
Abstract
Key message
Planting sites that are cooler, with more precipitation, early springs, high TD and hot dry summers reduce bole taper in young Douglas-fir trees. Seed-source climates that are drier, with high TD and low snowfall produce seed-sources with lower taper.
Abstract
Analysis of a 10 year reciprocal transplant study was performed to determine the influence of seed-source and local planting site climates on the bole taper of 8995 Douglas-fir (Pseudotsuga menziesii var. menziesii) trees. Trees were planted at 9 sites across Oregon and Washington, with 120 known families taken from 12 seed-source regions across California, Oregon, and Washington. Diameter at breast height (DBH) ranged from 0.1 to 22.9 cm, with tree ages ranging from 2 year-old seedlings to 12 year-old trees. Changes in Gini coefficients (∆G) of diameters along tree boles, as a surrogate for changes in taper, were modeled as a function of age, site climate, and seed-source climate (Wang et al., in PLoS One 11, 2016) using universal response functions (URF) (Wang et al., in Ecol Appl 20:153–163, 2010). Lower Gini coefficients come from less tapered boles, i.e., more cylindrically shaped boles. There was significant influence on taper from five site climate variables: mean annual temperature (MAT), mean annual precipitation (MAP), beginning of frost-free period (bFFP), difference between minimum and maximum monthly temperatures (TD), and summer heat-moisture index (SHM). These results suggest that cooler years with more precipitation, early springs, large TD and hot dry summers reduce tree taper. There was significant influence on taper from three seed-source climate variables: percent precipitation as snow (PAS), Hargrave’s climate moisture deficit (CMD), and TD. These results suggest dry regions with large ranges in monthly temperatures and low snowfall will produce seed-sources with lower Gini coefficients. Projections under future climates using an ensemble model show areas where G estimates are currently highest showed increases in G, while areas with the lowest G estimates decreased. Most of the Douglas-fir region shows declines in Gini coefficients under high emissions scenarios. These predicted changes in taper have direct implications for wood volume and carbon mass estimates of trees under future climates.

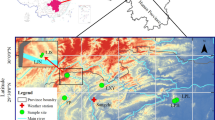
source locations are shown in the map on the left, and planting site locations are shown in the map on the right. Symbols are used to differentiate between seed-source regions. The seed-source regions are: CACST (light blue circles), CAKLA (purple triangles), CASIERRA (pink triangles), ORCSTS (green circles), ORSISL (yellow triangles), ORSISH (orange triangles), ORCSTN (dark blue circles), ORCASL (dark red triangles), ORCASH (red triangles), WACST (white circles), WACASL (black triangles), WACASH (white triangles)



Similar content being viewed by others

Data availability
Data available upon request.
Code availability
Code available upon request.
References
Bansal S, St. Clair JB, Harrington CA, Gould PJ (2015) Impact of climate change on cold hardiness of Douglas-fir (Pseudotsuga menziesii): environmental and genetic considerations. Glob Change Biol 21:3814–3826. https://doi.org/10.1111/gcb.12958
Beedlow PA, Waschmann RS, Lee EH, Tingey DT (2017) Seasonal patterns of bole water content in old growth Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco). Agric for Meteorol 242:109–119. https://doi.org/10.1016/j.agrformet.2017.04.017
Bruchert F, Gardiner B (2006) The effect of wind exposure on the tree aerial architecture and biomechanics of Sitka spruce (Picea sitchensis, Pinaceae). Am J Bot 93:1512–1521. https://doi.org/10.3732/ajb.93.10.1512
Campbell RK, Sugano AI (1993) Genetic variation and seed zones of Douglas-fir in the Siskiyou National Forest. U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station, Portland
Ceriani L, Verme P (2012) The origins of the Gini index: extracts from Variabilità e Mutabilità (1912) by Corrado Gini. J Econ Inequal 10:421–443. https://doi.org/10.1007/s10888-011-9188-x
Collins M, Knutti R, Arblaster J, et al (2013) Long-term Climate Change: Projections, Commitments and Irreversibility. Clim. Change 2013 Phys. Sci. Basis Contrib. Work. Group Fifth Assess. Rep. Intergov. Panel Clim. Change 1029–1136
del Río M, Bravo-Oviedo A, Ruiz-Peinado R, Condés S (2019) Tree allometry variation in response to intra- and inter-specific competitions. Trees 33:121–138. https://doi.org/10.1007/s00468-018-1763-3
Ford KR, Harrington CA, Bansal S et al (2016) Will changes in phenology track climate change? A study of growth initiation timing in coast Douglas-fir. Glob Change Biol 22:3712–3723. https://doi.org/10.1111/gcb.13328
Ford KR, Harrington CA, St. Clair JB (2017) Photoperiod cues and patterns of genetic variation limit phenological responses to climate change in warm parts of species’ range: modeling diameter-growth cessation in coast Douglas-fir. Glob Change Biol 23:3348–3362. https://doi.org/10.1111/gcb.13690
Gray LK, Hamann A (2013) Tracking suitable habitat for tree populations under climate change in western North America. Clim Change 117:289–303. https://doi.org/10.1007/s10584-012-0548-8
Hijmans RJ (2020) raster: geographic data analysis and modeling. Version 3.3–7URL https://CRAN.R-project.org/package=raster. Accessed 10 Jan 2019
King DA (1986) Tree form, height growth, and susceptibility to wind damage in Acer saccharum. Ecology 67:980–990
Krakowski J, Stoehr MU (2009) Coastal Douglas-fir provenance variation: patterns and predictions for British Columbia seed transfer. Ann For Sci 66:811–811. https://doi.org/10.1051/forest/2009069
Leites LP, Robinson AP, Rehfeldt GE et al (2012) Height-growth response to climatic changes differs among populations of Douglas-fir: a novel analysis of historic data. Ecol Appl 22:154–165. https://doi.org/10.1890/11-0150.1
McGown KI, O’Hara KL, Youngblood A (2016) Patterns of size variation over time in ponderosa pine stands established at different initial densities. Can J for Res 46:101–113. https://doi.org/10.1139/cjfr-2015-0096
Nigh G, Smith W (2012) Effect of climate on lodgepole pine stem taper in British Columbia, Canada. Forestry 85:579–587. https://doi.org/10.1093/forestry/cps063
Pinheiro J, Bates D, DebRoy S, et al (2019) Linear and nonlinear mixed effects models. Version 3.1–141URL https://CRAN.R-project.org/package=nlme. Accessed 10 Jan 2019
Poorter H, Niklas KJ, Reich PB et al (2012) Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental control: Tansley review. New Phytol 193:30–50. https://doi.org/10.1111/j.1469-8137.2011.03952.x
QGIS Development Team (2019) QGIS geographic information system. Version 3.6.3. Open source geospatial foundation. URL: http://qgis.org. Accessed 10 Jan 2019
R Core Team (2019) R: a language and environment for statistical computing. Version 3.5.3. Vienna, Austria. URL https://www.R-project.org/. Accessed 10 Jan 2019
Rehfeldt GE (1994) Evolutionary genetics, the biological species, and the ecology of the Interior cedar–hemlock forests. In: Proceedings of Interior cedar–hemlock–white pine forests: ecology and management. Washington State University, Pullman, WA, pp 91–100
Schneider R, Franceschini T, Fortin M, Saucier J-P (2018) Climate-induced changes in the stem form of 5 North American tree species. For Ecol Manag 427:446–455. https://doi.org/10.1016/j.foreco.2017.12.026
Sorensen FC (1983) Geographic variation in seedling Douglas-Fir (Pseudotsuga Menziesii) from the Western Siskiyou Mountains of Oregon. Ecology 64:696–702. https://doi.org/10.2307/1937191
St Clair JB, Mandel NL, Vance-Borland KW (2005) Genecology of Douglas Fir in Western Oregon and Washington. Ann Bot 96:1199–1214. https://doi.org/10.1093/aob/mci278
Valentine HT, Mäkelä A, Green EJ et al (2012) Models relating stem growth to crown length dynamics: application to loblolly pine and Norway spruce. Trees 26:469–478. https://doi.org/10.1007/s00468-011-0608-0
Vejpustková M, Čihák T (2019) Climate Response of Douglas Fir Reveals Recently Increased Sensitivity to Drought Stress in Central Europe. Forests 10:97. https://doi.org/10.3390/f10020097
Wang T, O’Neill GA, Aitken SN (2010) Integrating environmental and genetic effects to predict responses of tree populations to climate. Ecol Appl 20:153–163. https://doi.org/10.1890/08-2257.1
Wang T, Hamann A, Spittlehouse D, Carroll C (2016) Locally downscaled and spatially customizable climate data for historical and future periods for North America. PLoS One 11:e0156720. https://doi.org/10.1371/journal.pone.0156720
Warren JM, Meinzer FC, Brooks JR, Domec JC (2005) Vertical stratification of soil water storage and release dynamics in Pacific Northwest coniferous forests. Agric for Meteorol 130:39–58. https://doi.org/10.1016/j.agrformet.2005.01.004
Wensel LC (2000) The relationship between tree diameter growth and climate for coniferous species in northern California. Can J for Res 30:1463–1471
White TL, Ching KK (1985) Provenance study of Douglas-fir in the Pacific Northwest region IV. Field performance at age 25 years. Silvae Genet 34:84–90
Yun K, Hsiao J, Jung M-P et al (2017) Can a multi-model ensemble improve phenology predictions for climate change studies? Ecol Model 362:54–64. https://doi.org/10.1016/j.ecolmodel.2017.08.003
Funding
Funding provided by US Forest Service, and Bureau of Land Management.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Vospernik.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
468_2021_2254_MOESM3_ESM.png
Supplementary file3 Modeled Gini value estimates for populations experiencing the same shared climate but different historical climate norms. Shared climate values are based on values from the area encompassed by the green dot for RCP4.5 2020 data (PNG 2621 KB)
468_2021_2254_MOESM4_ESM.png
Supplementary file4 Gini value estimates from the ensemble model for populations grown in their native climate from 2010 to 2020 (PNG 1093 KB)
Rights and permissions
About this article

{kind=link}
{kind=link}
{kind=link}
Cite this article
Jones, D.A., Harrington, C.A. & St. Clair, J.B. Influence of climate on annual changes in Douglas-fir stem taper. Trees 36, 849–861 (2022). https://doi.org/10.1007/s00468-021-02254-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-021-02254-0