
Abstract
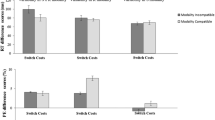
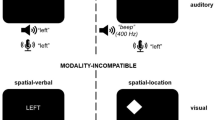
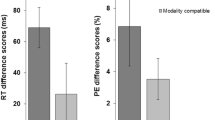
Previous research has indicated that modality switching is considerably affected by modality compatibility. It has been shown that switch costs are higher for switching between relatively incompatible sensory-motor modality mappings (i.e., auditory-manual and visual-vocal) compared to switching between compatible mappings (i.e., auditory-vocal and visual-manual). So far, however, it has been unclear whether these findings are influenced by learning processes resulting from very small stimulus sets and a large number of stimulus repetitions. In the present study, we investigated the role of learning concept-to-category associations (Experiment 1) as well as influences of learning concept-to-modality mappings (Experiment 2) on sensory-motor modality switching in semantic categorizations. The results of both experiments revealed shorter overall reaction times due to learning. Additionally, learning of concept-to-category associations (Experiment 1) led to a significant reduction of modality switch costs. Interestingly, however, modality-compatibility effects were neither significantly influenced by learning of concept-to-category associations nor by learning of concept-to-modality mappings. Thus, the present study provides first evidence that learning on the semantic level influences modality switching but it does not significantly affect modality compatibility.





Similar content being viewed by others
Notes
Sensory-motor modality switching represents a specific case of task switching. That is, the tasks that have to be switched are defined by their different sensory-motor modality mappings and not by the performed cognitive operation (e.g., semantic categorization). More precisely, participants switch between different sensory-motor modality mappings while performing cognitive operations that are fixed. Thus, the resulting switch costs are also termed as modality switch costs.
To further draw inferences about this null-effects, we calculated the Bayes factor (BF) that is a ratio that indicates how much more likely given data fit under one model than under another model. We conducted a Bayesian three-way repeated measures ANOVA with JASP, a freely available statistical program (Love et al., 2015; Rouder, Speckman, Sun, Morey, & Iverson, 2009). At this point, it has to be mentioned that Bayesian analyses always require the selection of prior probability distribution for the model parameters, always implicating a certain amount of subjectivity or prior knowledge about the effect that is to be studied. For the present analyses we used the default prior scales of JASP, which assumes a Cauchy distribution of possible effect sizes (cf. Rouder et al., 2009)–even though this slightly favors the null model because it assumes relatively large effect sizes for the alternative model. To draw inferences about the interaction modality compatibility × stimulus set, we compared the model including this interaction and the three main effects to the main effects-only model. The resulting BF01 = 3.5 indicates that the present data are about three times more likely under the main-effects-only model than under the interaction model. As regards the interaction transition × modality compatibility × stimulus set, the comparison of the full model to the model without the three-way interaction revealed the Bayes factor BF01 = 3.1, indicating that the data are more likely under the model without the three-way interaction than under the full model.
In order to evaluate the evidence of this numerical trend, which shows a reversed pattern compared to RT switch costs, we calculated the Bayes factor regarding the interaction modality transition × modality compatibility in the large stimulus set in a similar manner as described in Footnote 2. Specifically, we compared the model including the interaction modality transition × modality compatibility to the main-effects-only model. The resulting Bayes factor BF10 = 0.8 neither supports the alternative nor the null model, indicating that the error data of the large stimulus set are not very conclusive regarding this interaction.
In the practice blocks a consistent or variable concept-to-modality mapping was not possible because the sensory-motor modality mapping was fixed (e.g., a consistent auditory presentation of the stimuli in the auditory-vocal practice block).
Similar to Experiment 1 (see footnote 2), we calculated the Bayes factors to further draw inferences about these null-effects. As regards the interaction modality compatibility × concept-to-modality mapping, we compared the model including this interaction to a model without this interaction. The resulting Bayes factor BF01 = 5.0 indicates that the data were more likely under the model without the interaction than under the model including the interaction. A similar procedure with regard to the interaction transition × concept-to-modality revealed the Bayes factor BF01 = 5.2, indicating that the data are about five times as likely under the model without the interaction than under the interaction model. As regards the interaction transition × modality compatibility × concept-to-modality mapping, we compared the full model to the model without the three-way interaction. The analysis revealed the Bayes factor BF01 = 4.0, indicating that the data are about four times as likely under the model without the three-way interaction than under the full model.
References
Allport, A., & Wylie, G. (2000). Task-switching, stimulus-response bindings, and negative priming. In S. Monsell & J. Driver (Eds.), Control of cognitive processes: Attention and performance XVIII (pp. 35–70). Cambridge; MA: MIT Press.
Arrington, C. M., & Logan, G. D. (2004). Episodic and semantic components of the compound-stimulus strategy in the explicit task-cuing procedure. Memory Cogn, 32, 965–978.
Dobbins, I. G., Schnyer, D. M., Verfaellie, M., & Schacter, D. L. (2004). Cortical activity reductions during repetition priming can result from rapid response learning. Nature, 428, 316–319.
Greenwald, A. G. (1972). On doing two things at once: Time sharing as a function of ideomotor compatibility. Journal of Experimental Psychology, 94, 52–57.
Hazeltine, E., Ruthruff, E., & Remington, R. W. (2006). The role of sensory and output modality pairings in dual-task performance: Evidence for content-dependent central interference. Cognitive Psychology, 52, 291–345.
Hazeltine, E., Teague, D., & Ivry, R. B. (2002). Simultaneous dual-task performance reveals parallel response selection after practice. Journal of Experimental Psychology: Human Perception and Performance, 28, 527–545.
Horner, A. J., & Henson, R. N. (2011). Stimulus–response bindings code both abstract and reverses multiple levels of response representation. Memory & Cognition, 39, 1457–1471.
Kiesel, A., Steinhauser, M., Wendt, M., Falkenstein, M., Jost, K., Philipp, A. M., & Koch, I. (2010). Control and interference in task switching—a review. Psychological Bulletin, 136, 849–874.
Koch, I., & Allport, A. (2006). Cue-based preparation and stimulus-based priming of tasks in task switching. Memory & Cognition, 34, 433–444.
Koch, I., Prinz, W., & Allport, A. (2005). Involuntary retrieval in alphabet-arithmetic tasks: task-mixing and task-switching costs. Psychol Res, 69, 252–261.
Logan, G. D. (1988). Toward an instance theory of automatization. Psychological Review, 95, 492–527.
Logan, G. D. (1990). Repetition priming and automaticity: common underlying mechanisms? Cognitive Psychology, 22, 1–35.
Love, J., Selker, R., Verhagen, J., Marsman, M., Gronau, Q. F., Jamil, T., Smira, M., Epskamp, S., Wild, A., Morey, R., Rouder, J. & Wagenmakers, E. J. (2015). JASP (Version 0.8). Computer software.
Pashler, H. (1994). Dual-task interference in simple tasks: data and theory. Psychological Bulletin, 116, 220–244.
Rouder, J. N., Speckman, P. L., Sun, D., Morey, R. D., & Iverson, G. (2009). Bayesian t tests for accepting and rejecting the null hypothesis. Psychonomic Bulletin and Review, 16, 225–237.
Schaeffner, S., Koch, I., & Philipp, A.M. (2016a). The role of sensory-motor modality compatibility in language processing. Psychological Research, 80, 212-223.
Schaeffner, S., Koch, I., & Philipp, A. M. (2016b). Semantic effects on sensory-motor modality switching. Journal of Cognitive Psychology, 28, 726–742.
Schumacher, E. H., Seymour, T. L., Glass, J. M., Fencsik, D. E., Lauber, E. J., Kieras, D. E., & Meyer, D. E. (2001). Virtually perfect time sharing in dual-task performance: Uncorking the central cognitive bottleneck. Psychological Science, 12, 101–108.
Stephan, D. N., & Koch, I. (2010). Central cross-talk in task switching: Evidence from manipulating sensory-output modality compatibility. Journal of Experimental Psychology, Learning, Memory, and Cognition, 36, 1075–1081.
Stephan, D. N., & Koch, I. (2011). The role of sensory–output modality compatibility in task switching. Psychological Research, 75, 491–498.
Strobach, T., Liepelt, R., Pashler, H., Frensch, P. A., & Schubert, T. (2013). Effects of extensive dual-task practice on processing stages in simultaneous choice tasks. Attention, Perception, & Psychophysics, 75, 900–920.
Strobach, T., Liepelt, R., Schubert, T., & Kiesel, A. (2012). Task switching: Effects of practice on switch and mixing costs. Psychological Research, 76, 74–83.
Van Selst, M., Ruthruff, E., & Johnston, J. C. (1999). Can practice eliminate the psychological refractory period effect? Journal of Experimental Psychology: Human Perception and Performance, 25, 1268–1283.
Waszak, F., Hommel, B., & Allport, A. (2003). Task-switching and long-term priming: Role of episodic stimulus–task bindings in task-shift costs. Cognitive Psychology, 46, 361–413.
Wickens, C. D., Sandry, D. L., & Vidulich, M. (1983). Compatibility and resource competition between modalities of input, central processing, and output. Human Factors, 25, 227–248.
Acknowledgements
This research was funded by DFG Grant PH 156/3-2. We thank Eric Ruthruff and Wilfried Kunde for their helpful comments on an earlier version of this paper. Furthermore, we would like to thank Axel Mayer for his help in conducting the Bayesian analyses.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Funding
This research was funded by DFG grant PH 156/3-2.
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki Declaration and its later amendments or comparable ethical standards.
Informed consent
Informed consent was obtained from all individual participants included in the study.
Appendix
Appendix
Overview of all stimuli of the small and of the large stimulus set of Experiment 1 as well as their assignment to the two semantic categories.
Small stimulus set | Large stimulus set | ||
|---|---|---|---|
Natural | Manmade | Natural | Manmade |
Kind [child] | Geld [money] | Adler [eagle] | Anker [anchor] |
Rose [rose] | Sofa [sofa] | Affe [monkey] | Auto [car] |
Eule [owl] | Vase [vase] | Apfel [apple] | Bett [bed] |
Floh [flea] | Hemd [shirt] | Baum [tree] | Bild [picture] |
Biene [bee] | Boot [boat] | ||
Blume [flower] | Brief [letter] | ||
Delfin [dolphin] | Brille [glasses] | ||
Ente [duck] | Brücke [bridge] | ||
Esel [donkey] | Buch [book] | ||
Fisch [fish] | Dose [can] | ||
Frau [woman] | Eimer [bucket] | ||
Frosch [frog] | Fenster [window] | ||
Fuchs [fox] | Gürtel [belt] | ||
Gurke [cucumber] | Haus [house] | ||
Huhn [chicken] | Hose [trousers] | ||
Hund [dog] | Jacke [jacket] | ||
Kaktus [cactus] | Kette [chain] | ||
Kamel [camel] | Kissen [pillow] | ||
Katze [cat] | Knopf [button] | ||
Kiwi [Kiwi] | Korb [basket] | ||
Kürbis [pumpkin] | Krone [crown] | ||
Löwe [lion] | Lampe [lamp] | ||
Mann [man] | Mauer [wall] | ||
Nuss [nut] | Mütze [cap] | ||
Palme [palm] | Regal [shelf] | ||
Pfau [peacock] | Schal [scarf] | ||
Pferd [horse] | Schuh [shoe] | ||
Pilz [mushroom] | Seil [rope] | ||
Ratte [rat] | Stuhl [chair] | ||
Salat [salad] | Teller [plate] | ||
Schaf [sheep] | Teppich [carpet] | ||
Schwan [swan] | Tisch [table] | ||
Schwein [pig] | Topf [pot] | ||
Spinne [spider] | Treppe [stair] | ||
Tiger [tiger] | Turm [tower] | ||
Vogel [bird] | Tüte [bag] | ||
Wurm [worm] | Waage [balance] | ||
Zebra [zebra] | Weste [vest] | ||
Ziege [goat] | Wippe [seesaw] | ||
Zwiebel [onion] | Zaun [fence] | ||
Rights and permissions
About this article

Cite this article
Schaeffner, S., Koch, I. & Philipp, A.M. The role of learning in sensory-motor modality switching. Psychological Research 82, 955–969 (2018). https://doi.org/10.1007/s00426-017-0872-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00426-017-0872-8