
Abstract
Aims/hypothesis
Diabetes accelerates cardiovascular disease caused by atherosclerosis. Accordingly, diabetes accelerates atherosclerotic lesion progression and increases arterial smooth muscle cell proliferation. We hypothesized that diabetes can exert growth-promoting effects on smooth muscle cells via increased advanced glycation end-products or by dyslipidaemia.
Methods
Primary human arterial smooth muscle cells were stimulated with advanced glycation end-products, other ligands of the receptor for advanced glycation end-products or fatty acids common in triglycerides. Cell proliferation was measured as DNA synthesis, cell cycle distribution and cell number. Effects of oleate on cellular phospholipids, diacylglycerol, triglycerides and cholesterol esters were analyzed by thin-layer chromatography, and oleate accumulation into diacylglycerol was confirmed by gas chromatography.
Results
Human arterial smooth muscle cells express the receptor for advanced glycation end-products, but its ligands Nε-(carboxymethyl)lysine-modified proteins, methylglyoxal-modified proteins, S100B polypeptide and amyloid-β (1–40) peptide, exert no mitogenic action. Instead, oleate, one of the most common fatty acids in triglycerides, enhances platelet-derived growth factor-BB-mediated proliferation and oleate-containing 1,2-diacylglycerol formation in smooth muscle cells. This mitogenic effect of oleate depends on phospholipase D activity and is associated with an increased formation of oleate-enriched 1,2-diacylglycerol.
Conclusion/interpretation
Oleate, not ligands of the receptor for advanced glycation end-products, acts as an enhancer of human smooth muscle cell proliferation. Thus, lipid abnormalities, rather than hyperglycaemia, could be a major factor promoting proliferation of smooth muscle cells in atherosclerotic lesions.
Similar content being viewed by others
Atherosclerosis is accelerated by both Type I and Type 2 diabetes [1] by mechanisms that are poorly understood. Proliferation of arterial smooth muscle cells (SMCs) in lesions of atherosclerosis plays an important role in progression of early fatty streaks to fibroatheromas. We have recently reported that increased SMC proliferation in fibroatheromas occurs concomitantly with hyperglycaemia and an increased plasma triglyceride concentration in a porcine model of diabetes-accelerated atherosclerosis [2]. However, increased plasma glucose concentrations do not directly stimulate proliferation of SMCs isolated from these animals or from human subjects [2]. In this study, we investigated advanced glycation end-products (AGEs) and non-esterified fatty acids (NEFAs) as potential detrimental factors for SMCs.
AGEs are formed under hyperglycaemic and hyperlipidaemic conditions [3] and are present in atherosclerotic lesions [4]. These structures bind to a number of cellular proteins and have been shown to exert a plethora of cellular responses. AGE-binding proteins identified include the components of the AGE-receptor complex p60, p90 and galectin-3 [5, 6], receptor for AGEs (RAGE) [7], the class B scavenger receptor CD36 [8], and class A-type I and II scavenger receptors [9]. To date, the only AGE receptor with clear signalling capacities is RAGE, which can be activated by AGEs, but also by other ligands including amphoterin, amyloid peptides, S100 polypetides, and transthyretin [10]. Blocking interaction of RAGE with its ligands [11, 12, 13] can inhibit several processes involved in atherosclerosis.
Hypertriglyceridaemia is another risk factor for the development of atherosclerosis in people with and without diabetes [14] and is frequently seen in diabetic patients with cardiovascular disease [15] and in animal models of diabetes-accelerated atherosclerosis [16, 17]. NEFAs are liberated from triglycerides in the arterial wall by lipoprotein lipase [18]. The main fatty acids present in triglycerides in humans with and without diabetes are oleate and palmitate [19, 20]. Increased levels of circulating triglycerides in combination with hydrolysis of these triglycerides in the atherosclerotic plaque are likely to result in increased levels of NEFAs within the plaque.
The aim of this study was to compare the effects of AGEs to those of oleate and other common fatty acids on proliferation of primary human arterial SMCs. Non-transformed human arterial SMCs often respond differently than SMCs isolated from other species, such as rodent SMCs [21, 22], and studies of human SMCs are therefore of importance.
Materials and methods
Growth factors, peptides, proteins and lipids
BSA (endotoxin-free, grade VI), ovalbumin, human angiotensin II and human recombinant insulin were obtained from Sigma (St. Louis, Mo., USA). S100B was obtained from Calbiochem (La Jolla, Calif., USA). Wild-type amyloid-β (Aβ) peptide (1–40) was obtained from Biosource International (Camarillo, Calif., USA). The lyophilized peptide was dissolved in distilled H2O at 6 mg/ml, diluted to 1 mg/ml in calcium-free PBS and then incubated at 37°C for 36 h to induce peptide aggregation. Human recombinant platelet-derived growth factor B-chain homodimer (PDGF-BB) and insulin-like growth factor I (IGF-I) were from Upstate Biotechnology (Lake Placid, N.Y., USA). Human recombinant tumour necrosis factor-α (TNF-α) and interleukin 1β were obtained from PeproTech (Rocky Hill, N.J., USA). Sodium salts of palmitic acid (16:0), stearic acid (SA, 18:0), oleic acid (OA, cis 18:1), elaidic acid (EA, trans 18:1), linoleic acid (LA, cis 18:2), and conjugated linoleic acid (18:2) were from Nu-Chek Prep (Elysian, Minn., USA). Coenzyme A (CoA)-esters of OA, LA, SA and EA were obtained from Sigma. Propranolol was from Biomol Research Laboratories (Plymouth Meeting, Pa., USA) and 1-butanol and 2-butanol were obtained from Fisher Scientific Company (Houston, Tex., USA). [1-14C]-OA (2.22 GBq/mmol) was obtained from Amersham Biosciences (Piscataway, N.J., USA).
Cultured cells
Normal human aortic SMCs from four different donors were isolated from newborn aorta by an explant method [2]. SMC cultures were kept in Dulbecco’s Modified Eagle’s medium (GIBCO/BRL Life Technologies, Rockville, Md., USA) containing 5.6 mmol/l glucose and 0.5% human plasma-derived serum for 2 days prior to experiments to induce cellular quiescence. Cells were characterized as SMCs by morphologic criteria and by expression of smooth muscle α-actin, as shown by immunocytochemistry using a mouse monoclonal anti-smooth muscle α-actin antibody (clone 1A4; DAKO, Carpinteria, Calif., USA). Glucose levels in the culture media were monitored to avoid glucose depletion [23]. SMCs were used for experiments between passages 2 and 12.
Human umbilical vein endothelial cells (HUVECs) were isolated from donors without known disease [24]. HUVECs were cultured in RPMI (GIBCO/BRL Life Technologies) containing 20% fetal bovine serum and endothelial cell growth supplement (BD Biosciences, San Jose, Calif., USA), and were used for experiments between passages 1 and 3. All experiments were carried out in the absence of an endothelial cell growth supplement. Informed consent was obtained from all subjects according to the guidelines of the University of Washington. The studies were approved by the Human Subjects Committee at the University of Washington, and were carried out in accordance with the principles of the Declaration of Helsinki as revised in 2000.
Western blot analysis of RAGE expression
A rabbit polyclonal antibody against the N-terminal 16 amino acids (AQNITARIGEPLVLKC) of mature human RAGE conjugated to keyhole limpet haemocyanin was produced in rabbits (Genemed Synthesis, South San Francisco, Calif., USA). For Western blot analysis of RAGE expression, cell lysates were prepared and subjected to separation on 10% SDS polyacrylamide gels. The proteins were transferred to Immobilon-P membranes (Millipore, Bedford, Mass., USA). The membranes were blocked in 5% milk in PBS-Tween for 1 h at room temperature and the RAGE antiserum or preimmune serum was then incubated with the membrane at a 1:1000 dilution overnight at 4°C. For blocking experiments, the RAGE antiserum was pre-incubated with the antigen peptide (5 µg/ml) for 1 h at 37°C prior to incubation with membranes. This was followed by incubation with the secondary horseradish peroxidase-conjugated anti-rabbit antibody (Amersham Biosciences, Arlington Heights, Ill., USA) at a 1:5000 dilution. Western blots were developed by enhanced chemiluminescence according to the manufacturer’s instructions (Amersham Biosciences).
Production of AGE-modified proteins
BSA and ovalbumin (25 mg/ml) were glycated in endotoxin-free PBS (10 mmol/l, pH 7.4) containing 0.1 µg/ml pepstatin A, 0.5 µg/ml leupeptin, 2 µg/ml aprotinin, 1.5 mmol/l PMSF, 1 mmol/l EDTA, 1 mmol/l NaN3, 1 mol/l D-glucose and 0.5 mol/l glucose-6-phosphate for 8 weeks at 37°C under sterile aerobic conditions. The resulting proteins are referred to as AGE-BSA and AGE-ovalbumin. Control proteins were incubated under the same conditions in the absence of glucose and glucose-6-phosphate.
CML-modification of BSA was carried out according to a known method [25]. In short, BSA was incubated in endotoxin-free PBS (pH 7.8) in the presence of glyoxylic acid and NaCNBH3 (both from Sigma) at different ratios for 10 min to 24 h. Using a 10-min reaction and a ratio of glyoxylic acid:NaCNBH3 of 1:1, 1:2 and 1:7, we obtained CML-modified BSA at levels of 462, 277 and 97 mmoles modified NH2-residues/moles total NH2-residues, respectively. Control protein was incubated under the same conditions in the absence of glyoxylic acid. Minimal modification of BSA with 500 µmol/l methylglyoxal (MGO; Sigma) was carried out for 24 h at 37°C [26].
At the end of the glycation process the proteins were dialyzed (Slide-A-Lyzer Dialysis cassette, Mw cut-off 10,000 kDa; Pierce Chemical, Rockford, Ill., USA) against endotoxin-free PBS (pH 7.6) under sterile conditions for 72 h. Protein concentrations were then measured by the BCA-200 protein assay kit (Pierce).
Measurement of endotoxin levels
Endotoxin was removed, when needed, prior to modification of BSA or ovalbumin using Detoxi-Gel columns (Pierce). Endotoxin levels were measured using a chromogenic limulus amebocyte lysate test (QCL-1000; Whittaker M.A. Bioproducts, Walkersville, Md., USA). All samples showed endotoxin levels less than 5 pg/ml final concentration.
Characterization of AGE-modified proteins
SDS-PAGE analysis under reducing conditions was used to evaluate integrity of the modified proteins and mobility shifts caused by glycation. The 2,4,6-trinitrobenzenesulfonic acid (TNBS, Sigma) assay was used to quantify mmoles modified NH2-residues versus moles of total NH2-residues, using unmodified BSA as the calibration standard [27], thus giving the % modification. Fluorescence of AGEs (exc: 370 nm, em: 440 nm) was also measured. Finally, the amount of modification was determined by Matrix Assisted Laser Desorption/Ionization Time of Flight (MALDI-TOF) mass spectrometry for some AGE-modified proteins. Protein samples (1.5 µl, 10 pmol/µl) were diluted in 1.5 µl saturated dihydroxybenzoic acid, air dried on a MALDI sample plate and subjected to mass determination using a Bruker Biflex III MALDI-TOF mass spectrometer (Bruker Daltonics, Billerica, Mass., USA). The instrument was equipped with a 337 nm nitrogen laser.
Preparation of fatty acid-BSA complex
Sodium salts of fatty acids were diluted to a final concentration of 70 µmol/l, unless otherwise stated, in sterile Dulbecco’s Modified Eagle’s Medium containing 70 µmol/l fatty acid-free BSA (Sigma). This mixture was equilibrated for 1 h at 37°C, 5% CO2, prior to addition to the cells, allowing BSA-fatty acid complexes to form. This method has been estimated to result in an effective free fatty acid concentration in the nanomolar range [28]. The concentrations of fatty acid and BSA were based on a physiological ratio between fatty acid and the carrier protein and the concentration of albumin present in the extracellular fluid of the intimal portion of the arterial wall [29].
Measurement of SMC DNA synthesis, cell cycle distribution, and proliferation
DNA synthesis was measured as [3H]-thymidine incorporation into DNA [2]. Growth factors were added 3–4 h after stimulation with fatty acids. Cell cycle distribution was measured by flow cytometry. Following stimulation, SMCs were trypsinized, and DNA was then stained using 4,6-diamidino-2-phenylindole [30]. The fraction of cells in the G1, S and G2/M phases of the cell cycle was determined by using a Coulter Epics Elite flow cytometer (Coulter, Hialeah, Fla., USA) equipped with an argon laser (360 nm exc) and MPlus software [30]. Cell proliferation was also measured by determining cell number according to a known method [2].
VCAM-1 gene expression
Regulation of vascular adhesion molecule-1 (VCAM-1) mRNA was studied in HUVECs stimulated for 4 h with AGE-BSA or BSA control, or by TNF-α (10 nmol/l) as a positive control. Total RNA was extracted from the cells by using RNAeasy kits from Qiagen (Valencia, Calif., USA). VCAM-1 mRNA was quantified by semi-quantitative RT-PCR [31], using the Titan One Tube RT-PCR system (Roche Molecular Biochemicals, Indianapolis, Ind., USA). Custom made human VCAM-1 primers (Gibco BRL) were: 5′ primer 5′-CCCTTGACCGGCTGGAGATT-3′ and 3′ primer 5′-CTGGGGGCAACATTGACATAAAGTG-3′. The linearity of the reaction was optimized by using 0.3 µg RNA/reaction and 35 cycles. Annealing and elongation were achieved using a three-step program (30 s 94°C, 30 s 58°C, 60 s 68°C) that was concluded with 7 min at 68°C. The samples were then separated on 1% agarose gels, visualized by ethidium bromide (GIBCO/BRL), and photographed using an Eagle Eye II (Stratagene, Cedar Creek, Tex., USA). The reaction produced a single band of the expected size (241 bp). Human β-actin mRNA was quantified as a loading control. The primers for β-actin (Clontech Laboratories, Palo Alto, Calif., USA) were: 5′ primer 5′-ATCTGGCACCACACCTTCTACAATGAGCTGCG-3′ and 3′ primer 5′-CGTCATACTCCTGCTTGCTGATCCACATCTGC-3′. The reaction produced a single band of the expected size (838 bp). The intensities of the VCAM-1 bands were quantified by NIH Image 1.62 software (developed at the Research Services Branch of the National Institute of Mental Health) and normalized to the intensities of the β-actin bands.
Analysis of cellular lipids and accumulation of oleate in lipid pools
SMCs in 100-mm dishes were labelled with [14C]-OA (37 kBq/plate; 1 µmol/l) complexed to 70 µmol/l fatty acid-free BSA in the presence or absence of 70 µmol/l unlabelled OA. The cells were incubated at 37°C for 18 h and then stimulated with PDGF-BB (1 nmol/l) for 15 min. Membrane lipids were extracted using a modified method [32]. Lipid extracts were applied to unmodified silica gel G thin-layer chromatography plates and developed in a solvent system of hexane: diethyl ether:glacial acetic acid (105:45:3). Standards containing mono-, di-, and triglycerides (Nucheck Prep, Elysian Fields, Minn., USA), and cellular lipids were visualized by exposure of the plate to iodine vapour. The intensity of the iodine-visualized spots was quantified using the NIH Image 1.62 software. Plates were then dried and exposed to a phosphor screen for 24 h. A Molecular Dynamics Storm 860 PhosphorImager was used for detection and quantification of radioactive spots.
Determination of fatty acid chain composition of 1,2-diacylglycerol
The fatty acid composition of 1,2-diacylglycerol (1,2-DAG) was analyzed by gas chromatography. SMCs (3 million cells/sample) were incubated in the presence or absence of 70 µmol/l OA bound to BSA for 18 h and then stimulated with or without 1 nmol/l PDGF-BB for 15 min. Neutral lipids were then extracted and separated by thin-layer chromatography. Butylated hydroxytoluene (44 mg/l) was included to prevent oxidation of fatty acids. The 1,2-DAG spots were directly transesterified to prepare fatty acid methyl esters [33]. The fatty acid methyl esters were then separated by gas chromatography [34].
Statistical analysis
Statistical analysis was done using one-way analysis of variance followed by the Newman-Keuls multiple comparison test (Graph Pad Prism; Graph Pad Software, San Diego, Calif., USA). A p value less than 0.05 was considered statistically significant.
Results
RAGE is expressed by human SMCs
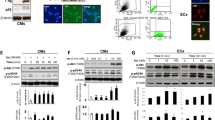
To verify that human arterial SMCs express RAGE, we generated a rabbit polyclonal antibody against human RAGE. This antibody detects the purified extracellular part of RAGE, whereas the preimmune serum does not detect RAGE (Fig. 1a). In extracts from human SMCs and HUVECs, the RAGE antibody detected a band of about 51 kDa corresponding to full-length RAGE (Fig. 1b). The specificity of the signal was verified by RAGE peptide competition experiments. Two non-specific bands were also detected by the RAGE antibody, and this signal was not blocked by the RAGE peptide.

RAGE is expressed by human arterial SMCs. A polyclonal anti-RAGE antibody was raised in rabbits. The pre-immune serum (PI) did not detect the extracellular part of purified soluble RAGE, sRAGE (generously provided by Dr. A.M. Schmidt), whereas the immune serum (I) recognized purified RAGE (a) as well as full-length RAGE (b) expressed in human SMCs and HUVECs (ECs). The antigen peptide completely blocked the ability of the RAGE antiserum to detect sRAGE (a) and full-length RAGE (b) in SMCs and endothelial cells. The experiment was carried out three times with similar results
Characterisation of AGE-modified proteins
As shown by SDS-PAGE, the glycation process caused an increase in the apparent molecular weight of BSA and ovalbumin. This was especially noticeable for AGE-BSA and AGE-ovalbumin, with smaller changes seen in MGO- and CML-modified proteins (Fig. 2a). AGE-BSA and AGE-ovalbumin also showed a marked increase in fluorescence compared to controls, whereas CML- and MGO-proteins had a weak or no fluorescence at 440 nm. However, the degree of modification of amino residues in these preparations, as evaluated by the TNBS assay, was increased by 40–64% compared to controls (Fig. 2a).

Analysis of glycated proteins. (a) The extent of glycation of BSA and ovalbumin (OVA) was analyzed by SDS-PAGE, the TNBS assay and by fluorescence (exc: 370 nm, em: 440 nm). The extent of modification of proteins and shifts in the apparent molecular weights due to the glycation process are shown. The experiment was repeated three times with similar results. (b) The extent of modification of CML-BSA modified using a 1:1 ratio of glyoxylic acid:NaCNBH3 for 24 h was analyzed using MALDI-TOF mass spectrometry. A 1:1 mixture of BSA and CML-BSA was analyzed. The double charged BSA and CML-BSA peaks are shown. The mass/charge ratio (m/z) of each peak is indicated
MALDI-TOF mass spectrometry was used to measure the number of modified residues compared to control proteins. This method indicated that the control BSA used did not exhibit basal modification. The maximal CML modification protocol used (a 1:1 ratio of glyoxylic acid:NaCNBH3 and a 24-h reaction) resulted in a single defined peak (Fig. 2b). The calculated number of modified residues in this CML-BSA preparation was 67. The results obtained by MALDI-TOF mass spectrometry closely resembled those obtained by the TNBS assay.
RAGE ligands do not induce proliferation of human SMCs
None of the glycated proteins induced DNA synthesis in human SMCs (Fig. 3a–c). Incubation of SMCs under high glucose (25 mmol/l) conditions did not affect their response to glycated proteins (data not shown). Furthermore, no effect of the different glycated proteins on PDGF-BB-induced DNA synthesis was observed (Fig. 4a). The glycated proteins also did not modulate the effects of IGF-I, insulin, angiotensin II, TNF-α or interleukin 1β. Two other RAGE ligands, S100B and amyloid-β peptide, did not stimulate DNA synthesis (Fig. 3d) or enhance the effects of PDGF-BB (Fig. 4b). The lack of growth-promoting effect of RAGE ligands was verified by determining the number of SMCs after a 6-day incubation.

RAGE ligands do not induce DNA synthesis in human SMCs. SMCs (50,000 cells/well) were stimulated for 20 h with AGE-BSA (a), CML-BSA (b), MGO-BSA (c), S100B peptide, or amyloid-β peptide (d) at indicated concentrations. Relevant BSA controls of the same batch were included in each experiment. [3H]-thymidine incorporation into DNA was subsequently measured as trichloroacetic acid-insoluble radioactive material. PDGF-BB was used as a positive control (black bars), and cells incubated without BSA were used as negative controls (open bars). The experiment was carried out three times with similar results. The values are presented as means±SEM of triplicate samples of representative experiments

RAGE ligands do not increase PDGF-BB induced DNA synthesis in human SMCs. (a) Human SMCs were incubated in the absence or presence of glycated proteins (100 µg/ml) and/or PDGF-BB (1 nmol/l). (b) Human SMCs were incubated with or without S100 (100 nmol/l), or amyloid-β peptide (1 or 5 µmol/l) and/or PDGF-BB. [3H]-thymidine incorporation into DNA was measured as described in Fig. 3. The results are presented as means±SEM of triplicate samples of representative experiments performed at least three times
In order to verify that the AGE-BSA used in this study was biologically active, its ability to induce VCAM-1 gene expression in HUVECs was investigated. AGE-BSA, but not control BSA, induced a clear increase in VCAM-1 gene expression after a 4-h stimulation, as did TNF-α (Fig. 5a). Quantification of the intensities of the VCAM-1 bands and normalization to the intensities of the corresponding β-actin bands (Fig. 5b) revealed that TNF-α induced a 3.7-fold induction of VCAM-1 mRNA, 100 µg/ml AGE-BSA induced a 2.0-fold increase in VCAM-1 mRNA compared to control while 300 µg/ml AGE-BSA induced a 2.6-fold increase in VCAM-1 mRNA.

AGE-BSA induces VCAM-1 gene expression in HUVECs. (a) HUVECs were stimulated with AGE-BSA or control BSA at indicated concentrations, or with TNF-α. RNA was extracted and VCAM-1 mRNA levels were analyzed by semi-quantitative RT-PCR. A negative control with all reagents except the RNA was included in each analysis (Neg. control). (b) Human β-actin mRNA levels were measured in the same samples as a loading control. The experiment was repeated twice with similar results
Oleate stimulates proliferation of human SMCs
OA, but not EA or conjugated LA, induced an increase in DNA synthesis (Fig. 6a). More strikingly, OA enhanced the mitogenic effect of PDGF-BB (Fig. 6a) and IGF-I (not shown). The effect of LA was smaller than that of OA, but it did reach statistical significance (p<0.001) in SMCs from two of the three donors investigated. SA and palmitic acid had no mitogenic activity. Since fatty acids require the esterification to a CoA-moiety prior to utilization in the cell, we also investigated the effect of CoA-modified fatty acids. OA-CoA stimulated DNA synthesis and enhanced the effects of PDGF-BB approximately five-fold (Fig. 6b). LA-CoA (70 µmol/l) induced an approximate two-fold increase of the effects of PDGF-BB, whereas EA-CoA and SA-CoA had only weak effects.

Oleate enhances the growth-promoting effects of PDGF-BB in human SMCs. (a) Human SMCs (40 000 cells/well) were incubated in the presence of 70 µmol/l linoleic acid (LA), conjugated linoleic acid (cLA), oleic acid (OA) or elaidic acid (EA) for 4 h. The cells were then stimulated with PDGF-BB (1 nmol/l) for an additional 20 h. [3H]-thymidine incorporation into DNA was measured as described in Fig. 3. (b) SMCs were stimulated with 70 µmol/l OA-CoA in the presence or absence of 1 nmol/l PDGF-BB. (c) SMCs (189,370±1861 cells/well, n=3, means±SEM) were stimulated with 70 µmol/l OA in the presence or absence of 1 nmol/l PDGF-BB for 3 days, and then counted. The results are presented as means±SEM of triplicate samples of a representative experiment carried out at least three times. # p<0.05 compared to basal; ### p<0.001 compared to basal; *** p<0.001 compared to PDGF-BB alone
OA also enhanced the effect of PDGF-BB on cell cycle progression. Thus, more cells were found in the S- and G2/M phases of the cell cycle, and fewer cells were in G1, after stimulation with OA plus PDGF-BB compared to stimulation with PDGF-BB alone (Fig. 7). Consistently, OA enhanced the effect of PDGF-BB on SMC number (Fig. 6c).

Oleate enhances cell cycle progression induced by PDGF-BB. Human SMCs were incubated without (a, c) or with 70 µmol/l OA (b, d) for 4 h, and then for an additional 24 h without (a, b) or with (c, d) 1 nmol/l PDGF-BB. Cell cycle distribution was analyzed by flow cytometry. The fractions of cells in G1, S and G2/M are shown as means of triplicate samples ± SEM of a representative experiment. The experiment was carried out three times with similar results. NS, non-significant compared to basal (a); *** p<0.001 compared to PDGF-BB alone (c)
Oleate increases levels of triglycerides and unesterified fatty acid in human SMCs
Stimulation of SMCs with OA for 18 h resulted in more than a four-fold increase in cellular triglyceride levels and a two- to three-fold increase in free fatty acid levels. Total levels of cholesterol esters and phospholipids were not affected by OA or PDGF-BB (Table 1).
We then examined the distribution of [14C]-OA in neutral lipid pools and phospholipids in SMCs exposed to 70 µmol/l BSA-bound OA compared to SMCs exposed to tracer amounts (1 µmol/l) of [14C]-OA bound to 70 µmol/l BSA. Tracer amounts of OA were added in order to visualize the lipid pools. However, SMCs stimulated with tracer amounts of OA cannot be compared to SMCs that have not been exposed to OA, because even nanomolar concentrations of carrier-bound OA are rapidly taken up by cells. An accumulation of [14C]-OA in the triglyceride and free fatty acid pools occured in SMCs that had been exposed to 70 µmol/l OA compared to SMCs that had been exposed to tracer amounts of [14C]-OA alone, reflecting the increase in total amounts of these lipid pools (Table 1). Conversely, accumulation of [14C]-OA in the phospholipid pool was lower in SMCs exposed to 70 µmol/l OA compared to SMCs exposed to tracer concentrations of [14C]-OA alone. There were no effects of a 15 min stimulation with PDGF-BB on the accumulation of [14C]-OA into these lipid pools (Table 1).
Oleate enhances the effects of PDGF-BB by stimulating generation of OA-enriched 1,2-DAG
We hypothesized that the effect of OA on PDGF-BB’s mitogenic effect could be due to PDGF-BB-induced activation of phospholipase D (PLD) and subsequent hydrolysis of OA-enriched phosphatidylcholine. To address this hypothesis, several experiments were carried out. Firstly, if PDGF-BB leads to PLD activation and hydrolysis of OA-containing phosphatidylcholine, PDGF-BB should give rise to formation of OA-containing 1,2-DAG in SMCs incubated with OA. The results show that this is indeed the case. PDGF-BB stimulation resulted in more [14C]-OA-containing 1,2-DAG compared to unstimulated SMCs (Fig. 8a). Levels of total 1,2-DAG were also increased by PDGF in SMCs exposed to 70 µmol/l OA (Fig. 8b).

Oleate enhances formation of OA-containing 1,2-DAG following PDGF-BB stimulation and increases PDGF-BB-induced DNA synthesis through a PLD-dependent mechanism. Human SMCs were incubated with tracer amounts of BSA-bound [14C]-OA in the presence or absence of unlabelled OA (70 µmol/l) for 18 h, and then for an additional 15 min in the presence or absence of PDGF-BB. Lipids were extracted and analyzed by thin-layer chromatography. Spots containing radioactive [14C]-OA were analyzed by using a PhosphorImager (a) whereas total lipid spots were visualized by iodine vapor (b). Values are represented as % of control SMCs incubated in the presence of tracer amouns of [14C]-OA (means±SEM; n=4). (c) Human SMCs were incubated in the presence or absence of OA (70 µmol/l) for 6 h, and then for an additional 20 h in the presence or absence of PDGF-BB. 1-butanol (1-but) and 2-butanol (2-but), both at 0.2% v/v, were added 30 min prior to stimulation with PDGF-BB. DNA synthesis was measured as described in Fig. 3. Values are represented as means±SEM of triplicate samples of a representative experiment (n=3)
Inhibition of PLD action by 1-butanol, but not by the 2-butanol control, reduced the effect of OA on PDGF-BB-induced DNA synthesis (Fig. 8c). In other experiments, propranolol (30 µmol/l) was used to inhibit phosphatidate phosphohydrolase, which catalyses the conversion of phosphatidic acid to DAG. Propranolol also inhibited the stimulatory effect of OA. Thus, the growth-promoting effects of OA are likely to be mediated by increased formation of OA-enriched 1,2-DAG mediated by the PLD pathway in human SMCs.
Exposure of human SMCs to oleate results in a dramatic change in 1,2-DAG fatty acid composition
In control SMCs, the saturated fatty acids myristic acid, stearic acid and palmitic acid made up 62% of the DAG acyl chains, whereas only 15% was comprised of OA. In contrast, in SMCs exposed to 70 µmol/l BSA-bound OA, OA content in DAG was increased two-fold and the content of the saturated fatty acids was decreased (Table 2). Thus, exposure of human SMCs to physiological levels of carrier-bound OA results in a change in fatty acid composition of 1,2-DAG.
Discussion
RAGE ligands do not stimulate human SMC proliferation
The results of this study show that AGEs and other RAGE ligands do not directly induce proliferation of human arterial SMCs, despite expression of RAGE in these cells. Analysis of biological effects of AGEs is complicated by the fact that various AGE structures are included under the term “AGEs”, such as hydroimidazolones derived from methylglyoxal, CML and bis(lysyl)imidazolium protein cross-links. Therefore, we investigated the effect of AGEs prepared by several different methods. The product termed AGE-BSA contains different AGE structures. CML- and MGO-modified BSA preparations were chosen as more defined products and for their ability to bind and activate RAGE [25, 35, 36]. The extent of AGE-modification of lysine and arginine residues in blood cells in vivo is in the range of 0.01 mmol–15 mol [37]. Levels of CML-modification in skin collagen and lens proteins are in the order of 2–5 mmol CML/mol lysine [38]. This is lower than the extents of modification used in this study. However, the extent of AGE-modification in lesions of atherosclerosis is unknown.
The effects of AGEs on proliferation of SMCs have been investigated with contradictory results in cells derived from rat, rabbit, porcine, and human arteries. The different results are likely to be due to differences in the extent of modification and/or AGE structures present in proteins modified by different methods. There may also be species-dependent differences in SMC responses to AGEs. Thus, glycated albumin increases proliferation of rat [39, 40] and rabbit [41] SMCs. In other studies, AGE-BSA did not stimulate proliferation of pig or rabbit SMCs [42, 43]. The lack of effects of AGEs on human SMCs is consistent with results of a previous study using BSA incubated for 1 week in 20 mmol/l glucose-6-phosphate [44].
Of the non-AGE RAGE ligands described, S100 peptides are particularly interesting in the context of atherosclerosis because of their asscoation with inflammation [10]. However, in this study S100B, as well as another RAGE ligand, amyloid-β peptide [10], failed to induce proliferation of human SMCs. Again, these findings in human SMCs are in contrast to results showing that similar concentrations of S100B can induce DNA synthesis in murine SMCs [45]. Thus, human SMCs may respond differently than rodent SMCs to RAGE stimulation.
In summary, RAGE ligands do not directly stimulate proliferation of isolated human SMCs. It is possible that human SMCs are quite resistant to increased oxidative stress, which mediates the effects of AGE-RAGE interaction in endothelial cells and monocytes/macrophages [46]. Therefore, direct effects of AGEs and S100 peptides on endothelial cells [13, 47] and/or monocytes/macrophages [48, 49] might be more pronounced than their effects on SMCs.
OA could mediate its mitogenic effects by causing increased formation of OA-enriched 1,2-DAG following PLD activation
This study shows that OA, one of the most common fatty acids in triglycerides in people and animals with and without diabetes [17, 19, 20], strongly enhances the mitogenic effects of PDGF-BB in primary human SMCs. Previous studies have shown that OA also enhances the growth-promoting activity of angiotensin II [50], endothelin-1 [51], and IGF-I [52] in SMCs derived from other species. Our results show that when carrier protein-bound OA is added to human SMCs at physiological concentrations, OA is incorporated mainly (~60%) into the phospholipid pool. OA is also incorporated into triglycerides (~35%), and causes a four-fold increase in total levels of triglycerides. However, the increase in triglycerides is unlikely to explain the mitogenic effects of OA because elaidic acid, a positional isomer of OA, results in similar increases in triglycerides [53] without affecting PDGF-BB-induced DNA synthesis. Instead, the mitogenic effect of OA appears to be dependent on PLD activity and conversion of phosphatidic acid to 1,2-DAG. PDGF-BB activates PLD in SMCs [54, 55]. Accordingly, at a time (15 min) when PLD activation is maximal [55], PDGF-BB results in an increase in OA-containing 1,2-DAG in SMCs exposed to 70 µmol/l BSA-bound OA. As OA is incorporated mainly into phosphatidylcholine, the major substrate of PLD, and not into phosphatidylinositol, the major substrate of phospholipase C, [52] and because of the differences in kinetics between PDGF-induced phospholipase C and PLD activation [55, 56], the increased levels of OA-containing 1,2-DAG are almost certainly due to PDGF-BB induced activation of PLD. Indeed, the effects of OA on PDGF-induced DNA synthesis are blocked by inhibition of PLD action. These results strongly suggest that in SMCs that have been exposed to physiologically relevant levels of OA, PDGF-BB causes increased formation of OA-containing 1,2-DAG through a PLD-dependent pathway, as shown schematically by Fig. 9. Consistent with this model and a previous study [57], PDGF causes more DAG in OA-treated SMCs compared to control SMCs. Furthermore, 1,2-DAG contains twice as much OA in OA-treated SMCs compared to controls. The increased OA levels in DAG were reflected by lower levels of saturated fatty acids. Interestingly, it has been suggested that DAG species enriched in monounsaturated or unsaturated fatty acids act as better activators of protein kinase Cα than DAG species enriched in saturated fatty acids [58]. Accordingly, saturated fatty acids, such as palmitate and stearate, stimulate formation of DAG in SMCs [53, 59], but do not enhance SMC proliferation. Thus, the results of our study strongly indicate that OA enhances the mitogenic effects of PDGF-BB by causing increased formation of OA-enriched 1,2-DAG following PLD activation.

Model of the mitogenic effects of OA in human SMCs. A large fraction (~99%) of OA taken up by SMCs is esterified to form OA-CoA. OA-CoA contributes to de novo synthesis of DAG, triglycerides and phospholipids by a pathway that involves glycerol 3-phosphate (glycerol 3-P), lysophosphatidic acid (LPA), and phosphatidic acid (PA). Exposure of human SMCs to carrier protein-bound OA results in increased levels of free, unesterified OA and total levels of triglycerides. OA is distributed mainly into the phospholipid pool (phosphatidylcholine, PC). In SMCs exposed to OA, PDGF-BB increases levels of OA-containing 1,2-DAG, most likely due to activation of phospholipase D (PLD). We propose that OA enhances the mitogenic effects of PDGF-BB by two mechanisms involving generation of OA-enriched 1,2-DAG. Firstly, PDGF-BB-induced activation of PLD gives rise to OA-enriched 1,2-DAG, which is a more efficient activator of classical protein kinase C (PKC) isoforms than DAG containing saturated acyl chains. Secondly, OA could inhibit the activation of DAG kinase (DGK) mediated by PDGF-BB [57], thereby further increasing levels of OA-enriched 1,2-DAG. PPH, phosphatidate phosphohydrolase
Increased levels of circulating OA-containing triglycerides could enhance SMC proliferation in diabetes
Oleate is the principal fatty acid in triglycerides within the arterial wall of people with and without diabetes [60]. Although relative levels of oleate (expressed as percentage of total fatty acids) in circulating triglycerides are often found to be similar in people and animals with and without Type 1 diabetes [17, 20], some studies have shown an increased level of oleate in triglycerides or total serum lipids from Type 2 diabetic patients compared to non-diabetic patients [20, 61]. Furthermore, circulating levels of free fatty acids, including oleate, have been reported to be increased in subjects with Type 2 diabetes [15, 62, 63]. Finally, oleate levels are increased in small arteries from diabetic subjects compared to non-diabetic subjects [64]. Although no direct measurements have been done to date, these studies suggest that the arterial wall is exposed to increased levels of oleate in people with Type 2 diabetes.
Expression of lipoprotein lipase in isolated macrophages is increased by increased glucose levels [65], suggesting that its expression could be increased in the diabetic environment. Oleate liberated from triglycerides by lipoprotein lipase may therefore act directly to enhance the mitogenic effects of growth factors, such as PDGF-BB on lesion SMCs. In line with a role of triglyceride hydrolysis in atherosclerosis are recent studies in which lipoprotein lipase in macrophages has been shown to contribute to atherosclerosis [66].
SMC proliferation is an important step in the progression of fatty streaks to fibroatheromas. This step is likely to convert a reversible fatty streak to an irreversible lesion. We have shown that hyperglycaemia [2] and AGEs have little direct effect on human SMC proliferation. We therefore propose that hyperglycaemia and AGEs can contribute to lesion initiation and progression by affecting the endothelium and monocytes/macrophages, but do not contribute directly to SMC proliferation. Instead, increased levels of OA-containing triglycerides associated with diabetes could contribute to SMC proliferation and accumulation in these lesions.
Abbreviations
- CML:
-
Nε-(carboxymethyl)lysine
- DAG:
-
diacylglycerol
- EA:
-
elaidic acid
- LA:
-
linoleic acid
- MALDI-TOF:
-
matrix assisted laser desorption/ionization time of flight
- MGO:
-
methylglyoxal
- OA:
-
oleic acid
- PDGF-BB:
-
platelet-derived growth factor B-chain homodimer
- PLD:
-
phospholipase D
- RAGE:
-
receptor for advanced glycation end-products
- SA:
-
stearic acid
- SMC:
-
smooth muscle cell
- TNBS:
-
2,4,6-trinitrobenzenesulfonic acid
- TNF-α:
-
tumour necrosis factor-α
- VCAM-1:
-
vascular cell adhesion molecule-1
References
Donahue RP, Orchard TJ (1992) Diabetes mellitus and macrovascular complications. An epidemiological perspective. Diabetes Care 15:1141–1155
Suzuki L, Poot M, Gerrity RG, Bornfeldt KE (2001) Diabetes accelerates smooth muscle accumulation in lesions of atherosclerosis: lack of direct growth promoting effects of high glucose levels. Diabetes 50:851–860
Singh R, Barden A, Mori T, Beilin L (2001) Advanced glycation end-products: a review. Diabetologia 44:129–146
Kume S, Takeya M, Mori T, Araki N, Suzuki H, Horiuchi S, Kodama T, Miyauchi Y, Takahashi K (1995) Immunohistochemical and ultrastructural detection of advanced glycation end products in atherosclerotic lesions of human aorta with a novel specific monoclonal antibody. Am J Pathol 147:654–667
Vlassara H, Li YM, Imani F, Wojciechowicz D, Yang Z, Liu F, Cerami A (1995) Identification of galectin-3 as a high-affinity binding protein for advanced glycation end products (AGE): a new member of AGE-receptor complex. Mol Med 1:634–646
Li YM, Mitsuhashi T, Wojciechowicz D, Shimizu N, Li J, Stitt A, He C, Banerjee D, Vlassara H (1996) Molecular identity and cellular distribution of advanced glycation endproduct receptors: relationship of p60 to OST-48 and p90 to 80K-H membrane proteins. Proc Natl Acad Sci USA 93:11047–11052
Neeper M, Schmidt AM, Brett J, Yan SD, Wang F, Pan YC, Elliston K, Stern D, Shaw A (1992) Cloning and expression of a cell surface receptor for advanced glycosylation end products of proteins. J Biol Chem 267:14998–15004
Ohgami N, Nagai R, Ikemoto M, Arai H, Kuniyasu A, Horiuchi S, Nakayama H (2001) CD36, a member of the class b scavenger receptor family, as a receptor for advanced glycation end products. J Biol Chem 276:3195–3202
Araki N, Higashi T, Mori T, Shibayama R, Kawabe Y, Kodama T, Takahashi K, Shichiri M, Horiuchi S (1995) Macrophage scavenger receptor mediates the endocytic uptake and degradation of advanced glycation end products of the Maillard reaction. Eur J Biochem 230:408–415
Stern D, Yan SD, Yan SF, Schmidt AM (2002) Receptor for advanced glycation endproducts: a multiligand receptor magnifying cell stress in diverse pathologic settings. Adv Drug Deliv Rev 54:1615–1625
Park L, Raman KG, Lee KJ, Lu Y, Ferran LJ Jr, Chow WS, Stern D, Schmidt AM (1998) Suppression of accelerated diabetic atherosclerosis by the soluble receptor for advanced glycation endproducts. Nat Med 4:1025–1031
Kislinger T, Tanji N, Wendt T, Qu W, Lu Y, Ferran LJ Jr, Taguchi A, Olson K, Bucciarelli L, Goova M, Hofmann MA, Cataldegirmen G, D’Agati V, Pischetsrieder M, Stern DM, Schmidt AM (2001) Receptor for advanced glycation end products mediates inflammation and enhanced expression of tissue factor in vasculature of diabetic apolipoprotein E-null mice. Arterioscler Thromb Vasc Biol 21:905–910
Basta G, Lazzerini G, Massaro M, Simoncini T, Tanganelli P, Fu C, Kislinger T, Stern DM, Schmidt AM, De Caterina R (2002) Advanced glycation end products activate endothelium through signal-transduction receptor RAGE: a mechanism for amplification of inflammatory responses. Circulation 105:816–822
Faergeman O (2000) Hypertriglyceridemia and the fibrate trials. Curr Opin Lipidol 11:609–614
Erkelens DW (1998) Diabetic dyslipidaemia. Eur Heart J 19 (Suppl H):H27–H40
Gerrity RG, Natarajan R, Nadler JL, Kimsey T (2001) Diabetes-induced accelerated atherosclerosis in swine. Diabetes 50:1654–1665
Dixon JL, Shen S, Vuchetich JP, Wysocka E, Sun GY, Sturek M (2002) Increased atherosclerosis in diabetic dyslipidemia swine: protection by atorvastatin involves decreased VLDL triglycerides but minimal effects on the lipoprotein profile. J Lipid Res 43:1618–1629
Pentikainen MO, Oksjoki R, Oorni K, Kovanen PT (2002) Lipoprotein lipase in the arterial wall: linking LDL to the arterial extracellular matrix and much more. Arterioscler Thromb Vasc Biol 22:211–217
Brynes AE, Edwards CM, Jadhav A, Ghatei MA, Bloom SR, Frost GS (2000) Diet-induced change in fatty acid composition of plasma triacylglycerols is not associated with change in glucagon-like peptide 1 or insulin sensitivity in people with type 2 diabetes. Am J Clin Nutr 72:1111–1118
Seigneur M, Freyburger G, Gin H, Claverie M, Lardeau D, Lacape G, Le Moigne F, Crockett R, Boisseau MR (1994) Serum fatty acid profiles in type I and type II diabetes: metabolic alterations of fatty acids of the main serum lipids. Diabet Res Clin Pract 23:169–177
Rybalkin SD, Bornfeldt KE, Sonnenburg WK, Rybalkina IG, Kwak KS, Hanson K, Krebs EG, Beavo JA (1997) Calmodulin-stimulated cyclic nucleotide phosphodiesterase (PDE1C) is induced in human arterial smooth muscle cells of the synthetic, proliferative phenotype. J Clin Invest 100:2611–2621
Hamad NM, Elconin JH, Karnoub AE, Bai W, Rich JN, Abraham RT, Der CJ, Counter CM (2002) Distinct requirements for Ras oncogenesis in human versus mouse cells. Genes Dev 16:2045–2057
Renard CB, Bornfeldt KE (2001) Human arterial smooth muscle cells rapidly deplete cell culture media of glucose. Diabetologia 44:1067–1068
Wautier JL, Paton C, Wautier MP, Pintigny D, Abadie E, Passa P, Caen JP (1981) Increased adhesion of erythrocytes to endothelial cells in diabetes mellitus and its relation to vascular complications. N Engl J Med 305:237
Kislinger T, Fu C, Huber B, Qu W, Taguchi A, Du Yan S, Hofmann M, Yan SF, Pischetsrieder M, Stern D, Schmidt AM (1999) Nε-(carboxymethyl)lysine adducts of proteins are ligands for receptor for advanced glycation end products that activate cell signaling pathways and modulate gene expression. J Biol Chem 274:31740–31749
Westwood ME, Argirov OK, Abordo EA, Thornalley PJ (1997) Methylglyoxal-modified arginine residues—a signal for receptor-mediated endocytosis and degradation of proteins by monocytic THP-1 cells. Biochim Biophys Acta 1356:84–94
Habeeb A (1966) Determination of free amino groups in proteins by trinitrobenzenesulfonic acid. Anal Biochem 14:328–336
Hamilton JA, Kamp F (1999) How are free fatty acids transported in membranes? Is it by proteins or by free diffusion through the lipids? Diabetes 48:2255–2269
Smith EB, Staples EM (1980) Distribution of plasma proteins across the human aortic wall-barrier functions of endothelium and internal elastic lamina. Atherosclerosis 37:579–590
Rabinovitch PS (1994) DNA content histogram and cell-cycle analysis. Methods Cell Biol 41:263–296
Zapolska-Downar D, Zapolski-Downar A, Markiewski M, Ciechanowicz A, Kaczmarczyk M, Naruszewicz M (2000) Selective inhibition by α-tocopherol of vascular cell adhesion molecule-1 expression in human vascular endothelial cells. Biochem Biophys Res Commun 274:609–615
Bligh E, Dyer W (1959) A rapid method of total lipid extraction and purification. Can J Biochem Physiol 37:911–917
Lepage G, Roy CC (1986) Direct transesterification of all lipids in a one step reaction. J Lipid Res 27:114–120
Satia JA, Patterson RE, Herrero R, Jin F, Dai Q, King IB, Chen C, Kristal AR, Prentice RL, Rossing MA (1999) Study of diet, biomarkers and cancer risk in the United States, China and Costa Rica. Int J Cancer 82:28–32
Thornalley PJ (2002) Glycation in diabetic neuropathy: characteristics, consequences, causes, and therapeutic options. Int Rev Neurobiol 50:37–57
Degenhardt TP, Thorpe SR, Baynes JW (1998) Chemical modification of proteins by methylglyoxal. Cell Mol Biol 44:1139–1145
Thornalley PJ, Battah S, Ahmed N, Karachalias N, Agalou S, Babaei-Jadidi R, Dawnay A (2003) Quantitative screening of advanced glycation endproducts in cellular and extracellular proteins by tandem mass spectrometry. Biochem J 375:581–592
Reddy S, Bichler J, Wells-Knecht K, Thorpe S, Baynes J (1995) Carboxymethyllysine is a dominant AGE antigen in tissue proteins. Biochemistry 34:10872–10878
Mizutani K, Ikeda K, Yamori Y (2000) Resveratrol inhibits AGEs-induced proliferation and collagen synthesis activity in vascular smooth muscle cells from stroke-prone spontaneously hypertensive rats. Biochem Biophys Res Commun 274:61–67
Wang R, Kudo M, Yokoyama M, Asano G (2001) Roles of advanced glycation endproducts (AGE) and receptor for AGE on vascular smooth muscle cell growth. J Nippon Med Sch 68:472–481
Satoh H, Togo M, Hara M, Miyata T, Han K, Maekawa H, Ohno M, Hashimoto Y, Kurokawa K, Watanabe T (1997) Advanced glycation endproducts stimulate mitogen-activated protein kinase and proliferation in rabbit vascular smooth muscle cells. Biochem Biophys Res Commun 239:111–115
Sakata N, Meng J, Takebayashi S (2000) Effects of advanced glycation end products on the proliferation and fibronectin production of smooth muscle cells. J Atheroscler Thromb 7:169–176
Higashi T, Sano H, Saishoji T, Ikeda K, Jinnouchi Y, Kanzaki T, Morisaki N, Rauvala H, Shichiri M, Horiuchi S (1997) The receptor for advanced glycation end products mediates the chemotaxis of rabbit smooth muscle cells. Diabetes 46:463–472
Iino K, Yoshinari M, Yamamoto M, Kaku K, Doi Y, Ichikawa K, Iwase M, Fujishima M (1996) Effect of glycated collagen on proliferation of human smooth muscle cells in vitro. Diabetologia 39:800–806
Sakaguchi T, Yan SF, Yan SD, Belov D, Rong LL, Sousa M, Andrassy M, Marso SP, Duda S, Arnold B, Liliensiek B, Nawroth PP, Stern DM, Schmidt AM, Naka Y (2003) Central role of RAGE-dependent neointimal expansion in arterial restenosis. J Clin Invest 111:959–972
Wautier MP, Chappey O, Corda S, Stern DM, Schmidt AM, Wautier JL (2001) Activation of NADPH oxidase by AGE links oxidant stress to altered gene expression via RAGE. Am J Physiol Endocrinol Metab 280:E685–E694
Sengoelge G, Fodinger M, Skoupy S, Ferrara I, Zangerle C, Rogy M, Horl WH, Sunder-Plassmann G, Menzel J (1998) Endothelial cell adhesion molecule and PMNL response to inflammatory stimuli and AGE-modified fibronectin. Kidney Int 54:1637–1651
Kirstein M, Brett J, Radoff S, Ogawa S, Stern D, Vlassara H (1990) Advanced protein glycosylation induces transendothelial human monocyte chemotaxis and secretion of platelet-derived growth factor: role in vascular disease of diabetes and aging. Proc Natl Acad Sci USA 87:9010–9014
Miyata T, Inagi R, Iida Y, Sato M, Yamada N, Oda O, Maeda K, Seo H (1994) Involvement of beta 2-microglobulin modified with advanced glycation end products in the pathogenesis of hemodialysis-associated amyloidosis. Induction of human monocyte chemotaxis and macrophage secretion of tumor necrosis factor-alpha and interleukin-1. J Clin Invest 93:521–528
Lu G, Meier KE, Jaffa AA, Rosenzweig SA, Egan BM (1998) Oleic acid and angiotensin II induce synergistic mitogenic response in vascular smooth muscle cells. Hypertension 31:978–985
Kwok CF, Shih KC, Hwu CM, Ho LT (2000) Linoleic acid and oleic acid increase the endothelin-1 binding and action in cultured rat aortic smooth muscle cells. Metabolism 49:1386–1389
Askari B, Carroll MA, Capparelli M, Kramer F, Gerrity RG, Bornfeldt KE (2002) Oleate and linoleate enhance the growth-promoting effects of insulin-like growth factor-I through a phospholipase D-dependent pathway in arterial smooth muscle cells. J Biol Chem 277:36338–36344
Lu X, Yang XY, Howard RL, Walsh JP (2000) Fatty acids modulate protein kinase C activation in porcine vascular smooth muscle cells independently of their effect on de novo diacylglycerol synthesis. Diabetologia 43:1136–1144
Inui H, Kitami Y, Tani M, Kondo T, Inagami T (1994) Differences in signal transduction between platelet-derived growth factor (PDGF) alpha and beta receptors in vascular smooth muscle cells. PDGF-BB is a potent mitogen, but PDGF-AA promotes only protein synthesis without activation of DNA synthesis. J Biol Chem 269:30546–30552
Shome K, Rizzo MA, Vasudevan C, Andresen B, Romero G (2000) The activation of phospholipase D by endothelin-1, angiotensin II, and platelet-derived growth factor in vascular smooth muscle A10 cells is mediated by small G proteins of the ADP-ribosylation factor family. Endocrinology 141:2200–2208
Bornfeldt KE, Raines EW, Nakano T, Graves LM, Krebs EG, Ross R (1994) Insulin-like growth factor-I and platelet-derived growth factor-BB induce directed migration of human arterial smooth muscle cells via signaling pathways that are distinct from those of proliferation. J Clin Invest 93:1266–1274
Du X, Jiang Y, Qian W, Lu X, Walsh JP (2001) Fatty acids inhibit growth-factor-induced diacylglycerol kinase α activation in vascular smooth-muscle cells. Biochem J 357:275–282
Sánchez-Pinera P, Micol V, Corbalan-Garcia S, Gomez-Fernandez JC (1999) A comparative study of the activation of protein kinase Cα by different diacylglycerol isomers. Biochem J 337:387–395
Yu HY, Inoguchi T, Kakimoto M, Nakashima N, Imamura M, Hashimoto T, Umeda F, Nawata H (2001) Saturated non-esterified fatty acids stimulate de novo diacylglycerol synthesis and protein kinase c activity in cultured aortic smooth muscle cells. Diabetologia 44:614–620
Singer P, Gnauck G, Honigmann G, Schliack V, Laeuter J (1977) The fatty acid composition of triglycerides in arteries, depot fat and serum of amputated diabetics. Atherosclerosis 28:87–92
Bohov P, Baláz V, Seböková E, Klimes I (1997) The effect of hyperlipidemia on serum fatty acid composition in type 2 diabetics. Ann NY Acad Sci 827:561–567
Kelley DE, Simoneau JA (1994) Impaired free fatty acid utilization by skeletal muscle in non-insulin-dependent diabetes mellitus. J Clin Invest 94:2349–2356
Schwertner HA, Mosser EL (1993) Comparison of lipid fatty acids on a concentration basis vs weight percentage basis in patients with and without coronary artery disease or diabetes. Clin Chem 39:659–663
Lecomte M, Claire M, Deneuville M, Wiernsperger N (1998) Fatty acid composition of phospholipids and neutral lipids from human diabetic small arteries and veins by a new TLC method. Prostaglandins Leukot Essent Fatty Acids 59:363–369
Sartippour MR, Lambert A, Laframboise M, St-Jacques P, Renier G (1998) Stimulatory effect of glucose on macrophage lipoprotein lipase expression and production. Diabetes 47:431–438
Babaev VR, Patel MB, Semenkovich CF, Fazio S, Linton MF (2000) Macrophage lipoprotein lipase promotes foam cell formation and atherosclerosis in low density lipoprotein receptor-deficient mice. J Biol Chem 275:26293–26299
Acknowledgements
We thank A. Wiczulis and A.M. Sherrid for excellent technical assistance during part of this study. We are grateful to I.B. King and F. Boric at the Fred Hutchinson Cancer Research Center, Seattle, for expert help in determining fatty acid composition of DAG. This work was supported by a grant from the National Institutes of Health, HL62887, an Innovation Award from the American Diabetes Association, and a grant from the Juvenile Diabetes Research Foundation International, 1-2003-33, to K.E.B. B.A. is supported by National Institutes of Health training grant T32 HL07312.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Renard, C.B., Askari, B., Suzuki, L.A. et al. Oleate, not ligands of the receptor for advanced glycation end-products, promotes proliferation of human arterial smooth muscle cells. Diabetologia 46, 1676–1687 (2003). https://doi.org/10.1007/s00125-003-1247-9
Received:
Revised:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00125-003-1247-9