
Abstract
Sequence variability in the cytochrome c oxidase I (COI) gene from 226 samples of the species previously considered Bathyraja parmifera (Rajidae) revealed three distinct haplotypes, one of which represents an undescribed species, the leopard skate. Further genetic examination of four closely related North Pacific and Bering Sea skate species, Bathyraja parmifera, B. simoterus, B. smirnovi, and the leopard skate in comparison with 19 related species indicates that together these four species comprise the subgenus Arctoraja. Phylogenetic analysis suggests that Arctoraja is monophyletic, but that the genus Bathyraja may be paraphyletic due to the phylogenetic position of Rhinoraja.
Similar content being viewed by others


Introduction
Morphological work has suggested that the Alaska skate, Bathyraja parmifera, is a species complex. Assessment surveys of groundfish resources conducted by the Alaska Fisheries Science Center (Seattle, WA, USA) have revealed a related undescribed species, now commonly known as the leopard skate (Stevenson et al. 2007), in the western Aleutian Islands; its scientific name and formal description will be published soon (Orr et al. in press). In addition, the relationship between B. parmifera and the closely related species Bathyraja simoterus and Bathyraja smirnovi, all members of the subgenus Arctoraja Ishiyama 1958, has not been fully resolved (see Orr et al. in press).
In this study we examine cytochrome c oxidase subunit I (COI) sequence variation within the subgenus Arctoraja and 19 related species: 15 other species of Bathyraja (10 from the North Pacific, 2 from the northwestern Pacific, 1 each from the North Atlantic, South Atlantic, and Southern Oceans), 1 species of Rhinoraja, 2 species of Raja endemic to the North Pacific and Bering Sea, and Amblyraja badia of the North Pacific [Electronic supplementary material (ESM) Table S1]. Cytochrome c oxidase subunit I (COI) has become the standard species-level “barcode” sequence, as 95% of species possess a distinctive sequence in this region (Hebert et al. 2003). Several studies have shown that COI sequence data can be highly accurate in the identification of skates and other elasmobranch species (Spies et al. 2006; Smith et al. 2008; Ward et al. 2008).
These COI sequences provide evidence regarding phylogenetic relationships of the subgenus Arctoraja as well as other North Pacific species of Bathyraja. In addition, we examine haplotypes of 226 individuals of the B. parmifera species complex. Analysis of these haplotypes provides information on the haplotype diversity of the species complex and corroborates the species-level status of the leopard skate, an undescribed species to be formally described by Orr et al. (in press).
Materials and methods
Sample collection. All samples of North Pacific Bathyraja were collected in Alaskan waters during research trawl surveys conducted by the Resource Assessment and Conservation Engineering Division (RACE) of the Alaska Fisheries Science Center, or during commercial fishing operations sampled by fisheries observers. Tissues of Bathyraja simoterus (n = 2) were obtained from Hokkaido University (HUMZ), and tissues of Bathyraja smirnovi (n = 2), Bathyraja matsubarai (n = 3), Bathyraja fedorovi (n = 2), and Rhinoraja longicauda (n = 2) were obtained from the National Science Museum (NSMT), Tokyo, Japan; all of these were collected in Japanese waters. Bathyraja richardsoni tissues (n = 2) from the North Atlantic were obtained from the University of Bologna. Tissues of B. simoterus were formalin-fixed, while all other tissues were preserved in nondenatured 95% ethanol prior to extraction. Consensus sequence data for the representatives of the South Atlantic and Southern Oceans [Bathyraja griseocauda (n = 8), Bathyraja maccaini (n = 18)] and an additional sequence of B. richardsoni from southern New Zealand waters were obtained from GenBank (Smith et al. 2008). All additional tissues and available voucher material are archived at the University of Washington Fish Collection (UW), Seattle, WA, USA, except for selected type material. See the “Materials Examined” section of Orr et al. (in press) for disposition of type material and other catalog information for tissue samples.
DNA extraction, amplification, and sequencing. For the ethanol-preserved specimens, including the leopard skate (n = 8), B. parmifera (n = 39), B. smirnovi (n = 2), B. matsubarai (n = 3), B. fedorovi (n = 2), R. longicauda (n = 2), Raja binoculata (n = 2), Raja rhina (n = 3), Bathyraja interrupta (n = 11), Bathyraja mariposa (n = 1), Bathyraja aleutica (n = 1), B. richardsoni (n = 2) and Bathyraja taranetzi (n = 6), DNA extraction was performed using Qiagen DNeasy extraction kits (Qiagen, Inc., Valencia, CA, USA). Subsequent amplification of COI via the polymerase chain reaction (PCR) was performed using COI_Raja primers (COI_RajaF; 5′-CCG CTT AAC TCT CAG CCA TC-3′ and COI_RajaR; 5′-TCA GGG TGA CCA AAG AAT CA-3′) according to the protocol in Spies et al. (2006). PCR products were diluted to 7 ng/μl with sterile dH2O, and sequenced with COI_Raja primers at the University of Washington High-Throughput Genomics Unit, Seattle, WA, USA (http//www.htseq.org). Forward and reverse sequences were aligned using Sequencher v4.2 (Gene Codes Corp., Ann Arbor, MI, USA) and all references to base pair location are based on alignment with sequences previously submitted to GenBank with the following accession numbers: DQ104880–104933, DQ665297, DQ665298, DQ385444 (Spies et al. 2006). Up to 720 bp of each sample were sequenced. Sequences for species of the subgenus Arctoraja were subsequently trimmed to 557 bp in order to have a complete alignment, while our alignment representative of the genus Bathyraja was trimmed to 498 bp. Final sequences were submitted to GenBank, as presented in ESM Table S1.
Five formalin-fixed specimens represent the known extant material of B. simoterus (Orr et al. in press). Due to their relative scarcity, fresh material was not available for genetic analysis. However, we were able to obtain two samples of B. simoterus (HUMZ 143363 and 143369) that had been stored in 10% formalin for at least 13 years. Although notoriously difficult, DNA extraction has been successfully performed from formalin-fixed tissue (France and Kocher 1996; Chase et al. 1998). Initial efforts to extract DNA and perform PCR as described above failed; therefore, two slightly different methods were tried on the two specimens in order to successfully obtain sequence data. For both formalin-fixed samples, tissue was soaked overnight in low Tris–EDTA buffer (10 mM Tris–HCl, 0.1 mM EDTA) at room temperature and subsequently extracted using the Qiagen DNeasy kit according to the standard protocol. After PCR amplification of the COI region as described above, PCR products were cloned using the Invitrogen TOPO TA cloning kit for sequencing (Invitrogen, Inc., Carlsbad, CA, USA). For sample HUMZ 143363, clones were grown overnight in 4 ml of LB-ampicillin broth, and then mini-prepped using the QIAprep Spin Miniprep kit (Qiagen, Inc.). The COI region was then amplified using universal plasmid primers (M13f-20 and M13r) in a reaction seeded with 0.5 μl of miniprep DNA in a total volume of 20 μl. Clones from sample HUMZ 143369 were not mini-prepped. Rather, the PCR was seeded by touching the clone to a pipette tip in a 20 μl volume. Four clones from each specimen were chosen for sequencing. For each clone, the PCR product was diluted to 7 ng/μl with sterile dH2O, and sequenced with COI_Raja primers at the University of Washington High-Throughput Genomics Unit as described above. A total of three forward and three reverse sequences were obtained for HUMZ 143363, and four forward and four reverse sequences were obtained for HUMZ 143369. Sequences were aligned using Sequencher v4.2 (Gene Codes Corp., Ann Arbor, MI, USA), and a 770-bp consensus sequence was obtained for each sample.
The initial sequencing of the leopard skate (n = 8) and B. parmifera (n = 39) indicated fixed haplotype differences between the two taxa, with two distinct haplotypes for B. parmifera. Therefore, a restriction fragment length polymorphism (RFLP) protocol was developed with the restriction enzyme TaqαI (New England Biolabs, Ipswich, MA, USA) to quickly and inexpensively distinguish haplotypes of 179 additional samples from the B. parmifera species complex, which includes the leopard skate, B. parmifera (common haplotype), and B. parmifera (rare haplotype). We extracted DNA using Qiagen DNeasy extraction kits as described above, amplified genomic DNA with COI_Raja primers, and digested with TaqαI in a 5 μl reaction containing 1× NEBuffer 4 and 2 μl PCR product at 65°C for 2 h. Product was electrophoresed on a 2% agarose gel in TE buffer at 50 mA for 2 h along with confirmed positive controls representing the three different haplotypes. Gels were stained in ethidium bromide for 20 min prior to visualization. All genetic screening was performed without regard to information on specimen morphology and collection location.
Phylogenetic analyses. Phylogenetic analyses were performed using COI consensus sequences from 23 species; all 15 valid Alaskan species of Bathyraja, Raja, and Amblyraja; the two putative western North Pacific species of the subgenus Arctoraja (B. smirnovi and B. simoterus); two additional species of Bathyraja from the western North Pacific (B. fedorovi and B. matsubarai); one from the North Atlantic (B. richardsoni); one from the South Atlantic (B. griseocauda); one from the Southern Ocean (B. maccaini); and one species of the western North Pacific genus Rhinoraja (R. longicauda) (ESM Table S1). Consensus sequences (557 bp) were derived from high-quality haplotype sequences and were aligned using BioEdit version 7.0.9.0 (Hall 1999).
Nucleotide diversity (π), haplotype diversity, and the number of polymorphic sites were calculated in DnaSP 4.50.3 (Rozas et al. 2003). Sequence divergence values between species were calculated in MEGA 4.0 (Tamura et al. 2007), using the Kimura two-parameter (K2P) distance model (Kimura 1980). The K2P distance was chosen because it incorporates proportions of both transitions and transversions, is a common choice for describing differentiation among species using COI (Hebert et al. 2003; Ward et al. 2008), and is considered a reliable statistic for describing sequence divergence when it is low (Nei and Kumar 2000) (Table 1).
Phylogenetic analysis was performed under both maximum parsimony and Bayesian criteria. Three species of the Rajinae (R. binoculata, R. rhina, and A. badia), the sister group of the Arhynchobatinae (McEachran and Dunn 1998), were selected as outgroup taxa. For the parsimony analysis, the matrix was analyzed with PAUP*4.0b (Swofford 2003), using the branch and bound search algorithm for an exact search (Hendy and Penny 1982). To evaluate branch support, a heuristic bootstrap analysis of 1,000 replicates was conducted, with simple addition sequence and TBR (tree bisection-reconnection) branch-swapping options. Bremer decay values, an error estimate similar to bootstrap values except that it is a measure of the number of steps between the score of the most parsimonius tree and the score of the same tree with that particular node removed (Bremer 1988), were calculated using TREEROT v.2 (Sorenson 1999).
For the Bayesian analysis, MrModeltest 2.3 (Nylander 2004) was used to select the highest likelihood model of nucleotide substitution. A hierarchical likelihood ratio test chose the HKY substitution model with gamma-distributed rate variation across sites (HKY + Γ) (Hasegawa et al. 1984). The model suggested by the Akaike information criterion (AIC) was the same, but with the addition of invariant sites (HKY + I + Γ). Both models were analyzed with MrBayes v3.1.2 (Huelsenbeck and Ronquist 2001; Ronquist and Huelsenbeck 2003). Posterior probability distributions were generated by running four Markov chains for 1.5 million generations and sampling every 100 generations, under the default of three heated chains and one cold. The initial 3,750 samples were discarded as burn-in, and the remaining 11,250 samples used to estimate tree topology, posterior probabilities, and branch lengths. The harmonic mean of sampled log likelihood values, which acts as a proxy for the likelihood of a particular model, was greater for the HKY + Γ model (–2174.20) than for the HKY + I + Γ model (–2181.58). Twice this difference, the Bayes factor, is 14.76, which is strong evidence that the HKY + Γ model is a better fit for this data set (Kass and Raftery 1995). Thus, only the results of the HKY + Γ model are presented here. Final tree topology was identical for both models and posterior probabilities at all nodes were identical, or nearly so.
Results
We observed four distinct haplotypes among the 230 samples from the four species of Arctoraja. Of the 557 bases, there were 10 variable sites, with a total of 11 mutations, 2 singleton sites, and 9 parsimony-informative sites. All mutations took place at the third codon position, and thus were synonymous sites. Eight of the informative sites had two variants, while one (at position 366) had three variants. The consensus sequences for the two individuals of Bathyraja simoterus were identical, except for one ambiguous nucleotide at position 21 of HUMZ 143369, which is not a variable site among the four species of Arctoraja. We observed a single haplotype within leopard skate samples, and B. smirnovi and B. simoterus shared a single haplotype.
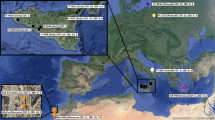
Within B. parmifera, substitutions at three nucleotide sites (positions 366, 453, and 555) yielded three haplotypes: the leopard skate with “TAC,” the common haplotype, Bpar1, with “GAT,” and the rare haplotype, Bpar2, with “AGT.” Within B. parmifera, nucleotide (sequence) diversity, π, was 0.0015 and haplotype diversity was 0.39, due to the presence of the two distinct haplotypes. The less common of the two B. parmifera haplotypes was found in 16.8% of the samples, and one of the seven rare haplotypes contained a singleton mutation, a C at position 393. The haplotype diagnostic for the leopard skate was found only in the western Aleutian Islands west of Amchitka Pass, while only the common B. parmifera haplotype was found in the central Aleutian Islands, from Amchitka Pass to Samalga Pass. Both haplotypes of B. parmifera were found throughout the eastern Aleutian Islands, Bering Sea, and Gulf of Alaska (Fig. 1).

Geographic locations of the leopard skate (squares), Bathyraja parmifera common haplotype (Bparm1, large black circles) and the B. parmifera rare haplotype (Bparm2, small white circles), B. smirnovi (Bsmi, triangles), and B. simoterus (Bsim, stars)
Sequence divergence, the percent difference in nucleotide sequence between related nucleic acid sequences, was 0.36% between the two haplotypes of B. parmifera and between B. parmifera and the leopard skate. Sequence divergence between the leopard skate and the rare haplotype of B. parmifera was 0.54%.
Phylogenetic results. Parsimony analysis resulted in 49 equally parsimonious trees of 286 steps [consistency index (CI) = 0.669, retention index (RI) = 0.818]. The 50% majority rule consensus tree (Fig. 2) did not recover a monophyletic Bathyraja. The single representative of Rhinoraja (R. longicauda) was nested deep within Bathyraja, rendering the latter paraphyletic. Within the ingroup, three significant clades were well supported: the subgenus Arctoraja, comprising the leopard skate, B. parmifera, B. smirnovi, and B. simoterus; a clade consisting of B. interrupta, B. violacea, and B. mariposa (subsequently referred to as the B. interrupta clade), and a clade including B. maculata, B. lindbergi, and B. matsubarai (referred to as the B. matsubarai clade). Within Arctoraja, the eastern Pacific species B. parmifera and the leopard skate are sister species, and as a clade are sister to the B. smirnovi + B. simoterus clade.

Consensus parsimony (left), with Bremer decay values above branches and bootstrap values below, and Bayesian (right) consensus tree, with posterior clade probabilities. Nodes with less than 50% bootstrap support (left) or 50% posterior probability (right) collapsed. Shaded area indicates the subgenus Arctoraja. Genus names are as follows: A. = Arctoraja, B. = Bathyraja, R. = Raja, Rhino. = Rhinoraja
The topology recovered in the Bayesian analysis is almost completely congruent with the parsminony analysis. The topology within the subgenus Arctoraja is identical in both analyses. The Arctoraja clade is well supported by Bremer values, bootstrap values, and posterior clade probabilities, as are the B. interrupta clade and the B. matsubarai clade. The genus Bathyraja is again recovered as paraphyletic due to the inclusion of R. longicauda. The only topological incongruence within the in-group between the two trees is the placement of B. fedorovi and B. trachura. In the Bayesian tree, Bathyraja fedorovi is recovered as sister to B. trachura plus the remainder of the ingroup with high posterior support (posterior clade probability = 1.0).
Discussion
Our genetic analysis of cytochrome oxidase (COI) in 226 individuals of the Bathyraja parmifera species complex revealed three distinct haplotypes, one of which is geographically distinct from the other two. Due to a unique combination of genetic, morphological, and meristic characters, specimens with the distinct haplotype will be described as a new species, now commonly referred to as the leopard skate (Stevenson et al. 2007; Orr et al. in press). The species presently recognized as B. parmifera exhibits two haplotypes, one much more common than the other, with broadly overlapping geographic ranges. Genetic differentiation among the haplotypes of the subgenus Arctoraja is notably low (K2P distance 0.000–0.015). In fact, we found identical sequences for the two western Pacific representatives of the subgenus (B. simoterus and B. smirnovi). Chondrichthyan mitochondrial DNA appears to be more highly conserved than in teleost fishes (Ward et al. 2008), and differentiation among species of Bathyraja is generally lower than for other chondrichthyans (Spies et al. 2006). Although it is rare for COI sequences from two different species to be identical, two other cases have been reported for elasmobranchs; Bathyraja lindbergi and Bathyraja maculata, Urolophus sufflavus and Urolophus cruciatus (Spies et al. 2006; Ward et al. 2008). The apparently fixed nucleotide differences between the leopard skate and the two haplotypes of B. parmifera provide a means of discriminating between the two species. In addition, the sequence divergence between the two haplotypes of B. parmifera is equivalent to that observed between B. parmifera and the leopard skate, which suggests the possibility of additional cryptic speciation in the genus. To date, photos but no morphological samples of the less common B. parmifera haplotype have been preserved, and further work will be required to evaluate their taxonomic status.
The hallmark of allopatric speciation is the presence of sister species distributed across a geographic barrier. This signature is clear in the distribution and close genetic and morphological relationship between the leopard skate and B. parmifera. Our results suggest that the leopard skate is restricted to the west of Amchitka Pass in the Aleutian Islands, while the common haplotype of B. parmifera is found east of Amchitka Pass in the central and eastern Aleutian Islands, and the rare haplotype is found only east of Samalga Pass in the eastern Aleutian Islands. Both haplotypes of B. parmifera are found in the Bering Sea and Gulf of Alaska. This distribution pattern may indicate that these species do not regularly cross deepwater passes. In general, the Aleutian Islands are characterized by narrow coastal areas surrounding islands with three major regions being demarcated by deepwater passes of 400 m depth or deeper (Logerwell et al. 2005). Skates reproduce by depositing demersal egg cases which remain in nursery areas until hatching (Hoff 2008). Because of a benthic lifestyle at all life history stages, and a bathymetric distribution typically more shallow than the passes, migration—and thus gene flow—of shallow-water skates may be restricted by deepwater passes.
Our analysis was based on samples of all species of Bathyraja from Alaska, several species from the western Pacific, representative species from each of the major oceans, and three outgroup species. Although COI is commonly used to distinguish between species, this region of the mtDNA genome provides a limited amount of DNA sequence data for phylogenetic analysis. Our phylogenetic analysis supports a monophyletic group within Bathyraja consisting of the leopard skate, B. parmifera, B. simoterus, and B. smirnovi, and therefore supports the recognition of the subgenus Arctoraja (Orr et al. in press). It also suggests that the B. matsubarai clade and the B. interrupta clade may merit consideration for subgeneric recognition due to the high level of support for these clades (Fig. 2). The morphological similarities of the B. matsubarai group were recognized by Ishiyama and Ishihara (1977), who noted that the configuration of the clasper cartilages in these species is remarkably similar. The B. interrupta group also exhibits some morphological similarities, most notably the complete absence of a pseudosiphon on the clasper (Ishiyama and Ishihara 1977; Stevenson et al. 2004).
Another interesting result of our phylogenetic analysis is the placement of Rhinoraja longicauda within Bathyraja, which renders the genus Bathyraja paraphyletic. Rhinoraja has been distinguished from Bathyraja on the basis of three characters: a long tail with a relatively high precaudal vertebral count (74–91), the presence of a generalized clasper with pseudosiphon, and the presence of a segmented rostral cartilage (Ishiyama 1952, 1958). The first two of these characters are also found in other members of the genus Bathyraja (Ishiyama 1958), and the segmented rostral cartilage is most likely an autapomorphic character; thus, there is no morphological evidence at present that would definitively place Rhinoraja outside the Bathyraja lineage. Rather, our analysis supports the hypothesis that Rhinoraja diverged from a lineage within the Bathyraja. Some authors have included B. taranetzi and even B. interrupta within the genus Rhinoraja (e.g., Raschi and McEachran 1991; Jamieson and Taylor 1997; McEachran and Dunn 1998; Compagno 1999; Dolganov 1999). This is plainly inappropriate with respect to B. interrupta, as the species does not have an elevated precaudal vertebral count, a pseudosiphon, or a segmented rostral cartilage, and the molecular phylogeny presented here indicates that B. interrupta is not closely related to Rhinoraja. With respect to B. taranetzi, the situation is less clear. This species does have a long tail with a high precaudal vertebral count (68–74) and a pseudosiphon on the clasper, but the presence of a segment in the rostral cartilage is questionable (see Stevenson et al. 2004). Unfortunately, the molecular phylogeny presented here does not provide any additional insight on this, as the positions of both B. taranetzi and R. longicauda are poorly resolved in both the parsimony and Bayesian analyses. A more intensive study, including more species of Bathyraja and Rhinoraja, as well as additional sequence data, is required to resolve this and other issues related to the phylogeny and classification of these fishes.
References
Bremer K (1988) The limits of amino acid sequence data in angiosperm phylogenetic reconstruction. Evolution 42:795–803
Chase MR, Etter RJ, Rex MA, Quattro JM (1998) Extraction and amplification of mitochondrial DNA from formalin-fixed deep sea mollusks. Biotechniques 24:243–247
Compagno LJV (1999) Checklist of living elasmobranchs. In: Hamlett WC (ed) Sharks, skates, and rays: the biology of elasmobranch fishes. The Johns Hopkins University Press, Baltimore, Maryland, pp 471–498
Dolganov VN (1999) Geographic and bathymetric distribution of the skates of the Rajidae family in the far eastern seas of Russia and adjacent waters. J Ichthyol 39:340–342
France SC, Kocher TD (1996) DNA sequencing of formalin-fixed crustaceans from archival research collections. Mar Mol Biol Biotechnol 5:304–313
Hall TA (1999) BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl Acids Symp Ser 41:95–98
Hasegawa M, Yano T, Kishino H (1984) A new molecular clock of mitochondrial-DNA and the evolution of hominoids. Proc Jpn Acad Ser B: Phys Biol Sci 60:95–98
Hebert P, Cywinska A, Ball S, deWaard J (2003) Biological identifications through DNA barcodes. Proc R Soc Lond 270:313–321
Hendy MD, Penny D (1982) Branch and bound algorithms to determine minimal evolutionary trees. Math Biosci 59:277–290
Hoff GR (2008) A nursery site of the Alaska skate (Bathyraja parmifera) in the eastern Bering Sea. Fish Bull 106:233–244
Huelsenbeck JP, Ronquist F (2001) MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 17:754–755
Ishiyama R (1952) Studies on the rays and skates belonging to the family Rajidae, found in Japan and adjacent regions. 4. A revision of three genera of Japanese rajids with descriptions of one new genus and four new species mostly occured in northern Japan. J Shimonoseki Coll Fish 2:1–34
Ishiyama R (1958) Studies on the rajid fishes (Rajidae) found in the waters around Japan. J Shimonoseki Coll Fish 7:193–394
Ishiyama R, Ishihara H (1977) Five new species of skates in the genus Bathyraja from the western North Pacific, with reference to their interspecific relationships. Jpn J Ichthyol 24:71–90
Jamieson A, Taylor SC (1997) Comparisons of three probability formulae for parentage exclusion. Animal Gen 28:397–400
Kass RE, Raftery AE (1995) Bayes factors. J Am Stat Assoc 90:773–795
Kimura M (1980) A simple method of estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J Mol Evol 16:111–120
Logerwell EA, Aydin K, Barbeaux S, Brown ES, Conners ME, Lowe S, Orr JW, Ortiz I, Reuter R, Spencer P (2005) Geographic patterns in the demersal ichthyofauna of the Aleutian Islands. Fish Oceanog 14:93–112
McEachran JD, Dunn KA (1998) Phylogenetic analysis of skates, a morphologically conservative clade of elasmobranchs (Chondrichthyes: Rajidae). Copeia 1998:271–290
Nei M, Kumar S (2000) Molecular evolution and phylogenetics. Oxford University Press, Oxford
Nylander JA (2004) MrModeltest v2 (program distributed by the author). Evolutionary Biology Centre, Uppsala University, Sweden
Orr JW, Stevenson DE, Hoff GR, Spies I, McEachran JD (in press) Bathyraja panthera, a new species of skate (Rajidae: Arhynchobatinae) from the western Aleutian Islands, and resurrection of the subgenus Arctoraja Ishiyama. NOAA Prof Pap NMFS
Raschi WG, McEachran JD (1991) Rhinoraja longi, a new species of skate from the outer Aleutian Islands, with comments on the status of Rhinoraja (Chondrichthys, Rajoidei). Can J Fish Aquat Sci 69:1889–1903
Ronquist F, Huelsenbeck JP (2003) MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 19:1572–1574
Rozas J, Sanchez-DelBarrio JC, Messeguer X, Rozas R (2003) DnaSP, DNA polymorphism analyses by the coalescent and other methods. Bioinformatics 19:2496–2497
Smith PJ, Steinke D, Mcveagh SM, Stewart AL, Struthers CD, Roberts CD (2008) Molecular analysis of Southern Ocean skates (Bathyraja) reveals a new species of Atlantic skate. J Fish Biol 73:1170–1182
Sorenson MD (1999) TreeRot, version 2. Boston University, Boston
Spies IB, Gaichas S, Stevenson DE, Orr JW, Canino M (2006) DNA-based identification of Alaska skates (Amblyraja, Bathyraja and Raja: Rajidae) using cytochrome c oxidase subunit I (coI) variation. J Fish Biol 69:283–292
Stevenson DE, Orr JW, Hoff GR, McEachran JD (2004) Bathyraja mariposa: a new species of skate (Rajidae: Arhynchobatinae) from the Aleutian islands. Copeia 2004:305–314
Stevenson DE, Orr JW, Hoff GR, McEachran JD (2007) Field guide to sharks, skates, and ratfish of Alaska. Anchorage, Alaska
Swofford DL (2003) PAUP*. Phylogenetic analysis using parsimony (*and other methods), version 4. Sinauer Associates, Sunderland
Tamura K, Dudley J, Nei M, Kumar S (2007) MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol Biol Evol 24:1596–1599
Ward RD, Holmes BH, White WT, Last PR (2008) DNA barcoding Australasian chondrichthyans: results and potential uses in conservation. Mar Freshw Res 59:57–71
Acknowledgments
We would like to thank the Alaska Fisheries Science Center and the University of Washington Fish Collection for their support, as well as those who contributed to sample collection. We would also like to thank Michael Canino (AFSC), Christopher Kenaley (UW), Beth Matta (AFSC), Bobette Dickerson (AFSC), Katherine Maslenikov (UW), Thomas Hollowed (AFSC), Theodore Pietsch (UW), Paola Pasolini (University of Bologna), Gento Shinohara (NSMT), Mamoru Yabe (HUMZ), and Kazuhiro Nakaya (HUMZ). This research is contribution EcoFOCI-0741 to the NOAA’s Fisheries-Oceanography Coordinated Investigations.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
About this article
Cite this article
Spies, I.B., Stevenson, D.E., Orr, J.W. et al. Molecular systematics of the skate subgenus Arctoraja (Bathyraja: Rajidae) and support for an undescribed species, the leopard skate, with comments on the phylogenetics of Bathyraja . Ichthyol Res 58, 77–83 (2011). https://doi.org/10.1007/s10228-010-0185-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10228-010-0185-z