
Abstract
Sodium deficiency elevates aldosterone, which in addition to epithelial tissues acts on the brain to promote dysphoric symptoms and salt intake. Aldosterone boosts the activity of neurons that express 11-beta-hydroxysteroid dehydrogenase type 2 (HSD2), a hallmark of aldosterone-sensitive cells. To better characterize these neurons, we combine immunolabeling and in situ hybridization with fate mapping and Cre-conditional axon tracing in mice. Many cells throughout the brain have a developmental history of Hsd11b2 expression, but in the adult brain one small brainstem region with a leaky blood–brain barrier contains HSD2 neurons. These neurons express Hsd11b2, Nr3c2 (mineralocorticoid receptor), Agtr1a (angiotensin receptor), Slc17a6 (vesicular glutamate transporter 2), Phox2b, and Nxph4; many also express Cartpt or Lmx1b. No HSD2 neurons express cholinergic, monoaminergic, or several other neuropeptidergic markers. Their axons project to the parabrachial complex (PB), where they intermingle with AgRP-immunoreactive axons to form dense terminal fields overlapping FoxP2 neurons in the central lateral subnucleus (PBcL) and pre-locus coeruleus (pLC). Their axons also extend to the forebrain, intermingling with AgRP- and CGRP-immunoreactive axons to form dense terminals surrounding GABAergic neurons in the ventrolateral bed nucleus of the stria terminalis (BSTvL). Sparse axons target the periaqueductal gray, ventral tegmental area, lateral hypothalamic area, paraventricular hypothalamic nucleus, and central nucleus of the amygdala. Dual retrograde tracing revealed that largely separate HSD2 neurons project to pLC/PB or BSTvL. This projection pattern raises the possibility that a subset of HSD2 neurons promotes the dysphoric, anorexic, and anhedonic symptoms of hyperaldosteronism via AgRP-inhibited relay neurons in PB.













Similar content being viewed by others

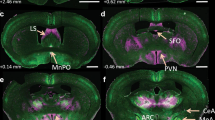
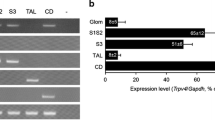
Notes
In preparing this manuscript, we also found evidence of Ai75 neurotoxicity in Slc17a7-IRES3-Cre mice (http://connectivity.brain-map.org/transgenic/experiment/304698566), which appears to be age-related and specific to regions that express that Cre-driver gene (hippocampus, cerebral cortex).
References
Aponte Y, Atasoy D, Sternson SM (2011) AGRP neurons are sufficient to orchestrate feeding behavior rapidly and without training. Nat Neurosci 14(3):351–355
Arriza JL, Simerly RB, Swanson LW, Evans RM (1988) The neuronal mineralocorticoid receptor as a mediator of glucocorticoid response. Neuron 1(9):887–900
Askew ML, Muckelrath HD, Johnston JR, Curtis KS (2015) Neuroanatomical association of hypothalamic HSD2-containing neurons with ERalpha, catecholamines, or oxytocin: implications for feeding? Front Syst Neurosci 9:91
Aston-Jones G, Delfs JM, Druhan J, Zhu Y (1999) The bed nucleus of the stria terminalis. A target site for noradrenergic actions in opiate withdrawal. Ann N Y Acad Sci 877:486–498
Bard P (1928) A diencephalic mechanism for the expression of rage with special reference to the sympathetic nervous system. Am J Physiol 84(3):490–515
Betley JN, Cao ZF, Ritola KD, Sternson SM (2013) Parallel, redundant circuit organization for homeostatic control of feeding behavior. Cell 155(6):1337–1350
Broadwell RD, Sofroniew MV (1993) Serum proteins bypass the blood-brain fluid barriers for extracellular entry to the central nervous system. Exp Neurol 120(2):245–263
Chang RB, Strochlic DE, Williams EK, Umans BD, Liberles SD (2015) Vagal sensory neuron subtypes that differentially control breathing. Cell 161(3):622–633
Chang SE, Smedley EB, Stansfield KJ, Stott JJ, Smith KS (2017) optogenetic inhibition of ventral pallidum neurons impairs context-driven salt seeking. J Neurosci 37(23):5670–5680
Chen S, Aston-Jones G (1995) Evidence that cholera toxin B subunit (CTb) can be avidly taken up and transported by fibers of passage. Brain Res 674(1):107–111
Craig AD (2002) How do you feel? Interoception: the sense of the physiological condition of the body. Nat Rev Neurosci 3(8):655–666
Dai JX, Hu ZL, Shi M, Guo C, Ding YQ (2008) Postnatal ontogeny of the transcription factor Lmx1b in the mouse central nervous system. J Comp Neurol 509(4):341–355
de Kloet ER, Otte C, Kumsta R, Kok L, Hillegers MH, Hasselmann H, Kliegel D, Joels M (2016) Stress and depression: a crucial role of the mineralocorticoid receptor. J Neuroendocrinol 28(8)
Denton DA, Sabine JR (1961) The selective appetite for Na ions shown by Na ion-deficient sheep. J Physiol 157:97–116
Diaz R, Brown RW, Seckl JR (1998) Distinct ontogeny of glucocorticoid and mineralocorticoid receptor and 11beta-hydroxysteroid dehydrogenase types I and II mRNAs in the fetal rat brain suggest a complex control of glucocorticoid actions. J Neurosci 18(7):2570–2580
Dong HW (2008) Allen reference atlas: a digital color brain atlas of the C57Black/6J male mouse. Wiley. ix, Hoboken, 366 p. p
Dong HW, Petrovich GD, Watts AG, Swanson LW (2001) Basic organization of projections from the oval and fusiform nuclei of the bed nuclei of the stria terminalis in adult rat brain. J Comp Neurol 436(4):430–455
Egli RE, Kash TL, Choo K, Savchenko V, Matthews RT, Blakely RD, Winder DG (2005) Norepinephrine modulates glutamatergic transmission in the bed nucleus of the stria terminalis. Neuropsychopharmacology 30(4):657–668
Epstein AN (1982) Mineralocorticoids and cerebral angiotensin may act together to produce sodium appetite. Peptides 3(3):493–494
Essner RA, Smith AG, Jamnik AA, Ryba AR, Trutner ZD, Carter ME (2017) AgRP neurons can increase food intake during conditions of appetite suppression and inhibit anorexigenic parabrachial neurons. J Neurosci 37(36):8678–8687
Evans LC, Ivy JR, Wyrwoll C, McNairn JA, Menzies RI, Christensen TH, Al-Dujaili EA, Kenyon CJ, Mullins JJ, Seckl JR, Holmes MC, Bailey MA (2016) Conditional deletion of Hsd11b2 in the brain causes salt appetite and hypertension. Circulation 133(14):1360–1370
Fitts DA (1991) Effects of lesions of the ventral ventral median preoptic nucleus or subfornical organ on drinking and salt appetite after deoxycorticosterone acetate or yohimbine. Behav Neurosci 105(5):721–726
Fluharty SJ, Epstein AN (1983) Sodium appetite elicited by intracerebroventricular infusion of angiotensin II in the rat: II. Synergistic interaction with systemic mineralocorticoids. Behav Neurosci 97(5):746–758
Formenti S, Bassi M, Nakamura NB, Schoorlemmer GH, Menani JV, Colombari E (2013) Hindbrain mineralocorticoid mechanisms on sodium appetite. Am J Physiol Regul Integr Comp Physiol 304(3):R252–R259
Franklin KBJ, Paxinos G (2013) Paxinos and Franklin’s The mouse brain in stereotaxic coordinates. Academic Press, Amsterdam
Funder J, Myles K (1996) Exclusion of corticosterone from epithelial mineralocorticoid receptors is insufficient for selectivity of aldosterone action: in vivo binding studies. Endocrinology 137(12):5264–5268
Funder JW, Pearce PT, Smith R, Smith AI (1988) Mineralocorticoid action: target tissue specificity is enzyme, not receptor, mediated. Science 242(4878):583–585
Garfield AS, Li C, Madara JC, Shah BP, Webber E, Steger JS, Campbell JN, Gavrilova O, Lee CE, Olson DP, Elmquist JK, Tannous BA, Krashes MJ, Lowell BB (2015) A neural basis for melanocortin-4 receptor-regulated appetite. Nat Neurosci 18(6):863–871
Geerling JC, Loewy AD (2006a) Aldosterone-sensitive neurons in the nucleus of the solitary tract: bidirectional connections with the central nucleus of the amygdala. J Comp Neurol 497(4):646–657
Geerling JC, Loewy AD (2006b) Aldosterone-sensitive neurons in the nucleus of the solitary tract: efferent projections. J Comp Neurol 497(2):223–250
Geerling JC, Loewy AD (2006c) Aldosterone-sensitive NTS neurons are inhibited by saline ingestion during chronic mineralocorticoid treatment. Brain Res 1115(1):54–64
Geerling JC, Loewy AD (2007a) 11beta-hydroxysteroid dehydrogenase 2 vs. transgene: discrepant loci of expression in the adult brain. Am J Physiol Renal Physiol 293(1):F440–F441 (author reply F442–F443)
Geerling JC, Loewy AD (2007b) Sodium depletion activates the aldosterone-sensitive neurons in the NTS independently of thirst. Am J Physiol Regul Integr Comp Physiol 292(3):R1338–R1348
Geerling JC, Loewy AD (2007c) Sodium deprivation and salt intake activate separate neuronal subpopulations in the nucleus of the solitary tract and the parabrachial complex. J Comp Neurol 504(4):379–403
Geerling JC, Loewy AD (2008) Central regulation of sodium appetite. Exp Physiol 93(2):177–209
Geerling JC, Loewy AD (2009) Aldosterone in the brain. Am J Physiol Renal Physiol 297(3):F559–F576
Geerling JC, Engeland WC, Kawata M, Loewy AD (2006a) Aldosterone target neurons in the nucleus tractus solitarius drive sodium appetite. J Neurosci 26(2):411–417
Geerling JC, Kawata M, Loewy AD (2006b) Aldosterone-sensitive neurons in the rat central nervous system. J Comp Neurol 494(3):515–527
Geerling JC, Chimenti PC, Loewy AD (2008) Phox2b expression in the aldosterone-sensitive HSD2 neurons of the NTS. Brain Res 1226:82–88
Geerling JC, Shin JW, Chimenti PC, Loewy AD (2010) Paraventricular hypothalamic nucleus: axonal projections to the brainstem. J Comp Neurol 518(9):1460–1499
Geerling JC, Stein MK, Miller RL, Shin JW, Gray PA, Loewy AD (2011) FoxP2 expression defines dorsolateral pontine neurons activated by sodium deprivation. Brain Res 1375:19–27
Gomez-Sanchez EP (1986) Intracerebroventricular infusion of aldosterone induces hypertension in rats. Endocrinology 118(2):819–823
Gong S, Zheng C, Doughty ML, Losos K, Didkovsky N, Schambra UB, Nowak NJ, Joyner A, Leblanc G, Hatten ME, Heintz N (2003) A gene expression atlas of the central nervous system based on bacterial artificial chromosomes. Nature 425(6961):917–925
Gonzalez AD, Wang G, Waters EM, Gonzales KL, Speth RC, Van Kempen TA, Marques-Lopes J, Young CN, Butler SD, Davisson RL, Iadecola C, Pickel VM, Pierce JP, Milner TA (2012) Distribution of angiotensin type 1a receptor-containing cells in the brains of bacterial artificial chromosome transgenic mice. Neuroscience 226:489–509
Grippo AJ, Moffitt JA, Beltz TG, Johnson AK (2006) Reduced hedonic behavior and altered cardiovascular function induced by mild sodium depletion in rats. Behav Neurosci 120(5):1133–1143
Gross PM, Wall KM, Pang JJ, Shaver SW, Wainman DS (1990) Microvascular specializations promoting rapid interstitial solute dispersion in nucleus tractus solitarius. Am J Physiol 259(6 Pt 2):R1131–R1138
Guyenet PG (2006) The sympathetic control of blood pressure. Nat Rev Neurosci 7(5):335–346
Haque M, Wilson R, Sharma K, Mills NJ, Teruyama R (2015) Localisation of 11beta-hydroxysteroid dehydrogenase type 2 in mineralocorticoid receptor expressing magnocellular neurosecretory neurones of the rat supraoptic and paraventricular nuclei. J Neuroendocrinol 27(11):835–849
Haskell-Luevano C, Monck EK (2001) Agouti-related protein functions as an inverse agonist at a constitutively active brain melanocortin-4 receptor. Regul Pept 99(1):1–7
Herbert H, Saper CB (1990) Cholecystokinin-, galanin-, and corticotropin-releasing factor-like immunoreactive projections from the nucleus of the solitary tract to the parabrachial nucleus in the rat. J Comp Neurol 293(4):581–598
Hlavacova N, Jezova D (2008) Chronic treatment with the mineralocorticoid hormone aldosterone results in increased anxiety-like behavior. Horm Behav 54(1):90–97
Hlavacova N, Wes PD, Ondrejcakova M, Flynn ME, Poundstone PK, Babic S, Murck H, Jezova D (2012) Subchronic treatment with aldosterone induces depression-like behaviours and gene expression changes relevant to major depressive disorder. Int J Neuropsychopharmacol 15(2):247–265
Holmes MC, Sangra M, French KL, Whittle IR, Paterson J, Mullins JJ, Seckl JR (2006) 11beta-Hydroxysteroid dehydrogenase type 2 protects the neonatal cerebellum from deleterious effects of glucocorticoids. Neuroscience 137(3):865–873
Jarvie BC, Palmiter RD (2017) HSD2 neurons in the hindbrain drive sodium appetite. Nat Neurosci 20(2):167–169
Jellinck PH, Monder C, McEwen BS, Sakai RR (1993) Differential inhibition of 11 beta-hydroxysteroid dehydrogenase by carbenoxolone in rat brain regions and peripheral tissues. J Steroid Biochem Mol Biol 46(2):209–213
Kang BJ, Chang DA, Mackay DD, West GH, Moreira TS, Takakura AC, Gwilt JM, Guyenet PG, Stornetta RL (2007) Central nervous system distribution of the transcription factor Phox2b in the adult rat. J Comp Neurol 503(5):627–641
Kawai Y (2018) Differential ascending projections from the male rat caudal nucleus of the tractus solitarius: an interface between local microcircuits and global macrocircuits. Front Neuroanat 12:63
Kawai Y, Senba E (1996) Organization of excitatory and inhibitory local networks in the caudal nucleus of tractus solitarius of rats revealed in in vitro slice preparation. J Comp Neurol 373(3):309–321
Koneru B, Bathina CS, Cherry BH, Mifflin SW (2014) Mineralocorticoid receptor in the NTS stimulates saline intake during fourth ventricular infusions of aldosterone. Am J Physiol Regul Integr Comp Physiol 306(1):R61–R66
Krashes MJ, Koda S, Ye C, Rogan SC, Adams AC, Cusher DS, Maratos-Flier E, Roth BL, Lowell BB (2011) Rapid, reversible activation of AgRP neurons drives feeding behavior in mice. J Clin Invest 121(4):1424–1428
Krashes MJ, Shah BP, Madara JC, Olson DP, Strochlic DE, Garfield AS, Vong L, Pei H, Watabe-Uchida M, Uchida N, Liberles SD, Lowell BB (2014) An excitatory paraventricular nucleus to AgRP neuron circuit that drives hunger. Nature 507(7491):238–242
Lesman-Leegte I, Jaarsma T, Sanderman R, Linssen G, van Veldhuisen DJ (2006) Depressive symptoms are prominent among elderly hospitalised heart failure patients. Eur J Heart Fail 8(6):634–640
Loewy A, Spyer K (1990) Central regulation of autonomic functions. Oxford University Press, New York
Luquet S, Perez FA, Hnasko TS, Palmiter RD (2005) NPY/AgRP neurons are essential for feeding in adult mice but can be ablated in neonates. Science 310(5748):683–685
Madisen L, Zwingman TA, Sunkin SM, Oh SW, Zariwala HA, Gu H, Ng LL, Palmiter RD, Hawrylycz MJ, Jones AR, Lein ES, Zeng H (2010) A robust and high-throughput Cre reporting and characterization system for the whole mouse brain. Nat Neurosci 13(1):133–140
Malinow KC, Lion JR (1979) Hyperaldosteronism (Conn’s disease) presenting as depression. J Clin Psychiatry 40(8):358–359
Matsuda T, Hiyama TY, Niimura F, Matsusaka T, Fukamizu A, Kobayashi K, Kobayashi K, Noda M (2017) Distinct neural mechanisms for the control of thirst and salt appetite in the subfornical organ. Nat Neurosci 20(2):230–241
McCance RA (1936) Experimental human salt deficiency. Lancet 1:823–830
McKinley MJ, Badoer E, Oldfield BJ (1992) Intravenous angiotensin II induces Fos-immunoreactivity in circumventricular organs of the lamina terminalis. Brain Res 594(2):295–300
Morris MJ, Na ES, Grippo AJ, Johnson AK (2006) The effects of deoxycorticosterone-induced sodium appetite on hedonic behaviors in the rat. Behav Neurosci 120(3):571–579
Murck H, Buttner M, Kircher T, Konrad C (2014) Genetic, molecular and clinical determinants for the involvement of aldosterone and its receptors in major depression. Nephron Physiol 128(1–2):17–25
Naray-Fejes-Toth A, Fejes-Toth G (2007) Novel mouse strain with Cre recombinase in 11beta-hydroxysteroid dehydrogenase-2-expressing cells. Am J Physiol Renal Physiol 292(1):F486–F494
Naray-Fejes-Toth A, Colombowala IK, Fejes-Toth G (1998) The role of 11beta-hydroxysteroid dehydrogenase in steroid hormone specificity. J Steroid Biochem Mol Biol 65(1–6):311–316
Nijenhuis WA, Oosterom J, Adan RA (2001) AgRP(83–132) acts as an inverse agonist on the human-melanocortin-4 receptor. Mol Endocrinol 15(1):164–171
Ollmann MM, Wilson BD, Yang YK, Kerns JA, Chen Y, Gantz I, Barsh GS (1997) Antagonism of central melanocortin receptors in vitro and in vivo by agouti-related protein. Science 278(5335):135–138
Palmiter RD (2018) The parabrachial nucleus: CGRP neurons function as a general alarm. Trends Neurosci 41(5):280–293
Pardridge WM, Mietus LJ (1979) Transport of steroid hormones through the rat blood-brain barrier. Primary role of albumin-bound hormone. J Clin Invest 64(1):145–154
Parvizi J, Damasio A (2001) Consciousness and the brainstem. Cognition 79(1–2):135–160
Price JL, Drevets WC (2012) Neural circuits underlying the pathophysiology of mood disorders. Trends Cogn Sci 16(1):61–71
Reincke M (2018) Anxiety, depression, and impaired quality of life in primary aldosteronism: why we shouldn’t ignore it! J Clin Endocrinol Metab 103(1):1–4
Resch JM, Fenselau H, Madara JC, Wu C, Campbell JN, Lyubetskaya A, Dawes BA, Tsai LT, Li MM, Livneh Y, Ke Q, Kang PM, Fejes-Toth G, Naray-Fejes-Toth A, Geerling JC, Lowell BB (2017) Aldosterone-sensing neurons in the NTS exhibit state-dependent pacemaker activity and drive sodium appetite via synergy with angiotensin II signaling. Neuron 96(1):190–206 (e197)
Robson AC, Leckie CM, Seckl JR, Holmes MC (1998) 11 Beta-hydroxysteroid dehydrogenase type 2 in the postnatal and adult rat brain. Brain Res Mol Brain Res 61(1–2):1–10
Roland BL, Li KX, Funder JW (1995) Hybridization histochemical localization of 11 beta-hydroxysteroid dehydrogenase type 2 in rat brain. Endocrinology 136(10):4697–4700
Rowland NE, Fregly MJ (1988) Characteristics of thirst and sodium appetite in mice (Mus musculus). Behav Neurosci 102(6):969–974
Rutledge T, Reis VA, Linke SE, Greenberg BH, Mills PJ (2006) Depression in heart failure a meta-analytic review of prevalence, intervention effects, and associations with clinical outcomes. J Am Coll Cardiol 48(8):1527–1537
Sakai RR, Ma LY, Zhang DM, McEwen BS, Fluharty SJ (1996) Intracerebral administration of mineralocorticoid receptor antisense oligonucleotides attenuate adrenal steroid-induced salt appetite in rats. Neuroendocrinology 64(6):425–429
Sakai RR, McEwen BS, Fluharty SJ, Ma LY (2000) The amygdala: site of genomic and nongenomic arousal of aldosterone-induced sodium intake. Kidney Int 57(4):1337–1345
Saper CB (2016) The house alarm. Cell Metab 23(5):754–755
Sawchenko PE, Brown ER, Chan RK, Ericsson A, Li HY, Roland BL, Kovacs KJ (1996) The paraventricular nucleus of the hypothalamus and the functional neuroanatomy of visceromotor responses to stress. Prog Brain Res 107:201–222
Sequeira SM, Geerling JC, Loewy AD (2006) Local inputs to aldosterone-sensitive neurons of the nucleus tractus solitarius. Neuroscience 141(4):1995–2005
Shields AD, Wang Q, Winder DG (2009) alpha2A-adrenergic receptors heterosynaptically regulate glutamatergic transmission in the bed nucleus of the stria terminalis. Neuroscience 163(1):339–351
Shin JW, Geerling JC, Loewy AD (2008) Inputs to the ventrolateral bed nucleus of the stria terminalis. J Comp Neurol 511(5):628–657
Shin JW, Geerling JC, Loewy AD (2009) Vagal innervation of the aldosterone-sensitive HSD2 neurons in the NTS. Brain Res 1249:135–147
Shin JW, Geerling JC, Stein MK, Miller RL, Loewy AD (2011) FoxP2 brainstem neurons project to sodium appetite regulatory sites. J Chem Neuroanat 42(1):1–23
Simpson JB, Routtenberg A (1978) Subfornical organ: a dipsogenic site of action of angiotensin II. Science 201(4353):379–381
Song K, Allen AM, Paxinos G, Mendelsohn FA (1992) Mapping of angiotensin II receptor subtype heterogeneity in rat brain. J Comp Neurol 316(4):467–484
Sonino N, Fallo F, Fava GA (2006) Psychological aspects of primary aldosteronism. Psychother Psychosom 75(5):327–330
Sonino N, Tomba E, Genesia ML, Bertello C, Mulatero P, Veglio F, Fava GA, Fallo F (2011) Psychological assessment of primary aldosteronism: a controlled study. J Clin Endocrinol Metab 96(6):E878–E883
Stornetta RL, Hawelu-Johnson CL, Guyenet PG, Lynch KR (1988) Astrocytes synthesize angiotensinogen in brain. Science 242(4884):1444–1446
Sunn N, McKinley MJ, Oldfield BJ (2003) Circulating angiotensin II activates neurones in circumventricular organs of the lamina terminalis that project to the bed nucleus of the stria terminalis. J Neuroendocrinol 15(8):725–731
Szabo NE, da Silva RV, Sotocinal SG, Zeilhofer HU, Mogil JS, Kania A (2015) Hoxb8 intersection defines a role for Lmx1b in excitatory dorsal horn neuron development, spinofugal connectivity, and nociception. J Neurosci 35(13):5233–5246
Terenzi MG, Ingram CD (1995) A combined immunocytochemical and retrograde tracing study of noradrenergic connections between the caudal medulla and bed nuclei of the stria terminalis. Brain Res 672(1–2):289–297
Tindell AJ, Smith KS, Pecina S, Berridge KC, Aldridge JW (2006) Ventral pallidum firing codes hedonic reward: when a bad taste turns good. J Neurophysiol 96(5):2399–2409
Tong Q, Ye CP, Jones JE, Elmquist JK, Lowell BB (2008) Synaptic release of GABA by AgRP neurons is required for normal regulation of energy balance. Nat Neurosci 11(9):998–1000
Ueda K, Okamura N, Hirai M, Tanigawara Y, Saeki T, Kioka N, Komano T, Hori R (1992) Human P-glycoprotein transports cortisol, aldosterone, and dexamethasone, but not progesterone. J Biol Chem 267(34):24248–24252
Uhr M, Holsboer F, Muller MB (2002) Penetration of endogenous steroid hormones corticosterone, cortisol, aldosterone and progesterone into the brain is enhanced in mice deficient for both mdr1a and mdr1b P-glycoproteins. J Neuroendocrinol 14(9):753–759
Velema MS, de Nooijer AH, Burgers VWG, Hermus A, Timmers H, Lenders JWM, Husson O, Deinum J (2017) Health-related quality of life and mental health in primary aldosteronism: a systematic review. Horm Metab Res 49(12):943–950
Verstegen AMJ, Vanderhorst V, Gray PA, Zeidel ML, Geerling JC (2017) Barrington’s nucleus: neuroanatomic landscape of the mouse “pontine micturition center”. J Comp Neurol
Walker DL, Toufexis DJ, Davis M (2003) Role of the bed nucleus of the stria terminalis versus the amygdala in fear, stress, and anxiety. Eur J Pharmacol 463(1–3):199–216
Wu Q, Boyle MP, Palmiter RD (2009) Loss of GABAergic signaling by AgRP neurons to the parabrachial nucleus leads to starvation. Cell 137(7):1225–1234
Zardetto-Smith AM, Beltz TG, Johnson AK (1994) Role of the central nucleus of the amygdala and bed nucleus of the stria terminalis in experimentally-induced salt appetite. Brain Res 645(1–2):123–134
Zeisel A, Hochgerner H, Lonnerberg P, Johnsson A, Memic F, van der Zwan J, Haring M, Braun E, Borm LE, La Manno G, Codeluppi S, Furlan A, Lee K, Skene N, Harris KD, Hjerling-Leffler J, Arenas E, Ernfors P, Marklund U, Linnarsson S (2018) Molecular architecture of the mouse nervous system. Cell 174(4):999–1014 (e1022)
Zhang ZH, Kang YM, Yu Y, Wei SG, Schmidt TJ, Johnson AK, Felder RB (2006) 11beta-hydroxysteroid dehydrogenase type 2 activity in hypothalamic paraventricular nucleus modulates sympathetic excitation. Hypertension 48(1):127–133
Acknowledgements
We thank Richard Palmiter and Aniko Fejes-Toth for sharing Hsd11b2 Cre-driver mice; David Olson for L10GFP Cre-reporter mice; and Justin Grobe, Huxing Cui, and Kenji Saito for Agtr1a-GFP mice and brainstem tissue. Hideki Enomoto of Kobe University generously provided an aliquot of GP-anti-Phox2b, and Carmen Birchmeier generously provided an aliquot of GP-anti-Lmx1b antiserum. Finally, we thank Brad Lowell for mentorship and for material support for much of this work.
Funding
Grant sponsors: NIH F32 DK103387 (JMR). NIH K08 NS099425 (JCG). Aging Mind and Brain Initiative, University of Iowa Center for Aging (JCG).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethical approval
All procedures performed in studies involving animals were in accordance with the ethical standards of the institution or practice at which the studies were conducted.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplemental figure A1
: Whole-slide imaging of a representative brain after immunohistochemical staining for HSD2 using nickel-DAB for maximum sensitivity (1-in-3, 40 µm tissue sections through a full mouse brain). No brain regions contain HSD2 immunoreactivity visible at low magnification (PDF 619 KB)
Supplemental figure A2
: High magnification images of every brain region previously reported to express Hsd11b2 mRNA or contain immunoreactivity for HSD2 (page 2). The NTS contains intense HSD2 immunoreactivity, but no other region contains labeling above background levels. Scale bars are 200 µm and apply to all panels (PDF 1302 KB)
Supplemental figure A3
: Whole-slide imaging of a representative brain with tdTomato nuclear reporter for Hsd11b2 (Hsd11b2Cre;Ai75-lsl-Tomato, 1-in-3 series of 40 µm sections). This Ai75 Cre-reporter is expressed in sparse, small cells in every brain region, at every level, and in neurons in a variety of regions outside the NTS, including the cerebellar granule cell layer and some of its connections (pontine nuclei, lateral reticular nucleus, external cuneate nucleus), superior colliculus, and several parts of the diencephalon (PDF 10583 KB)
Supplemental figure A3
: Whole-slide imaging of a representative brain with tdTomato nuclear reporter for Hsd11b2 (Hsd11b2Cre;Ai75-lsl-Tomato, 1-in-3 series of 40 µm sections). This Ai75 Cre-reporter is expressed in sparse, small cells in every brain region, at every level, and in neurons in a variety of regions outside the NTS, including the cerebellar granule cell layer and some of its connections (pontine nuclei, lateral reticular nucleus, external cuneate nucleus), superior colliculus, and several parts of the diencephalon (PDF 3241 KB)
Supplemental figure A4
: Examples of Syp-mCherry labeling in HSD2 axons and boutons in brain tissue sections from an Hsd11b2-Cre mouse injected with AAV-DIO-Syp-mCherry into the NTS (injection site in Figure 6). We immunolabeled mCherry using nickel-DAB (black) and added a light Nissl counterstain (blue-gray) for cytoarchitectural reference. For each image, an inset (adapted from Figure 7) shows the level and location (red box). (A) Lightly labeled axons pass through the intermediate reticular formation of the caudal medulla; (B) dense axon-terminal field in the pre-locus coeruleus (pLC); (C) less-dense axons and boutons in the rostral pLC and medial parabrachial nucleus (PB); (D) dense axon-terminal field in the central lateral PB (PBcL), bordering the superior cerebellar peduncle; (E) lightly labeled axons course dorsally around the sensory and motor trigeminal nuclei before turning caudally to reach the PB; (F) light axonal branching and bouton labeling in the lateral hypothalamic area (LHA)/parasubthalamic nucleus (PSTN); (G) small cluster of boutons formed by a single branching axon in the central nucleus of the amygdala (CeA); (H) few, sparse axon branches and boutons in the ventral midbrain; (I-J) caudal and middle levels of the dense, focal axon-terminal field in the ventrolateral bed nucleus of the stria terminalis (BSTvL). Scale bars are 50 µm (PDF 28465 KB)
Rights and permissions
About this article

Cite this article
Gasparini, S., Resch, J.M., Narayan, S.V. et al. Aldosterone-sensitive HSD2 neurons in mice. Brain Struct Funct 224, 387–417 (2019). https://doi.org/10.1007/s00429-018-1778-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00429-018-1778-y