Sw. trees in La Sierra Dominican Republic. For Syst 19:170–183. https://doi.org/10.5424/FS/2010192-01312
Cajander AK (1949) Forest types and their significance. Acta For Fenn 56(5):1–71
Charru M, Seynave I, Morneau F, Bontemps JD (2010) Recent changes in forest productivity: an analysis of national forest inventory data for common beech (Fagus sylvatica L.) in north-eastern France. For Ecol Manag 260:864–874. https://doi.org/10.1016/J.FORECO.2010.06.005
Dakskobler I (2008) Pregled bukovih rastišč v Sloveniji (A review of beech sites in Slovenia). Zb Gozdarstva Lesar 87:3–14
Dănescu A, Albrecht AT, Bauhus J (2016) Structural diversity promotes productivity of mixed, uneven-aged forests in southwestern Germany. Oecologia 182:319–333. https://doi.org/10.1007/s00442-016-3623-4
Dulamsuren C, Hauck M, Kopp G et al (2016) European beech responds to climate change with growth decline at lower, and growth increase at higher elevations in the center of its distribution range (SW Germany). Trees 31:673–686. https://doi.org/10.1007/S00468-016-1499-X
Di Filippo A, Biondi F, Čufar K, De Luis M, Grabner M, Maugeri M, Presutti Saba E, Schirone B, Piovesan G (2007) Bioclimatology of beech (Fagus sylvatica L.) in the Eastern Alps: spatial and altitudinal climatic signals identified through a tree-ring network. J Biogeogr 34(11):1873–1892. https://doi.org/10.1111/j.1365-2699.2007.01747.x
Ellenberg H, Leuschner C (2010) Nährstoffumsätze. Veg. Mitteleuropas mit den Alpen ökologischer, dynamischer und Hist. Leverkusen, Germany
Etzold S, Ferretti M, Reinds GJ et al (2020) Nitrogen deposition is the most important environmental driver of growth of pure, even-aged and managed European forests. For Ecol Manag 458:117762. https://doi.org/10.1016/J.FORECO.2019.117762
Forrester DI (2015) Transpiration and water-use efficiency in mixed-species forests versus monocultures: effects of tree size, stand density and season. Tree Physiol 35:289–304. https://doi.org/10.1093/TREEPHYS/TPV011
Forrester DI (2019) Linking forest growth with stand structure: tree size inequality, tree growth or resource partitioning and the asymmetry of competition. For Ecol Manag 447:139–157. https://doi.org/10.1016/J.FORECO.2019.05.053
Forrester DI, Bauhus J (2016) A review of processes behind diversity-productivity relationships in forests. Curr For Rep 2:45–61. https://doi.org/10.1007/S40725-016-0031-2/FIGURES/3
Friedrichs DA, Trouet V, Büntgen U et al (2009) Species-specific climate sensitivity of tree growth in Central-West Germany. Trees 23:729–739. https://doi.org/10.1007/S00468-009-0315-2
GURS (2014) Prostorske podatkovne zbirke Republike Slovenije (Spatial dataset of Republic of Slovenia). Minstrstvo za okolje in prostor. Geodetska uprava Republike Slovenije, Ljubljana
Goisser M, Geppert U, Rötzer T et al (2016) Does belowground interaction with Fagus sylvatica increase drought susceptibility of photosynthesis and stem growth in Picea abies? For Ecol Manag 375:268–278. https://doi.org/10.1016/J.FORECO.2016.05.032
Hanewinkel M, Hummel S, Albrecht A (2010) Assessing natural hazards in forestry for risk management: a review. Eur J For Res 130:329–351. https://doi.org/10.1007/S10342-010-0392-1
Hartl-Meier C, Dittmar C, Zang C, Rothe A (2014) Mountain forest growth response to climate change in the Northern Limestone Alps. Trees 28:819–829. https://doi.org/10.1007/s00468-014-0994-1
Hilmers T, Avdagić A, Bartkowicz L et al (2019) The productivity of mixed mountain forests comprised of Fagus sylvatica, Picea abies, and Abies alba across Europe. For Int J For Res 92:512–522. https://doi.org/10.1093/FORESTRY/CPZ035
Hökkä H, Alenius V, Penttilä T (1997) Individual-tree basal area growth models for Scots pine, pubescent birch and Norway spruce on drained peatlands in Finland. Silva Fenn 31:161–178. https://doi.org/10.14214/SF.A8517
Hu X, Duan G, Zhang H (2021) Modelling individual tree diameter growth of Quercus mongolica secondary forest in the northeast of China. Sustainability 13:4533. https://doi.org/10.3390/SU13084533
Huber MO, Sterba H, Bernhard L (2014) Site conditions and definition of compositional proportion modify mixture effects in Picea abies–Abies alba stands. Can J For Res 44:1281–1291. https://doi.org/10.1139/CJFR-2014-0188/SUPPL_FILE/CJFR-2014-0188SUPPL.PD
Jourdan M, Lebourgeois F, Morin X (2019) The effect of tree diversity on the resistance and recovery of forest stands in the French Alps may depend on species differences in hydraulic features. For Ecol Manag 450:117486. https://doi.org/10.1016/J.FORECO.2019.117486
Jump AS, Hunt JM, Pen̈uelas J (2006) Rapid climate change-related growth decline at the southern range edge of Fagus sylvatica. Glob Chang Biol 12:2163–2174. https://doi.org/10.1111/J.1365-2486.2006.01250.X
Kirchen G, Calvaruso C, Granier A et al (2017) Local soil type variability controls the water budget and stand productivity in a beech forest. For Ecol Manag 390:89–103. https://doi.org/10.1016/J.FORECO.2016.12.024
Lüdecke et al (2021) Package performance: an R package for assessment, comparison and testing of statistical models R package v (0.8.0.). https://easystats.github.io/performance/
Lenth RV (2021) Package emmeans: estimated marginal means, aka least-squares means. R package v (1.7.1-1). https://github.com/rvlenth/emmeans
Lei X, Wang W, Peng C (2009) Relationships between stand growth and structural diversity in spruce-dominated forests in New Brunswick. Can J For Res 39(10):1835–1847. https://doi.org/10.1139/X09-089
Lhotka JM, Loewenstein EF (2011) An individual-tree diameter growth model for managed uneven-aged oak-shortleaf pine stands in the Ozark Highlands of Missouri, USA. For Ecol Manag 261:770–778. https://doi.org/10.1016/J.FORECO.2010.12.008
Liang J, Buongiorno J, Monserud RA et al (2007) Effects of diversity of tree species and size on forest basal area growth, recruitment, and mortality. For Ecol Manag 243:116–127. https://doi.org/10.1016/J.FORECO.2007.02.028
Liang J, Crowther TW, Picard N et al (2016) Positive biodiversity-productivity relationship predominant in global forests. Science. https://doi.org/10.1126/SCIENCE.AAF8957
Mayer H (1974) Wälder des Ostalpenraumes. Stuttgart, Germany
MAFF (2021) Pedological map. Ministry of Agriculture, Forestry and Food, Ljubljana
Martinez del Castillo E, Zang CS, Buras A et al (2022) (2022) Climate-change-driven growth decline of European beech forests. Commun Biol 5(1):1–9. https://doi.org/10.1038/s42003-022-03107-3
Mina M, Huber MO, Forrester DI et al (2018) Multiple factors modulate tree growth complementarity in Central European mixed forests. J Ecol 106:1106–1119. https://doi.org/10.1111/1365-2745.12846
Monserud RA, Sterba H (1996) A basal area increment model for individual trees growing in even- and uneven-aged forest stands in Austria. For Ecol Manag 80:57–80. https://doi.org/10.1016/0378-1127(95)03638-5
Myung IJ (2003) Tutorial on maximum likelihood estimation. J Math Psychol 47:90–100. https://doi.org/10.1016/S0022-2496(02)00028-7
Pretzsch H (1997) Analysis and modeling of spatial stand structures. Methodological considerations based on mixed beech-larch stands in lower Saxony. For Ecol Manag 97:237–253. https://doi.org/10.1016/S0378-1127(97)00069-8
Pretzsch H, Schütze G (2021) Tree species mixing can increase stand productivity, density and growth efficiency and attenuate the trade-off between density and growth throughout the whole rotation. Ann Bot 128:767–786. https://doi.org/10.1093/AOB/MCAB077
Pretzsch H, Rötzer T, Matyssek R et al (2014) Mixed Norway spruce (Picea abies [L.] Karst) and European beech (Fagus sylvatica [L.]) stands under drought: from reaction pattern to mechanism. Trees 28:1305–1321. https://doi.org/10.1007/S00468-014-1035-9
Pretzsch H, Hilmers T, Uhl E et al (2020) European beech stem diameter grows better in mixed than in mono-specific stands at the edge of its distribution in mountain forests. Eur J For Res 1401(140):127–145. https://doi.org/10.1007/S10342-020-01319-Y
Pretzsch H, Hilmers T, Biber P et al (2020) Evidence of elevation-specific growth changes of spruce, fir, and beech in European mixed mountain forests during the last three centuries. Can J For Res 50:689–703. https://doi.org/10.1139/CJFR-2019-0368/ASSET/IMAGES/LARGE/CJFR-2019-0368F9.JPEG
Pretzsch H (2009) Forest dynamics, growth and yield. From measurement to model. Springer Berlin, Heidelberg
Ray D (2001) Ecological site classification decision support system V1.7. Forestry Commission, Edinburgh
Roberts DW (2015) Potential natural vegetation and environment: a critique of Kusbach, Shaw and Long. Appl Veg Sci 18:733–738. https://doi.org/10.1111/AVSC.12177
Scharnweber T, Manthey M, Criegee C, Bauwe A, Schröder A, Wilmking M (2011) Drought matters—declining precipitation influences growth of Fagus sylvatica L. and Quercus robur L. in north-eastern Germany. For Ecol Manag 262:947–961
Schelhaas MJ, Hengeveld GM, Heidema N et al (2018) Species-specific, pan-European diameter increment models based on data of 2.3 million trees. For Ecosyst 5:21. https://doi.org/10.1186/s40663-018-0133-3
SEA (2021) Spatial climate data of Slovenia 1971–2000. Slovenian Environment Agency, Ljubljana. https://gis.arso.gov.si/geoportal/catalog/main/home.page. Accessed 26 June 2021
Seltmann CT, Wernicke J, Petzold R et al (2021) The relative importance of environmental drivers and their interactions on the growth of Norway spruce depends on soil unit classes: a case study from Saxony and Thuringia Germany. For Ecol Manag 480:118671. https://doi.org/10.1016/j.foreco.2020.118671
SFS, 2014: Forestry data collection. Slovenia Forest Service, Ljubljana
Spellerberg IF (2008) Shannon-Wiener Index. In: Encyclopedia of Ecology. Oxford
Tegel W, Seim A, Hakelberg D et al (2014) A recent growth increase of European beech (Fagus sylvatica L.) at its Mediterranean distribution limit contradicts drought stress. Eur J For Res 33:61–71. https://doi.org/10.1007/S10342-013-0737-7
Tenzin J, Tenzin K, Hasenauer H (2017) Individual tree basal area increment models for broadleaved forests in Bhutan. For Int J For Res 90:367–380. https://doi.org/10.1093/FORESTRY/CPW065
Thurm EA, Pretzsch H (2016) Improved productivity and modified tree morphology of mixed versus pure stands of European beech (Fagus sylvatica) and Douglas-fir (Pseudotsuga menziesii) with increasing precipitation and age. Ann For Sci 73:1047–1061. https://doi.org/10.1007/S13595-016-0588-8/TABLES/2
Trotsiuk V, Hartig F, Cailleret M et al (2020) Assessing the response of forest productivity to climate extremes in Switzerland using model-data fusion. Glob Chang Biol 26:2463–2476. https://doi.org/10.1111/GCB.15011
Trouillier M, van der Maaten-Theunissen M, Scharnweber T, Wilmking M (2020) A unifying concept for growth trends of trees and forests—the “potential natural forest.” Front For Glob Chang 3:113. https://doi.org/10.3389/FFGC.2020.581334/BIBTEX
Uzoh FCC, Oliver WW (2008) Individual tree diameter increment model for managed even-aged stands of ponderosa pine throughout the western United States using a multilevel linear mixed effects model. For Ecol Manag 256:438–445. https://doi.org/10.1016/j.foreco.2008.04.046
Vospernik S (2021) Basal area increment models accounting for climate and mixture for Austrian tree species. For Ecol Manag 480:118725. https://doi.org/10.1016/j.foreco.2020.118725
Wang W, Chen X, Zeng W et al (2019) Development of a mixed-effects individual-tree basal area increment model for oaks (Quercus spp.) considering forest structural diversity. Forests 10(6):474. https://doi.org/10.3390/f10060474
Weiner J, Solbrig OT (1984) The meaning and measurement of size hierarchies in plant populations. Oecologia 61:334–336. https://doi.org/10.1007/BF00379630
Weiskittel A, Kuehne C, McTague JP, Oppenheimer M (2016) Development and evaluation of an individual tree growth and yield model for the mixed species forest of the Adirondacks Region of New York, USA. For Ecosyst 3:1–17. https://doi.org/10.1186/S40663-016-0086-3/FIGURES/4
Wu H (2015) The relationship between terrain factors and spatial variability of soil nutrients for pine-oak mixed forests in Qinlang Mountains. J Nat Res 30:858–869. https://doi.org/10.11849/zrzyxb.2015.05.013
Wykoff WR (1990) A basal area increment model for individual conifers in the Northern Rocky mountains. For Sci 36:1077–1104. https://doi.org/10.1093/FORESTSCIENCE/36.4.1077
Wykoff WR, Crookston NL, Stage AR (1982) User’s guide to the stand prognosis model (forestry). US Department of Agriculture, Forest Service, General Technical Report, INT-133
Zar JH (2010) Biostatistical analysis. New Jersey
Zeller L, Pretzsch H (2019) Effect of forest structure on stand productivity in Central European forests depends on developmental stage and tree species diversity. For Ecol Manag 434:193–204. https://doi.org/10.1016/j.foreco.2018.12.024
Zimmermann J, Hauck M, Dulamsuren C, Leuschner C (2015) Climate warming-related growth decline affects Fagus sylvatica, but not other broad-leaved tree species in Central European mixed forests. Ecosystems 18:560–572. https://doi.org/10.1007/s10021-015-9849-x
Zlatník A (1976) Lesnická fytologie. Forest phytology. SZN, Praha
Acknowledgements
We thank the Slovenia Forest Service for providing the measurement data from the permanent sampling plots. We thank the anonymous reviewers for their comments and suggestions. The study was supported by the Slovenian Ministry of Agriculture, Forestry and Food (projects: The development of forest models for Slovenia, Grant no. V4-2014; Managing forest risks due to climate change; Grant no. V4-2211) and the Slovenian Research Agency (research core funding Grant no. P4-0059).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Peter Biber.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Appendices
Appendix 1
See Table
6.
Appendix 2
See Table
7.
Appendix 3
See Table
8.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

{kind=link}
{kind=link}
{kind=link}
Cite this article
Bončina, A., Trifković, V., Ficko, A. et al. Diameter growth of European beech on carbonate: a regional versus forest type perspective. Eur J Forest Res 142, 917–932 (2023). https://doi.org/10.1007/s10342-023-01562-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-023-01562-z