
Abstract
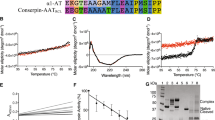
Serpin constitute about 10% of blood protein and are associated with mutations that results in aberrant intermolecular linkages which leads to polymer formation. Studies with short peptides have shown promise in depolymerization of serpins however a reactive center loop based peptide also makes the serpin inactive. A chemical chaperone based approach is a better option in terms of maintaining activity and retarding polymerization but not much is known about its binding and mechanism. Specific target for chemical chaperones and its effectiveness across many serpin is not known. We did an analysis of serpin cavity using CASTp and show that cavities are distributed throughout the molecule where the largest cavities are generally present in areas of major conformational change like shutter region, helix D and helix F. An analysis of different conformational states of serpins showed that this large cavity undergoes increase in size in latent and cleaved states as compared to native state. We targeted serpins with a variety of carbohydrate, methylamine and amino acid based chemical chaperones and selected those that have highest binding energy across different serpins to assess their ability to bind large cavities. The results show that carbohydrate based chemical chaperone like sorbitol, sucrose, arabitol and trehalose and amino acid based chaperones like dopamine, phenylalanine, arginine and glutamic acid are the most effective in binding serpins. Most of these chemical chaperone interacted with residues in the shutter region and the helix D arm at the C-terminal which are part of the largest cavities. We selected the carbohydrate based chemical chaperone with best binding energies and did experimental study under the condition that induce polymerization and show that indeed they were able to retard polymer formation with moderate effect on inhibition rates. However a fluorometric study with native antithrombin showed that chemical chaperone may effect the conformation of the proteins. Our study shows that chemical chaperones have the best binding affinities for the cavities around shutter region and helix D and that a cavity targeting based approach seems to be a better option for retarding polymerization in serpins, but a thorough analysis of its effect on folding, inhibition and cofactor binding is required.



Similar content being viewed by others


Abbreviations
- Serpin:
-
Serine protease inhibitors
- ATIII:
-
Antithrombin
- HCFII:
-
Heparin cofactor II
- PAI:
-
Plasminogen activator inhibitor
- PC1:
-
Protein C1-inhibitor
- ADT-Vina:
-
Autodock tools vina
- CASTp:
-
Computed atlas of surface topography of proteins
- RCL:
-
Reactive center loop
- TMAO:
-
Trimethylamine N-oxide
References
Devlin GL, Bottomley SP (2005) A protein family under ‘stress’-serpin stability, folding and misfolding. Front Biosci 10:288–299
Stein PE, Carrell RW (1995) What do dysfunctional serpins tell us about molecular mobility and disease? Struct Biol 2:96–113
Huntington JA, Read RJ, Carrell RW (2000) Structure of a serpin-protease complex shows inhibition by deformation. Nature 407:923–926
Mast AE, Enghild JJ, Pizzo S, Salvesen G (1991) Analysis of the plasma elimination kinetics and conformational stabilities of native, proteinase-complexed, and reactive site cleaved serpins: comparison of alpha 1-proteinase inhibitor, alpha 1-antichymotrypsin, antithrombin III, alpha 2-antiplasmin, angiotensinogen and ovalbumin. Biochemistry 30:1723–1730
Silverman GA, Bird PI, Carrell RW, Church FC, Coughlin PB, Gettins PG, Irving JA, Lomas DA, Luke CJ, Moyer RW, Pemberton PA, Remold-O’Donnell E, Salvesen GS, Travis J, Whisstock JC (2001) The serpins are an expanding superfamily of structurally similar but functionally diverse proteins. J Biol Chem 276:33293–33296
Carrell RW, Tavis J (1985) Alpha 1 antitrypsin and the serpins variation and counter variation. Trends Biochem Sci 8:20–24
Lomas DA, Evans DL, Finch JT, Carrell RW (1992) The mechanism of Z α-1 antitrypsin accumulation in the liver. Nature 357:605–607
Mast AE, Enghild JJ, Salvesen G (1992) Conformation of the reactive site loop of alpha 1-proteinase inhibitor probed by limited proteolysis. Biochemistry 31:2720–2728
Beauchamp NJ, Pike RN, Daly M, Butler L, Makris M, Dafforn TR, Zhou A, Fitton HL, Preston FE, Peake IR, Carrell RW (1998) Antithrombin Wibble and Wobble (T85M/K):archetypal conformational diseases with in vivo latent-transition, thrombosis, and heparin activation. Blood 92:2696–706
Burrows JA, Willis LK, Perlmutter DH (2000) Chemical chaperones mediate increased secretion of mutant α-1 antitrypsin (a1-AT) Z: a potential pharmacological strategy for prevention of liver injury and emphysema in α-1 antitrypsin deficiency. Proc Natl Acad Sci 97:1796–1801
Zhou A, Stein PE, Huntington JA, Carrell RW (2003) Serpin polymerisation is prevented by a hydrogen-bond network which is centred on His 334 and stabilised by glycerol. J Biol Chem 278:15116–15122
Sharp LK, Mallya M, Kinghorn KJ, Wang Z, Crowther DC, Huntington JA, Belorgey D, Lomas DA (2006) Sugar and alcohol molecules provide a therapeutic strategy for the serpinopathies that cause dementia and cirrhosis. FEBS J 273:2540–2552
Lee C, Maeng JS, Kocher JP, Lee B, Yu MH (2001) Cavities of α-1 antitrypsin that play structural and functional roles. Protein Sci 10:1446–1453
Binkowski TA, Naghibzadeh S, Liang J (2003) CASTp: Computed Atlas of Surface Topography of proteins. Nucleic Acids Res 31:3352–3355
Liang J, Edelsbrunner H, Fu P, Sudhakar PV, Subramaniam S (1998) Analytical shape computing of macromolecules I: molecular area and volume through alpha shape. Proteins 33:1–17
Trott O, Olson AJ (2010) AutoDock Vina: improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J Comput Chem 31:455–461
Morris GM, Huey R, Lindstrom W, Sanner MF, Belew RK, Goodsell DS, Olson AJ (2009) AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J Comput Chem 30:2785–2791
Jairajpuri MA, Lu A, Desai U, Olson ST, Bjork I, Bock SC (2003) Antithrombin III phenylalanines 122 and 121 contribute to its high affinity for heparin and its conformational activation. J Biol Chem 278:15941–15950
Jairajpuri MA, Lu A, Bock SC (2002) Elimination of P1 arginine 393 interactions with underlying glutamic acid 255 partially activates antithrombin III for thrombin inhibition but not factor Xa inhibition. J Biol Chem 277:24460–24465
Krem MM, Di Cera E (2003) Conserved Ser residues, the Shutter Region, and Speciation in Serpin Evolution. J Biol Chem 278:3810–3814
Lee KN, Park SD, Yu MH (1996) Probing the native strain in alpha1-antitrypsin. Nat Struct Biol 3:497–500
Potempa J, Korzus E, Travis J (1994) The serpin superfamily of proteinase inhibitors: structure, function and regulation. J Mol Biol 269:15957–1560
Whisstock JC, Bottomley SP (2006) Molecular gymnastics: serpin structure, folding and misfolding. Curr Opin Struct Biol 16:761–768
Gooptu B, Lomas DA (2009) Conformational pathology of the serpins: themes, variations, and therapeutic strategies. Annu Rev Biochem 78:147–176
Parfrey H, Mahadeva R, Ravenhill NA, Zhou A, Dafforn TR, Foreman RC, Lomas DA (2003) Targeting a surface cavity of α1-antitrypsin to prevent conformational disease. J Biol Chem 278:33060–33066
Im H, Seo EJ, Yu MH (1999) Metastability in the inhibitory mechanism of human α-1 antitrypsin. J Biol Chem 274:11072–11077
Belzar KJ, Zhou A, Carrell RW, Gettins PG, Huntington JA (2002) Helix D elongation and allosteric activation of antithrombin. J Biol Chem 277:8551–8558
Acknowledgments
This research was supported by grants from Department of Biotechnology and University Grant Commission, Government of India. PS is supported by a grant from Rajiv Gandhi National Fellowship. AN is supported by an Innovation in Science Pursuit for Inspired Research (INSPIRE) fellowship from Department of Science and Technology, Government of India. SK is supported by a fellowship from University Grant Commission.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Singh, P., Khan, M.S., Naseem, A. et al. Analysis of surface cavity in serpin family reveals potential binding sites for chemical chaperone to reduce polymerization. J Mol Model 18, 1143–1151 (2012). https://doi.org/10.1007/s00894-011-1110-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00894-011-1110-8