
Abstract
Riparian forest soils can be highly dynamic, due to frequent fluvial disturbance, erosion, and sediment deposition, but effects of dams on riparian soils are poorly understood. We examined soils along toposequences within three river segments located upstream, between, and downstream of two dams on the Elwha River to evaluate relationships between riparian soil development and forest age, succession, and channel proximity, explore dam effects on riparian soils, and provide a baseline for the largest dam removal in history. We found that older, later-successional forests and geomorphic surfaces contained soils with finer texture and greater depth to cobble, supporting greater forest floor mass, mineral soil nutrient levels, and cation exchange. Forest stand age was a better predictor than channel proximity for many soil characteristics, though elevation and distance from the channel were often also important, highlighting how complex interactions between fluvial disturbance, sediment deposition, and biotic retention regulate soil development in this ecosystem. Soils between the dams, and to a lesser extent below the lower dam, had finer textures and higher mineral soil carbon, nitrogen, and cation exchange than above the dams. These results suggested that decreased fluvial disturbance below the dams, due to reduced sediment supply and channel stabilization, accelerated soil development. In addition, reduced sediment supply below the dams may have decreased soil phosphorus. Soil δ 15N suggested that salmon exclusion by the dams had no discernable effect on nitrogen inputs to upstream soils. Recent dam removal may alter riparian soils further, with ongoing implications for riparian ecosystems.
Similar content being viewed by others

Introduction
Riparian soils along unregulated rivers often display high spatial and temporal variability in texture, organic matter, and nutrient availability, reflecting variation in proximity to the channel, time since disturbance, woody debris, sediment deposition, and successional stage (Naiman and others 2005; Naiman and others 2010). Primary-successional riparian surfaces are created frequently by channel change and bar formation during large floods (Meyer 2001; Daniels 2003). Following major fluvial disturbances, riparian soils can develop rapidly because they receive large water, sediment, and nutrient inputs from the river and surrounding watershed, creating hotspots of nutrient availability and cycling, primary productivity, and organic carbon stores (Pinay and others 2002; McClain and others 2003; Harms and Grimm 2008; Cabezas and Comin 2010; Ricker and others 2013).
On young fluvial surfaces, fine sediment deposition during floods is often the primary source of organic matter and nutrients for soil formation (Steiger and Gurnell 2003; Adair and others 2004; Bechtold and Naiman 2009; Cabezas and Comin 2010; Batz and others 2015), although floods also erode riparian sediments (Walker 1989; Toda and others 2005; Saint-Laurent and others 2014). Early-successional, nitrogen-fixing plants also contribute substantial nitrogen to riparian soils in many regions (Luken and Fonda 1983; Rhoades and others 2008; Scott and others 2008), and surface water and groundwater provide additional inputs (Andersen and others 2003; Morris and others 2010; Koyama and Kielland 2011). Over time, early-successional vegetation can accelerate succession, by slowing flood flows and increasing fine sediment deposition, which increases surface elevations, nutrient availability, and plant growth, further slowing floods and enhancing fine sediment deposition (Gurnell and Petts 2006; Griffin and others 2014; Le Bouteiller and Venditti 2014). As higher floodplain surfaces develop and flood less often, detrital inputs from riparian plants become the primary source of organic matter and nutrients (Bechtold and Naiman 2009; Cabezas and Comin 2010; Batz and others 2015). However, riparian soils often do not exhibit clear successional chronosequences, both because large floods bury or remove patches of soil on older surfaces (Hill 2011; Appling and others 2014; Batz and others 2015) and because patchiness in fine sediment deposition and erosion on young surfaces continues to affect organic matter and nutrient retention as surfaces age (Bechtold and Naiman 2009; Southwell and Thoms 2011; Batz and others 2015).
Dams have the potential to alter riparian soils substantially, particularly along rivers with historically large floods or sediment transport. Dams often reduce flood frequency and intensity, capture sediment and woody debris, and alter the timing and magnitude of low flows, affecting riparian spatial and temporal dynamics, community composition, and ecosystem function (Nilsson and Berggren 2000; Graf 2006; Poff and Zimmerman 2010). These changes may alter riparian soils in numerous and contrasting ways. Both lower flood magnitudes and lower sediment supply below dams may slow accumulation of soil nutrients and organic matter by reducing fine sediment, organic matter, and nutrient inputs to both young and older surfaces (Pinay and others 2002; Beauchamp and others 2007; Kunz and others 2011a; Kunz and others 2011b), especially where sediment from downstream tributaries or erodible floodplains is limited (Phillips and others 2004). At the same time, however, lower flood magnitudes and lower sediment supply below dams may result in more rapid and spatially uniform accumulation of soil nutrients and organic matter, because lower flood magnitudes can reduce fluvial disturbance and erosion of developing soils, and lower sediment supply can induce channel armoring, incision, and stabilization and reduce coarse sediment deposits that bury developing soils (Friedman and others 1998; Adair and others 2004; Grams and Schmidt 2005; Hupp and others 2015). Reduced fluvial disturbance below dams also may reduce abundance of early-successional nitrogen-fixing plants, reducing nitrogen inputs (Mallik and Richardson 2009; Perakis and others 2011). In regions with large anadromous fish populations (for example, salmon in northwestern North America), dams that prevent fish passage may reduce nutrient inputs from decomposing fish carcasses (Helfield and Naiman 2001; Drake and others 2005; Morris and Stanford 2011). Finally, changes in low flows due to dams may affect soil moisture and primary productivity, and hence decomposition and nutrient cycling (Adair and others 2004). Different effects may predominate on different geomorphic surfaces, especially given the large impacts of frequent fluvial disturbance and coarse sediment deposition on young pioneer bars and greater importance of fine sediment deposition in developing floodplain forests. However, effects of dams on riparian soils have rarely been studied.
To gain insights into riparian soils along regulated rivers, we examined soils upstream, between, and downstream of two moderately sized dams along the Elwha River in the Pacific Coastal Ecoregion of northwestern North America (Figure 1). Effects of the dams on fluvial processes differed between the segments downstream of the dams, which allowed us to evaluate the support for different mechanisms of dam effects. Neither dam altered peak flows substantially during their 85-year (upper dam) and 98-year (lower dam) operations, but the upper dam sometimes diminished or augmented low flows (Pohl 1999; Duda and others 2008). More importantly, both dams restricted downstream sediment and wood transport, trapping an estimated 21 ± 4 million m3 of sediment (Randle and others 2015). Low sediment supply downstream of the dams contributed to armoring and stabilization of the active channel between the dams and immediately downstream of the lower dam, thus restricting channel migration and avulsion (Pohl 2004; Draut and others 2011). In contrast, farther downstream of the lower dam, within 5 km of the river mouth, local sediment recruitment was sufficient to allow considerable channel mobility during high flow events, with greatest mobility within 1 km of the mouth (Pohl 2004; Draut and others 2011). Consequently, riparian surfaces between the dams, and to a lesser extent below the lower dam, tended to be composed of fewer pioneer bars and more terraces than above the dams (Shafroth and others 2017). Further, riparian forests between the dams tended to be older, with higher abundance of mid- and later-successional communities, lower abundance of early-successional communities, and fewer areas for early-successional seedling recruitment than above the dams (Kloehn and others 2008; Shafroth and others 2017).

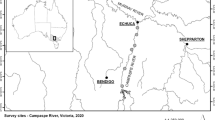
Locations of 15 transects sampled on the upper, middle, and lower segments of the Elwha River, in relation to the Elwha and Glines Canyon dams and associated reservoirs. Upper segment transect locations were 31.7, 30.6, 30.4, 29.0, and 28.3 km upstream of the river mouth, with cross valley widths of 322, 437, 412, 390, and 319 m, respectively. Middle segment transect locations were 21.2, 19.5, 18.5, 17.1, and 15.8 km upstream of the river mouth, with cross valley widths of 319, 401, 557, 534, and 550 m, respectively. Lower segment transect locations were 6.0, 4.5, 3.4, 2.6, and 1.8 km upstream of the river mouth, with cross valley widths of 430, 476, 1174, 819, and 1027 m, respectively. Only perennial tributaries are shown.
Understanding the effects of the Elwha River dams on riparian soils first requires understanding how fluvial processes and forest succession affect soils along this particular river. Riparian soil development varies considerably among rivers depending on complex interactions between flow regime, sediment supply, nutrient budgets, and floodplain geomorphology. To gain insights into the primary drivers of spatial variation in riparian soils along the Elwha River, we examined relationships between riparian soil chemistry and texture and forest age, successional stage, and channel proximity along 15 toposequences. Further, we used model selection criteria to determine the most useful predictors of riparian soil characteristics.
To explore potential dam effects on riparian soils, we compared soil characteristics upstream, between, and downstream of the dams, and compared relationships between soil characteristics and forest age among the three river segments. Higher nutrient levels and finer sediments upstream of the dams would support the hypothesis that lower sediment supply below the dams predominantly slowed soil development, by reducing fine sediment and nutrient inputs. In contrast, higher nutrient levels and finer sediments between the dams would support the hypothesis that lower sediment supply, and hence reduced channel mobility, between the dams predominantly promoted soil development, by increasing retention of developed riparian soils. Larger differences in soils between younger and older surfaces upstream of the dams than downstream would support the hypothesis that lower sediment supply below the dams slowed soil development more on younger surfaces than on older surfaces, by reducing fine sediment and nutrient inputs. In contrast, smaller differences in soils between younger and older surfaces upstream of the dams would support the hypothesis that lower sediment supply below the dams predominantly promoted soil development on younger surfaces by reducing channel mobility and increasing soil retention, and also may have slowed soil development on older surfaces by reducing fine sediment and nutrient inputs. Higher δ 15N and nutrient levels downstream of the dams would support the hypothesis that salmon exclusion reduced upstream nutrient inputs, as δ 15N is elevated in anadromous salmon (Mathisen and others 1988). The dams have since been removed, constituting the largest dam removal to date, so our results also provide a baseline for future studies of dam removal effects on riparian soils and ecosystem recovery.
Methods
Study Area
The Elwha River drains 833 km2 of mountainous, forested terrain primarily in Olympic National Park, Washington, USA, dropping approximately 1,370 m along its 72-km length to the Strait of Juan de Fuca (Figure 1). Annual precipitation declines from about 550 to 100 cm from the headwaters to the mouth (Phillips and Donaldson 1972). Average instantaneous discharge is 43 m3 s−1, the 2-year flood discharge is 400 m3 s−1, and the 100-year flood discharge is 1240 m3 s−1 (Duda and others 2011b). Large-magnitude, short-duration floods occur during winter rainstorms, whereas smaller-magnitude, longer-duration floods occur during snowmelt in late spring and early summer (Draut and others 2011).
The Elwha Dam, 32-m high, was constructed 7.9 km from the river mouth in 1913. Glines Canyon Dam, 64-m high, was constructed 21.6 km from the mouth in 1927. Elwha Dam operations were always predominantly “run-of-the-river,” with roughly equal inflows and outflows. In contrast, Glines Canyon Dam operations resulted in significant flow fluctuations from 1927 to 1975, were largely run-of-the-river from 1975 to 1999, and were adjusted to augment September and October low flows to benefit native fish beginning in 2000 (Duda and others 2011b). Both dams were removed between 2011 and 2014 (Warrick and others 2015).
We measured riparian soil properties in August 2007 along three river segments: an unregulated segment above both dams, a regulated segment between the dams, and a regulated segment below both dams (hereafter the “upper,” “middle,” and “lower” segments, respectively). The upper segment flowed about 4 km through an alluvial valley between two bedrock canyons about 32–28 river km above the mouth, with a predominantly island-braided channel. The middle segment flowed about 6 km through an alluvial valley beginning approximately 200 m downstream of Glines Canyon Dam, below a short, narrow bedrock canyon, and ending at the Olympic National Park boundary, with a single or anabranching gravel-bed channel. The lower segment flowed about 5 km through an alluvial valley beginning about 1 km downstream of Elwha Dam, below a narrow bedrock canyon, and ending about 2 km upstream of the mouth, with an anabranching gravel-bed channel. The upper segment valley was entirely unconfined, and the middle and lower segment valleys were largely unconfined, with narrower valley widths (<150 m) along only about 400 and 200 m of their river lengths, respectively.
Soil Measurements
Soils were sampled in 55 forested plots, a representative subset of 117 plots established to study riparian ecosystem dynamics along the Elwha River (Shafroth and others 2017). The plots were located along five cross-valley transects within each segment, selected to sample toposequences representing the range of geomorphic conditions common to the Elwha River (Figure 1). All transects were located within unconfined reaches. Plots were placed randomly along the transects within patches distinguished by overstory vegetation and geomorphic position at elevations 0.3–6.7 m above the main channel water surface. Plots varied in size from 50 to 250 m2 (5 × 10–10 × 25 m), with the long side placed perpendicular to the transect line; smaller plots were placed in homogeneous patches with high tree densities. N = 16, 20, and 19 for the upper, middle, and lower segments, respectively.
Soil samples were collected from two points within each plot, midway between the upstream and downstream plot edges, and either 10 or 20 m apart. At each point, a forest floor (that is, O horizon) sample of all dead, less-than-1-cm diameter, organic material was collected from a 20 × 20-cm2 subplot. Then, a 10-cm-deep mineral soil sample was collected, in most cases with a 6.9-cm diameter corer. In gravelly soils, a 50 × 50-cm2 pit was excavated; the sediment was divided into three size classes (<16-, 16–64-, and >64-mm) and weighed; and 1 kg of the less-than-16-mm fraction was retained for analysis. Soil samples were sealed in plastic bags, refrigerated, shipped to the laboratory, and processed within 6 d of collection. Pit volume was estimated from direct measurements, or for irregularly shaped pits by lining the pit with a plastic sheet and measuring the water volume required to fill the pit. The depth of soil above cobble and gravel deposits was estimated by depressing a 0.6-cm diameter, 1.2-m steel probe as deeply as possible in two to four locations near each sampling point; measurements were averaged for each plot. In some cases, the probe may have stopped at coarse roots.
In the laboratory, samples from each plot were pooled and sieved into 2–16-mm and less-than-2-mm size classes. The 2–16-mm class was separated into rocks and organic detritus. Laboratory analyses were conducted on the forest floor and less-than-2-mm mineral soil fraction. Moisture content was determined for forest floor samples by drying at 65°C for 48 h, and for mineral soil by drying at 105°C for 48 h. Mineral soil texture was determined using the hydrometer method. Forest floor and mineral soil total [C] and [N] were determined by drying subsamples at 65°C, grinding to fine powder, and analyzing with a Costech ECS-4010 elemental combustion analyzer. Forest floor [P], [K], [Ca], [Mg], and [Na] were determined by digesting ground samples in nitric acid and analyzing with Perkin Elmer Optima inductively coupled plasma optical emission spectrometry. Because forest floor samples were often contaminated with mineral soil, they were ashed to determine % organic matter. Forest floor nutrient concentrations were calculated per unit mass organic matter. Forest floor nutrient pools were calculated as the product of organic matter-adjusted forest floor mass and nutrient concentrations. On mineral soil samples, potential net N mineralization and percent nitrification of mineralized N were determined by incubating 10 g fresh soil at 25°C and 60% water holding capacity for 28 d; both initial and final NH4 + and NO3 − were extracted from 7 g soil for 1 h with 35 ml 0.5 M K2SO4, filtered through Whatman 42 filters, and analyzed colorimetrically using a Lachat 8000 flow-injection autoanalyzer. Mineral soil Bray-1 P was extracted from 5 g field-moist soil for 1 min with 25 ml 0.3 N NH4F and 0.025 N HCl, centrifuged at 3400 rpm for 5 min, filtered through Whatman 42 filters, and analyzed colorimetrically for soluble reactive P via autoanalyzer. Forest floor and mineral soil C:N (mass-based) were calculated from total [C] and [N]. Forest floor N:P was calculated from total [N] and [P], whereas mineral soil N:P was calculated from initial inorganic [N] and available Bray-1 [P]. Mineral soil Ca, Mg, K, and Na were extracted from 2 g field-moist soil with 40 ml unbuffered 1 M ammonium acetate, and analyzed using inductively coupled plasma optical emission spectrometry. After base cation extraction, cation exchange capacity (CEC) was determined by displacing ammonium with 0.1 N HCl, followed by colorimetric ammonium analysis. Base saturation was estimated as the sum of extractable Ca, Mg, K, and Na, divided by CEC. Field-moist mineral soil solution pH (2 water:1 soil) was assayed using an Accumet pH probe.
Mineral soil total bulk density (hereafter BDT) was calculated as the total sample mass (including all particle sizes), corrected for soil moisture, and divided by the total sample volume. Fine bulk density (hereafter BDF) was calculated as the dry mass of the less-than-2-mm fraction, divided by the total sample volume. Mineral soil nutrient pools to a 10-cm depth were calculated as the product of BDF, 10 cm, and nutrient concentrations.
Throughout, forest floor and mineral soil measurements are distinguished with subscripts “FF” and “MS,” nutrient concentrations are indicated with square brackets, and nutrient pools are indicated with subscript “(pool).”
Plant Community and Topographic Measurements
Percent cover, stem density, and basal area of overstory trees in each plot were measured in 2003 and 2004. Ocular, ground-based estimates of overstory cover for each species were recorded using the midpoints of Daubenmire cover classes: 0–5% = 2.5%, 5–25% = 15.5%, 25–50% = 37.5%, 50–75% = 62.5%, 75–95% = 85%, 95–100% = 97.5% (Daubenmire 1959). Woody stems greater-than-5-cm DBH were identified to species, counted, and measured to calculate stem densities and basal areas. Overstory species importance was calculated as the sum of relativized cover, relativized stem density, and relativized basal area in each plot, with each measure relativized as a percent of the total for all species in the plot. Stand age was estimated by counting annual rings on increment bores collected from the largest individuals of characteristic species within or adjacent to the plots, with years added to account for time required for individuals to grow from the ground surface to the coring height. For six plots from which cores were not collected, ages of the largest trees were estimated using size–age regressions for Alnus rubra Bong. (red alder), Populus balsamifera L. ssp. trichocarpa (Torr. & A. Gray ex Hook.) Brayshaw (black cottonwood), or Pseudotsuga menziesii (Mirb.) Franco (Douglas fir) (Shafroth and others 2017).
In a separate paper (Shafroth and others 2017), the 117 plots of the larger study were assigned to 11 overstory community types identified by hierarchical cluster analysis of log-transformed overstory species importance. The cluster analysis was performed using Sørenson (Bray–Curtis) distances and flexible beta = −0.25. The choice of where to “prune” the resulting tree was made such that all clusters represented clear and ecologically informative differences in composition. Misclassified plots were identified using non-parametric discriminant analysis. The resulting community types represented a successional gradient from early-successional stands dominated by Salix spp. L. (willow) and A. rubra to later-successional stands dominated by conifers (P. menziesii, Thuja plicata Donn ex D. Don (western redcedar), Tsuga heterophylla (Raf.) Sarg. (western hemlock)) and/or maples (Acer macrophyllum Pursh (bigleaf maple), Acer circinatum Pursh (vine maple)). Community types were named based on indicator species analysis, for species with indicator values above 15. Nomenclature follows the USDA plants database (http://plants.usda.gov).
For the present study, to accommodate our smaller sample size, the 11 community types were lumped into seven (Table 1; Figure S1); a total of six plots were reclassified, by combining the two earliest-successional types (Alnus–Salix + no overstory), combining two later-successional types dominated by A. macrophyllum and T. plicata (Thuja–Acer + Acer–Populus–Thuja), and combining three later-successional types dominated by P. menziesii and either A. grandis, A. circinatum, or T. heterophylla (Pseudotsuga–Abies + Pseudotsuga–Acer + Pseudotsuga–Tsuga). These lumpings were supported by proximity of the community types in NMDS ordination space for log-transformed overstory species importance (Shafroth and others 2017). NMDS ordination of the 117 plots, using Sørenson (Bray–Curtis) distances and Varimax axis rotation, identified separate axes for late-successional P. menziesii-dominated plots, late-successional A. macrophyllum-dominated plots, and late-successional P. balsamifera-dominated plots, describing 38, 34, and 19% of the variance, respectively. Plots containing the Pseudotsuga–Abies, Pseudotsuga–Acer, and Pseudotsuga–Tsuga community types were tightly grouped in NMDS ordination space, as were plots containing the Thuja–Acer, Acer–Populus–Thuja, and to a lesser extent Acer community types.
Among the resulting seven community types, Alnus–Salix plots contained Salix spp. and were usually dominated by Salix spp., and/or A. rubra. Alnus plots were dominated by A. rubra alone. Alnus–Populus plots were co-dominated by P. balsamifera and A. rubra. Pseudotsuga–Alnus plots were co-dominated by P. menziesii and A. rubra; P. balsamifera and A. grandis were also abundant in some plots. Pseudotsuga–Abies plots were dominated by P. menziesii without A. rubra; P. balsamifera, A. grandis, A. circinatum, A. macrophyllum, and T. heterophylla were abundant in some plots. Acer plots were co-dominated by A. macrophyllum, A. circinatum, and sometimes A. rubra. Thuja–Acer plots contained T. plicata and were usually co-dominated by T. plicata and A. macrophyllum; A. circinatum, A. rubra, and P. balsamifera were also abundant in some plots.
Elevation above and distance from the nearest primary channel and the nearest channel with perennial streamflow were measured for each plot using a Leica TCR705 total station in 2003 and 2004. We interpreted proximity to the nearest primary channel as indicative of proximity to fluvial disturbance (hereafter elevationdisturbance and distancedisturbance), and proximity to the nearest channel with perennial flow as indicative of proximity to the water table (hereafter elevationgroundwater and distancegroundwater). Because initial surveys occurred across a range of stream discharges, the relative elevation of surveyed points along the transects was tied to a final survey during a low flow period in August 2005, during which instantaneous discharge varied by <0.03 m3 s−1 at our transects.
Soil Stable Isotopes
In a separate sampling effort aimed at characterizing sources and movement of soil C and N along the river, natural abundance isotopes of soil C and N were measured at 42 sampling points within 11 reaches along the three segments in 2006 and 2007 (Table 1). Sampling points were located less than 150 m from the active channel within three vegetation types: (1) gravel bars with establishing A. rubra and Salix spp. saplings (<10-m tall, with silt or sand soils), (2) mature A. rubra stands (>10-m tall, >20-cm DBH, with organic soils), and (3) mature conifer stands without A. rubra. At each point, three widely spaced, 10-cm-deep, mineral soil samples were collected with a corer and composited. A subsample was dried at 65°C and analyzed at the Colorado Plateau Stable Isotope Laboratory at Northern Arizona University for δ 13C and δ 15N, with values expressed using standard ‘delta’ (δ) notation relative to international standards.
Statistical Analyses
To condense the 49 soil measures (listed in Tables 2, 3) into fewer, easily interpreted variables, the measures were ordinated with non-metric multidimensional scaling (NMDS) in PC-ORD 5.1, using Sørenson (Bray–Curtis) distances, random starting configurations, 250 runs with real data, 250 iterations with the best starting configuration for the final solution, and Varimax rotation to maximize correspondence between ordination axes and soil measures. Prior to ordination, the data were relativized by the maximum for each measure. Scree plots were examined to select between one and six dimensions.
To examine relationships between riparian soils and other ecosystem properties, correlations between individual soil measures, NMDS ordination scores, stand age, channel proximity, and overstory species importance were evaluated using Kendall’s tau analyses, with |τ| > 0.3 indicating strong correlations.
To explore possible dam effects, individual soil measures and NMDS ordination scores were compared among the three segments using PROC MIXED in SAS 9.3 with the maximum likelihood method, with segment as a fixed effect, transect (nested within segment), and patch (nested within segment and transect) as random effects, degrees of freedom calculated using the Kenward–Roger method, and post hoc means comparisons conducted with Tukey-adjusted LSmeans. In separate analyses, (1) overstory community type was included as an additional fixed effect to test for differences among community types, (2) stand age was included as a fixed effect to test for effects of stand age on soil characteristics and to test whether differences in soils among segments were robust to accounting for differences in stand age, and (3) both stand age and segment × stand age were included as fixed effects to test for differences among segments in the relationship between soil characteristics and stand age. Analyses of stable isotopes were similar but included segment and vegetation type as fixed effects and year and reach (nested within segment) as random effects.
To meet model assumptions, stand age, elevationdisturbance, elevationgroundwater, distancedisturbance, distancegroundwater, depth to cobble, forest floor mass, [K]FF, CFF(pool), NFF(pool), PFF(pool), KFF(pool), CaFF(pool), MgFF(pool), NaFF(pool), [C]MS, [N]MS, [P]MS, [Mg]MS, CMS(pool), NMS(pool), PMS(pool), CaMS(pool), MgMS(pool), and BDT were log-transformed for analysis. N mineralization, [K]MS, [Ca]MS, and KMS(pool) were square-root transformed. Percent nitrification was arcsin, square-root transformed. Percent composition by 16–64-mm particles and less-than-64-mm particles included many zeros, so were log-transformed and analyzed using PROC GLIMMIX in SAS 9.3 with a Poisson distribution and the Laplace method.
Finally, the relative importance of stand age, elevationdisturbance, elevationgroundwater, distancedisturbance, and distancegroundwater for predicting each ordination axis and soil measure was evaluated by comparing AICc scores (Akaike Information Criterion corrected for finite sample size) among 18 models (listed in Table 4), using the same statistical methods as the segment comparisons described above. Models that included both measures of elevation or distance were excluded because of substantial collinearity. Variance inflation factors calculated using PROC REG in SAS 9.3 indicated negligible collinearity (VIF <1.7) in the 18 models for all soil measures. To avoid discussing poorly supported models, model selection results were not reported for soil variables with evidence ratios below 10 for the null model relative to the best model. Generally, w i of the best models were not sufficiently greater than those of other models to select a best model with confidence. Therefore, the relative importance of proximity to the nearest primary channel versus the nearest active channel, and of elevation above versus distance from the channel, also was evaluated by comparing \(\sum {w_{i} }\) for the different predictors.
Results
NMDS Ordination
NMDS ordination yielded three axes describing 58% (axis 1), 29% (axis 2), and 4% (axis 3) of the variance (total R 2 = 91%), with a final stability of less than 0.000005 and final stress of 11.4 (Figure 2). Both axis 1 and axis 2 captured variation in forest floor mass and nutrient pools and mineral soil nutrient concentrations; high axis 1 and 2 scores corresponded to high forest floor mass and nutrient pools (except KFF(pool)), and high mineral soil C:NMS (mass-based), [C]MS, [N]MS, [Ca]MS, [Mg]MS, CEC, and moisture (Tables 2, 3). In addition, axis 1 captured variation in mineral soil nutrient pools and texture; high axis 1 scores corresponded to high CMS(pool), NMS(pool), KMS(pool), CaMS(pool), MgMS(pool), KFF(pool), and [K]MS, as well as high depth to cobble, clay, silt, and less-than-2-mm particles and low BDT, sand, and 16–64 and greater-than-64-mm particles. In contrast, axis 2 captured variation in N cycling; high axis 2 scores corresponded to high N mineralization and low BDF and PMS(pool). Axis 3 described variation in Na and fine soil texture; high axis 3 scores corresponded to low [Na]FF, [Na]MS, NaMS(pool), base saturation, pH and sand, and high silt.

Varimax-rotated scores from NMDS ordination of relativized soil measurements: A Axis 1 versus Axis 2, B Axis 1 versus Axis 3, and C Axis 3 versus Axis 2. Overlays of seven overstory community types and three river segments are shown, as well as joint plots indicating correlations (Kendall’s |τ| > 0.3) between the ordination axes and soil measurements, as well as correlations with forest age, community composition, and channel proximity. Non-soil measures are shown in bold: age = stand age, elev = both elevationdisturbance and elevationgroundwater, Acer = both Acer macrophyllum and Acer circinatum overstory importance, Salix = Salix spp. overstory importance, and Thuja = Thuja plicata overstory importance. Among soil measures, cobble = depth to cobble, mass = forest floor mass, NaFF = [Na]FF, poolsFF = forest floor C, N, P, K, Ca, Mg, and Na pools, C:N = C:NMS (mass-based), C = [C]MS, N = [N]MS, K = [K]MS, Ca = [Ca]MS, Mg = [Mg]MS, Na = [Na]MS, Cp = mineral soil C pool, Np = mineral soil N pool, Pp = mineral soil P pool, Kp = mineral soil K pool, Cap = mineral soil Ca pool, Mgp = mineral soil Mg pool, Nap = mineral soil Na pool, CEC = mineral soil cation exchange capacity, base = mineral soil base saturation, pH = mineral soil pH, mineralization = mineral soil N mineralization rate, moisture = mineral soil moisture, BDT = mineral soil coarse bulk density, BDF = mineral soil fine bulk density, clay = percent composition by clay, silt = percent composition by silt, sand = percent composition by sand, 2 = percent composition by <2-mm particles, 16 = percent composition by 16–64-mm particles, and 64 = percent composition by > 64-mm particles.
NMDS axis 1 scores, and most of the individual soil measures associated with axis 1 scores, were positively correlated with stand age, elevationdisturbance, elevationgroundwater, and late-successional Acer macrophyllum and Acer circinatum importance (axis 1, τ = 0.58, 0.34, 0.44, 0.50, and 0.43, respectively), and negatively correlated with early-successional Salix spp. importance (axis 1, τ = −0.46) (Tables 2, 3; Figures 2, S2). These results suggested that fine sediments, forest floor mass, and mineral soil and forest floor nutrient concentrations and pools generally increased with time since major fluvial disturbance, and that NMDS axis 1 captured much of this variation in soil developmental stage.
NMDS axis 3 scores were not correlated with stand age or channel proximity, but were negatively correlated with Thuja plicata importance (τ = −0.31). T. plicata importance was positively correlated with [Na]MS, [Mg]FF, [Ca]MS, and [Mg]MS. NMDS axis 2 scores were not strongly correlated with stand age, channel proximity, or overstory species.
Overstory Community Comparisons
Overstory communities differed predictably in stand age and elevation above the channel, following a typical successional trajectory for Pacific Northwest riparian forests. Alnus–Salix communities were youngest, followed by Alnus and Alnus–Populus communities; Pseudotsuga–Alnus communities were intermediate, and Pseudotsuga–Abies, Acer, and Thuja–Acer communities were oldest (F 6,55 = 25.0, p < 0.0001; Figure S1). Alnus–Salix, Alnus, and Alnus–Populus communities occurred at lower elevations above the channel than Pseudotsuga–Abies and Thuja–Acer communities (disturbance, F 4,46 = 8.0, p < 0.0001; groundwater, F 4,49 = 9.0, p < 0.0001). Overstory communities did not differ significantly in distance from the channel.
Soil characteristics tended to differ between the younger, lower versus older, higher communities, corresponding to typical changes in riparian soils with time since major fluvial disturbance. Specifically, older communities tended to have higher depth to cobble, forest floor mass and nutrient pools, [P]FF, [Ca]FF, and mineral soil nutrient pools and concentrations, C:NMS (mass-based), CEC, moisture, silt, and less-than-2-mm particles than younger communities, as well as lower N:PFF (mass-based) and sand (Figures. 3, 4). Accordingly, NMDS axis 1 scores, which were correlated with all of those individual measures except [P]FF and N:PFF, were higher in Thuja–Acer, Acer, Pseudotsuga–Abies, Alnus–Populus, and Alnus communities than in Alnus–Salix communities, and higher in Pseudotsuga–Abies communities than in Pseudotsuga–Alnus and Alnus communities (Figure 2; F 6,53 = 13.5, p < 0.0001), further supporting interpretation of axis 1 as indicating soil development.

Depth to cobble and forest floor variables that differed significantly among seven overstory community types on the Elwha River: A depth to cobble (F 6,55 = 4.7, p = 0.0006), B forest floor mass (F 6,43 = 10.3, p < 0.0001), C forest floor C:N (F 6,37 = 2.1, p = 0.03), D forest floor N:P (F 6,49 = 5.9, p = 0.0001), E forest floor [P] (F 6,53 = 6.2, p < 0.0001), F forest floor [Ca] (F 6,54 = 4.6, p = 0.0008), G forest floor [Mg] (F 6,54 = 2.4, p = 0.04), H forest floor C pools (F 6,43 = 10.2, p < 0.0001), I forest floor N pools (F 6,42 = 7.4, p < 0.0001), J forest floor P pools (F 6,40 = 9.2, p < 0.0001), (k) forest floor K pools (F 6,54 = 4.6, p = 0.0007), L forest floor Ca pools (F 6,42 = 6.2, p = 0.0001), M forest floor Mg pools (F 6,41 = 6.0, p = 0.0002), and N forest floor Na pools (F 6,25 = 5.2, p = 0.001). Error bars are 1 SEM. Within each graph, different letters indicate significant differences among overstory community types (Tukey-adjusted L Smeans, p < 0.05).

Mineral soil variables that differed significantly among seven overstory community types on the Elwha River: A mineral soil C:N (F 6,44 = 9.1, p < 0.0001), B CEC (F 6,44 = 13.2, p < 0.0001), C pH (F 6,48 = 2.6, p = 0.03), D N mineralization (F 6,45 = 5.3, p = 0.0003), E moisture (F 6,43 = 3.3, p = 0.009), F mineral soil [C] (F 6,44 = 10.0, p < 0.0001), G mineral soil [N] (F 6,44 = 8.0, p < 0.0001), H mineral soil [P] (F 6,54 = 6.3, p < 0.0001), I mineral soil [K] (F 6,50 = 5.3, p = 0.0003), J mineral soil [Ca] (F 6,41 = 7.0, p < 0.0001), K mineral soil [Mg] (F 6,51 = 10.2, p < 0.0001), L mineral soil C pools (F 6,54 = 6.9, p < 0.0001), M mineral soil N pools (F 6,54 = 5.1, p = 0.0003), N mineral soil P pools (F 6,48 = 2.9, p = 0.02), O mineral soil K pools (F 6,46 = 4.4, p = 0.001), P mineral soil Ca pools (F 6,54 = 4.3, p = 0.001), Q mineral soil Mg pools (F 6,54 = 6.5, p < 0.0001), R percent composition by silt (F 6,55 = 3.4, p = 0.006), S percent composition by sand (F 6,55 = 2.6, p = 0.03), and T percent composition by < 2-mm particles (F 6,55 = 2.5, p = 0.04). Error bars are 1 SEM. Within each graph, different letters indicate significant differences among overstory community types (Tukey-adjusted LSmeans, p < 0.05).
A few differences in soils among overstory communities were less clearly related to successional stage. N mineralization was higher in Acer, Alnus, and Alnus–Populus communities than Alnus–Salix and Pseudotsuga–Abies communities. Further, [Mg]FF was higher in Thuja–Acer communities than Pseudotsuga–Abies communities.
Relative Importance of Stand Age and Channel Proximity
We evaluated the relative importance of stand age and different measures of channel proximity as predictors of soil characteristics by comparing AICc scores among models that included different combinations of stand age, elevationdisturbance, elevationgroundwater, distancedisturbance, and distancegroundwater. The best models included stand age alone for nine measures, either elevationdisturbance or elevationgroundwater alone for only three measures, stand age together with either elevation above or distance from the channel for 13 measures, and stand age, elevation, and distance for nine measures (Tables 4 and S1). For most soil variables, there was relatively strong support for other models besides the best model. However, for 16 soil measures, models that did not include stand age had very little support (w i ≤ 0.03, evidence ratio >10): depth to cobble, forest floor mass, CFF(pool), NFF(pool), PFF(pool), and KFF(pool), mineral soil C, N, and Mg concentrations and pools, Ca pools, CEC, base saturation, and, accordingly, NMDS axis 1 scores, which were correlated with all of these measures except base saturation (Table S1). The null model was the best model or close to the best model (evidence ratio <10) for 18 measures, including N cycling and forest floor nutrient concentrations, indicating poor support for stand age and channel proximity as predictors of those soil characteristics (Tables 4, S2).
Summing Akaike w i s across models containing elevationdisturbance, elevationgroundwater, distancedisturbance, or distancegroundwater suggested that proximity to the nearest disturbance channel was more often a better predictor than proximity to the nearest groundwater channel (Table S1). \(\sum {w_{i} } s\) were substantially greater for proximity to the nearest disturbance channel for forest floor mass and nutrient pools, for mineral soil [C]MS, [N]MS, [Ca]MS, [Mg]MS, MgMS(pool), CEC, pH, moisture, BDT, silt, sand, and less-than-2- and 16–64-mm particles, and, accordingly, for NMDS axis 1 scores (correlated with all of these measures but pH) and axis 3 scores (correlated with pH, silt, and sand). In contrast, \(\sum {w_{i} } s\) for proximity to the nearest groundwater channel were substantially greater only for depth to cobble, N:PFF (mass-based), and 2–16-mm particles.
Summing Akaike w i s across models also suggested that elevation above the channel was more often a better predictor than distance from the channel (Table S1). \(\sum {w_{i} } s\) were substantially greater for elevation than distance for depth to cobble, forest floor mass and nutrient pools (except KFF(pool)), N:PFF (mass-based), for mineral soil C:NMS (mass-based), [C]MS, [P]MS, KMS(pool), MgMS(pool), CEC, BDT, and less-than-2 and 16–64-mm particles, and accordingly, for NMDS axis 1 scores (which were correlated with all of these measures except [P]MS). \(\sum {w_{i} } s\) for distance were substantially greater only for [N]MS, [Mg]MS, pH, moisture, silt, sand, and, accordingly, NMDS axis 3 scores (which were correlated with pH, silt, and sand).
River Segment Comparisons
Soils on the upper segment tended to have characteristics associated with younger stand ages and overstory communities compared to soils on the middle segment, with intermediate levels on the lower segment (Table 5; Figure 5), supporting the hypothesis that the dams accelerated downstream riparian soil development. In accordance with earlier studies, plots on the upper segment contained significantly younger stands than the middle segment, with intermediate-aged stands on the lower segment. Soils on the upper segment had lower [Ca]MS, [Mg]MS, CMS(pool), CaMS(pool), MgMS(pool), CEC, moisture, clay, and, accordingly, NMDS axis 1 scores (which were correlated with all of these soil measures) than both the middle and lower segments, as well as lower [C]MS, [N]MS, NMS(pool) and silt, and higher sand, than the middle segment. An exception to the pattern of younger soil characteristics on the upper segment was that [P]MS and PMS(pool) were higher, and N:PMS (mass-based) was lower, on the upper segment than on the middle and lower segments. The differences in [C]MS, [N]MS, CMS(pool), NMS(pool), CEC, moisture, silt, sand, and NMDS axis 1 scores did not persist when stand age was included in the analyses (Table S3), suggesting that differences in stand age may explain these differences among segments. In contrast, differences persisted in [Ca]MS, [Mg]MS, CaMS(pool), MgMS(pool), clay, [P]MS, PMS(pool), and N:PMS (mass-based) when stand age was included in the analyses, suggesting that other factors contributed to those differences.

Mean A coarse and B fine soil particle composition of the three segments of the Elwha River. In B different letters indicate significant differences among segments within each particle size class (Tukey-adjusted LSmeans, p < 0.05): clay, F 2,13 = 11.4, p = 0.001; silt, F 2,55 = 3.6, p = 0.03; and sand, F 2,55 = 5.6, p = 0.006. Coarse soil particle composition in A did not differ significantly among segments.
Soils on the lower segment also had distinct characteristics, with higher [Ca]FF, [Mg]FF, [Na]FF, NaFF(pool), [Na]MS, base saturation, and, accordingly, NMDS axis 3 scores (which were correlated with [Na]FF, NaFF(pool), [Na]MS, and base saturation), than both the middle and upper segments, higher CaFF(pool), MgFF(pool), and sand than the middle segment, and higher [K]MS, NaMS(pool), and pH than the upper segment. With the exception of [K]MS and sand, these differences persisted when stand age was included in the analyses (Table S3).
We also tested for differences among segments in the slopes of relationships between stand age and soil characteristics, to identify possible dam effects on soil development rates. Segment × stand age interactions were significant for elevationdisturbance, C:NMS and C:NFF (mass-based), [N]FF, [P]FF, and [Mg]FF (Figure 6). Post hoc pairwise comparisons (Bonferroni-adjusted α = 0.017) indicated that for elevationdisturbance and C:NMS, effects of stand age differed between the upper and middle segments; both increased with stand age on the upper segment but did not change significantly with stand age on the middle segment. For C:NFF, [N]FF, [P]FF, and [Mg]FF, the upper segment differed from both the middle and lower segments. C:NFF decreased and [N]FF, [P]FF, and [Mg]FF increased with stand age on the lower and middle segments (although for the middle segment only the [P]FF trend was significant), whereas C:NFF increased and [N]FF, [P]FF, and [Mg]FF decreased (although not significantly) with stand age on the upper segment.

Stand age effects on plot and soil characteristics on the three segments of the Elwha River. Only variables with significant segment x stand age interactions are shown: A elevationdisturbance (F 2,40 = 5.8, p = 0.006), B C:Nmineral (F 2,51 = 6.9, p = 0.002), C C:Nforest-floor (F 2,33 = 9.9, p = 0.0004), D [N]forest-floor (F 2,23 = 9.4, p = 0.001), E [P]forest-floor (F 2,53 = 5.7, p = 0.006), and F [Mg]forest-floor (F 2,54 = 7.9, p = 0.001). Lines were estimated with linear regression. Significant lines were A lower, F 1,19 = 11.7, p = 0.003, slope = 0.4, intercept = −0.5, R 2 = 0.38; upper, F 1,15 = 9.3, p = 0.008, slope = 0.5, intercept = −0.4, R 2 = 0.38; B upper, F 1,16 = 26.9, p < 0.0001, slope = 11.6, intercept = −4.6, R 2 = 0.63; C lower, F 1,18 = 10.1, p = 0.005, slope = −10.7, intercept = 50.1, R 2 = 0.30; upper, F 1,15 = 9.7, p = 0.007, slope = 14.3, intercept = 9.5, R 2 = 0.37; D lower, F 1,18 = 11.7, p = 0.003, slope = 7.0, intercept = 5.5, R 2 = 0.63; E lower, F 1,19 = 16.2, p = 0.0007, slope = 0.5, intercept = 0.2, R 2 = 0.46; middle, F 1,19 = 8.3, p = 0.01, slope = 0.8, intercept = −0.4, R 2 = 0.30; and F lower, F 1,19 = 8.2, p = 0.01, slope = 1.2, intercept = 0.4, R 2 = 0.30.
Soil Stable Isotopes
Soil δ 13C and δ 15N were higher on gravel bars than mature Alnus rubra and conifer stands (Figure 7). In addition, δ 13C was higher on the upper segment than the middle segment [−25.7 ± 0.3 (S.E.M.) compared to −26.7 ± 0.2 (S.E.M.)‰; F 2,9 = 10.2, p = 0.004]. δ 15N did not differ among segments.

Mean soil δ 13C and δ 15N within three vegetation types along the Elwha River. Different letters indicate significant differences among vegetation types (Tukey-adjusted L Smeans, p < 0.05): δ 13C, F 2,37 = 45.9, p < 0.0001; δ 15N, F 2,36 = 12.6, p < 0.0001.
Discussion
Riparian Soils, Stand Age, and Channel Proximity
Riparian soils are spatially and temporally dynamic, with infrequent large floods scouring away floodplain communities and soils, transporting and depositing sediments and creating new geomorphic surfaces, and more frequent, smaller floods depositing fine sediments that contribute organic matter and nutrients (Naiman and others 2005; Naiman and others 2010). Soil characteristics across toposequences along the Elwha River were strongly correlated with forest stand age, which can be considered a proxy for time since the most recent stand-replacing flood. Older, later-successional riparian forests contained soils with finer texture, lower bulk density, greater depth to cobble, and higher forest floor mass, forest floor and mineral soil nutrient concentrations and pools, and mineral soil C:N, CEC, and moisture.
Increased time since fluvial disturbance often leads to finer soil texture and higher nutrient content in riparian soils (Johnson and others 1976; Schwendenmann 2000; Bagstad and others 2006; Bechtold and Naiman 2006; Morris and others 2010; Cierjacks and others 2011). In Pacific Northwest riparian forests, soil C and N accumulate rapidly for approximately 100 years after major fluvial disturbance (Entry and Emmingham 1995; Latterell and others 2006; Bechtold and Naiman 2009; Naiman and others 2010), but C and N accumulation often slows after 100–200 years (Walker 1989; Chapin and others 1994; Bechtold and Naiman 2009). The continued accumulation of soil C and N with time in our sampling likely reflects the relatively young age of sampled forests, with only four stands more than 110 years old, and suggests that dam effects on soil C and N accumulation occurred during a dynamic period of soil development in these forests.
Several different measurements of riparian geomorphic surfaces, including stand age, elevation above and distance from the primary channel, and elevation above and distance from side channels, can serve as proxy measurements for time since fluvial disturbance, sedimentation processes, and moisture regimes (for example, van Coller and others 2000; Cierjacks and others 2011; Bedison and others 2013). At our sites, model selection suggested that stand age was a more consistently useful predictor than channel proximity for a diverse array of soil characteristics. Stand age may be a better proxy for time since major disturbance than channel proximity at our sites because fluvial disturbance occurs at different elevations and distances from the channel depending on channel and floodplain geometry and river stage–discharge relationships. The greater importance of stand age than channel proximity also may reflect high forest productivity and the potential for forests in this region to operate together with fluvial inputs to shape soil development (Bechtold and Naiman 2009). However, the best models included both stand age and channel proximity for 22 physical and chemical soil measures, highlighting how interactions between fluvial disturbance, sediment deposition, and biotic retention regulate soil development in this dynamic ecosystem.
Proximity to primary, “disturbance” channels was more informative than proximity to secondary, “groundwater” channels for most variables, suggesting that effects of channel proximity on fluvial disturbance or sediment deposition (independent of stand age) were more important than effects on water availability. Elevation above the channel also tended to be more informative than distance from the channel, probably because elevation determines inundation during floods more consistently than distance.
Soil C and N Sources During Riparian Succession
Changes in soil δ 15N and δ 13C with succession can reflect changes in the sources and/or processing of N and C. The rapid declines we observed in both soil δ 15N and δ 13C during the transition from gravel bars to mature Alnus rubra stands, with slower changes thereafter, suggest that organic matter sources changed more than N and C processing with succession along the Elwha River. Specifically, while microbial processing of soil N and C typically discriminates against heavy isotopes and thus increases soil δ 15N and δ 13C over time during succession (Handley and Raven 1992; Šantrůčková and others 2000), we observed the opposite pattern. Instead, the declining soil δ 15N and δ 13C during succession suggest a more important role for 15N-depleted inputs from symbiotic N-fixation and 13C-depleted inputs from photosynthesis (Farquhar and others 1989; Handley and Raven 1992).
Freshly created gravel bars had low mineral soil C:N (<10) and δ 15N [1.7 ± 0.1 (S.E.M.)‰; Figure 7b], as expected from suspended river sediments of eroded mineral soil (~1.4‰, Hatten and others 2012). Soil δ 15N then declined significantly from gravel bars to mature Alnus rubra and conifer stands [0.7 ± 0.1 and 0.6 ± 0.4 (S.E.M.)‰, respectively], approaching atmospheric N2 (0‰), consistent with symbiotic N-fixation by A. rubra, and sustained retention of fixed N during succession to conifer stands (Scott and others 2008; Bechtold and Naiman 2009; Perakis and others 2015). Soil δ 15N along the Elwha River was somewhat lower than in other young Pacific Northwest riparian forests containing A. rubra (for example, 2.2, 2.2, 3.0‰; Bartz and Naiman 2005; Scott and others 2008; D’Amore and others 2011) and considerably lower than in older Pacific Northwest terraces and upland forests (for example, 3.7, 4.3, 6.1‰; Trudell and others 2004; D’Amore and others 2011; Perakis and others 2011; Perakis and others 2015), perhaps indicating higher A. rubra N-fixation, lower denitrification, or lower 15N-enriched marine inputs along the Elwha River.
Similarly, elevated soil δ 13C on gravel bars [−25.2 ± 0.2 (S.E.M.)‰] was consistent with deposition of C derived from suspended mineral sediments (~ −26‰; Hatten and others 2012), older organic matter (−25 to −26‰; Sanderman and others 2009), and carbonates (−4 to −11‰; Stevenson and others 2005). Soil δ 13C declined in mature A. rubra and conifer stands [−27.1 ± 0.2 and −26.5 ± 0.1 (S.E.M.)‰, respectively], perhaps indicating a shift towards greater contributions of 13C-depleted riparian plant organic matter produced in situ [−29.8 ± 0.2 (S.E.M.)‰; K. Jenkins, unpublished data]. Higher δ 13C on gravel bars was also likely due to higher riparian foliar δ 13C in these habitats than in A. rubra and conifer stands [−28.7 ± 0.3 (S.E.M.) compared to −30.6 ± 0.2 (S.E.M.)‰; K. Jenkins, unpublished data). Plants on gravel bars may have discriminated less against 13C because of higher light availability and photosynthetic capacity (Yakir and Israeli 1995; Duursma and Marshall 2006), or, less likely, because of higher water stress and lower stomatal conductance (Farquhar and others 1989). Soil δ 13C in A. rubra and conifer stands was similar to soil δ 13C in upland old-growth conifer forests in the area [−26.0 ± 0.2 and −26.8 ± 0.2 (S.E.M.)‰; Trudell and others 2004], suggesting more balanced 13C-depleted gains and losses from plant inputs and microbial respiration during later succession.
Taken together, these results support the idea that N and C sources can change rapidly with development of gravel bars and succession to mature A. rubra stands, with slower changes during subsequent succession to conifers.
Riparian Soils and Overstory Community Composition
Nitrogen cycling along the Elwha River was more closely related to overstory community composition than to time since major fluvial disturbance. Although higher river-floodplain hydrological connectivity and sedimentation often increase riparian N mineralization (Noe and others 2013), we found that stand age and channel proximity were not correlated with potential N mineralization along the Elwha River. Instead, mineral soil N cycling may have been influenced more by variation in N-fixation and litterfall content. Rapid N mineralization in Alnus and Alnus–Populus communities was likely due to A. rubra N-fixation (Luken and Fonda 1983; Perakis and others 2012). In contrast, relatively low [N]MS in young Alnus–Salix communities, and relatively high C:NMS in Pseudotsuga–Alnus communities, may explain the slower N mineralization in these two Alnus community types. High Acer macrophyllum and Acer circinatum foliage and litterfall N content, and low litterfall C:N and lignin:N, compared to Pseudotsuga menziesii, Tsuga heterophylla, and Thuja plicata (Ogden and Schmidt 1997; Tashe and Schmidt 2003; Klopatek 2008; Cross and Perakis 2011; Hamdan and Schmidt 2012; but see Turk and others 2008; van Huysen and others 2013), all of which contribute to higher litter quality and decomposability, may explain the faster N mineralization in Acer communities than Pseudotsuga–Abies and Thuja–Acer communities.
Several soil nutrients were also strongly associated with particular overstory species. Correlations between A. macrophyllum and A. circinatum importance and mineral soil nutrient concentrations and pools support the hypothesis that A. macrophyllum and A. circinatum increase soil [N], [K], [Ca], and [Mg] relative to P. menziesii, T. heterophylla, and T. plicata (Ogden and Schmidt 1997; Tashe and Schmidt 2003; Chandler and others 2008; Turk and others 2008; Cross and Perakis 2011; Hamdan and Schmidt 2012). High [P]FF in Acer communities likely reflect biological cycling of inorganic P from mineral soil (Tashe and Schmidt 2003) to the forest floor via high P uptake into foliage and litterfall in the Acer spp. (Cross and Perakis 2011). Similarly, high forest floor and mineral soil [Ca] and [Mg] in Thuja–Acer communities, and correlations with high T. plicata importance, support the hypothesis that T. plicata promotes Ca and Mg accumulation in biomass and soils relative to P. menziesii and T. heterophylla (Turner and Franz 1985; Prescott and others 2000; Cross and Perakis 2011; Hawkins and Robbins 2014).
Riparian Soils Upstream, Between, and Downstream of the Dams
Long-term reduction in sediment supply below dams has the potential to influence riparian soils in contrasting ways. Lower sediment supply may slow soil development by reducing deposition of fine sediments, organic matter, and nutrients (Pinay and others 2002). At the same time, it may promote soil development by increasing retention of fine sediments, organic matter, and nutrients on riparian surfaces downstream of dams. This increased retention can occur because low sediment supply below dams can induce armoring, incision, and stabilization of the active channel (Williams and Wolman 1984), thus reducing channel mobility and floodplain avulsion, and hence erosion of developed riparian soils.
Differences in channel bed grain size and armoring among segments suggest that sediment supply below the Elwha River dams was most limited on the middle segment and less limited on the lower segment, where there was some local sediment recruitment (Pohl 2004). On the upper segment, abundant sediment and large woody debris, in part from a 1967 natural landslide dam failure (Acker and others 2008), have contributed to considerable channel mobility and floodplain turnover, whereas on the middle segment and upstream portions of the lower segment, low sediment and woody debris supplies have resulted in channel stabilization and low floodplain turnover (Pohl 2004; Kloehn and others 2008; Draut and others 2011). We found that [C]MS, [N]MS, CEC, silt, clay, and NMDS axis 1 scores (which were correlated with each of these variables) were all highest on the middle segment, lowest on the upper segment, and intermediate on the lower segment, whereas sand (that is, percent of <2-mm sediment made up of sand) was lowest on the middle segment and highest on the upper segment. These results suggest that reduced sediment supply may have promoted riparian soil development below the dams, by reducing channel mobility and thus increasing fine sediment and nutrient retention. These effects of lower channel mobility may have overwhelmed the direct effects of reduced sediment supply on fine sediment deposition below the dams.
Given the importance of stand age in predicting riparian soil characteristics, the large differences in forest dynamics among segments, due at least in part to the dams, were likely to be associated with differences in soils. As a result of channel stabilization below the dams, the middle segment, and to a lesser extent the lower segment, has experienced less riparian forest disturbance and turnover than the upper segment over the last century (Kloehn and others 2008; Shafroth and others 2017), resulting in fewer opportunities for erosional losses of fine sediment and organic matter. A broad survey of riparian forests along the Elwha River indicated that stand ages on the middle segment were considerably older than on the upper and lower segments [86 ± 5 compared to 50 ± 7 and 56 ± 6 (S.E.M.) years, respectively] (Shafroth and others 2017), similar to the stand ages sampled in the present study (Table 5). Further, aerial photograph interpretation suggested that the mean age of riparian forests remained stable on the upper segment due to natural floodplain dynamics, but increased on the middle and lower segments between 1940 and 2000 due to reduced fluvial disturbance (Kloehn and others 2008). The finer soil texture and higher soil [C]MS, [N]MS, and CEC on the middle segment than on the upper segment may have been a simple result of higher abundance of older geomorphic surfaces due to lower channel mobility on this segment. Accounting for differences in stand age among segments eliminated these differences in soil properties, except for clay.
We saw little evidence that reduced sediment supply decreased fine sediments, organic matter, or nutrients on floodplains and terraces of the middle and lower segments, in contrast to the increased bed grain size of the active mainstem channel below the dams (Pohl 2004; Draut and others 2011) and in contrast to reports of coarser sediments and limited nutrients below dams in the arid and semiarid western U.S. (Graf 2000; Phillips and others 2004; Beauchamp and others 2007; Rood and others 2011). Almost all soil characteristics correlated with fine sediments were either higher below the dams or did not differ among segments. Further, there were few significant interactions between segment and stand age effects that might have suggested slower rates of soil development on older surfaces of the middle and lower segments due to reduced fine sediment deposition. Given that the Elwha River dams captured a large proportion of the river’s fine sediment (Randle and others 2015), much of the silt and clay on older surfaces of the middle segment may have been deposited largely before Glines Canyon Dam was installed; 55% of plots on the middle segment existed prior to the dam. Subsequent soil development on these older surfaces may have been driven mainly by autochthonous inputs from the riparian plant community (Bechtold and Naiman 2009; Cabezas and Comin 2010; Batz and others 2015).
However, there was some evidence to suggest that lower sediment supply did reduce sediment and nutrient inputs to riparian soils below the dams. First, mineral soil P concentrations and pools were highest on the upper segment, which could not be explained by differences in stand age. Instead, this pattern may be attributable to the greater abundance of P-rich shale bedrock in the upper watershed (Duda and others 2008; Moran and others 2013; Porder and Ramachandran 2013), combined with reduced fluvial P inputs to the middle and lower segments. P is less soluble than most nutrients and thus more tightly bound to the sediment and organic matter that accumulates behind dams (Kennedy and Walker 1990). Previous research has reported reduced stream water P concentrations below the Elwha River dams (Duda and others 2011a), as well as in other dammed rivers (Kelly 2001; Paul 2003; Bosch and others 2009). Second, the lack of significantly higher depth to cobble, forest floor mass, and less-than-2-mm particles on the middle segment despite its older stand ages could reflect counteracting effects of low sediment inputs on the otherwise greater soil development on the middle segment. In particular, depth to cobble was highest on the upper segment (although not significantly), suggesting that soils on the middle segment may have been relatively shallow for their age. Third, elevationdisturbance increased more slowly with stand age on the middle and lower segments than on the upper segment, suggesting slower sediment accumulation below the dams. Finally, on younger surfaces, [N]FF and [P]FF were lower, and C:NMS and C:NFF were higher, on the middle and lower segments than on the upper segment, which may have been due to reduced fluvial N and P inputs to young surfaces below the dams.
Soil δ 15N did not differ among segments, suggesting that the dams had little discernable effect on soil N sources or 15N-depleting soil processes. In particular, δ 15N was not higher on the lower segment, in contrast to expectations that nearly a century of restricted N transport in anadromous salmon (δ 15N = 10.4–12.5‰; Mathisen and others 1988) would lead to elevated soil δ 15N below, and not above, the dams. Several explanations may account for the lack of elevated soil δ 15N below the dams, including diminished salmon populations since dam construction (Pess and others 2008) and/or dominance of ecosystem soil N budgets by A. rubra N-fixation (Helfield and Naiman 2002; Scott and others 2008).
Other factors besides the dams also may have contributed to differences in riparian soil characteristics among river segments. Effects of dams on downstream riverine and riparian ecosystems can be difficult to distinguish from natural longitudinal variation. Upstream and downstream areas often differ naturally in sediment parent material, climate, valley width and geometry, stream gradient, and more, especially above and below the naturally narrow points where dams are often constructed. Further, river sediment and nutrient regimes commonly change longitudinally, as fine sediments and nutrients are transported from upstream and accumulate downstream (Newbold and others 1982; Kuglerova and others 2015). The river segments examined in this study differed in gradient, declining from a mean of 0.0149 m m−1 on the upper segment to 0.0065 and 0.0030 m m−1 on the middle and lower segments, respectively (Kloehn and others 2008). As rivers decline in gradient from headwaters to mouth, flow velocity also declines, and suspended fine sediments are deposited on the river bed, channel margins, and floodplain (Knighton 1984). Lower gradients on the middle and lower segments may have increased silt and clay deposition before dam emplacement, perhaps especially clay, as differences in clay among segments could not be explained by differences in stand age. However, the differences in gradient, along with downstream sediment transport and sorting, would not be expected to result in the higher observed silt and clay on the middle segment than on the lower segment. Instead, lower channel mobility on the middle segment likely contributed at least in part to its finer sediment texture.
Differences among segments in mineral soil and forest floor Ca, Mg, and Na, and hence base saturation, pH, and NMDS axis 3 scores (which were correlated with Na, pH, and base saturation) may have resulted mainly from natural longitudinal trends. These soil characteristics were lowest on the upper segment and highest on the lower segment, and their differences could not be explained by differences in stand age among segments. Downstream movement and accumulation of clay may have increased Ca, Mg, and Na retention on the lower and middle segments, even though the low C:NMS (13–15 g g−1) and high relative nitrification (84–94%) measured across segments should promote intense base cation leaching from soils (Van Miegroet and Cole 1984; Perakis and others 2006; Perakis and others 2013). The maintenance of relatively high base saturation in all segments (62–85%) suggests that local weathering and/or riverine transport of base cations was sufficient to maintain high fertility in riparian soils across all segments.
Implications for Riparian Plant Communities
Differences in riparian soils among river segments may have contributed to differences in riparian plant community composition or productivity. Higher [N]MS and finer soil texture on the middle segment had the potential to increase forest productivity (Edmonds and Chappell 1994; Kayahara and others 1995; Graff and others 1999) and relative success of species better adapted to fine-textured or N-rich soils. Differences in soil δ 13C among segments may indicate higher productivity on the middle segment. The lower soil δ 13C on the middle segment may have resulted from a higher proportional contribution of 13C-depleted plant organic matter to the total C pool on the middle segment, perhaps due to higher plant production, leading to a lower proportional contribution of relatively 13C-enriched river sediments to the total C pool on the middle segment than on the upper segment. Foliar δ 13C was not lower on the middle segment for three woody riparian species (K. Jenkins, unpublished data), suggesting that low soil δ 13C on the middle segment was not due to more intense plant 13C discrimination.
On the other hand, reduced P concentrations and pools on the middle and lower segments may have constrained riparian plant responses to higher N (Boggs and Weaver 1994). P sometimes limits growth of P. menziesii, T. heterophylla, and perhaps other species in Pacific Northwest forests (Radwan and others 1991; Blevins and others 2006; Mainwaring and others 2014). The Elwha River watershed is relatively poor in P, as a result of low P in soil parent material and low human P inputs to the watershed (Duda and others 2011a; Cavaliere and Homann 2012). P capture by the dams intensified this P-limitation in riparian soils, increasing available N:Pmineral by greater than 60% on the middle and lower segments. More intense P-limitation may have reduced riparian plant productivity, reduced A. rubra N-fixation (Uliassi and Ruess 2002), or favored species with lower P requirements, such as T. plicata (Cade-Menun and Berch 1997; Blevins and others 2006,;Cross and Perakis 2011).
Dam Removal and Riparian Soils
Removal of the Elwha River dams will likely have ongoing effects on riparian soils, with further implications for riparian communities. In the five years since dam removal began, more than 1.2 million tons of reservoir sediment have accumulated in the main channel, side channels, and channel margins downstream, forming new bars, prompting channel avulsion, and creating a braided channel morphology (Draut and Ritchie 2015; East and others 2015; Warrick and others 2015). Winter floods in 2014 and 2015 (300–450 m3 s−1 instantaneous peak flow; USGS gage 12045500, Elwha River at McDonald Bridge) deposited reservoir sediments on floodplain surfaces (Shafroth, personal observation). Future monitoring of the spatial extent, depth, texture and nutrient content of these deposits, and their effects on channel mobility, will reveal long-term effects of dam removal on Elwha River riparian soils.
References
Acker SA, Beechie TJ, Shafroth PB. 2008. Effects of a natural dam-break flood on geomorphology and vegetation on the Elwha River, Washington, USA. Northwest Sci 82:210–23.
Adair EC, Binkley D, Andersen DC. 2004. Patterns of nitrogen accumulation and cycling in riparian floodplain ecosystems along the Green and Yampa rivers. Oecologia 139:108–16.
Andersen DC, Nelson SM, Binkley D. 2003. Flood flows, leaf breakdown, and plant-available nitrogen on a dryland river floodplain. Wetlands 23:180–9.
Appling AP, Bernhardt ES, Stanford JA. 2014. Floodplain biogeochemical mosaics: a multidimensional view of alluvial soils. J. Geophys Res-Biogeosci 119:1538–53.
Bagstad KJ, Lite SJ, Stromberg JC. 2006. Vegetation, soils, and hydrogeomorphology of riparian patch types of a dryland river. West N Am Nat 66:23–44.
Bartz KK, Naiman RJ. 2005. Effects of salmon-borne nutrients on riparian soils and vegetation in southwest Alaska. Ecosystems 8:529–45.
Batz N, Verrecchia EP, Lane SN. 2015. Organic matter processing and soil evolution in a braided river system. Catena 126:86–97.
Beauchamp VB, Stromberg JC, Stutz JC. 2007. Flow regulation has minimal influence on mycorrhizal fungi of a semi-arid floodplain ecosystem despite changes in hydrology, soils, and vegetation. J Arid Environ 68:188–205.
Bechtold JS, Naiman RJ. 2006. Soil texture and nitrogen mineralization potential across a riparian toposequence in a semi-arid savanna. Soil Biol Biochem 38:1325–33.
Bechtold JS, Naiman RJ. 2009. A quantitative model of soil organic matter accumulation during floodplain primary succession. Ecosystems 12:1352–68.
Bedison JE, Scatena FN, Mead JV. 2013. Influences on the spatial pattern of soil carbon and nitrogen in forested and non-forested riparian zones in the Atlantic Coastal Plain of the Delaware River Basin. For Ecol Manag 302:200–9.
Blevins LL, Prescott CE, Van Niejenhuis A. 2006. The roles of nitrogen and phosphorus in increasing productivity of western hemlock and western redcedar plantations on northern Vancouver Island. For Ecol Manag 234:116–22.
Boggs K, Weaver T. 1994. Changes in vegetation and nutrient pools during riparian succession. Wetlands 14:98–109.
Bosch NS, Johengen TH, Allan JD, Kling GW. 2009. Nutrient fluxes across reaches and impoundments in two southeastern Michigan watersheds. Lake Reserv Manag 25:389–400.
Cabezas A, Comin FA. 2010. Carbon and nitrogen accretion in the topsoil of the Middle Ebro River floodplains (NE Spain): implications for their ecological restoration. Ecol Eng 36:640–52.
Cade-Menun BJ, Berch SM. 1997. Response of mycorrhizal western red cedar to organic phosphorus sources and benomyl. Can J Bot 75:1226–35.
Cavaliere E, Homann P. 2012. Elwha river sediments: phosphorus characterization and dynamics under diverse environmental conditions. Northwest Sci 86:95–107.
Chandler JR, Schmidt MG, Dragicevic S. 2008. Spatial patterns of forest floor properties and litterfall amounts associated with bigleaf maple in conifer forest of southwestern British Columbia. Can J Soil Sci 88:295–313.
Chapin FS, Walker LR, Fastie CL, Sharman LC. 1994. Mechanisms of primary succession following deglaciation at Glacier Bay, Alaska. Ecol Monogr 64:149–75.
Cierjacks A, Kleinschmit B, Kowarik I, Graf M, Lang F. 2011. Organic matter distribution in floodplains can be predicted using spatial and vegetation structure data. River Res Appl 27:1048–57.
Cross A, Perakis SS. 2011. Tree species and soil nutrient profiles in old-growth forests of the Oregon Coast Range. Can J For Res 41:195–210.
D’Amore DV, Bonzey NS, Berkowitz J, Ruegg J, Bridgham S. 2011. Holocene soil-geomorphic surfaces influence the role of salmon-derived nutrients in the coastal temperate rainforest of southeast Alaska. Geomorphology 126:377–86.
Daniels JM. 2003. Floodplain aggradation and pedogenesis in a semiarid environment. Geomorphology 56:225–42.
Daubenmire RF. 1959. A canopy-coverage method of vegetational analysis. Northwest Sci 33:43–64.
Drake DC, Smith JV, Naiman RJ. 2005. Salmon decay and nutrient contributions to riparian forest soils. Northwest Sci 79:61–71.
Draut AE, Logan JB, Mastin MC. 2011. Channel evolution on the dammed Elwha River, Washington, USA. Geomorphology 127:71–87.
Draut AE, Ritchie AC. 2015. Sedimentology of new fluvial deposits on the Elwha River, Washington, USA, formed during large-scale dam removal. River Res Appl 31:42–61.
Duda JJ, Coe HJ, Morley SA, Kloehn KK. 2011a. Establishing spatial trends in water chemistry and stable isotopes (δ 15N and δ 13C) in the Elwha River prior to dam removal and salmon recolonization. River Res Appl 27:1169–81.
Duda JJ, Freilich JE, Schreiner EG. 2008. Baseline studies in the Elwha River ecosystem prior to dam removal: introduction to the special issue. Northwest Sci 82:1–12.
Duda JJ, Warrick JA, Magirl CS. 2011b. Coastal and lower Elwha River, Washington, prior to dam removal - history, status, and defining characteristics. Duda JJ, Warrick JA, Magirl CS, editors. Coastal habitats of the Elwha River: biological and physical patterns and processes prior to dam removal. U.S. Geological Survey scientific investigations report 2011-5120. Reston (VA): U.S. Geological Survey. pp 1–26.
Duursma RA, Marshall JD. 2006. Vertical canopy gradients in δ 13C correspond with leaf nitrogen content in a mixed-species conifer forest. Trees-Struct Funct 20:496–506.
East AE, Pess GR, Bountry JA, Magirl CS, Ritchie AC, Logan JB, Randle TJ, Mastin MC, Minear JT, Duda JJ, Liermann MC, McHenry ML, Beechie TJ, Shafroth PB. 2015. Large-scale dam removal on the Elwha River, Washington, USA: river channel and floodplain geomorphic change. Geomorphology 228:765–86.
Edmonds RL, Chappell HN. 1994. Relationships between soil organic-matter and forest productivity in western Oregon and Washington. Can J For Res 24:1101–6.
Entry JA, Emmingham WH. 1995. Influence of forest age on nutrient availability and storage in coniferous soils of the Oregon Coast Range. Can J For Res 25:114–20.
Farquhar GD, Ehleringer JR, Hubick KT. 1989. Carbon isotope discrimination and photosynthesis. Ann Rev Plant Physiol Plant Mol Biol 40:503–37.
Friedman JM, Osterkamp WR, Scott ML, Auble GT. 1998. Downstream effects of dams on channel geometry and bottomland vegetation: regional patterns in the Great Plains. Wetlands 18:619–33.
Graf WL. 2000. Locational probability for a dammed, urbanizing stream: Salt River, Arizona, USA. Environ Manag 25:321–35.
Graf WL. 2006. Downstream hydrologic and geomorphic effects of large dams on American rivers. Geomorphology 79:336–60.
Graff JE, Hermann RK, Zaerr JB. 1999. Dry matter and nitrogen allocation in western redcedar, western hemlock, and Douglas fir seedlings grown in low- and high-N soils. Ann For Sci 56:529–38.
Grams PE, Schmidt JC. 2005. Equilibrium or indeterminate? Where sediment budgets fail: sediment mass balance and adjustment of channel form, Green River downstream from Flaming Gorge Dam, Utah and Colorado. Geomorphology 71:156–81.
Griffin ER, Perignon MC, Friedman JM, Tucker GE. 2014. Effects of woody vegetation on overbank sand transport during a large flood, Rio Puerco, New Mexico. Geomorphology 207:30–50.
Gurnell A, Petts G. 2006. Trees as riparian engineers: the Tagliamento River, Italy. Earth Surf Proc Land 31:1558–74.
Hamdan K, Schmidt M. 2012. The influence of bigleaf maple on chemical properties of throughfall, stemflow, and forest floor in coniferous forest in the Pacific Northwest. Can J For Res 42:868–78.
Handley LL, Raven JA. 1992. The use of natural abundance of nitrogen isotopes in plant physiology and ecology. Plant, Cell Environ 15:965–85.
Harms TK, Grimm NB. 2008. Hot spots and hot moments of carbon and nitrogen dynamics in a semiarid riparian zone. J Geophys Res-Biogeosci. doi:10.1029/2007JG000588.
Hatten JA, Goni MA, Wheatcroft RA. 2012. Chemical characteristics of particulate organic matter from a small, mountainous river system in the Oregon Coast Range, USA. Biogeochemistry 107:43–66.
Hawkins BJ, Robbins S. 2014. Contrasts in growth and nitrogen nutrition of species in the Cupressaceae and Pinaceae in response to calcium. Plant Soil 380:315–25.
Helfield JM, Naiman RJ. 2001. Effects of salmon-derived nitrogen on riparian forest growth and implications for stream productivity. Ecology 82:2403–9.
Helfield JM, Naiman RJ. 2002. Salmon and alder as nitrogen sources to riparian forests in a boreal Alaskan watershed. Oecologia 133:573–82.
Hill AR. 2011. Buried organic-rich horizons: their role as nitrogen sources in stream riparian zones. Biogeochemistry 104:347–63.
Hupp CR, Schenk ER, Kroes DE, Willard DA, Townsend PA, Peet RK. 2015. Patterns of floodplain sediment deposition along the regulated lower Roanoke River, North Carolina: annual, decadal, centennial scales. Geomorphology 228:666–80.
Johnson WC, Burgess RL, Keammerer WR. 1976. Forest overstory vegetation and environment on the Missouri River floodplain in North Dakota. Ecol Monogr 46:59–84.
Kayahara GJ, Carter RE, Klinka K. 1995. Site index of western hemlock (Tsuga heterophylla) in relation to soil nutrient and foliar chemical measures. For Ecol Manag 74:161–9.
Kelly VJ. 2001. Influence of reservoirs on solute transport: a regional-scale approach. Hydrol Process 15:1227–49.
Kennedy RH, Walker WW. 1990. Reservoir nutrient dynamics. In: Thornton KW, Kimmel BL, Payne FE, Eds. Reservoir limnology: ecological perspectives. New York: Wiley. p 109–31.
Kloehn KK, Beechie TJ, Morley SA, Coe HJ, Duda JJ. 2008. Influence of dams on river-floodplain dynamics in the Elwha River, Washington. Northwest Sci 82:224–35.
Klopatek JM. 2008. Litter decomposition contrasts in second- and old-growth Douglas-fir forests of the Pacific Northwest, USA. Plant Ecol 196:123–33.
Knighton D. 1984. Fluvial forms and processes. London: Edward Arnold. p 218p.
Koyama L, Kielland K. 2011. Plant physiological responses to hydrologically mediated changes in nitrogen supply on a boreal forest floodplain: a mechanism explaining the discrepancy in nitrogen demand and supply. Plant Soil 342:129–39.
Kuglerova L, Jansson R, Sponseller RA, Laudon H, Malm-Renofalt B. 2015. Local and regional processes determine plant species richness in a river-network metacommunity. Ecology 96:381–91.
Kunz MJ, Anselmetti FS, Wuest A, Wehrli B, Vollenweider A, Thuring S, Senn DB. 2011a. Sediment accumulation and carbon, nitrogen, and phosphorus deposition in the large tropical reservoir Lake Kariba (Zambia/Zimbabwe). J Geophys Res-Biogeosci. doi:10.1029/2010JG001538.
Kunz MJ, Wuest A, Wehrli B, Landert J, Senn DB. 2011b. Impact of a large tropical reservoir on riverine transport of sediment, carbon, and nutrients to downstream wetlands. Water Resour Res. doi:10.1029/2011WR010996.
Latterell JJ, Bechtold JS, O’Keefe TC, Van Pelt R, Naiman RJ. 2006. Dynamic patch mosaics and channel movement in an unconfined river valley of the Olympic Mountains. Freshw Biol 51:523–44.
Le Bouteiller C, Venditti JG. 2014. Vegetation-driven morphodynamic adjustments of a sand bed. Geophys Res Lett 41:3876–83.
Luken JO, Fonda RW. 1983. Nitrogen accumulation in a chronosequence of red alder communities along the Hoh River, Olympic National Park, Washington. Can J For Res 13:1228–37.
Mainwaring DB, Maguire DA, Perakis SS. 2014. Three-year growth response of young Douglas-fir to nitrogen, calcium, phosphorus, and blended fertilizers in Oregon and Washington. For Ecol Manag 327:178–88.
Mallik AU, Richardson JS. 2009. Riparian vegetation change in upstream and downstream reaches of three temperate rivers dammed for hydroelectric generation in British Columbia, Canada. Ecol Eng 35:810–19.
Mathisen OA, Parker PL, Goering JJ, Kline TC, Poe PH, Scalan RS. 1988. Recycling of marine elements transported into freshwater by anadromous salmon. Verh der Intern Ver Limnol 23:2249–58.
McClain ME, Boyer EW, Dent CL, Gergel SE, Grimm NB, Groffman PM, Hart SC, Harvey JW, Johnston CA, Mayorga E, McDowell WH, Pinay G. 2003. Biogeochemical hot spots and hot moments at the interface of terrestrial and aquatic ecosystems. Ecosystems 6:301–12.
Meyer GA. 2001. Recent large-magnitude floods and their impact on valley-floor environments of northeastern Yellowstone. Geomorphology 40:271–90.
Moran PW, Cox SE, Embrey SS, Huffman RL, Olsen TD, Fradkin SC. 2013. Sources and sinks of nitrogen and phosphorus in a deep oligotrophic lake, Lake Cresent, Olympic National Park, Washington: U.S. Geological Survey scientific investigations report 2012-5107. Reston (VA): U.S. Geological Survey. 56p.
Morris MR, Brouwer BO, Caves JK, Harner MJ, Stanford JA. 2010. Successional changes in soil and hyporheic nitrogen fertility on an alluvial flood plain: implications for riparian vegetation. Aquat Sci 72:519–32.
Morris MR, Stanford JA. 2011. Floodplain succession and soil nitrogen accumulation on a salmon river in southwestern Kamchatka. Ecol Monogr 81:43–61.
Naiman RJ, Bechtold JS, Beechie TJ, Latterell JJ, Van Pelt R. 2010. A process-based view of floodplain forest patterns in coastal river valleys of the Pacific Northwest. Ecosystems 13:1–31.
Naiman RJ, Bechtold JS, Drake DC, Latterell JJ, O’Keefe TC, Balian EV. 2005. Origins, patterns, and importance of heterogeneity in riparian systems. In: Lovett GM, Turner MG, Jones CG, Weathers KC, Eds. Ecosystem function in heterogeneous landscapes. New York: Springer. p 279–309.
Newbold JD, O’Neill RV, Elwood JW, Van Winkle W. 1982. Nutrient spiralling in streams: implications for nutrient limitation and invertebrate activity. Am Nat 120:628–52.
Nilsson C, Berggren K. 2000. Alterations of riparian ecosystems caused by river regulation. Bioscience 50:783–92.
Noe GB, Hupp CR, Rybicki NB. 2013. Hydrogeomorphology influences soil nitrogen and phosphorus mineralization in floodplain wetlands. Ecosystems 16:75–94.
Ogden AE, Schmidt MG. 1997. Litterfall and soil characteristics in canopy gaps occupied by vine maple in a coastal western hemlock forest. Can J Soil Sci 77:703–11.
Paul L. 2003. Nutrient elimination in pre-dams: results of long term studies. Hydrobiologia 504:289–95.
Perakis S, Maguire D, Bullen T, Cromack K, Waring R, Boyle J. 2006. Coupled nitrogen and calcium cycles in forests of the Oregon Coast Range. Ecosystems 9:63–74.
Perakis SS, Matkins JJ, Hibbs DE. 2012. N2-fixing red alder indirectly accelerates ecosystem nitrogen cycling. Ecosystems 15:1182–93.
Perakis SS, Sinkhorn ER, Catricala CE, Bullen TD, Fitzpatrick JA, Hynicka JD, Cromack K. 2013. Forest calcium depletion and biotic retention along a soil nitrogen gradient. Ecol Appl 23:1947–61.
Perakis SS, Sinkhorn ER, Compton JE. 2011. δ 15N constraints on long-term nitrogen balances in temperate forests. Oecologia 167:793–807.
Perakis SS, Tepley AJ, Compton JE. 2015. Disturbance and topography shape nitrogen availability and δ 15N over long-term forest succession. Ecosystems 18:573–88.
Pess GR, McHenry ML, Beechie TJ, Davies J. 2008. Biological impacts of the Elwha River dams and potential salmonid responses to dam removal. Northwest Science 82:72–90.
Phillips EL, Donaldson WR. 1972. Washington climate for these counties: Clallam, Grays Harbor, Jefferson, Pacific, and Wahkiakum counties. Cooperative extension service publication EM 3708. Pullman (WA): Washington State University. 91p.
Phillips JD, Slattery MC, Musselman ZA. 2004. Dam-to-delta sediment inputs and storage in the lower Trinity River, Texas. Geomorphology 62:17–34.
Pinay G, Clement JC, Naiman RJ. 2002. Basic principles and ecological consequences of changing water regimes on nitrogen cycling in fluvial systems. Environ Manag 30:481–91.
Poff NL, Zimmerman JKH. 2010. Ecological responses to altered flow regimes: a literature review to inform the science and management of environmental flows. Freshw Biol 55:194–205.
Pohl MP. 1999. The dams of the Elwha River: downstream impacts and policy implications. PhD dissertation. Tempe (AZ): Arizona State University. 296p.
Pohl MP. 2004. Channel bed mobility downstream from the Elwha dams, Washington. Prof Geogr 56:422–31.
Porder S, Ramachandran S. 2013. The phosphorus concentration of common rocks—a potential driver of ecosystem P status. Plant Soil 367:41–55.
Prescott CE, Vesterdal L, Pratt J, Venner KH, de Montigny LM, Trofymow JA. 2000. Nutrient concentrations and nitrogen mineralization in forest floors of single species conifer plantations in coastal British Columbia. Can J For Res 30:1341–52.
Radwan MA, Shumway JS, Debell DS, Kraft JM. 1991. Variance in response of pole-size trees and seedlings of Douglas-fir and western hemlock to nitrogen and phosphorus fertilizers. Can J For Res 21:1431–8.
Randle TJ, Bountry JA, Ritchie AC, Wille K. 2015. Large-scale dam removal on the Elwha River, Washington, USA: erosion of reservoir sediment. Geomorphology 246:709–28.
Rhoades C, Binkley D, Oskarsson H, Stottlemyer R. 2008. Soil nitrogen accretion along a floodplain terrace chronosequence in northwest Alaska: influence of the nitrogen-fixing shrub Shepherdia canadensis. Ecoscience 15:223–30.
Ricker MC, Stolt MH, Donohue SW, Blazejewski GA, Zavada MS. 2013. Soil organic carbon pools in riparian landscapes of southern New England. Soil Sci Soc Am J 77:1070–9.
Rood SB, Goater LA, Gill KM, Braatne JH. 2011. Sand and sandbar willow: a feedback loop amplifies environmental sensitivity at the riparian interface. Oecologia 165:31–40.
Saint-Laurent D, Beaulac-Gervais V, Berthelot JS. 2014. Comparison of soil organic carbon and total nitrogen contents in inundated and non-inundated zones in southern Quebec, Canada. Catena 113:1–8.
Sanderman J, Lohse KA, Baldock JA, Amundson R. 2009. Linking soils and streams: sources and chemistry of dissolved organic matter in a small coastal watershed. doi:10.1029/2008WR006977.
Šantrůčková H, Bird MI, Lloyd J. 2000. Microbial processes and carbon-isotope fractionation in tropical and temperate grassland soils. Funct Ecol 14:108–14.
Schwendenmann L. 2000. Soil properties of boreal riparian plant communities in relation to natural succession and clear-cutting, Peace River Lowlands, Wood Buffalo National Park, Canada. Water Air Soil Pollut 122:449–67.
Scott EE, Perakis SS, Hibbs DE. 2008. δ 15N patterns of Douglas-fir and red alder riparian forests in the Oregon coast range. For Sci 54:140–7.
Shafroth PB, Hartt C, Perry LG, Braatne JH. 2017. Effects of dams and geomorphic context on riparian forests of the Elwha River, Washington. Ecosphere.
Southwell M, Thoms M. 2011. Patterns of nutrient concentrations across multiple floodplain surfaces in a large dryland river system. Geogr Res 49:431–43.
Steiger J, Gurnell AM. 2003. Spatial hydrogeomorphological influences on sediment and nutrient deposition in riparian zones: observations from the Garonne River, France. Geomorphology 49:1–23.
Stevenson BA, Kelly EF, McDonald EV, Busacca AJ. 2005. The stable carbon isotope composition of soil organic carbon and pedogenic carbonates along a bioclimatic gradient in the Palouse region, Washington State, USA. Geoderma 124:37–47.
Tashe NC, Schmidt MG. 2003. The influence of understory vine maple on forest floor mineral soil properties in coastal temperate forests. Can J Soil Sci 83:35–44.
Toda Y, Ikeda S, Kumagai K, Asano T. 2005. Effects of flood flow on flood plain soil and riparian vegetation in a gravel river. J Hydraul Eng-Asce 131:950–60.
Trudell SA, Rygiewicz PT, Edmonds RL. 2004. Patterns of nitrogen and carbon stable isotope ratios in macrofungi, plants and soils in two old-growth conifer forests. New Phytol 164:317–35.
Turk TD, Schmidt MG, Roberts NJ. 2008. The influence of bigleaf maple on forest floor and mineral soil properties in a coniferous forest in coastal British Columbia. For Ecol Manag 255:1874–82.
Turner DP, Franz EH. 1985. The influence of western hemlock and western redcedar on microbial numbers, nitrogen mineralization, and nitrification. Plant Soil 88:259–67.
Uliassi DD, Ruess RW. 2002. Limitations to symbiotic nitrogen fixation in primary succession on the Tanana River floodplain. Ecology 83:88–103.
van Coller AL, Rogers KH, Heritage GL. 2000. Riparian vegetation-environment relationships: complimentarity of gradients versus patch hierarchy approaches. J Veg Sci 11:337–50.
van Huysen TL, Harmon ME, Perakis SS, Chen H. 2013. Decomposition and nitrogen dynamics of 15N-labeled leaf, root, and twig litter in temperate coniferous forests. Oecologia 173:1563–73.
Van Miegroet H, Cole DW. 1984. The impact of nitrification on soil acidification and cation leaching in a red alder ecosystem. J Environ Qual 13:586–90.
Walker LR. 1989. Soil-nitrogen changes during primary succession on a floodplain in Alaska, USA. Arct Alp Res 21:341–9.
Warrick JA, Bountry JA, East AE, Magirl CS, Randle TJ, Gelfenbaum G, Ritchie AC, Pess GR, Leung V, Duda JJ. 2015. Large-scale dam removal on the Elwha River, Washington, USA: source-to-sink sediment budget and synthesis. Geomorphology 246:729–50.
Williams GP, Wolman MG. 1984. Downstream Effects of Dams on Alluvial Rivers. Washington, D.C.: U.S. Geological Survey.
Yakir D, Israeli Y. 1995. Reduced solar irradiance effects on net primary productivity (NPP) and the δ 13C and δ 18O values in plantations of Musa sp., Musaceae. Geochim Et Cosmochim Acta 59:2149–51.
Acknowledgements
Financial support for this work was provided through the U.S. Geological Survey’s Park Oriented Biological Support Program. We thank Olympic National Park for permission to sample within the park. Kurt Jenkins, USGS FRESC, provided the stable isotope samples. Hillary Eichler, Jennifer Lee, Shea McDonald, and Brittany Wilcox with the Peninsula College NSF REU program, and Steven Acker, Sarah Beldin, Cynthia Pritekel, and Jean Snyder assisted with sample collection. Christina Catricala assisted with sample analyses. Aaron Freeman produced Figure 1. Vanessa Beauchamp, Emily Bernhardt, and three anonymous reviewers provided useful comments on previous versions of the manuscript. Any use of trade names is for descriptive purposes only and does not imply endorsement by the US Government.
Author information
Authors and Affiliations
Corresponding author
Additional information
Author contributions
LGP analyzed the data and wrote the paper. PBS conceived of and designed the study, performed the research, and contributed to the paper SSP designed the study, performed the research, and contributed to the paper.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Perry, L.G., Shafroth, P.B. & Perakis, S.S. Riparian Soil Development Linked to Forest Succession Above and Below Dams Along the Elwha River, Washington, USA. Ecosystems 20, 104–129 (2017). https://doi.org/10.1007/s10021-016-0080-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-016-0080-1