
Abstract
Myocardial connexin 43 (Cx43) forms gap junctions and hemichannels, and is also present within subsarcolemmal mitochondria. The protein is phosphorylated by several kinases including mitogen-activated protein kinase (MAPK), protein kinase C (PKC), and casein kinase 1 (CK1). A reduction in Cx43 content abrogates myocardial infarct size reduction by ischemic preconditioning (IPC). The present study characterizes the contribution of Cx43 phosphorylation towards mitochondrial function, hemichannel activity, and the cardioprotection by IPC in wild-type (WT) mice and in mice in which Cx43-phosphorylation sites targeted by above kinases are mutated to non-phosphorylatable residues (Cx43MAPKmut, Cx43PKCmut, and Cx43CK1mut mice). The amount of Cx43 in the left ventricle and in mitochondria was reduced in all mutant strains compared to WT mice and Cx43 phosphorylation was altered at residues not directly targeted by the mutations. Whereas complex 1 respiration was reduced in all strains, complex 2 respiration was decreased in Cx43CK1mut mice only. In Cx43 epitope-mutated mice, formation of reactive oxygen species and opening of the mitochondrial permeability transition pore were not affected. The hemichannel open probability was reduced in Cx43PKCmut and Cx43CK1mut but not in Cx43MAPKmut cardiomyocytes. Infarct size in isolated saline-perfused hearts after ischemia/reperfusion (45 min/120 min) was comparable between genotypes and was significantly reduced by IPC (3 × 3 min ischemia/5 min reperfusion) in WT, Cx43MAPKmut, and Cx43PKCmut, but not in Cx43CK1mut mice, an effect independent from the amount of Cx43 and the probability of hemichannel opening. Taken together, our study shows that alterations of Cx43 phosphorylation affect specific cellular functions and highlights the importance of Cx43 phosphorylation by CK1 for IPC’s cardioprotection.
Similar content being viewed by others

Introduction
Cell–cell communication in the ventricular myocardium is mediated via gap junctions, which are mainly formed by connexin 43 (Cx43). Six Cx43 proteins assemble into hexamers, the so-called connexons or hemichannels, and are transported to the sarcolemma, where two hemichannels from adjacent cells dock to each other and form gap junctions, which allow the passage of molecules up to 1.5 kDa in size and mediate electrical and metabolic cell coupling. Cx43 is part of a family comprising 21 members in humans with molecular weights ranging from 26 to 60 kDa and which are expressed in different organs and cell types. The connexin secondary structure consists of four transmembrane domains and a cytosolic amino-and carboxyterminus. Within the carboxyterminus of Cx43, at least 21 serine (S) residues are targeted and phosphorylated by different kinases, among them protein kinase B (PKB or AKT), protein kinase C (PKC), casein kinase 1 (CK1), calcium/calmodulin kinase II (CamKII), mitogen-activated protein kinase (MAPK), p34cdc2/cyclin B kinase (p34cdc2), and the pp60src kinase (src) (for review, see [69]).
S262 (similar to S279 and S282) is commonly suggested to be targeted by the extracellular-signal-related kinase (ERK) and the mutation of S262 to alanine increases gap junction permeability and inhibits desoxyribonucleic acid (DNA) synthesis. CK1 phosphorylates Cx43 at S325, S328, and S330 and mice in which S325/328/330 are mutated to alanines display disturbed gap junction formation and are susceptible to arrhythmias [54]. After phosphorylation of S373 by AKT, the interaction between Cx43 and zonula occludens 1 (ZO-1) is limited and the size of gap junctions is increased which may represent a first step in the disassembly process of gap junctions. Cx43 phosphorylation at S365 is also involved in the protein–protein interaction with ZO-1. Dephosphorylation of Cx43 at S365 is essential for the phosphorylation of Cx43 at the PKC-target site S368; therefore, the Cx43 phosphorylation at S365 is suggested to function as a “gatekeeper” [71]. Cx43 S365 phosphorylation is increased under conditions where the activity of protein kinase A (PKA) is enhanced; however, Cx43 is a poor substrate for a direct phosphorylation by PKA. PKC directly phosphorylates Cx43 at S368 and this phosphorylation reduces the intercellular communication via gap junctions. The role of Cx43 phosphorylation for gap junction dynamics, which is not only controlled by kinases but also by phosphatases [29], is reviewed in [69].
In addition to its localization at the sarcolemma, Cx43 is also present at the inner membrane of mitochondria isolated from ventricular tissue [55, 73], especially in subsarcolemmal mitochondria (SSM) and to a far lesser extent in interfibrillar mitochondria (IFM), which are located between the myofibrils [9, 75]. Compared to the detailed analysis of the phosphorylation of gap junctional Cx43, the phosphorylation of mitochondrial Cx43 is relatively unknown; however, phosphorylation at S262 and S368 has been detected [73, 76]. Within mitochondria, Cx43 influences mitochondrial respiration [7], formation of reactive oxygen species (ROS) [24, 68, 77], uptake of potassium ions [10], and opening of the mitochondrial permeability transition pore (MPTP) [20, 73].
Acute myocardial ischemia/reperfusion (I/R) injury elicits death of cardiomyocytes via necrosis, apoptosis, necroptosis, and pyroptosis (for review, see [26]). Both ischemic preconditioning (IPC) and ischemic postconditioning reduce myocardial infarct size after prolonged ischemia followed by reperfusion. In IPC, cycles of short, non-lethal episodes of ischemia/reperfusion (I/R) precedes the long-lasting ischemia, whereas in ischemic postconditioning, the short periods of I/R are applied at the onset of reperfusion following the long-lasting ischemic episode. Adenosine, bradykinin, opioids, and cytokines function as mediators or receptor-dependent triggers of cardioprotection. After a preconditioning stimulus, three major signaling pathways become activated, i.e., a) a pathway involving eNOS, protein kinase G and PKC; b) the RISK pathway (reperfusion injury salvage kinase pathway) including activation of AKT, ERK, and glycogen synthase kinase 3β; and c) the SAFE pathway (survival activating factor enhancement), in which cytokine receptors and janus-kinase/signal transducer and activator of transcription proteins are induced [25, 26]. Besides targeting the sarcoplasmic reticulum, the nucleus, as well as the sarcolemma (including gap junctions and hemichannels), mitochondria represent a common endpoint of cardioprotective maneuvers [6]. Whereas mitochondrial respiration declines after I/R, oxygen consumption is maintained after IPC. The role of ROS in the cardioprotection by IPC is complex: whereas IPC reduces the detrimental burst of ROS at the onset of reperfusion, small amounts of ROS also function as trigger molecules in the signaling cascade of IPC [66]. In addition, opening of mitochondrial ATP-dependent potassium channels is important to reduce myocardial I/R injury and the opening of these channels leads to ROS formation which activate protein kinases involved in cardioprotective signaling (for review, see [25]). Conditions favoring opening of the MPTP, which elicits mitochondrial swelling, rupture, and finally cell death, are induced by ischemia [47], and both ischemic pre- and postconditioning induce cardioprotection via the inhibition of MPTP opening at reperfusion. Also, mitochondrial dynamics, i.e., fusion and fission, are involved in I/R injury, whereby the inhibition of fission—and thereby a decreased fragmentation of mitochondrial networks—is effective in reducing I/R damage [49].
Cx43 is involved in myocardial I/R injury and also represents an important signaling molecule in the cardioprotection by IPC. The role of Cx43 in myocardial I/R damage comprises all aspects of Cx43 function: posttranslational modifications, protein amount, alterations of gap junction, and hemichannel opening as well as mitochondrial function [60]. Whereas ischemia is often associated with a dephosphorylation and lateralization of Cx43 at the cardiomyocyte plasma membrane [60], Cx43 phosphorylation, and localization is maintained after IPC [56, 61]. The cardioprotection by IPC depends on the presence of Cx43, since the genetic reduction of Cx43 abrogates the infarct size reduction by IPC [59, 62, 63]. Gap junctional coupling seems to be part of IPC’s cardioprotection, since blocking of Cx43-formed channels during the trigger phase abolishes the infarct size reduction of IPC [39] and IPC blocks chemical gap junctional coupling during ischemia (for review, see [46]). Plasmalemmal Cx43-formed hemichannels are predominantly closed under physiological conditions [1, 22], but they open after prolonged ischemia, which leads to cell swelling and finally cell death [65, 81]. Accordingly, the inhibition of Cx43-formed hemichannels by the peptide Gap19 reduces myocardial infarction after I/R [81] demonstrating the importance of the prevention of hemichannel opening for the reduction of myocardial damage after I/R. Since Cx43 affects mitochondrial function in several ways that influence cardioprotection [6, 12, 32], Cx43 within the the mitochondria is of special interest for myocardial I/R injury. Following I/R, phosphorylation of mitochondrial Cx43 is decreased [76]. Mitochondrial Cx43 is also involved in IPC, since a specific reduction of mitochondrial Cx43 abrogates preconditioning [55] and overexpression of mitochondrial Cx43 is sufficient to induce cell protection [43].
It is noteworthy that although ischemic postconditioning alters Cx43 phosphorylation [5, 23, 50], infarct size is effectively reduced in heterozygous Cx43-deficient mice [27]. Thus, despite its relevance for the cardioprotection by IPC, ischemic postconditioning is not dependend on Cx43.
Whereas Cx43 clearly contributes to I/R response, the role of Cx43 phosphorylation on the regulation of mitochondrial function, cardiomyocyte hemichannel activity, and cardioprotection by IPC is heretofore not well studied and, thus, is the goal of the present study.
Materials and methods
Animals
The generation of the mouse strains Cx43MAPKmut, Cx43PKCmut, and Cx43CK1mut has been described previously [30, 34]. In the line Cx43MAPKmut, serines 255, 262, 279, and 282 (which are targeted by MAPK) were replaced by non-phosphorylatable alanine residues; in the line Cx43PKCmut, the PKC-targeted residue serine 368 was replaced by alanine, and in the line Cx43CK1mut, serines at positions 325, 328, and 330 were replaced by alanine, tyrosine, and alanine, respectively. Mutated mice were backcrossed with C57Bl6/J mice for 10 generations (Fred Hutchinson Cancer Research Center). At the University of Giessen, mutated mice were backcrossed with C57Bl6/J mice after every 6–8 generations (Cx43MAPKmut), after 10 generations (Cx43PKCmut), and after 4–6 generations (Cx43CK1mut). C57Bl6/J mice (Janvier, Le Genest-Saint-Isles, France) served as controls. In addition, inducible Cx43 knockout mice (Cx43Cre−ER(T)/fl) which express 50% of Cx43 were studied [16]. Both male and female mice of 12–22 weeks of age (18–34 g) were analyzed. Mice were kept in dark/light cycles of 12 h each and had free access to standard chow and drinking water. The present study conforms to the Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health (NIH publication No. 85–23, revised 1996) and is in accordance with the ARRIVE (Animal research: Reporting in vivo experiments) guidelines. The study was approved by the animal welfare office of the Justus-Liebig-University Giessen, Germany (522_M) and by the committee on ethical usage of animals of Ghent University (ECD16-24.)
Isolation of subsarcolemmal mitochondria
Subsarcolemmal mitochondria (SSM) were isolated from left-ventricular (LV) tissue by differential centrifugation as already described [11] and were used to study mitochondrial function immediately after isolation. For Western Blot analysis, mitochondria were further purified by Percoll-gradient ultracentrifugation and were stored at − 80 °C. The purity of all mitochondrial preparations was verified by Western Blot analysis (Supplementary Fig. 3).
Western Blot analysis
Protein extraction from LV tissue samples and isolated mitochondria is described in detail in the online supplement. Antibodies were used against total Cx43, Cx43 phosphorylated at S262, S325/328/330, S365, S368, and S373. Antibodies against glyceraldehyde 3-phosphate dehydrogenase (GAPDH) and manganese superoxide dismutase (MnSOD) were used for normalization of total and mitochondrial proteins, respectively.
Mitochondrial respiration
Mitochondrial oxygen consumption was measured from 100 µg/ml mitochondrial proteins with a Clark-type electrode (Strathkelvin, Glasgow, UK) at 25 °C using glutamate and malate as substrates for complex 1 and succinate (in the presence of rotenone to inhibit complex 1) as substrate for complex 2. In addition, complex 4-mediated respiration was analyzed in the presence of N,N,N′,N′-tetramethyl-p-phenylenediamine (TMPD) and ascorbate, and uncoupling of oxidative phosphorylation was induced by the addition of FCCP (cyanide 4-(trifluoromethoxy)phenylhydrazone). Respiration was stimulated by the addition of 40 µmol/L ADP. Oxygen consumption is shown in nmol O2*min−1*mg protein−1 [4].
Formation of reactive oxygen species
ROS formation was detected from 50 µg mitochondrial proteins using 50 µmol/L AmplexUltraRed. Background fluorescence without mitochondria was subtracted and the slope of the fluorescence signal was calculated for 2 min [4].
Calcium-induced MPTP opening
The calcium retention capacities of 100 µg/ml SSM with glutamate and malate were measured at 25 °C in the presence of 0.5 µmol/L Calcium Green 5 N. Five µmol/L CaCl2 were added every third minute until a sudden increase in Calcium Green 5 N fluorescence occurred reflecting MPTP opening [4]. As a positive control, measurements were performed in the presence of 1 µmol/L Cyclosporine A (CsA), which inhibits MPTP opening.
Cardiomyocyte isolation and electrophysiological recordings
The isolation of LV cardiomyocytes from wild-type (WT), Cx43MAPKmut, Cx43PKCmut, and Cx43CK1mut mice was performed as previously described [42]. Mice were sacrificed by cervical dislocation. Following thoracotomy, the heart was quickly excised and transferred to a Langendorff apparatus and perfused at a constant flow (~ 3 ml/min) and temperature (37 °C). Ca2+-tolerant cells were used for the experiment within 6 h after isolation. Cardiomyocytes were studied using whole-cell voltage-clamp to record membrane currents. Patch-clamp experiments were conducted at room temperature (22 ± 1 °C).
Ischemia/reperfusion in vitro
Isolated mouse hearts were Langendorff-perfused as already described [4]. The I/R protocol comprised of a 5 min stabilization phase, followed by 45 min ischemia and 120 min reperfusion. IPC was performed by three preceding cycles of 3 min ischemia and 5 min reperfusion. At the end of the protocol, myocardial infarction was delineated by triphenyltetrazolium chloride (TTC) staining and quantified by planimetry [4].
Statistics
Data are presented as mean ± SEM. Western Blot data on the amount of Cx43 in total and mitochondrial protein extracts were compared by non-parametric Rank Sum test, and Western Blot data on phosphorylated Cx43, ROS formation, opening of Cx43-formed hemichannels, and sodium/calcium exchanger (NCX) parameters were analyzed by unpaired two-sided t test. The amount of phosphorylated and total Cx43 in isolated cardiomyocytes was compared by paired t test. The amount of mitochondrial protein isolated from LV tissue was compared by one-way ANOVA. Data on heart weight and heart weight/body weight, mitochondrial respiration, calcium-induced MPTP opening, myocardial infarction, end-diastolic pressure (EDP), and left-ventricular developed pressure (LVDP) were compared by two-way ANOVA followed by Holm–Sidak test. Analyses were done with SigmaStat 3.5 (Systat, Software GmbH, Erkrath, Germany) and OriginLab software (OriginLab Corporation, USA). A p value < 0.05 was considered to indicate a significant difference.
A detailed description of the Methods is given in the online supplement.
Results
The amount of Cx43 was measured by Western Blot analysis in total LV protein extracts of WT, Cx43MAPKmut, Cx43PKCmut, and Cx43CK1mut mice, respectively (Table 1). Whereas Cx43 was moderately reduced in Cx43MAPKmut mice, a more severe reduction of the protein was detected in Cx43CK1mut mice and especially in Cx43PKCmut mice (all strains p < 0.05). The phosphorylation of Cx43 was analyzed by Western blot at S262, S325/328/330, S365, S368, and S373. Antibodies against Cx43 phosphorylated at S255, S279, and S282 could not be validated in initial Western Blot analyses; therefore, the Cx43 phosphorylation status at residues targeted by MAPK was based on S262 only. In total LV protein extracts of Cx43MAPKmut mice, the ratio of phosphorylated over total Cx43 was not altered. In Cx43PKCmut and Cx43CK1mut mice, Cx43 phosphorylation was significantly decreased at S365, but increased at S262. Additionally, Cx43 phosphorylation was diminished at S325/328/330 and S373 in Cx43PKCmut and at S368 in Cx43CK1mut mice. Treatment of isolated cardiomyocytes from wild-type mice for 2 h or 6 h with inhibitors against PKC (GF109203X) or CK1 (CKI-7) did not increase Cx43 S262 phosphorylation or reduce Cx43 expression as in Cx43PKCmut or Cx43CK1mut mice (Supplementary Fig. 1). As expected, no immunoreactivity was detected using antibodies against S262 in Cx43MAPKmut, against S368 in Cx43PKCmut, and against S325/328/330 in Cx43CK1mut mice, which proves the specificity of the antibodies and the genotype of the mice (Supplementary Fig. 2).
SSM were isolated from LV tissue of WT, Cx43MAPKmut, Cx43PKCmut, and Cx43CK1mut mice, and the amount of isolated proteins per mg LV tissue was quantified. An exemplary Western Blot demonstrating the purity of mitochondrial preparations is demonstrated in Supplementary Fig. 3. A trend towards reduced amounts of isolated SSM was found in Cx43CK1mut mice; however, there was no significant difference (amount of isolated SSM in µg protein/mg LV tissue: WT: 2.34 ± 0.18, n = 8; Cx43MAPK: 1.72 ± 0.42, n = 5; Cx43PKCmut: 2.13 ± 0.25, n = 5; Cx43CK1mut: 1.50 ± 0.17, n = 6, p = ns). Western Blot analysis was performed to characterize the amount of Cx43 in SSM isolated from LV tissue of WT, Cx43MAPKmut, Cx43PKCmut, and Cx43CK1mut mice (Table 1). Compared to total LV proteins, similar reductions of mitochondrial Cx43 were detected in Cx43MAPKmut and Cx43CK1mut, whereas a further decrease to 9 ± 2% was measured in Cx43PKCmut SSM (all strains p < 0.05). The analysis of the phosphorylation status of mitochondrial Cx43 displayed decreased phosphorylation at S325/328/330 and S373 in Cx43MAPKmut mice. In mitochondria from Cx43PKCmut mice, the Cx43 phosphorylation at S325/328/330 was reduced, but it was increased at S262, S365, and S373. In SSM isolated from Cx43CK1mut mice, S262 was increasingly phosphorylated compared to WT SSM. Original Western blots and single values of the data presented in Table 1 are shown in the online supplement (Supplementary Figs. 4 –11).
Since the relative phosphorylation of Cx43 and also the absolute amount of the phosphorylated protein may be important, Western blot data of phosphorylated Cx43 were also normalized to the housekeeping proteins GAPDH (LV proteins) or MnSOD (SSM proteins, Table 2). Due to the low amount of Cx43 in the Cx43PKCmut mitochondria, the amount of Cx43 phosphorylated at S262 and S365 was reduced when normalized to MnSOD, whereas it was increased when normalized to total Cx43. The enhanced phosphorylation of S262 in Cx43PKCmut and Cx43CK1mut as seen by normalization to total Cx43 was not detected by normalization to GAPDH or MnSOD.
To investigate whether or not mutations at MAPK, PKC, and CK1 target sites within Cx43 influence mitochondrial function, respiration, ROS formation and MPTP opening were measured in SSM isolated from WT, Cx43MAPKmut (Fig. 1), Cx43PKCmut (Fig. 2), and Cx43CK1mut mice (Fig. 3). Whereas basal respiration (both complex 1 und 2 substrates) was not affected by mutating Cx43 phosphorylation sites, ADP-stimulated oxygen consumption of SSM respiring on complex 1 substrates was decreased in SSM from the three analyzed mouse strains, with the highest reduction in SSM of Cx43CK1mut mice. Using complex 2 substrates, ADP-stimulated respiration was diminished in Cx43CK1mut SSM only. Despite a trend towards a reduced respiratory control rate (RCR; ADP-stimulated respiration/basal respiration) in Cx43CK1mut SSM, there was no significant difference in the RCR between SSM isolated from WT mice and mice with mutated Cx43 phosphorylation sites. There was a trend towards a reduced oxygen consumption driven by N,N,N′,N′-tetramethyl-p-phenylenediamine (TMPD)/ascorbate (complex 4 respiration) and a significantly decreased uncoupled respiration using carbonyl cyanide 4-(trifluoromethoxy)phenylhydrazone (FCCP) in Cx43CK1mut compared to WT SSM (Table 3). The protein amounts of complexes 1–5 of the electron transport chain were not altered in Cx43MAPKmut and Cx43PKCmut, whereas the level of complex 1 was significantly enhanced in Cx43CK1mut mice (Supplementary Fig. 12). ROS formation was similar between SSM isolated from WT, Cx43MAPKmut, Cx43PKCmut, and Cx43CK1mut mice. In addition, the analysis of calcium-induced MPTP opening revealed no differences between the analyzed mouse lines. The use of cyclosporine A (CsA) as an inhibitor of MPTP opening significantly enhanced the calcium retention capacity in all analyzed strains.

Mitochondrial function in Cx43MAPKmut mice. a Mitochondrial oxygen consumption of SSM isolated from wild-type (WT) and Cx43MAPKmut mice respiring on complex 1 or complex 2 substrates without (basal) and with ADP to stimulate respiration. WT: n = 9, Cx43MAPKmut n = 9, *: p < 0.05 vs ADP WT as analyzed by two-way ANOVA. b Original traces showing ROS formation detected by Amplex UltraRed fluorescence in arbitrary units (a.u.) from WT and Cx43MAPKmut SSM. c Quantification of ROS formation (slope of Amplex UltraRed fluorescence in a.u./min) from WT (n = 5) and Cx43MAPKmut SSM (n = 5), p = ns as analyzed by unpaired t test. d Original traces showing Calcium green 5 N fluorescence (in a.u.) of SSM isolated from WT and Cx43MAPKmut mice. Every third minute, 5 µmol/L CaCl2 was added until a sudden increase in Calcium green 5 N fluorescence occurred (↓) which reflects MPTP opening. Measurements were performed without (con) and with Cyclosporine A (CsA), which was used as a positive control to inhibit MPTP opening. e The number of calcium pulses which was necessary to induce MPTP opening was calculated from SSM of WT and Cx43MAPKmut mice without (con) and with CsA. WT (n = 7), Cx43MAPKmut (n = 7). Data were compared by two-way ANOVA

Mitochondrial function in Cx43PKCmut mice. a Mitochondrial oxygen consumption of SSM isolated from wild-type (WT) and Cx43PKCmut mice respiring on complex 1 or complex 2 substrates without (basal) and with ADP to stimulate respiration. WT: n = 11, Cx43PKCmut n = 11, *p < 0.05 vs ADP WT as analyzed by two-way ANOVA. b Original traces showing ROS formation detected by Amplex UltraRed fluorescence in arbitrary units (a.u.) from WT and Cx43PKCmut SSM. c Quantification of ROS formation (slope of Amplex UltraRed fluorescence in a.u./min) from WT (n = 7) and Cx43PKCmut SSM (n = 7), p = ns, data were compared by unpaired t-test. d Original traces showing Calcium green 5 N fluorescence (in a.u.) of SSM isolated from WT and Cx43PKCmut mice. Every third minute, 5 µmol/L CaCl2 was added until a sudden increase in Calcium green 5 N fluorescence occurred (↓) which reflects MPTP opening. Measurements were performed without (con) and with Cyclosporine A (CsA), which was used as a positive control to inhibit MPTP opening. e The number of calcium pulses which was necessary to induce MPTP opening was calculated from SSM of WT and Cx43PKCmut mice without (con) and with CsA. WT (n = 7), Cx43PKCmut (n = 7). Data were compared by two-way ANOVA

Mitochondrial function in Cx43CK1mut mice. a Mitochondrial oxygen consumption of SSM isolated from wild-type (WT) and Cx43CK1mut mice respiring on complex 1 or complex 2 substrates without (basal) and with ADP to stimulate respiration. WT: n = 6, Cx43CK1mut n = 6, *p < 0.05 vs ADP WT as analyzed by two-way ANOVA. b Original traces showing ROS formation detected by Amplex UltraRed fluorescence in arbitrary units (a.u.) from WT and Cx43CK1mut SSM. c Quantification of ROS formation (slope of Amplex UltraRed fluorescence in a.u./min) from WT (n = 8) and Cx43CK1mut SSM (n = 8), p = ns, data were compared by unpaired t test. d Original traces showing calcium green 5 N fluorescence (in a.u.) of SSM isolated from WT and Cx43CK1mut mice. Every third minute, 5 µmol/L CaCl2 was added until a sudden increase in Calcium green 5 N fluorescence occurred (↓) which reflects MPTP opening. Measurements were performed without (con) and with Cyclosporine A (CsA), which was used as a positive control to inhibit MPTP opening. e The number of calcium pulses which was necessary to induce MPTP opening was calculated from SSM of WT and Cx43CK1mut mice without (con) and with CsA. WT (con: n = 6; CsA: n = 5) and Cx43CK1mut (n = 6). Data were compared by two-way ANOVA
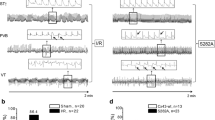
Cx43 hemichannel activity was compared between WT, Cx43MAPKmut, Cx43PKCmut, and Cx43CK1mut mice using patch-clamp experiments. LV cardiomyocytes were voltage-clamped at the resting potential of − 70 mV. Short caffeine applications were used to trigger Cx43 hemichannel activity, which is characterized by spiking unitary events superimposed on the macroscopic sodium/calcium exchanger (NCX) current [42] (Fig. 4a). Altering Cx43 phosphorylation on the MAPK, PKC, and CK1 target sites significantly reduced the amplitude and slowed down the kinetics of the NCX current compared to control (Fig. 4b, c). However, the changes in NCX transporter kinetics did not reflect an alteration of the subsarcolemmal calcium content (Fig. 4d), for which NCX is considered to be a valuable indicator [79]. Cx43 hemichannel activity was significantly reduced in Cx43CK1mut and in Cx43PKCmut mice, but not in Cx43MAPKmut mice (Fig. 4e).

Effect of altered Cx43 phosphorylation on hemichannel gating. a Representative sodium/calcium exchanger (NCX) current traces with superimposed Cx43 unitary current events triggered by caffeine (10 mmol/L) at Vh = − 70 mV. b Average data showing significantly reduced normalized NCX peak amplitudes in Cx43MAPKmut, Cx43PKCmut, and Cx43CK1mut cardiomyocytes compared to wild-type (WT) cells. NCX currents were normalized on the cell capacity to minimize the difference due to cell size. c Summary data illustrating the time constant of the recovery phase in cardiomyocytes of WT, Cx43MAPKmut, Cx43PKCmut, and Cx43CK1mut mice. d Overall sarcoplasmic reticulum calcium equilibrium. e Quantification of the Cx43 hemichannel open probability demonstrating reduced single-channel activity in Cx43PKCmut and Cx43CK1mut mice compared to control conditions. WT (N = 6; n = 31), (Cx43MAPKmut (N = 9; n = 26), Cx43PKCmut (N = 3; n = 19), Cx43CK1mut (N = 4; n = 20),). *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001 (unpaired Student’s t test)
To characterize the importance of Cx43 phosphorylation for myocardial I/R injury, isolated hearts were Langendorff-perfused and underwent a protocol of I/R (45 min ischemia and 120 min reperfusion) without and with IPC. Heart rate returned to 600 bpm in all groups within 10 min reperfusion and hemodynamic data are shown in Table 4. Hearts of WT, Cx43MAPKmut, Cx43PKCmut, and Cx43CK1mut mice revealed similar infarct sizes after I/R (Fig. 5). Following IPC, infarct size was reduced in WT, Cx43MAPKmut and Cx43PKCmut hearts, whereas IPC did not protect hearts of Cx43CK1mut mice. In Cx43Cre−ER(T)/fl hearts, which express 50% of Cx43 (an amount comparable to that in Cx43CK1mut mice), infarct size after I/R with IPC was 77.8 ± 7.4% of the area at risk (n = 4, p = ns vs. WT), thereby indicating a loss of cardioprotection by IPC also in this strain.

Influence of Cx43 phosphorylation on the cardioprotection by IPC in vitro. Infarct sizes (in % of the area at risk) are shown for wild-type (WT), Cx43MAPKmut, Cx43PKCmut, and Cx43CK1mut mice subjected to ischemia/reperfusion (I/R) or ischemic preconditioning (IPC) in vitro. In addition to the individual infarct sizes of each animal, the mean value ± SEM of each group is presented, WT (I/R: n = 9, IPC: n = 8), Cx43MAPKmut (I/R: n = 6, IPC; n = 6), Cx43PKCmut (I/R: n = 6, IPC; n = 6), Cx43CK1mut (I/R: n = 6, IPC; n = 6), data were compared by two-way ANOVA
Discussion
The present study demonstrates that the specific phosphorylation of Cx43 at MAPK, PKC, or CK1 target sites influenced the phosphorylation and the amount of Cx43 in LV and mitochondrial protein extracts. The respective mutations in the three mouse lines altered mitochondrial oxygen consumption without effects on ROS formation and MPTP opening. Hemichannel activity was reduced in cardiomyocytes of Cx43PKCmut and Cx43CK1mut mice, whereas the cardioprotection by IPC was abrogated in Cx43CK1mut mice only.
At first, we analyzed the expression and phosphorylation of Cx43 in total LV proteins extracted from Cx43MAPKmut, Cx43PKCmut, and Cx43CK1mut mice. Compared to WT proteins, we detected a moderate decrease in total Cx43 in Cx43MAPKmut mice. The analysis of the amount of Cx43 in this strain in vascular smooth muscle cells did not demonstrate reduced levels of Cx43, which may be due to the different tissue/cell types analyzed [34]. In Cx43PKCmut and Cx43CK1mut mice, we found a similar decrease of Cx43 in mouse hearts as previously published [30, 54, 70]. Whereas others suggested that the decreased Cx43 protein levels in Cx43CK1mut mice are due to posttranscriptional alterations [70], the reason for the reduced Cx43 protein amount in Cx43MAPKmut, Cx43PKCmut, and Cx43CK1mut mice remains unclear. Pharmacological inhibition of PKC or CK1 did not reduce Cx43 expression as in Cx43PKCmut and Cx43CK1mut mice, indicating that those interventions are not able to mimic the complex situation in vivo, where reduced gap junctional stability and Cx43 down-regulation are observed [54, 67, 70]. A failure to reduce Cx43 phosphorylation and amount of the whole-cell protein under baseline conditions using PKC and CK1 inhibitors has already been shown in different cell types, including cardiomyocytes [13, 15, 58, 69, 78, 82].
We addressed the question whether the mutation of phosphorylation sites within Cx43 influences the phosphorylation status at other serine residues. The analyses of LV total proteins of Cx43PKCmut and Cx43CK1mut mice demonstrated alterations of Cx43 phosphorylation at all residues analyzed, whereas such distinct phosphorylation was not observed in Cx43MAPKmut mice. Our data confirm a previous study showing enhanced phosphorylation of Cx43 at S262 in Cx43CK1mut mice [70]. Therefore, mutations of S325/328/330 or S368 are sufficient to induce significant changes in Cx43 phosphorylation at multiple residues including both enhanced and reduced phosphorylation at other amino acids. Possibly, the phosphorylation status of Cx43 has an impact on the binding of kinases or phosphatases and may thereby effect the phosphorylation of other target proteins [70].
In Cx43MAPKmut and Cx43CK1mut mice, we found comparable reductions of Cx43 in total and in mitochondrial protein extracts, whereas in Cx43PKCmut mice, Cx43 was further reduced. Thus, the phosphorylation of Cx43 at S368 is important for the mitochondrial amount of Cx43. Yet, the mechanisms that prevent a mitochondrial translocation of Cx43 not phosphorylated at S368 remain unclear.
Previous studies indicated a phosphorylation of mitochondrial Cx43 at S262 [73] and S368 [64, 73] and these findings are confirmed in the present analysis. Additionally, we detected a phosphorylation of mitochondrial Cx43 at S325/328/330, S365, and S373, which demonstrates that under physiological conditions, the mitochondrial fraction of the protein is phosphorylated at multiple residues.
The pattern of Cx43 phosphorylation differed between total LV and mitochondrial proteins in Cx43MAPKmut, Cx43PKCmut, and Cx43CK1mut mice. We cannot judge from the present data if only Cx43 with a specific phosphorylation pattern is imported into the mitochondria, or if the phosphorylation status of the protein is achieved within the organelle. Since several kinases and phosphatases have been detected in mitochondria [41], the latter is also possible.
We addressed the question whether the mutation of Cx43 phosphorylation sites alters mitochondrial function. Compared to wild-type mitochondria, mitochondria from Cx43MAPKmut and Cx43PKCmut mice revealed a moderate, but significant reduction of ADP-stimulated oxygen consumption when respiring on complex 1 substrates, whereas mitochondria from Cx43CK1mut showed decreased oxygen consumption when respiring on complex 1 and complex 2 substrates. Also, the uncoupled respiration in the presence of FCCP, which indicates the maximum rate of respiration that can be achieved within the mitochondrial preparation, was diminished in Cx43CK1mut mitochondria and there was also a trend towards a reduced TMPD/ascorbate-stimulated (complex 4) respiration. Therefore, the mutation of CK1 phosphorylation sites within Cx43 generally impairs mitochondrial oxygen consumption. We already showed a specific decrease of complex 1-mediated respiration in mitochondria isolated from inducible Cx43-knockout mice [7], which contain about 10% Cx43, and hypothesized that the mitochondrial amount of Cx43 is important for mitochondrial function. Interestingly, in the present study, we detected similar reductions of oxygen consumption in mitochondria isolated from Cx43MAPKmut and Cx43PKCmut mice containing either 73% or 9% Cx43, whereas the highest limitation in complex 1-mediated respiration was found in mitochondria from Cx43CK1mut mice containing about 62% Cx43. Therefore, it is unlikely that the influence of Cx43 on mitochondrial respiration is caused by a reduction in the amount of the protein only; rather, the phosphorylation status of Cx43 is involved in the regulation of mitochondrial oxygen consumption. Since Cx43 phosphorylation at S262 is absent in Cx43MAPKmut mice and the phosphorylation at this residue is increased in mitochondria of Cx43PKCmut and Cx43CK1mut mice, the importance of Cx43 S262 for respiration can presumably be neglected. The confinement of respiration correlated with the phosphorylation status at S325/328/330, which was higher in Cx43MAPKmut than in Cx43PKCmut and absent in Cx43CK1mut mitochondria. The finding that the amount of complex 1 of the electron transport chain is enhanced in Cx43CK1mut mice suggests that the efficiency of the electron transport chain is decreased in this mouse strain.
ROS formation is decreased after inhibition of Cx43 by carbenoxolone [68] or small interfering RNAs [77]. Mitochondrial Cx43 is involved in the regulation of MPTP (which occurs at the onset of reperfusion, leads to mitochondrial swelling, and finally cell death) as shown by an accelerated calcium-induced MPTP opening in cardiac mitochondria after inhibition of Cx43 with the mimetic peptide Gap27 or 18 alpha-glycyrrhetinic acid [8, 73]. In contrast, increased mitochondrial calcium retention capacity was measured in cardiac mitochondria upon Cx43 inhibition; however, these experiments were performed while blocking the mitochondrial calcium uniporter [20]. In the present study, ROS formation and calcium-induced MPTP opening were similar in WT, Cx43MAPKmut, Cx43PKCmut, and Cx43CK1mut mitochondria. Taking into account that the mitochondrial amount of Cx43 significantly differs between the three mouse lines without having consequences for ROS formation and MPTP opening, the present study again emphasizes that not only the amount of Cx43 but also posttranslational modifications are important for mitochondrial function.
The role of Cx43 phosphorylation in cardiac I/R injury has been addressed in several studies. Whereas an alteration of the phosphorylation status of Cx43 is observed with ischemia, the specific phosphorylation of Cx43 on S325/328/330 [2, 37], S365 [2, 71], S368 [17, 76], and S373 [2, 14] depends on the model and the duration of ischemia. However, several studies demonstrate that ischemia is associated with dephosphorylation of Cx43, a lateralization of the protein from the gap junctions and a subsequent internalization of the protein [60]. As a consequence of I/R, the interactions between Cx43 and its binding partners are altered [44], an effect potentially contributing to decreased electrical and chemical gap junctional coupling during ischemia [46]. Yet, the present study did not directly investigate the effect of Cx43 phosphorylation on gap junctional coupling in myocardial I/R injury. An opening of Cx43-formed hemichannels is known to contribute to I/R injury [65, 81]. Hemichannel opening is regulated via Cx43 phosphorylation, e.g., the mutation of the PKC site S368 to alanine reduces hemichannel opening (for review see [52]) and astrocytes of Cx43MAPKmut mice display diminished hemichannel activity [19]. Recently, it was shown that in a model of Duchenne muscular dystrophy the mutation of S325/328/330 to phospho-mimicking glutamic acids increased hemichannel opening [28]. However, the replacement of S325/328/330 to alanine or aspartate residues both enhances hemichannel opening [18]. Here, we describe decreased hemichannel activity in Cx43PKCmut and Cx43CK1mut, but not in Cx43MAPKmut cardiomyocytes. Therefore, we confirm previous data on the role of PKC-mediated Cx43 phosphorylation for hemichannel opening, whereas the contribution of MAPK and CK1 signaling on Cx43 hemichannel activity seems to be dependent on the model/cell type analyzed. It is interesting to note, that although probability of hemichannel opening was partially reduced, the extent of I/R injury was similar between the different mouse strains.
It is likely that Cx43 phosphorylation is involved in cardioprotection, since pharmacological strategies which reduce myocardial damage following I/R are often associated with preserved Cx43 phosphorylation [60]. Moreover, the mutation of S368 to alanine abolishes the cardioprotection by sphingosine-1-phosphate [48]. However, the similar infarct sizes following I/R we observed in all strains analyzed argue against an influence of Cx43 phosphorylation at MAPK, PKC, or CK1 target sites on myocardial I/R injury per se. Similar infarct sizes in WT and Cx43CK1mut mice have already been demonstrated after myocardial I/R in animals less than 1 year old [70]. However, it must be considered that in the analyzed mouse strains—especially in Cx43PKCmut mice—not only the phosphorylation of Cx43 but also the total amount of the protein is reduced. Previous studies in mice with 50% Cx43 demonstrate decreased myocardial infarction following permanent coronary ligation [36], but no influence of the reduced protein amount on infarct size after acute I/R [59, 62].
Data showing that ischemia induces dephosphorylation of Cx43 lead to the hypothesis that the cardioprotection by IPC may prevent such dephosphorylation, and indeed, IPC enhances Cx43 phosphorylation (S262 and S368) both in vitro and in vivo [56, 61, 72]. It is suggested that the cardioprotection by IPC is mediated at least in part via suppressing chemical gap junctional communication [46, 60], but also mitochondrial Cx43 seems to play a role [40, 55]. A reduction of Cx43 to about 50% in constitutive knockout mice is sufficient to abrogate IPC’s cardioprotection in vivo [62] and in vitro (present study with Cx43Cre−ER(T)/fl mice); however, others demonstrate that IPC reduces infarct size in vitro in inducible Cx43 knockout mice expressing half the level of Cx43 [59]. In the present study, we showed that Cx43PKCmut mice expressing around 40% of normal Cx43 levels were effectively protected by IPC in vitro, whereas in Cx43CK1mut mice with 54%, the level of Cx43 of the cardioprotection by IPC was lost. These data argue that Cx43 expression levels alone cannot explain IPC effects and indicate the importance of posttranslational modifications. Our data indicate that the phosphorylation of Cx43 at MAPK and PKC-target sites is not essential for the cardioprotection by IPC, but highlight the role of CK1-targeted residues for the reduction of myocardial I/R damage. Moreover, the loss of infarct size reduction by IPC in Cx43CK1mut mice correlated with the highest S262 phosphorylation of mitochondrial Cx43. Whereas opening of Cx43-formed hemichannels during ischemia is known to contribute to myocardial I/R damage, our finding of reduced hemichannel activity in Cx43CK1mut mice makes it unlikely that hemichannel opening is involved in the loss of cardioprotection by IPC in these mice. Cardiomyocytes of Cx43CK1mut mice—similar to Cx43MAPKmut and Cx43PKCmut mice—displayed reduced NCX currents, suggesting that targeting phosphorylation on the Cx43 C-terminal residues can control channel gating and affects the kinetic of the NCX, one of the regulators of intracellular calcium homeostasis. Ca2+ entry via reverse mode NCX contributes to myocardial I/R injury [31]. Since our data demonstrated reduced NCX currents in all three mouse lines with mutated Cx43 phosphorylation sites, NCX-mediated currents are unlikely to explain the loss of cardioprotection specifically in Cx43CK1mut mice.
Mitochondria are important targets in the transduction of myocardial conditioning [6]. Reduction of mitochondrial Cx43 (without affecting gap junctional Cx43) abrogates pharmacological preconditioning [55], whereas the overexpression of mitochondrial Cx43 induces cytoprotection [43], pointing to an important role of the amount of mitochondrial Cx43 in cardioprotection. However, such direct correlation between the level of mitochondrial Cx43 and cardioprotection by IPC is not supported by the present data. Heterozygous Cx43-deficient mice have a specific deficit in ROS formation in response to diazoxide, and accordingly, the cardioprotection by pharmacological preconditioning with diazoxide is lost [24]. Yet, the present study demonstrates that neither the reduction of mitochondrial Cx43 by more than 50% nor the mutation of Cx43 phosphorylation sites modified ROS formation per se. In line with these findings, myocardial infarct size following I/R was similar among all mice strains. MPTP opening is regulated by Cx43, since pharmacological inhibition of Cx43 decreases the calcium retention capacity and thereby accelerates MPTP opening [8, 73]. In I/R injury, MPTP opening at reperfusion contributes to irreversible myocardial damage; a similar MPTP opening was found in Cx43MAPKmut, Cx43PKCmut, and Cx43CK1mut mitochondria which went along with an unaltered infarct size following I/R but also argues against its importance for the attenuation of cardioprotection by IPC in Cx43CK1mut mice. With reperfusion after prolonged ischemia, the combination of still elevated calcium concentrations and restored ATP levels via oxidative phosphorylation leads to cardiomyocyte hypercontracture and finally to cell death [21]. The decreased oxygen consumption measured in Cx43CK1mut mitochondria might possibly limit the generation of ATP at reperfusion and may thereby be beneficial; however, similar infarct sizes in WT and Cx43CK1mut hearts do not favor an involvement of CK1-mediated Cx43 phosphorylation in cardiomyocyte hypercontracture. Several studies show that in the cardioprotection by IPC preserved mitochondrial energetics by maintaining mitochondrial oxygen consumption limits cell death after myocardial I/R injury [6]. In the present study, impaired respiration was measured especially in Cx43CK1mut mitochondria. Whereas this finding suggests that limited oxygen consumption in Cx43CK1mut mitochondria may be important for the loss of infarct size reduction by IPC in Cx43CK1mut mice in vitro, the exact contribution of mitochondrial Cx43 (total amount and phosphorylated Cx43) towards the cardioprotection by IPC remains unclear.
Study limitations—1. In the present study, we used genetically modified mice in which Cx43 phosphorylation sites are constitutively altered. These mice may carry some unrecognized additional mutations, which may also influence cellular and/or mitochondrial function.
2. Myocardial infarction after I/R injury may differ between isolated buffer-perfused hearts—as performed in the present study—and the in vivo situation. Therefore, we cannot exclude that Cx43MAPKmut, Cx43PKCmut, and Cx43CK1mut mice will show a different response towards a preconditioning stimulus in the in vivo situation. Thus, the evaluation of infarct size without and with IPC in vivo will represent an important extension of the present study.
3. In addition, it has to be noted that mitochondrial function and electrophysiological measurements were performed at 25 °C or room temperature. Whereas mitochondrial oxygen consumption is generally lower at 25 °C compared to 37°, the temperature sensitivity of mitochondrial respiration is dependent on the type of substrates and is lowest for glutamate and malate [38], the complex 1 substrates used in the present study. While data show that Cx26-formed hemichannels are sensitive to changes in the temperature [74, 80], Cx43 hemichannel gating mechanisms are not affected by temperature [51, 57], and therefore, the majority of studies investigating Cx43 hemichannels were commonly performed at room temperature [3, 33, 35, 45, 53, 60]. Importantly, since experiments on mitochondrial function and electrophysiological recordings were carried out at the same temperatures, differences in respiration and hemichannel activity between the mouse strains cannot be attributed to temperature variances.
Taken together, our data show that the mutation of Cx43 phosphorylation sites targeted by MAPK, PKC, or CK1 to non-phosphorylatable residues affects the amount and the phosphorylation of Cx43 at amino acids not directly targeted by the mutations and thereby emphasizes the complex interplay between posttranslational Cx43 modifications and the amount of the Cx43 protein. The alterations of the amounts/phosphorylation of Cx43 have consequences for specific cellular function, since mutation of Cx43 phosphorylation sites for MAPK, PKC, and CK1 sites reduces mitochondrial respiration, mutation of PKC and CK1 sites decreases hemichannel activity, and the mutation of Cx43 residues targeted by CK1 abrogates the infarct size reduction by IPC (Fig. 6). Therefore, in addition to Cx43 phosphorylation at S262 and S368, phosphorylation at S325/328/330 is involved in mediating IPC’s cardioprotection, an effect independent from the protein amount of Cx43 and Cx43-mediated hemichannel opening.

The role of Cx43 phosphorylation for specific cellular functions. The scheme shows the topology of Cx43 including the transmembrane domains (1–4) and important phosphorylation sites, which are indicated by filled circles and are numbered according to their positions. ↓: decreased/impaired function, ✓: normal function
Data availability
Not applicable.
Code availability
Not applicable.
Abbreviations
- AKT:
-
Protein kinase B
- CamKII:
-
Ca2+/calmodulin kinase II
- CK1:
-
Casein kinase 1
- Cx43:
-
Connexin 43
- CsA:
-
Cyclosporine A
- EDP:
-
End-diastolic pressure
- ERK:
-
Extracellular-signal related kinase
- GAPDH:
-
Glyceraldehyde 3-phosphate dehydrogenase
- IFM:
-
Interfibrillar mitochondria
- IPC:
-
Ischemic preconditioning
- I/R:
-
Ischemia/reperfusion
- LV:
-
Left ventricle
- LVDP:
-
Left-ventricular developed pressure
- NCX:
-
Sodium/calcium exchanger
- MAPK:
-
Mitogen-activated protein kinase
- MnSOD:
-
Manganese superoxide dismutase
- MPTP:
-
Mitochondrial permeability transition pore
- p34cdc2:
-
P34cdc2/cyclin B kinase
- PKA:
-
Protein kinase A
- PKC:
-
Protein kinase C
- ROS:
-
Reactive oxygen species
- S:
-
Serine
- SSM:
-
Subsarcolemmal mitochondria
- Src:
-
Pp60src kinase
- TTC:
-
Triphenyltetrazolium chloride
- WT:
-
Wild type
- ZO-1:
-
Zonula occludens 1
References
Andelova K, Egan Benova T, Szeiffova Bacova B, Sykora M, Prado NJ, Diez ER, Hlivak P, Tribulova N (2020) Cardiac connexin-43 hemichannels and pannexin1 channels: provocative antiarrhythmic targets. Int J Mol Sci 22. https://doi.org/10.3390/ijms22010260
Axelsen LN, Stahlhut M, Mohammed S, Larsen BD, Nielsen MS, Holstein-Rathlou NH, Andersen S, Jensen ON, Hennan JK, Kjolbye AL (2006) Identification of ischemia-regulated phosphorylation sites in connexin43: a possible target for the antiarrhythmic peptide analogue rotigaptide (ZP123). J Mol Cell Cardiol 40:790–798. https://doi.org/10.1016/j.yjmcc.2006.03.005
Bao X, Reuss L, Altenberg GA (2004) Regulation of purified and reconstituted connexin 43 hemichannels by protein kinase C-mediated phosphorylation of Serine 368. J Biol Chem 279:20058–20066. https://doi.org/10.1074/jbc.M311137200
Boengler K, Bulic M, Schreckenberg R, Schluter KD, Schulz R (2017) The gap junction modifier ZP1609 decreases cardiomyocyte hypercontracture following ischaemia/reperfusion independent from mitochondrial connexin 43. Br J Pharmacol 174:2060–2073. https://doi.org/10.1111/bph.13804
Boengler K, Heusch G, Schulz R (2011) Nuclear-encoded mitochondrial proteins and their role in cardioprotection. Biochim Biophys Acta 1813:1286–1294. https://doi.org/10.1016/j.bbamcr.2011.01.009
Boengler K, Lochnit G, Schulz R (2018) Mitochondria “THE” target of myocardial conditioning. Am J Physiol Heart Circ Physiol 315:H1215–H1231. https://doi.org/10.1152/ajpheart.00124.2018
Boengler K, Ruiz-Meana M, Gent S, Ungefug E, Soetkamp D, Miro-Casas E, Cabestrero A, Fernandez-Sanz C, Semenzato M, Lisa FD, Rohrbach S, Garcia-Dorado D, Heusch G, Schulz R, Mercola M (2012) Mitochondrial connexin 43 impacts on respiratory complex I activity and mitochondrial oxygen consumption. J Cell Mol Med 16:1649–1655. https://doi.org/10.1111/j.1582-4934.2011.01516.x
Boengler K, Schulz R (2017) Connexin 43 and mitochondria in cardiovascular health and disease. Adv Exp Med Biol 982:227–246. https://doi.org/10.1007/978-3-319-55330-6_12
Boengler K, Stahlhofen S, van de Sand A, Gres P, Ruiz-Meana M, Garcia-Dorado D, Heusch G, Schulz R (2009) Presence of connexin 43 in subsarcolemmal, but not in interfibrillar cardiomyocyte mitochondria. Basic Res Cardiol 104:141–147. https://doi.org/10.1007/s00395-009-0007-5
Boengler K, Ungefug E, Heusch G, Leybaert L, Schulz R (2013) Connexin 43 impacts on mitochondrial potassium uptake. Front Pharmacol 4:73. https://doi.org/10.3389/fphar.2013.00073
Botker HE, Hausenloy D, Andreadou I, Antonucci S, Boengler K, Davidson SM, Deshwal S, Devaux Y, Di Lisa F, Di Sante M, Efentakis P, Femmino S, Garcia-Dorado D, Giricz Z, Ibanez B, Iliodromitis E, Kaludercic N, Kleinbongard P, Neuhauser M, Ovize M, Pagliaro P, Rahbek-Schmidt M, Ruiz-Meana M, Schluter KD, Schulz R, Skyschally A, Wilder C, Yellon DM, Ferdinandy P, Heusch G (2018) Practical guidelines for rigor and reproducibility in preclinical and clinical studies on cardioprotection. Basic Res Cardiol 113:39. https://doi.org/10.1007/s00395-018-0696-8
Cadenas S (2018) ROS and redox signaling in myocardial ischemia-reperfusion injury and cardioprotection. Free Radic Biol Med 117:76–89. https://doi.org/10.1016/j.freeradbiomed.2018.01.024
Cooper CD, Lampe PD (2002) Casein kinase 1 regulates connexin-43 gap junction assembly. J Biol Chem 277:44962–44968. https://doi.org/10.1074/jbc.M209427200
Dunn CA, Lampe PD (2014) Injury-triggered Akt phosphorylation of Cx43: a ZO-1-driven molecular switch that regulates gap junction size. J Cell Sci 127:455–464. https://doi.org/10.1242/jcs.142497
Duquesnes N, Derangeon M, Métrich M, Lucas A, Mateo P, Li L, Morel E, Lezoualc’h F, Crozatier B (2010) Epac stimulation induces rapid increases in connexin43 phosphorylation and function without preconditioning effect. Pflugers Arch 460:731–741. https://doi.org/10.1007/s00424-010-0854-9
Eckardt D, Theis M, Degen J, Ott T, van Rijen HV, Kirchhoff S, Kim JS, de Bakker JM, Willecke K (2004) Functional role of connexin43 gap junction channels in adult mouse heart assessed by inducible gene deletion. J Mol Cell Cardiol 36:101–110. https://doi.org/10.1016/j.yjmcc.2003.10.006
Ek-Vitorin JF, King TJ, Heyman NS, Lampe PD, Burt JM (2006) Selectivity of connexin 43 channels is regulated through protein kinase C-dependent phosphorylation. Circ Res 98:1498–1505. https://doi.org/10.1161/01.RES.0000227572.45891.2c
Ek-Vitorin JF, Pontifex TK, Burt JM (2018) Cx43 channel gating and permeation: multiple phosphorylation-dependent roles of the carboxyl terminus. Int J Mol Sci 19:1659. https://doi.org/10.3390/ijms19061659
Freitas-Andrade M, Wang N, Bechberger JF, De Bock M, Lampe PD, Leybaert L, Naus CC (2019) Targeting MAPK phosphorylation of Connexin43 provides neuroprotection in stroke. J Exp Med 216:916–935. https://doi.org/10.1084/jem.20171452
Gadicherla AK, Wang N, Bulic M, Agullo-Pascual E, Lissoni A, De Smet M, Delmar M, Bultynck G, Krysko DV, Camara A, Schluter KD, Schulz R, Kwok WM, Leybaert L (2017) Mitochondrial Cx43 hemichannels contribute to mitochondrial calcium entry and cell death in the heart. Basic Res Cardiol 112:27. https://doi.org/10.1007/s00395-017-0618-1
Garcia-Dorado D, Ruiz-Meana M, Inserte J, Rodriguez-Sinovas A, Piper HM (2012) Calcium-mediated cell death during myocardial reperfusion. Cardiovasc Res 94:168–180. https://doi.org/10.1093/cvr/cvs116
Goodenough DA, Paul DL (2003) Beyond the gap: functions of unpaired connexon channels. Nat Rev Mol Cell Biol 4:285–294. https://doi.org/10.1038/nrm1072
He H, Li N, Zhao Z, Han F, Wang X, Zeng Y (2015) Ischemic postconditioning improves the expression of cellular membrane connexin 43 and attenuates the reperfusion injury in rat acute myocardial infarction. Biomed Rep 3:668–674. https://doi.org/10.3892/br.2015.485
Heinzel FR, Luo Y, Li X, Boengler K, Buechert A, Garcia-Dorado D, Di Lisa F, Schulz R, Heusch G (2005) Impairment of diazoxide-induced formation of reactive oxygen species and loss of cardioprotection in connexin 43 deficient mice. Circ Res 97:583–586. https://doi.org/10.1161/01.RES.0000181171.65293.65
Heusch G (2015) Molecular basis of cardioprotection: signal transduction in ischemic pre-, post-, and remote conditioning. Circ Res 116:674–699. https://doi.org/10.1161/circresaha.116.305348
Heusch G (2020) Myocardial ischaemia-reperfusion injury and cardioprotection in perspective. Nat Rev Cardiol 17:773–789. https://doi.org/10.1038/s41569-020-0403-y
Heusch G, Buchert A, Feldhaus S, Schulz R (2006) No loss of cardioprotection by postconditioning in connexin 43-deficient mice. Basic Res Cardiol 101:354–356. https://doi.org/10.1007/s00395-006-0589-0
Himelman E, Lillo MA, Nouet J, Gonzalez JP, Zhao Q, Xie LH, Li H, Liu T, Wehrens XH, Lampe PD, Fishman GI, Shirokova N, Contreras JE, Fraidenraich D (2020) Prevention of connexin-43 remodeling protects against Duchenne muscular dystrophy cardiomyopathy. J Clin Invest 130:1713–1727. https://doi.org/10.1172/jci128190
Hood AR, Ai X, Pogwizd SM (2017) Regulation of cardiac gap junctions by protein phosphatases. J Mol Cell Cardiol 107:52–57. https://doi.org/10.1016/j.yjmcc.2017.05.002
Huang GY, Xie LJ, Linask KL, Zhang C, Zhao XQ, Yang Y, Zhou GM, Wu YJ, Marquez-Rosado L, McElhinney DB, Goldmuntz E, Liu C, Lampe PD, Chatterjee B, Lo CW (2011) Evaluating the role of connexin43 in congenital heart disease: Screening for mutations in patients with outflow tract anomalies and the analysis of knock-in mouse models. J Cardiovasc Dis Res 2:206–212. https://doi.org/10.4103/0975-3583.89804
Imahashi K, Pott C, Goldhaber JI, Steenbergen C, Philipson KD, Murphy E (2005) Cardiac-specific ablation of the Na+-Ca2+ exchanger confers protection against ischemia/reperfusion injury. Circ Res 97:916–921. https://doi.org/10.1161/01.RES.0000187456.06162.cb
Jasova M, Kancirova I, Waczulikova I, Ferko M (2017) Mitochondria as a target of cardioprotection in models of preconditioning. J Bioenerg Biomembr 49:357–368. https://doi.org/10.1007/s10863-017-9720-1
John SA, Kondo R, Wang SY, Goldhaber JI, Weiss JN (1999) Connexin-43 hemichannels opened by metabolic inhibition. J Biol Chem 274:236–240. https://doi.org/10.1074/jbc.274.1.236
Johnstone SR, Kroncke BM, Straub AC, Best AK, Dunn CA, Mitchell LA, Peskova Y, Nakamoto RK, Koval M, Lo CW, Lampe PD, Columbus L, Isakson BE (2012) MAPK phosphorylation of connexin 43 promotes binding of cyclin E and smooth muscle cell proliferation. Circ Res 111:201–211. https://doi.org/10.1161/circresaha.112.272302
Kang J, Kang N, Lovatt D, Torres A, Zhao Z, Lin J, Nedergaard M (2008) Connexin 43 hemichannels are permeable to ATP. J Neurosci 28:4702–4711. https://doi.org/10.1523/jneurosci.5048-07.2008
Kanno S, Kovacs A, Yamada KA, Saffitz JE (2003) Connexin43 as a determinant of myocardial infarct size following coronary occlusion in mice. J Am Coll Cardiol 41:681–686. https://doi.org/10.1016/s0735-1097(02)02893-0
Lampe PD, Cooper CD, King TJ, Burt JM (2006) Analysis of Connexin43 phosphorylated at S325, S328 and S330 in normoxic and ischemic heart. J Cell Sci 119:3435–3442. https://doi.org/10.1242/jcs.03089
Lemieux H, Blier PU, Gnaiger E (2017) Remodeling pathway control of mitochondrial respiratory capacity by temperature in mouse heart: electron flow through the Q-junction in permeabilized fibers. Sci Rep 7:2840. https://doi.org/10.1038/s41598-017-02789-8
Li G, Whittaker P, Yao M, Kloner RA, Przyklenk K (2002) The gap junction uncoupler heptanol abrogates infarct size reduction with preconditioning in mouse hearts. Cardiovasc Pathol 11:158–165
Li X, Heinzel FR, Boengler K, Schulz R, Heusch G (2004) Role of connexin 43 in ischemic preconditioning does not involve intercellular communication through gap junctions. J Mol Cell Cardiol 36:161–163. https://doi.org/10.1016/j.yjmcc.2003.10.019
Lim S, Smith KR, Lim ST, Tian R, Lu J, Tan M (2016) Regulation of mitochondrial functions by protein phosphorylation and dephosphorylation. Cell Biosci 6:25. https://doi.org/10.1186/s13578-016-0089-3
Lissoni A, Hulpiau P, Martins-Marques T, Wang N, Bultynck G, Schulz R, Witschas K, Girao H, De Smet M, Leybaert L (2021) RyR2 regulates Cx43 hemichannel intracellular Ca2+-dependent activation in cardiomyocytes. Cardiovasc Res 117:123–136. https://doi.org/10.1093/cvr/cvz340
Lu G, Haider H, Porollo A, Ashraf M (2010) Mitochondria-specific transgenic overexpression of connexin-43 simulates preconditioning-induced cytoprotection of stem cells. Cardiovasc Res 88:277–286. https://doi.org/10.1093/cvr/cvq293
Martins-Marques T, Anjo SI, Pereira P, Manadas B, Girao H (2015) Interacting network of the gap junction (GJ) protein connexin43 (Cx43) is modulated by ischemia and reperfusion in the heart. Mol Cell Proteomics 14:3040–3055. https://doi.org/10.1074/mcp.M115.052894
Meunier C, Wang N, Yi C, Dallerac G, Ezan P, Koulakoff A, Leybaert L, Giaume C (2017) Contribution of astroglial Cx43 hemichannels to the modulation of glutamatergic currents by D-serine in the mouse prefrontal cortex. J Neurosci 37:9064–9075. https://doi.org/10.1523/jneurosci.2204-16.2017
Miura T, Miki T, Yano T (2010) Role of the gap junction in ischemic preconditioning in the heart. Am J Physiol Heart Circ Physiol 298:H1115-1125. https://doi.org/10.1152/ajpheart.00879.2009
Morciano G, Bonora M, Campo G, Aquila G, Rizzo P, Giorgi C, Wieckowski MR, Pinton P (2017) Mechanistic role of mPTP in ischemia-reperfusion injury. Adv Exp Med Biol 982:169–189. https://doi.org/10.1007/978-3-319-55330-6_9
Morel S, Christoffersen C, Axelsen LN, Montecucco F, Rochemont V, Frias MA, Mach F, James RW, Naus CC, Chanson M, Lampe PD, Nielsen MS, Nielsen LB, Kwak BR (2016) Sphingosine-1-phosphate reduces ischaemia-reperfusion injury by phosphorylating the gap junction protein Connexin43. Cardiovasc Res 109:385–396. https://doi.org/10.1093/cvr/cvw004
Ong SB, Subrayan S, Lim SY, Yellon DM, Davidson SM, Hausenloy DJ (2010) Inhibiting mitochondrial fission protects the heart against ischemia/reperfusion injury. Circulation 121:2012–2022. https://doi.org/10.1161/circulationaha.109.906610
Penna C, Perrelli MG, Raimondo S, Tullio F, Merlino A, Moro F, Geuna S, Mancardi D, Pagliaro P (2009) Postconditioning induces an anti-apoptotic effect and preserves mitochondrial integrity in isolated rat hearts. Biochim Biophys Acta 1787:794–801. https://doi.org/10.1016/j.bbabio.2009.03.013
Peracchia C (2004) Chemical gating of gap junction channels; roles of calcium, pH and calmodulin. Biochim Biophys Acta 1662:61–80. https://doi.org/10.1016/j.bbamem.2003.10.020
Pogoda K, Kameritsch P, Retamal MA, Vega JL (2016) Regulation of gap junction channels and hemichannels by phosphorylation and redox changes: a revision. BMC Cell Biol 17(Suppl 1):11. https://doi.org/10.1186/s12860-016-0099-3
Ponsaerts R, De Vuyst E, Retamal M, D’Hondt C, Vermeire D, Wang N, De Smedt H, Zimmermann P, Himpens B, Vereecke J, Leybaert L, Bultynck G (2010) Intramolecular loop/tail interactions are essential for connexin 43-hemichannel activity. FASEB J 24:4378–4395. https://doi.org/10.1096/fj.09-153007
Remo BF, Qu J, Volpicelli FM, Giovannone S, Shin D, Lader J, Liu FY, Zhang J, Lent DS, Morley GE, Fishman GI (2011) Phosphatase-resistant gap junctions inhibit pathological remodeling and prevent arrhythmias. Circ Res 108:1459–1466. https://doi.org/10.1161/circresaha.111.244046
Rodriguez-Sinovas A, Boengler K, Cabestrero A, Gres P, Morente M, Ruiz-Meana M, Konietzka I, Miro E, Totzeck A, Heusch G, Schulz R, Garcia-Dorado D (2006) Translocation of connexin 43 to the inner mitochondrial membrane of cardiomyocytes through the heat shock protein 90-dependent TOM pathway and its importance for cardioprotection. Circ Res 99:93–101. https://doi.org/10.1161/01.RES.0000230315.56904.de
Rong B, Xie F, Sun T, Hao L, Lin MJ, Zhong JQ (2016) Nitric oxide, PKC-epsilon, and connexin43 are crucial for ischemic preconditioning-induced chemical gap junction uncoupling. Oncotarget 7:69243–69255. https://doi.org/10.18632/oncotarget.12087
Sáez JC, Retamal MA, Basilio D, Bukauskas FF, Bennett MV (2005) Connexin-based gap junction hemichannels: gating mechanisms. Biochim Biophys Acta 1711:215–224. https://doi.org/10.1016/j.bbamem.2005.01.014
Salameh A, Wustmann A, Karl S, Blanke K, Apel D, Rojas-Gomez D, Franke H, Mohr FW, Janousek J, Dhein S (2010) Cyclic mechanical stretch induces cardiomyocyte orientation and polarization of the gap junction protein connexin43. Circ Res 106:1592–1602. https://doi.org/10.1161/circresaha.109.214429
Sanchez JA, Rodriguez-Sinovas A, Barba I, Miro-Casas E, Fernandez-Sanz C, Ruiz-Meana M, Alburquerque-Bejar JJ, Garcia-Dorado D (2013) Activation of RISK and SAFE pathways is not involved in the effects of Cx43 deficiency on tolerance to ischemia-reperfusion injury and preconditioning protection. Basic Res Cardiol 108:351. https://doi.org/10.1007/s00395-013-0351-3
Schulz R, Gorge PM, Gorbe A, Ferdinandy P, Lampe PD, Leybaert L (2015) Connexin 43 is an emerging therapeutic target in ischemia/reperfusion injury, cardioprotection and neuroprotection. Pharmacol Ther 153:90–106. https://doi.org/10.1016/j.pharmthera.2015.06.005
Schulz R, Gres P, Skyschally A, Duschin A, Belosjorow S, Konietzka I, Heusch G (2003) Ischemic preconditioning preserves connexin 43 phosphorylation during sustained ischemia in pig hearts in vivo. FASEB J 17:1355–1357. https://doi.org/10.1096/fj.02-0975fje
Schwanke U, Konietzka I, Duschin A, Li X, Schulz R, Heusch G (2002) No ischemic preconditioning in heterozygous connexin43-deficient mice. Am J Physiol Heart Circ Physiol 283:H1740-1742. https://doi.org/10.1152/ajpheart.00442.2002
Schwanke U, Li X, Schulz R, Heusch G (2003) No ischemic preconditioning in heterozygous connexin 43-deficient mice–a further in vivo study. Basic Res Cardiol 98:181–182. https://doi.org/10.1007/s003950300002
Shan H, Wei J, Zhang M, Lin L, Yan R, Zhang R, Zhu YH (2015) Suppression of PKCepsilon-mediated mitochondrial connexin 43 phosphorylation at serine 368 is involved in myocardial mitochondrial dysfunction in a rat model of dilated cardiomyopathy. Mol Med Rep 11:4720–4726. https://doi.org/10.3892/mmr.2015.3260
Shintani-Ishida K, Uemura K, Yoshida K (2007) Hemichannels in cardiomyocytes open transiently during ischemia and contribute to reperfusion injury following brief ischemia. Am J Physiol Heart Circ Physiol 293:H1714-1720. https://doi.org/10.1152/ajpheart.00022.2007
Skyschally A, Schulz R, Gres P, Korth HG, Heusch G (2003) Attenuation of ischemic preconditioning in pigs by scavenging of free oxyradicals with ascorbic acid. Am J Physiol Heart Circ Physiol 284:H698-703. https://doi.org/10.1152/ajpheart.00693.2002
Slavi N, Toychiev AH, Kosmidis S, Ackert J, Bloomfield SA, Wulff H, Viswanathan S, Lampe PD, Srinivas M (2018) Suppression of connexin 43 phosphorylation promotes astrocyte survival and vascular regeneration in proliferative retinopathy. Proc Natl Acad Sci USA 115:E5934–E5943. https://doi.org/10.1073/pnas.1803907115
Soetkamp D, Nguyen TT, Menazza S, Hirschhauser C, Hendgen-Cotta UB, Rassaf T, Schluter KD, Boengler K, Murphy E, Schulz R (2014) S-nitrosation of mitochondrial connexin 43 regulates mitochondrial function. Basic Res Cardiol 109:433. https://doi.org/10.1007/s00395-014-0433-x
Solan JL, Lampe PD (2018) Spatio-temporal regulation of connexin43 phosphorylation and gap junction dynamics. Biochim Biophys Acta 1860:83–90. https://doi.org/10.1016/j.bbamem.2017.04.008
Solan JL, Marquez-Rosado L, Lampe PD (2019) Cx43 phosphorylation-mediated effects on ERK and Akt protect against ischemia reperfusion injury and alter the stability of the stress-inducible protein NDRG1. J Biol Chem 294:11762–11771. https://doi.org/10.1074/jbc.RA119.009162
Solan JL, Marquez-Rosado L, Sorgen PL, Thornton PJ, Gafken PR, Lampe PD (2007) Phosphorylation at S365 is a gatekeeper event that changes the structure of Cx43 and prevents down-regulation by PKC. J Cell Biol 179:1301–1309. https://doi.org/10.1083/jcb.200707060
Srisakuldee W, Jeyaraman MM, Nickel BE, Tanguy S, Jiang ZS, Kardami E (2009) Phosphorylation of connexin-43 at serine 262 promotes a cardiac injury-resistant state. Cardiovasc Res 83:672–681. https://doi.org/10.1093/cvr/cvp142
Srisakuldee W, Makazan Z, Nickel BE, Zhang F, Thliveris JA, Pasumarthi KB, Kardami E (2014) The FGF-2-triggered protection of cardiac subsarcolemmal mitochondria from calcium overload is mitochondrial connexin 43-dependent. Cardiovasc Res 103:72–80. https://doi.org/10.1093/cvr/cvu066
Steffens M, Göpel F, Ngezahayo A, Zeilinger C, Ernst A, Kolb HA (2008) Regulation of connexons composed of human connexin26 (hCx26) by temperature. Biochim Biophys Acta 1778:1206–1212. https://doi.org/10.1016/j.bbamem.2008.01.016
Sun J, Nguyen T, Aponte AM, Menazza S, Kohr MJ, Roth DM, Patel HH, Murphy E, Steenbergen C (2015) Ischaemic preconditioning preferentially increases protein S-nitrosylation in subsarcolemmal mitochondria. Cardiovasc Res 106:227–236. https://doi.org/10.1093/cvr/cvv044
Totzeck A, Boengler K, van de Sand A, Konietzka I, Gres P, Garcia-Dorado D, Heusch G, Schulz R (2008) No impact of protein phosphatases on connexin 43 phosphorylation in ischemic preconditioning. Am J Physiol Heart Circ Physiol 295:H2106-2112. https://doi.org/10.1152/ajpheart.00456.2008
Tu RH, Li QJ, Huang Z, He Y, Meng JJ, Zheng HL, Zeng ZY, Zhong GQ (2017) Novel functional role of heat shock protein 90 in mitochondrial connexin 43-mediated hypoxic postconditioning. Cell Physiol Biochem 44:982–997. https://doi.org/10.1159/000485399
Turner MS, Haywood GA, Andreka P, You L, Martin PE, Evans WH, Webster KA, Bishopric NH (2004) Reversible connexin 43 dephosphorylation during hypoxia and reoxygenation is linked to cellular ATP levels. Circ Res 95:726–733. https://doi.org/10.1161/01.RES.0000144805.11519.1e
Varro A, Negretti N, Hester SB, Eisner DA (1993) An estimate of the calcium content of the sarcoplasmic reticulum in rat ventricular myocytes. Pflugers Arch 423:158–160. https://doi.org/10.1007/bf00374975
Wang H, Stahl F, Scheper T, Steffens M, Warnecke A, Zeilinger C (2019) Microarray-based screening system identifies temperature-controlled activity of Connexin 26 that is distorted by mutations. Sci Rep 9:13543. https://doi.org/10.1038/s41598-019-49423-3
Wang N, De Vuyst E, Ponsaerts R, Boengler K, Palacios-Prado N, Wauman J, Lai CP, De Bock M, Decrock E, Bol M, Vinken M, Rogiers V, Tavernier J, Evans WH, Naus CC, Bukauskas FF, Sipido KR, Heusch G, Schulz R, Bultynck G, Leybaert L (2013) Selective inhibition of Cx43 hemichannels by Gap19 and its impact on myocardial ischemia/reperfusion injury. Basic Res Cardiol 108:309. https://doi.org/10.1007/s00395-012-0309-x
Yu L, Zhao Y, Xu S, Ding F, Jin C, Fu G, Weng S (2013) Advanced glycation end product (AGE)-AGE receptor (RAGE) system upregulated connexin43 expression in rat cardiomyocytes via PKC and Erk MAPK pathways. Int J Mol Sci 14:2242–2257. https://doi.org/10.3390/ijms14022242
Acknowledgements
The authors want to thank Anna Reis, Sabrina Böhme, Elvira Ungefug, Nadine Woitasky, and Martin Heisler for excellent technical assistance.
Funding
Open Access funding enabled and organized by Projekt DEAL. The present study was funded by the German Research Foundation (BO 2955/4–1) and the US National Institutes of Health (PDL-GM55632).
Author information
Authors and Affiliations
Contributions
CH, AL, and PMG conducted experiments and analyzed data, PDL provided mice and antibodies, PDL, KDS, LL, JH RS, and KB designed research and analyzed data, KB drafted the manuscript, and all authors contributed to the final version of the manuscript.
Corresponding author
Ethics declarations
Conflicts of interest
RS received honoraries for lectures provided by Sanofi.
Ethics approval
All animal experiments were performed in accordance to guidelines of the local animal ethics committees.
Consent to participate
Not applicable.
Consent for publication
All authors have read and approved the submission of the manuscript.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hirschhäuser, C., Lissoni, A., Görge, P.M. et al. Connexin 43 phosphorylation by casein kinase 1 is essential for the cardioprotection by ischemic preconditioning. Basic Res Cardiol 116, 21 (2021). https://doi.org/10.1007/s00395-021-00861-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00395-021-00861-z