
Abstract
The basal non-mammaliaform cynodonts from the late Permian (Lopingian) and Early Triassic are a major source of information for the understanding of the evolutionary origin of mammals. Detailed knowledge of their anatomy is critical for understanding the phylogenetic transition toward mammalness and the paleobiological reconstruction of mammalian precursors. Using micro-computed tomography (μCT), we describe the internal morphology of the interorbital region that includes the rarely fossilized orbitosphenoid elements in four basal cynodonts. These paired bones, which are positioned relatively dorsally in the skull, contribute to the wall of the anterior part of the braincase and form the floor for the olfactory lobes. Unlike procynosuchids and the more basal therapsids in which the orbitosphenoids are well developed, dense, and bear a ventral keel, the basal epicynodonts Cynosaurus, Galesaurus, and Thrinaxodon display cancellous, reduced, and loosely articulated orbitosphenoids, a condition shared with many eucynodonts. The hemi-cylindrical orbitosphenoid from which the mammalian condition is derived re-evolved convergently in traversodontid and some probainognathian cynodonts.
Similar content being viewed by others


Introduction
The late Permian (Lopingian)/Early Triassic cynodonts Procynosuchus delaharpeae, Cynosaurus suppostus, Galesaurus planiceps, and Thrinaxodon liorhinus (along with a handful of less abundant taxa) are the basal-most offshoots of the Cynodontia, the clade including Mammalia and their ancestors (Hopson and Kitching 2001; Liu and Olsen 2010; Ruta et al. 2014). As basal members of the cynodont clade, they are important for understanding the emergence of mammalian characters as well as the paleobiology of mammalian forerunners (Brink 1959; Rubidge and Sidor 2001; Abdala 2003; Kemp 2005; Fernandez et al. 2013; Jasinoski et al. 2015; Benoit et al. 2015, 2016a). Procynosuchus, Cynosaurus, Galesaurus, and Thrinaxodon are found in the South African Beaufort Group of the Karoo Supergroup, which certainly is the richest fossiliferous area documenting the evolution of non-mammaliaform (NM) therapsids (Rubidge and Sidor 2001). Unlike other basal cynodonts, they are abundant and with a good representation of cranial material, which makes them even more critical for any discussion about the evolution of NM cynodonts and the ancestry of mammals. This is particularly true for Thrinaxodon, which has a more widespread distribution and is represented by over a hundred well-preserved specimens (Brink 1959; Estes 1961; Fourie 1974; Colbert and Kitching 1977; Fernandez et al. 2013; Abdala et al. 2013; Jasinoski et al. 2015; Benoit et al. 2016a). Therefore, thorough knowledge of basal cynodont anatomy is crucial for unfolding the radiation of NM cynodonts and the deep evolutionary root of mammals.
The orbitosphenoid in mammals is an endochondral bone that forms the lateral wall and floor of the anterior braincase supporting the anterior-most part of the brain. This element is usually cartilaginous in extant sauropsids (de Beer 1985); however, it is present in some squamates (e.g., Lacerta, Shinisaurus), as well as in archosaurs (i.e., dinosaurs, including birds, and crocodiles) due to the ossification of the metotic pila (de Beer 1985; Currie 1997; Bever et al. 2005). In mammals, this bone is an independent element that ossifies on the orbital cartilage from the nasal capsule to the proximity of the auditory capsule (de Beer 1985). It shows different degrees of fusion with the remaining sphenoidal elements. In mammaliaforms, the orbitosphenoid is always present and ventrally closes the anterior part of the braincase participating of the orbital mosaic (Kielan-Jaworowska et al. 2004). A discrete orbitosphenoid has also been documented in most NM therapsids (e.g., Olson 1938; Boonstra 1968; Kemp 1969; Sigogneau 1970; Cluver 1971; Kemp 1972; Fourie 1993; Kielan-Jaworowska et al. 2004; Sidor and Smith 2007; Sigurdsen et al. 2012; Castanhinha et al. 2013; Laaß 2015; Benoit et al. 2016b; Crompton et al. 2017). However, its presence has never been formally recognized in the basal epicynodonts Cynosaurus, Galesaurus, and Thrinaxodon, even though numerous skulls have been described. For instance, despite the large number of specimens investigated using computed tomography (CT) scans (e.g., Rowe et al. 1995; Jasinoski et al. 2015), serial-sectioning (e.g., Broom 1938; Fourie 1974), and gross anatomical observations (e.g., Estes 1961), there is no unequivocal report of the orbitosphenoid in the endocranial cavity of Thrinaxodon. This would make the condition in basal cynodonts more similar to that of sauropsids than to mammals and other NM therapsids. This presumed absence of the orbitosphenoid in the ancestors of mammaliaforms is paradoxical and has important phylogenetic implications (Luo 1994).
Here, we describe in detail the previously unrecognized or neglected orbitosphenoid of Cynosaurus, Galesaurus, and Thrinaxodon using micro-computed tomography (μCT) scanning.
Material and methods
The μCT technique is a non-destructive technique that allows the observation of the internal structures of fossil skulls and the in silico reconstruction of otherwise inaccessible anatomical details. This technique and regular comparative anatomy was used on 51 NM therapsid skulls (Online Resource 1). The majority of specimens were μCT scanned at the Evolutionary Studies Institute (University of the Witwatersrand, South Africa) using a Nikon Metrology XTH 225/320 LC dual-source CT system. Two specimens were scanned using the 225 LC at DebTech (DeBeers, Johannesburg, ZA) (Online Resource 1). Thrinaxodon specimens BPI/1/7199 and BP/1/5905 and Microgomphodon specimen SAM-PK-10160 were scanned at the European Synchrotron Radiation Facility, Grenoble (see Fernandez et al. 2013 for details of Thrinaxodon). Three-dimensional renderings were obtained using manual segmentation under Avizo 8 (FEI VSG, Hillsboro OR, USA). We also used published descriptions and serial sections from the literature for comparison (see Online Resource 1).
To reconstruct the evolution of the morphology of the orbitosphenoid throughout therapsid phyogeny, we applied a parsimony model of ancestral state reconstruction on a character matrix created for this study (Online Resource 2) under Mesquite 2.75 and 3.2 (Maddison and Maddison 2009). The phylogenetic tree used for the reconstruction of ancestral states is based on Sigurdsen et al. (2012) and Ruta et al. (2014) for therocephalian and NM cynodont, respectively. The tree is rooted with Gorgonopsia that show little variation in the gross morphology of their orbitosphenoid (see below).
Institutional abbreviations
BP, Evolutionary Studies Institute (formerly Bernard Price Institute for Palaeontological Research), University of the Witwatersrand, Johannesburg, South Africa; NMQR, National Museum, Bloemfontein, South Africa; RC, Rubidge Collection, Wellwood, Graaff Reinet, South Africa; SAM, Iziko South African Museum, Cape Town, South Africa.
Results and description
In the gorgonopsian Aelurosaurus, the orbitosphenoids are fused ventrally into a long and hemi-cylindrical bone that encapsulates a tubular braincase (Fig. 1a). The orbitosphenoid is V-shaped in cross section, thin, and not cancellous (Fig. 1a). This bone is articulated with the mesethmoid rostrally and with the interorbital septum ventrally (Fig. 1a). These characters represent the plesiomorphic condition for cynodonts according to the ancestral character state reconstruction (Fig. 2).

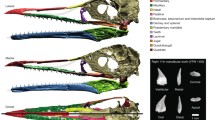
Digital reconstruction and μCT section of the orbitosphenoids in a gorgonopsian and some basal cynodonts. From left to right digital reconstruction in anterior, dorsal, and lateral views; the position of the orbitosphenoid on an oblique view of the skull (translucent); and a μCT cross section of the orbitosphenoid. Solid arrows show the orbitosphenoid bones; dotted arrow shows the ventral keel. Scale bar = 10 mm

Results of the ancestral character state reconstruction (based on the character matrix in Online Resource 2). Names of the clades are indicated on the tree on the bottom right corner of the figure
Orbitosphenoids are present in five of the seven Procynosuchus specimens that were examined (BP/1/650, BP/1/3747, RC 5, BP/1/3748, and RC 92). The paired orbitosphenoids loosely articulate with the frontals that form the roof of the endocranial cavity, and in some cases, articulate with each other ventromedially (Fig. 1b). The bones are shorter anteroposteriorly than in Aelurosaurus, thin, and do not have large cancellous spaces (Fig. 1b). A ventral keel only is demonstrably present on specimen RC 92 (Fig. 3), whereas a complete interorbital septum separating the internal cranial cavity medially appears to be present in two other specimens (BP/1/650, BP/1/3747; Online Resource 1). Specimen BP/1/3748 noticeably shows no evidence of an interorbital septum or fusion between the two orbitosphenoids (Fig. 1b), two characters that are reconstructed as apomorphic for Epicynodontia (Fig. 2e). According to the results of the ancestral character state analysis, Procynosuchus is either plesiomorphic or variable for most characters of the orbitosphenoid (Fig. 2). It nevertheless shares with epicynodonts a short orbitosphenoid and the absence of direct (bony) articulation with the cranial roof (Fig. 2a, d).

The phylogeny and evolution of the orbitosphenoid in Cynodontia (μCT photo of specimens in cross section). 1 anteroposteriorly short and loosely articulated orbitosphenoids (at least in some specimens), direct (bony) articulation with the cranial vault lost, ventral keel and interorbital septum variably present; 2 cancellous bone present, ventral keel absent, interorbital septum absent (large interorbital vacuity); 3 large fused orbitosphenoid articulated with the cranial vault, ventral keel present; 4 anteroposteriorly elongated orbitosphenoid, interorbital vacuity closed. Rdg ridge, Spt interorbital septum, Sed sediment filling. Solid arrow, orbitosphenoid; dotted arrow, ventral keel. Specimens (top to bottom, left to right) BP/1/3776, BP/1/4658, BP/1/4245, BP/1/1669, BP/1/4778, BP/1/1563, BP/1/10468, BP/1/7199, RC 92, BP/1/3849, SAM-PK-10160, BP/1/216. Images taken using Avizo 8 (FEI VSG, Hillsboro OR, USA). Not to scale. Phylogeny after Ruta et al. (2014)
A paired ossification was discovered in four of the μCT scanned specimens of Cynosaurus, but it was in situ in only two specimens, SAM-PK-4333 and BP/1/1563 (Fig. 1c). In these specimens, the ossification consisted of two thin plate-like structures that appear to articulate ventromedially (Fig. 1c). The cross section of the structure is U-shaped (Fig. 1c). It is located ventral to the frontals but does not directly articulate with the cranial roof (a connection was likely maintained by soft tissues). In the smallest specimen of Cynosaurus (BP/1/4469), the two bones are separated and loose inside the endocranial space, which suggests that the fusion between the two orbitosphenoids did not always occur. They are thin and dense posteriorly but they become slightly thicker and more cancellous anteriorly (Fig. 1c). In both SAM-PK-4333 and BP/1/1563, the cancellous bone is filled with metallic nodules (Fig. 1c).
Disarticulated orbitosphenoid bones were observed inside several skulls of Galesaurus (RC 845, NMQR 135, NMQR 3542, displaced but not far from in situ position in NMQR 860; see Online Resource 1), but it is in situ only in SAM-PK-K10468 (Fig. 1d). In cross section, these narrow curved bones form a U-shaped structure similar to that in Cynosaurus (Fig. 1d), but the paired bones do not contact each other ventrally and so the U-shaped structure remains mostly open ventrally. The bones are completely filled with cancellous spaces. In SAM-PK-K10468, the bones are in close contact with the ventral surface of the frontal, but they only loosely articulate (Fig. 1d). Noticeably, the space that separates the ventral margins of both orbitosphenoids appears smaller caudally in Galesaurus, whereas in Procynosuchus (BP/1/3748) and Cynosaurus, this separation is smaller rostrally (Fig. 1b–d).
Here, for the first time, we report the definitive presence of a paired ossification in three specimens of Thrinaxodon. In specimen BP/1/7199, there are two ex situ bones that form a U-shaped structure (Fig. 1e), like in Galesaurus and Cynosaurus. The bones are slightly displaced relative to each other (Fig. 1e), and in life would not have articulated ventromedially. The orbitosphenoids are thick and cancellous, similar to Galesaurus (Fig. 1). In specimen BP/1/4263, two curved and cancellous detached bones are preserved near the dorsal part of the pterygoids (Fig. 1f). Specimen BP/1/5372 also displays one possible isolated and displaced orbitosphenoid (Online Resource 1). In addition, an incomplete Thrinaxodon skull (UCMP 42880), broken at the level of the postorbital bar, revealed an extremely thin bone continuous with the frontal on the left side that might represent an isolated orbitosphenoid (Online Resource 1).
Our survey found that Cynosaurus, Galesaurus, and Thrinaxodon display the apomorphic conditions of orbitosphenoid morphology that characterize Epicynodontia. The only exception is the ventral articulation of the orbitosphenoids, which is retained in Cynosaurus, but not in the other two taxa (Fig. 2b). However, most of these characters then reverse back to their plesiomorphic state at some point in the Probainognathia and some Cynognathia (Fig. 2).
Discussion
The orbitosphenoids, here described for the first time in Cynosaurus, Galesaurus, and Thrinaxodon, tend to become displaced during fossilization, which can make their identification difficult (Fig. 1). Fortunately, the paired ossification is located in situ in one specimen of Cynosaurus (AM 4947) and Galesaurus (SAM-PK-K10468) (Fig. 1c, d). The short hemi-cylindrical structure they form in the anterior-most part of the braincase, below the frontal, resembles the paired bones that support the tubular forebrain in Procynosuchus, and other NM therapsids (Figs. 1a, b and 3 and Online Resource 1). This paired ossification is unequivocally identified as the orbitosphenoid in Cynognathia (von Huene 1936; Brink 1955; Bonaparte 1962; Crompton et al. 2015), and Probainognathia (Crompton 1958, 1964; Bonaparte and Barberena 2001; Bonaparte et al. 2003; Kemp 2009; Soares et al. 2011), including tritylodontids (Kühne 1956; Hopson 1964; Sun 1984; Sues 1986; Gow 1986; Kielan-Jaworowska et al. 2004; Liu and Olsen 2010), and Brasilodon (Bonaparte et al. 2003, 2005).
Except for two reports in procynosuchids (BP/1/1821; Brink 1961, 1963), orbitosphenoid bones have not been described for any basal cynodont, including the abundant and thoroughly studied Thrinaxodon. Thus, when examining the sister group relationships of mammaliaforms, Luo (1994) considered the orbitosphenoid as unossified in Thrinaxodontidae. There are, nevertheless, equivocal reports of the presence of orbitosphenoid bones in basal cynodonts. Broom (1938, Fig. 2, Sect. 11) figured two unlabeled fragments of thin bone occurring posterior to the orbits and ventral to the frontal bones on a serial section of Thrinaxodon. It is possible that the longer of the two bones he interpreted as a sclerotic plate, a structure never corroborated in hundreds of specimens of Thrinaxodon, might represent an orbitosphenoid because it has a similar shape and position as those reported here (Fig. 1e, f).
Haughton (1918, p. 201) briefly described in Cynosaurus (SAM-PK-4333) two curved ossifications located close to each other rostrally and then separating caudally identified as “crushed sphenoids or orbitosphenoids.” His associated figure (Haughton 1918, Fig. 53c) of a V-shaped structure in cross section comprised of two distinct bones in close contact but not fused ventrally indeed corresponds to the morphology of the orbitosphenoids observed here in the μCT scanned Cynosaurus specimen BP/1/4469 (Online Resource 1).
The fact that the orbitosphenoid remained unrecognized in most basal epicynodonts for so long might be explained in three ways. First, there is a lack of thorough description of the braincase of adult specimens, based either on CT scans or serial sections (with the exception of Thrinaxodon). Second, the postmortem disarticulation of orbitosphenoids, in which they are found “floating” within the endocranial cavity, can lead to their misidentification or a lack of recognition (e.g., Haughton 1918; Broom 1938). Lastly, the orbitosphenoid in basal epicynodonts tends to be a thin bone with a cancellous structure, which might make it difficult to differentiate it from the surrounding sediment during preparation of specimens, leading to its destruction. This last point illustrates the importance of non-destructive μCT investigation prior to fossil preparation. These three explanations might also account for inconsistencies in the interpretation of the presence of this bone in other basal cynodont taxa such as Dvinia (see Ivakhnenko 2013, contra Tatarinov 1974).
Evolution of the orbitosphenoid in NM cynodonts
A ventral ossification of the anterior-most part of the braincase is absent in “pelycosaurs,” and the homology of the orbitosphenoid with one or more of the bones of the sphenethmoid region in these basal taxa is still not certain (Romer and Price 1940; Hopson 1979; Kielan-Jaworowska et al. 2004). Aside from cynodonts, the orbitosphenoid is known in representatives of all the other NM therapsid lineages such as the biarmosuchians (=sphenethmoid; Sidor and Smith 2007; Day et al. 2016), dinocephalians (Boonstra 1968; Benoit et al. 2016b), anomodonts (Cluver 1971; Fourie 1993; Castanhinha et al. 2013; Laaß 2015), gorgonopsians (Olson 1938; Kemp 1969; Sigogneau 1970), and therocephalians (Kemp 1972; Sigurdsen et al. 2012; Benoit et al. 2016b). In all of these taxa, the orbitosphenoid bones appear to be fused ventrally forming a long and hemi-cylindrical V-shaped structure in cross section (Figs. 1a and 3). This structure itself is indistinguishably fused to the mesethmoid rostrally (the resulting bone is often called the anterior plate or septosphenoid in anomodonts; Boonstra 1968; Cluver 1971; Castanhinha et al. 2013) and to the interorbital septum ventrally (Fig. 3). Together with the epipterygoid, they form the sphenethmoidal complex (Kielan-Jaworowska et al. 2004). The dorsal margins articulate firmly with the frontals, except in some therocephalians where the orbitosphenoids are often loose in the endocranial cavity (Benoit et al. 2016b; Fig. 3 and Online resource 1). The orbitosphenoid is not cancellous, except in dinocephalians (Boonstra 1968; Benoit et al. 2016b), some dicynodonts (Laaß 2015), and the therocephalians Hofmeyria and Olivierosuchus (Fig. 3 and Online Resource 1). According to the ancestral character state reconstruction (Fig. 2), anteroposterioly elongated, paired, and dense orbitosphenoids, attached dorsally to the cranial roof and fused ventrally with the interorbital septum via a prominent ventral keel, is the plesiomorphic condition of Cynodontia (Fig. 2).
In procynosuchids, the orbitosphenoids are not closely connected to the ventral surface of the frontal and parietal bones (Brink 1961, 1963; Figs. 1b and 3), like in some therocephalians and other NM cynodonts (Fig. 2d). Thus, they can easily be lost during the fossilization process or being mistakenly identified as unimportant loose bones and destroyed during preparation. This may explain the absence of any ventral ossification of the forebrain in the otherwise thorough cranial description of a Zambian Procynosuchus specimen (Kemp 1979). Nevertheless, the orbitosphenoids are in situ in four out of seven of our specimens and still form a U-shaped structure (Figs. 1b and 3 and Online Resource 1), which suggests that the soft-tissue connection to the underside of the skull roof was quite strong. In BP/1/3748, the paired orbitosphenoids seem to articulate loosely with each other, like in epicynodonts (Haughton 1918; Figs. 1b and 2b), but they are thin and devoid of cancellous spaces like in non-cynodont therapsids (Figs. 1b, 2c, and 3). The presence/absence of a ventral keel and its articulation with the interorbital septum remain unclear in Procynosuchus because of the considerable variation of these characters (Figs. 1b, 2e, and 3 and Online Resource 1; Brink 1963). Perhaps this is the result of the cartilaginous nature or partial mineralization of the interorbital septum in Procynosuchus. Therefore, in many aspects, the orbitosphenoid of Procynosuchus appears to be intermediate between the anatomy observed in therocephalians and those of the basal epicynodonts (Figs. 2 and 3, node 1).
Most NM epicynodonts depart from the plesiomorphic conditions described in Procynosuchus because their orbitosphenoids never display a ventral keel and are composed of delicate bone filled with large cancellous spaces (Figs. 2c, e and 3, node 2). As the basal-most epicynodont of this study, Cynosaurus display cancellous spaces only in the anterior part of the orbitosphenoid. In addition, the interobital septum is absent in most epicynodont taxa (Fig. 2e). The orbitosphenoids of NM epicynodonts form a hemi-cylindrical structure that is anteroposteriorly shorter than in other, more basal NM therapsids (Figs. 1 and 2a). The structure is U-shaped in cross section, and opens ventrally as a result of the absence of articulation between the paired orbitosphenoids, except in some specimens of Cynosaurus, in which both orbitosphenoids remain articulated (Figs. 1c–f, 2b, and 3). The absence of ossification of the interorbital septum leaves a large interorbital vacuity between the orbits (Kielan-Jaworowska et al. 2004). As a consequence, orbitosphenoids are often found loose in the endocranial cavity or disarticulated, especially in basal epicynodonts (Figs. 1c, e, f and 3). The presence of in situ orbitosphenoids, at least in some specimens, suggests that they were supported by the cartilaginous interorbital septum and walls of braincase (a piece of a possible interorbital septum is ossified in a single Galesaurus specimen AMNH 2223; Online Resource 1).
Among the Cynognathia, some Late Triassic traversodontids such as Massetognathus and Exaeretodon re-developed a strong attachment to the cranial vault and a ventral keel below the elongated and medially fused orbitosphenoids (Figs. 2b, d and 3, mark 3; von Huene 1936; Brink 1955; Bonaparte 1962; Crompton et al. 2015). There is no visible interorbital septum, which still leaves the large interorbital vacuity widely opened, but a prominent ridge, produced by the palatine, is present on the floor of the vacuity (Fig. 3), suggesting the presence of a cartilaginous septum.
As in basal epicynodonts, an anteroposteriorly short and cancellous orbitosphenoid, and a large interorbital vacuity is maintained in the Probainognathia Lumkuia, Prozostrodon, Tritheledontidae, Brasilodon, and possibly Therioherpeton (Figs. 2 and 3; Hopson 1964; Kielan-Jaworowska et al. 2004; Bonaparte and Barberena 2001; Bonaparte et al. 2003, 2005; Soares et al. 2011). On the contrary, a long hemi-cylindrical orbitosphenoid with a ventral keel resembling that of Massetognathus and Exaeretodon is present in Chiniquodon theotonicus (Kemp 2009), Probainognathus (Crompton et al. 2017) (though the bone is rarely preserved and, when present, displays variable anatomy in this taxon; see, e.g., Romer 1970; Kielan-Jaworowska et al. 2004; Crompton et al. 2017), and possibly in Ecteninion (Rowe 2002, Digimorph.org; note that the exact boundaries of the orbitosphenoid are difficult to trace on the CT scan; Martinez et al. 1996), which suggests that the mineralization of the interorbital septum among Probainognathia was likely subject to variability and homoplasy (Fig. 2). Unlike other cynodonts, the orbitosphenoid, when present, is always articulated to the frontal in Probainognathia and the two Traversodontidae represented in our sample (Fig. 2d).
In tritylodontids, the orbitosphenoid contacts the palatine rostroventrally and is expanded caudally so that it contacts the epipterygoid posterodorsally and the prootic posteroventrally, which results in the loss of the interorbital vacuity and the consequent development of an interorbital mosaic formed by the orbitosphenoid, palatine, and frontal (Figs. 2e and 3, node 4; Hopson 1964; Sun 1984; Sues 1986; Gow 1986; Kielan-Jaworowska et al. 2004). This condition is essentially identical to that in stem mammaliaforms and represents a significant departure from that represented in other NM cynodonts (Hopson 1964; Sun 1984; Sues 1986; Gow 1986; Kielan-Jaworowska et al. 2004). In tritylodontids, the orbitosphenoids still form a long and tubular structure, but in early mammaliaforms, they become divergent and separated medially by the presphenoid as the brain hemispheres grow larger (Kermack et al. 1981; Kielan-Jaworowska et al. 2004; Rowe et al. 2011; Rowe 2002, Digimorph.org). In Brasilitherium, the orbitosphenoid as interpreted by Ruf et al. (2014, Fig. 5a) on CT scans images, covers the orbital lamina of the lacrimal medially, and does not contribute to the braincase. However, the peculiar location of the bone is probably due to a postmortem displacement (Crompton et al. 2017). This is supported by (i) the position of the orbitosphenoid in Brasilodon, the sister taxon of Brasilitherium, in which it is located in its regular place (Bonaparte et al. 2003, 2005), and (ii) the ancestral character state analysis that reconstructs the condition in Brasilitherium as being similar to that in tritylodontids and mammaliaforms (Fig. 2e).
As evidenced from the discussion above, eucynodonts show a wide range of variation in the pattern of ossification of the orbitosphenoid and they even tend to re-develop the plesiomorphic condition of an anteroposteriorly elongated and fused orbitosphenoid with a ventral keel in a convergent manner in traversodontids and some probainognathians (Figs. 2 and 3). The retention of a cartilaginous support for the orbitosphenoid in basal epicynodonts probably provided the basis for this homoplasic evolution (Fig. 3). This transition from a developed orbitosphenoid in procynosuchids to its reduction in basal epicynodonts (accompanied by a large interorbital vacuity), followed by the re-acquisition of the plesiomorphic condition and formation of a complete interorbital wall and the concomitant loss of the vacuity in derived eucynodonts, is puzzling. The orbitosphenoid is usually cartilaginous in extant sauropsids (de Beer 1937), which led Kemp (1979) to hypothesize that the acquisition of a large interorbital vacuity through the reduction of the orbitosphenoid in NM cynodonts could result from the supposed persistence of a primitive cranial kinetic mechanism. Complete ossification of the braincase, on the other hand, may have served to protect the central nervous system in eucynodonts (both among probainognathians and cynognathians), in which the brain became an increasingly important organ to support improvements in sensorial inputs and a more flexible behavior in those likely nocturnal species (Rowe et al. 2011; Benoit et al. 2016b). If true, then this is not reflected in endocranial cast size since the encephalization quotient (a measure of relative brain size) does not vary significantly among NM therapids (Jerison 1973; Rowe et al. 2011; Castanhinha et al. 2013; Laaß 2015; Rodrigues et al. 2013). The orbitosphenoid serves as a support for the olfactory bulbs which began to grow larger in the clade unifying Brasilitherium and Mammaliaformes (Rodrigues et al. 2013). However, the ancestral character state reconstruction does not reveal any particular change in the anatomy of the orbitosphenoid at this level of the cladogram (Fig. 2). In addition, the ossification of the orbitosphenoid into a solid braincase provides support for the development of the internal adductor musculature (e.g., levator pterygoidei), which might have increased the power of the bite and efficiency of mastication, particularly in herbivorous taxa such as traversodontids and tritylodontids (Kemp 2005; Jones et al. 2009). This would, at least partially, account for the convergent evolution of a longer and ventrally keeled orbitosphenoid in these two groups (Figs. 2 and 3). These preliminary hypotheses are thus not completely satisfactory but they may be relevant for some of the cases of appearance and disappearance of the orbitosphenoid in NM cynodonts. Further work is necessary to address this baffling evolutionary transition.
Conclusion
The orbitosphenoid is an important component of the braincase that supports the olfactory region of the brain. This bone is represented in all the therapsid lineages, generally as a hemi-cylindrical element. For years, it was believed that the orbitosphenoid was not ossified in basal epicynodonts (e.g., Thrinaxodon; Luo 1994), but here, for the first time, we demonstrate the presence of thin, short, and cancellous orbitosphenoids. This finding illustrates the importance of non-destructive μCT investigation prior to physical preparation and shows that innovative imaging techniques can still reveal new anatomical details, even in a well-known taxon such as Thrinaxodon.
References
Abdala F (2003) Galesaurid cynodonts from the Early Triassic of South Africa: another example of conflicting distribution of characters in non-mammalian cynodonts. S Afr J Sci 99:95–96
Abdala F, Jasinoski SC, Fernandez V (2013) Ontogeny of the Early Triassic cynodont Thrinaxodon liorhinus (Therapsida): dental morphology and replacement. J Vertebr Paleontol 33:1408–1431
Benoit J, Abdala F, Van den Brandt MJ, Manger PR, Rubidge BS (2015) Physiological implications of the abnormal absence of the parietal foramen in a late Permian cynodont (Therapsida). Sci Nat 102(11–12):69
Benoit J, Manger PR, Rubidge BS (2016a) Palaeoneurological clues to the evolution of defining mammalian soft tissue traits. Sci Rep 6:25604. doi:10.1038/srep25604
Benoit J, Manger PR, Fernandez V, Rubidge B (2016b) Cranial bosses of Choerosaurus dejageri (Therapsida, Therocephalia): earliest evidence of cranial display structures in eutheriodonts. PLoS One 11:e0161457
Bever GS, Bell CJ, Maisano JA (2005) The ossified braincase and cephalic osteoderms of Shinisaurus Crocodilurus (Squamata, Shinisauridae). Palaeontol Electron 8.1(4A):1–36
Bonaparte JF (1962) Descripción del craneo y mandíbula de Exaeretodon frenguellii, Cabrera. Pub Mus Cienc Mar del Plata 1:135–202
Bonaparte JF, Barberena MC (2001) On two advanced carnivorous cynodonts from the Late Triassic of southern Brazil. Bull Mus Comp Zool 156:59–80
Bonaparte JF, Martinelli A, Schultz CL, Rubert R (2003) The sister group of mammals: small cynodonts from the Late Triassic of southern Brazil. Rev Bras Paleontol 5:5–27
Bonaparte JF, Martinelli A, Schultz CL, Rubert R (2005) New information on Brasilodon and Brasilitherium (Cynodontia, Probainognathia) from the Late Triassic, southern Brazil. Rev Bras Paleontol 8:25–46
Boonstra LD (1968) The braincase, basicranial axis and median septum in the Dinocephalia. Ann S Afr Mus 50:195–273
Brink AS (1955) A study on the skeleton of Diademodon. Palaeontol Afr 3:3–39
Brink AS (1959) Note on a new skeleton of Thrinaxodon liorhinus. Palaeontol Afr 6:15–22
Brink AS (1961) A new type of primitive cynodont. Palaeontol Afr 7:119–154
Brink AS (1963) A new skull of the procynosuchid cynodont Leavachia duvenhagei broom. Palaeontol Afr 8:57–75
Broom R (1938) On the structure of the skull of the cynodont, Thrinaxodon liorhinus, Seeley. Ann Transv Mus 19:263–269
Castanhinha R, Araújo R, Júnior LC, Angielczyk KD, Martins GG, Martins RMS, Chaouiya C, Beckmann F, Wilde F (2013) Bringing dicynodonts back to life: paleobiology and anatomy of a new emydopoid genus from the Upper Permian of Mozambique. PLoS One 8(12):e80974
Cluver MA (1971) The cranial morphology of the dicynodont genus Lystrosaurus. Ann S Afr Mus 56:155–274
Colbert EH, Kitching JW (1977) Triassic cynodont reptiles from Antarctica. Am Mus Novit 2611:1–30
Crompton AW (1958) The cranial morphology of a new genus and species of ictidosaurian. Proc Zool Soc London 130:183–216
Crompton AW (1964) On the skull of Oligokyphus. Bull Brit Mus Nat Hist Geol 9(4):70–82
Crompton AW, Musinsky C, Owerkowicz T (2015) The evolution of the mammalian nose. In: Dial K, Shubin NH, Brainerd EL (eds) Great transformations in vertebrate evolution. University of Chicago Press, Chicago, pp 189–203
Crompton AW, Owerkowicz T, Bhullar B, Musinsky C (2017) Origin of the Lateral Wall of the Mammalian Skull: Fossils, Monotremes and Therians Revisited. J Mammal Evol. doi:10.1007/s10914-017-9388-7
Currie PJ (1997) Braincase anatomy. In: Currie PJ, Padian K (eds) Encyclopedia of dinosaurs. Academic Press, New York, pp 81–85
Day MO, Rubidge BS, Abdala F (2016) A new mid-Permian burnetiamorph specimen from the Main Karoo Basin of South Africa and a phylogenetic review of Burnetiamorpha. Acta Paleontol Pol 61:701–719
De Beer GR (1937) The development of the vertebrate skull. The Clarendon Press, Oxford
De Beer GR (1985) The development of the vertebrate Skull. The University of Chicago Press, Chicago
Estes R (1961) Cranial anatomy of the cynodont reptile Thrinaxodon liorhinus. Bull Mus Comp Zool 125:165–180
Fernandez V, Abdala F, Carlson KJ, Cook DC, Rubidge BS, Yates A, Tafforeau P (2013) Synchrotron reveals Early Triassic odd couple: injured amphibian and aestivating therapsid share burrow. PLoS One 8(6):e64978
Fourie S (1974) The cranial morphology of Thrinaxodon liorhinus Seeley. Ann S Afr Mus 65:337–400
Fourie H (1993) A detailed description of the internal structure of the skull of Emydops (Therapsida: Dicynodontia). Palaeontol Afr 30:103–111
Gow CE (1986) The side wall of the braincase in cynodont therapsids, and a note on the homology of the mammalian promontorium. S Afr J Zool 21:136–148
Haughton (1918) Some new carnivorous Therapsida, with notes upon the brain-case in certain species. Ann S Afr Mus 12:175–216
Hopson JA (1964) The braincase of the advanced mammal-like reptile Bienotherium. Postilla 87:1–30
Hopson JA (1979) Paleoneurology. In: Glans C, Northcutt RG, Ulinski P (eds) Biology of the Reptilia. Academic Press, New York, pp 39–146
Hopson JA, Kitching JW (2001) A probainognathian cynodont from South Africa and the phylogeny of non-mammalian cynodonts. Bull Mus Comp Zool 156:5–35
Ivakhnenko MF (2013) Cranial morphology of Dvinia prima Amalitzky (Cynodontia, Theromorpha). Paleontol J 47:210–222
Jasinoski SC, Abdala F, Fernandez V (2015) Ontogeny of the Early Triassic cynodont Thrinaxodon liorhinus (Therapsida): cranial morphology. Anat Rec 298:1440–1464
Jerison HJ (1973) Evolution of the brain and intelligence. Academic Press, New York
Jones MEH, Curtis N, O’Higgins P, Fagan M, Evans SE (2009) The head and neck muscles associated with feeding in Sphenodon (Reptilia: Lepidosauria: Rhynchocephalia). Palaeontol Electron 12.2(7A):1–56
Kemp TS (1969) On the functional morphology of the gorgonopsid skull. Phil Trans R Soc B Biol Sci 256:1–83
Kemp TS (1972) Whaitsiid Therocephalia and the origin of cynodonts. Phil Trans R Soc B Biol Sci 264:1–54
Kemp TS (1979) The primitive Cynodont Procynosuchus: functional anatomy of the skull and relationships. Phil Trans R Soc B Biol Sci 285:73–122
Kemp TS (2005) The origin and evolution of mammals. Oxford University Press, Oxford
Kemp TS (2009) The endocranial cavity of a nonmammalian Eucynodont, Chiniquodon theotenicus, and its implications for the origin of the mammalian brain. J Vertebr Paleontol 29:1188–1198
Kermack KA, Mussett F, Rigney HW (1981) The skull of Morganucodon. Zool J Linnean Soc 71:1–158
Kielan-Jaworowska Z, Cifelli RL, Luo ZX (2004) Mammals from the age of dinosaurs origins, evolution, and structure. Columbia University Press, New York
Kühne WG (1956) The Liassic Therapsid Oligokyphus.Trustees of the British museum (natural history). London
Laaß M (2015) Virtual reconstruction and description of the cranial Endocast of Pristerodon mackayi (Therapsida, Anomodontia). J Morphol 276:1089–1099
Liu J, Olsen P (2010) The phylogenetic relationships of Eucynodontia (Amniota: Synapsida). J Mamm Evol 17:151–176
Luo Z (1994) Sister-group relationships of mammals and transformations of diagnostic mammalian characters. In: Fraser NC, Sues HD (eds) In the shadow of the dinosaurs: early Mesozoic Tetrapods. Cambridge University Press, Cambridge, pp 98–128
Maddison WP, Maddison DR (2009) Mesquite: a modular system for evolutionary analysis Version 2.6 http://mesquiteproject.org
Martinez RN, May CL, Forster CA (1996) A new carnivorous cynodont from the Ischigualasto formation (Late Triassic, Argentina), with comments on eucynodont phylogeny. J Vertebr Paleontol 16:271–284
Olson EC (1938) The occipital, otic, basicranial and pterygoid regions of the Gorgonopsia. J Morphol 62:141–175
Rodrigues PG, Ruf I, Schultz CL (2013) Study of a digital cranial endocast of the non-mammaliaform cynodont Brasilitherium riograndensis (Later Triassic, Brazil) and its relevance to the evolution of the mammalian brain. Paläontol Z 88:329–352
Romer AS (1970) The Chañares (Argentina) Triassic reptilian fauna VI. A chiniquodontid cynodont with an incipient squamosal-dentary jaw articulation. Breviora 344:1–18
Romer AS, Price LW (1940) Review of the Pelycosauria. GSA Spec Pap 28:1–534
Rowe T (2002-2005) Digimorph, digital morphology library, University of Texas Digital Morphology. Group. http://Digimorph.Org. Accessed: 25/11/2016
Rowe T, Carlson W, Bottorff W (1995) Thrinaxodon: digital atlas of the skull, Second edn. University of Texas Press, Austin
Rowe TB, Macrini TE, Luo ZX (2011) Fossil evidence on origin of the mammalian brain. Science 332:955–957
Rubidge BS, Sidor CA (2001) Evolutionary patterns among Permo-Triassic therapsids. Annu Rev Ecol Syst 32:449–480
Ruf I, Maier W, Rodrigues PG, Schultz CL (2014) Nasal anatomy of the nonmammaliaform cynodont Brasilitherium riograndensis (Eucynodontia, Therapsida) reveals new insight into mammalian evolution. Anat Rec 297:2018–2030
Ruta M, Botha-Brink J, Mitchell SA, Benton MJ (2014) The radiation of cynodonts and the ground plan of mammalian morphological diversity. Phil Trans R Soc B Biol Sci 280:20131865. doi:10.1098/rspb.2013.1865
Sidor CA, Smith RMH (2007) A second burnetiamorph therapsid from the Permian Teekloof formation of South Africa and its associated fauna. J Vertebr Paleont 27:420–430
Sigogneau D (1970) Revision systematique des gorgonopsiens Sud-Africains. Cahiers de Paleontologie, Paris
Sigurdsen T, Huttenlocker A, Modesto SP, Rowe TB, Damiani R (2012) Reassessment of the morphology and Paleobiology of the Therocephalian Tetracynodon darti (Therapsida), and the phylogenetic relationships of Baurioidea. J Vertebr Paleontol 32:1113–1134
Soares MB, Schultz CL, Horn BLD (2011) New information on Riograndia guaibensis Bonaparte, Ferigolo & Ribeiro 2001 (Eucynodontia, Tritheledontidae) from the Late Triassic of southern Brazil: anatomical and biostratigraphic implications. An Acad Bras Cienc 83:329–354
Sues HD (1986) The skull and dentition of two tritylodontid synapsids from the Lower Jurassic of western North America. Bull Mus Comp Zool 151:217–268
Sun AL (1984) Skull morphology of the tritylodontid genus Bienotheroides of Sichuan. Sci Sin B 27:270–284
Tatarinov LP (1974) Terriodont of USSR. Trudy Paleontologicheskii Zhurnal 143:1–226
von Huene F (1936) Die fossilen Reptilien des südamerikanischen Gondwanalandes. Ergebnisse der Sauriergrabungen in Südbrasilien 1928/29. Beck'sche Verlagsbuchhandlung, Munich
Acknowledgements
We thank K. Jakata (ESI, University of the Witwatersrand, ZA), P. Keanly (X-Sight, ZA), C. Churms (DebTech, DeBeers, ZA), G. Rougier (University of Louisville), and the European Synchrotron Radiation Facility for the provision of synchrotron radiation facilities and P. Tafforeau for the assistance in using beamline ID17. We also thank ESI preparators T. Nemavhundi and C. Dube. This research was conducted with the financial support from PAST and its Scatterlings projects, the National Research Foundation of South Africa, the Claude Leon Foundation, and the DST-NRF Centre of Excellence in Palaeosciences (CoE in Palaeosciences). Opinions expressed and conclusions arrived at, are those of the authors and are not necessarily to be attributed to the CoE in Paleosciences.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by: Robert R. Reisz
Electronic supplementary material
ESM 1
(XLSX 56 kb).
Rights and permissions
About this article

Cite this article
Benoit, J., Jasinoski, S.C., Fernandez, V. et al. The mystery of a missing bone: revealing the orbitosphenoid in basal Epicynodontia (Cynodontia, Therapsida) through computed tomography. Sci Nat 104, 66 (2017). https://doi.org/10.1007/s00114-017-1487-z
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00114-017-1487-z