
Abstract
Polyploidization is pivotal in plant speciation, affecting adaptability, ecological tolerance and specific geographical distribution patterns. While cytotype diversity has been extensively studied in angiosperms and ferns, knowledge in homosporous lycophytes remains very limited. Our study addresses this gap, focusing on the homosporous lycophyte Huperzia selago in Central Europe. The genome size of 1330 individuals from 208 populations were assessed via flow cytometry, revealing five distinct cytotypes (2x, 3x, 4x, 5x, 6x). Challenging chromosome counting using gemmae roots yielded average counts of 140 (2x), 204 (3x), and 262 (4x) chromosomes. The hexaploid genome size (29 pg) approached the upper limits reported for lycophytes. Tetraploids were the most abundant (72.7%), while triploids (21.3%) were less common, while the remaining cytotypes comprised less than 5% of the dataset. Geographical separation of cytotypes was not observed. However, uncommon cytotypes were associated with the Alps and triploids also occured in the highest parts of Western Carpathians. Around 27.3% of populations exhibited multiple cytotypes. Except for atypical diploids, spore abortion differed between even (21.8%) and odd ploidies (65.2%). Microcharacter (stoma, spore) proportions did not linearly correspond with increasing ploidy levels. The high ploidy-level diversity and cytotype coexistence in Central European H. selago match the well documented patterns in ferns and angiosperms. These findings provide valuable insights into lycophyte polyploidy, underscoring the necessity for broader geographical sampling and application of molecular studies to elucidate phylogenetic relationships and taxonomic classifications within the genus Huperzia.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Polyploidization is one of the main mechanisms of the plant speciation process. This phenomenon has been implicated in 15% of flowering plants and 31% of homosporous fern speciation events (Wood et al. 2009). Polyploidy confers several advantages, including the restoration of fertility even when different parental genomes are involved, increased gene diversity linked to the attenuation recessive allele effects by dominant types, alteration in gene expression leading to novel adaptations, and facilitation of selfing and asexual reproduction (Comai 2005). Consequently, polyploids are known to exhibit greater ecological tolerance (Ramsey 2011), adapt more readily to harsh and colder environmental conditions (Kirchheimer et al. 2016), demonstrate enhanced potential for evolutionary novelty and changes in reproductive mechanisms, and display increased reproductive success under suboptimal conditions (te Beest et al. 2012). Conversely, higher ploidy levels may pose challenges such as the risk of gene expression modification or aneuploidy due to changes in cellular structure and errors during meiosis (Comai 2005), as well as "unintended" gene silencing (Mittelsten Scheid et al. 2003).
Changes in ploidy level may be influenced by temporary changes in species distribution and abundance driven by climatic changes, secondary contact of isolated lineages (Hewitt 2004) or due to cold-induced increased production of unreduced gametes (Brochmann et al. 2004). The Pleistocene’s cyclical shifts between glacial and interglacial periods, resulting in the recurrent isolation of lineages adapted to warmer climates and the expansion of species adapted to colder conditions, were pivotal in the evolutionary trajectory of numerous plant species (Hewitt 2004). As species retreated to lowland areas due to cooling, disjunctive ranges emerged, leading to allopatric speciation (Boucher et al. 2016). Additionally, polyploidization can be commonly associated with sympatric speciation, where ecological speciation and the formation of reproductive barriers take place (Ostevik et al. 2012).
However, both reproductive and geographic barriers among plants of different ploidy levels may not be absolute, with their intensity varying between species and cytotypes (Sonnleitner et al. 2013). Consequently, cytotypic variation can persist even in mixed populations without clear evolutionary significance (Hanušová et al. 2019).
While these mechanisms of speciation are well-studied in angiosperms and ferns, they remain understudied in lycophytes, the earliest diverging vascular plant group. In homosporous lycophyte it has been studied sporadically, e.g. in genus Lycopodium (Takamiya and Tanaka 1982), and Huperzia (Beitel and Wagner 1982; Takamiya and Kurita 1983). In contrast, in the heterosporous genus Isoëtes, polyploidy is a common phenomenon connected with varying degrees of fertility associated meiosis irregularities and aneuploidy across taxa (Suissa et al. 2022). Additionally, in Isoëtes, there are indications that polyploidy played a key role in speciation from isolated lineages (Liu et al. 2004). In Europe, a similar example involves the diversified heterosporous lineages of the Isoëtes lacustris complex, in which recently diversified species have been recognized based on chromosome counts and distinctive morphological characteristics (Brunton et al. 2020).
Our study deals with polyploidy in lycophytes of Central Europe, with a particular focus on the homosporous genus Huperzia. We extensively examined this phenomenon in the relatively common mountain lycophyte, H. selago (L.) Schrank and Mart., the sole species of the genus reported thus far in Central Europe. This circumboreal taxon occurs in a very wide range of habitats including mountain forest understories, subalpine and alpine meadows, moist rocks, stony screes and Pinus mugo scrubs (Kaplan et al. 2019). Its chromosome number is reported to be 2n = 264 (Manton 1950). In contrast, H. arctica (Tolm.) Sipliv is reported from Northern Europe. It is a circumpolar taxon growing in tundra and alpine meadows (Blockeel 2006) and is believed to have a chromosome number of 2n = 90 (Sorsa 1962; Valentine and Moore 1993; Blockeel 2006). Furthermore, several other taxa were recently reported from Northern Europe such as H. continentalis Testo, A. Haines & A. V. Gilman (Testo et al. 2016), H. europaea Björk, and H. acicularis Björk (Björk 2020). Their distribution, habitat requirements and ploidy level are not sufficiently known as they have not been recognized until recently. In Europe, the genus also includes the Macaronesian endemic H. suberecta (Lowe) Tardieu, reported from Madeira and the Azores (Blockeel 2006).
In the genus Huperzia, various chromosome numbers have been reported for several Asian species: 2n = 134, 136, 204, 264, 272, 276 and 528 (Ghatak 1965; Beitel and Wagner 1982; Takamiya and Kurita 1983; Takamiya 1984; Li et al. 2024), highlighting significant cytotype variability within the genus. Notably, conflicting chromosome counts have been published for H. selago: 2n = 68 (Hagerup in Hagerup and Peterson 1960; Löve and Löve 1961), 2n = 88 (Harmsen in Löve and Löve 1948), 2n = 90 (Sorsa 1962), and 2n = 264 (Manton 1950). However, some values were later deemed mistaken, corresponding to the chromosome number of Lycopodium in the same location, or were published as single numbers without proper documentation (Löve and Löve 1965). Studying this material is challenging, as indicated by the literature (Manton 1950; Wagner 1992), therefore chromosome counts from Europe are relatively scarce.
As an alternative to chromosome counting, flow cytometry is currently the most widely used method for assessing ploidy levels and genome size (e.g., Sliwinska et al. 2005). However, common practice requires fresh tissues to be used. In contrast, some studies report the successful use of dried tissues of angiosperms (Tomaszewska et al. 2021), ferns (Wolf et al. 2015), mosses (Bainard et al. 2020), and lycophytes (Little et al. 2007; Bainard et al. 2011b). However, the results obtained from dried tissues should be limited to the ploidy level estimation, interpreting fine (within-ploidy) genome size differences is not recommended at all (Sliwinska et al. 2022). Most of these successful measurements were done from silica gel-dried tissues, but air-dried samples (mosses; Bainard et al. 2020) and herbarium vouchers, including lycophytes (Selaginella; Little et al. 2007; Bainard et al. 2011b), were successfully measured as well. We adopted a comprehensive approach in our study, comparing fresh, silica-dried, and air-dried (herbarium) H. selago specimens. This allowed us to assess the applicability of DNA ploidy level estimation in study materials lacking fresh tissues, such as those from remote areas or existing herbarium specimens.
In samples for which flow cytometry is not feasible or possible (old material, laboratory equipment not available to the researcher), traditionally accepted micromorphological characters such as spore abortion, spore size and stomata size, which often correlate with genome size and ploidy, can be employed (Wagner et al. 1986; Ekrt et al. 2021). However, the validity of these characteristics should be tested via calibration.
In this study, we focused on the cytotype diversity and distribution patterns of the widespread mountain lycopod H. selago in a broader area of Central Europe. We attempt to answer the following questions: (1) What is the cytotype variation in H. selago in Central Europe? (2) Can individual cytotypes be assigned to ploidy levels corresponding with chromosome counts? (3) Are genome size measurements from dried material (silica-dried, herbarium vouchers) reliable? (4) Do genome size, stoma size, spore size and spore abortion allow the determination of ploidy level?
Materials and methods
Field sampling
Plant material was sampled during 2016–2023. The sampling of 1330 plants from 208 populations covered the diversity of all typical habitats (forests, spring areas, alpine grasslands, rocks, screes, etc.) occupied by the H. selago across Central Europe with particular emphasis on Central European mountains. Depending on the plant’s abundance at a given locality, we sampled around 10 plants per population (if possible) about 2 m apart, occasionally more to capture ploidy variability. Each site was characterized by geographical coordinates, habitat, and elevation (see Supplementary Data SI1). Whole plants or a subset of shoots per plant were collected to make herbarium vouchers and for flow cytometry (FCM). Samples for FCM were either kept fresh until the analysis or a ca 4 cm long part of one shoot was desiccated using silica gel within a day. Herbarium vouchers are deposited in CBFS. Distribution of samples and discovered ploidies and occurrence of the ploidies on elevation scale were displayed in ArcGIS Pro 3.0.3 (ESRI, Los Angeles, USA).
Flow cytometry
Flow cytometry was applied for ploidy identification and genome size estimation in H. selago. We used the two-step protocol with Otto buffers (Otto 1992) following the best practice recommendations (Sliwinska et al. 2022). Individual plants or pooled samples (usually two individuals) were chopped with the internal standard (amount of the material about 5:4 as sample:standard) in 400 μl of Otto I buffer using a sharp razor blade in a Petri dish and filtered through 42 μm nylon mesh. After 3–5 min 800 μl of Otto II buffer with 2-mercaptoethanol (2 mg/ml) and fluorochrome DAPI (4,6-diamidino-2-phenylindole, final concentration 4 μl/ml) was added into the sample. Stained samples were analysed using a CyFlowSpace instrument (Sysmex-Partec) equipped with 365 nm UV-LED as a light source. Fluorescence intensity of 3000 particles was recorded. Flow cytometry histograms were evaluated using Flowing Software 2.5.1 (P. Terho, University of Turku, freeware available at https://bioscience.fi/services/cell-imaging/flowing-software/). The relationship between sample relative fluorescence (i.e., ratio of sample/standard mean fluorescence) and ploidy level was calibrated using chromosome counting (see below).
As DAPI stain selectively binds to AT bases of DNA, genome size differences may seemingly appear between samples differing strongly in their genomic AT/GC content. Due to possible differences in AT/GC content between the sample and the standard, it is impossible to calculate sample genome size based on DAPI staining. To address this issue, genome size was determined for a selected subset of samples representing all cytotypes using propidium iodide (PI) staining, which stains all bases indiscriminately. The sample preparation was identical, only using PI at final concentration 50 μg/mL as fluorochrome instead of DAPI. As internal standards Pisum sativum ‘Ctirad’ (2C DNA = 9.09 pg; Doležel et al. 1998) was used for the PI-stained measurements, and Vicia faba ‘Inovec’ (2C DNA = 26.90 pg; Doležel et al. 1992) for the DAPI-stained ones. Measurements with other standards used because of possible overlaps were recalculated to these standards, based on calibration of all standards against each other. Fluorescence intensity of 5000 particles was analyzed using a CyFlowSpace instrument (Sysmex-Partec) equipped with a green solid-state laser (Cobolt Samba 532 nm, 100 mW). In PI analyses, samples were always processed individually and a mean value of three measurements on different days was used for genome size calculation.
Most samples were measured from fresh tissue, but silica-dried or herbarium samples were also used. To compare analysis quality using fresh, silica-dried and herbarium tissue, 51 samples of the two most common cytotypes (3 × and 4x, see below) were measured by DAPI-staining from each type of tissues and ten samples (3 × a 4x) were measured by PI-staining three times from each type of tissue. For calibration purposes, all samples measured by PI-staining were also measured by DAPI-staining. Differences in the genome size and coefficients of variation (CV) of a sample peak (as a common measure of the analysis quality) were compared between types of tissue preservation.
Statistical analyses were performed in R 4.1.2 (R Core Team 2021). A linear mixed-effect model with sample identity as a random factor was used to compare the relative genome size and sample CV between the tissue types (separately for each cytotype). Differences among groups were tested using Tukey contrasts as implemented in the function glth from the multcomp package (Hothorn 2008).
Karyology
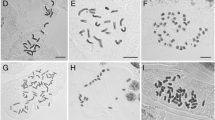
Plants of the three lowest cytotypes were used for chromosome counting: diploids (2x) from population ID 108 (Rasen-Antholz, the Alps, Italy), triploids (3x) from population ID 102 (Tatranská Lomnica, Vysoké Tatry Mts, Slovakia), tetraploids (4x) from populations ID 45 (Uricani, Retezat Mts, Romania) and 117 (Matrei in Osttirol, Hohe Tauern Mts, Austria), see Supplementary Data SI1 for the locality details. Because of very slow growth of H. selago plants, instead of roots of mature plants we used gemmae (vegetative propagules) to get fresh roots with cells undergoing mitosis. The gemmae were placed on wet filter paper in a Petri dish and grown in a cultivation box at 20 °C and 12 h of light. After several days, 2–3 mm long roots were collected and put in 1,4-dichlorbenzene and incubated in darkness at room temperature for 2–3 h. After pre-treatment, the roots were rinsed in water and placed into a fixative solution 3:1 ethanol:acetic acid and the material was stored at −20 °C. Further processing followed the modified protocol of Mandáková and Lysak (2016). The roots were washed in distilled water twice for 5 min, then in cold 1 × citrate buffer (0.1 M citric acid monohydrate + 0.1 M trisodium citrate dihydrate mixed in 2:3 ratio and 10 × diluted) twice for 5 min and then the buffer was changed for an enzymatic mix (0.3% pectolyase, 0.3% cellulase, 0.3% cytohelicase in citrate buffer). The roots were digested at 37 °C for 1–1.5 h. After digestion, the enzyme mix was changed for cold 1 × citrate buffer for at least 15 min. Then each root was placed onto slide, the meristematic tissue was separated, and the rest of material was discarded. The meristem was disintegrated with a preparation needle and 20 µl of 60% acetic acid was added. After 2 min, the slide was placed on a heating block at 50 °C and the suspension was spread with a preparation needle for 30 s. Then, 100 µl of ethanol:acetic acid solution was applied on the suspension and the liquid was strained from the slide. The dried slide was stored at 4 °C. As a staining solution, 15 µl of Vectashield with DAPI was added and the preparation was covered with cover slip and fixed with nail polish. The mitotic nuclei were observed under Nikon Eclipse E600 fluorescence microscope equipped with Nikon DS-Qi1Mc camera and the picture was taken with NIS-Elements AR software.
Spore size and abortion
Spore size was measured using ten plants per each cytotype, with the exception of rare diploids and hexaploids, for which only five and two plants were analyzed, respectively. Unripe unseparated tetrads were not counted or measured. Furthermore, the number of well-developed regular spores and putative diplospores (henceforth, we mark with this term spores which are spherical and visibly bigger than others), and the number of aborted regular spores and diplospores were evaluated in all measured samples (Supplementary data Fig. S1).
The yield of the H. selago spores from herbarium vouchers is usually relatively small, but where possible, spore length (“spore size”) of regular spores was measured in the proximal view. Considering the spore as an equilateral triangle, on the side with the trilete mark upwards (i.e. with the proximal vertex visible), spore length is defined as distance the perpendicular to the center of the any distal edge towards the opposite vertex of the imaginary "triangle" (Supplementary data Fig. S2). If the spore “triangle” was isosceles (instead of equilateral), the height perpendicular to the base (i.e. the longest of the “triangle” heights) was measured. Diplospore size was measured between the most distant points of the shape because of the oval shape of diplospores. Where possible, 20 regular spores and 10 diplospores were measured for each plant. All measurements were done at 400 × magnification. Differences in the regular spores and diplospores size between ploidies were tested using one-way ANOVA with Tukey HSD (honestly significant difference), based on mean values for each individual.
Spore abortion rates (SAI; Hornych and Ekrt 2017) were determined for all plants used for spore size measurement. In total, 500 (in case this was not possible, only 200 or 250) spores were assessed at 100 × magnification in 1% acetocarmine solution (Supplementary data Fig. S1), in which aborted spores with differently shaped exospore and without cell content have different color than ones with well-developed content. The number of regular spores and diplospores was recorded from the whole number. One-way ANOVA with Tukey HSD test was used to compare the proportion of aborted regular spores and diplospores between the cytotypes; % of the aborted spores was arcsine-transformed.
Stomata size
For this measurement, clear nail polish was first applied to the underside of microphylls, and a transparent adhesive tape was used to make imprint, which was transferred to the slide. Stomata were measured at 400 × magnification. Ten stomata per plant were measured, and from each stoma one guard cell was measured longitudinally. Ten plants of each cytotype (the same plant as for SAI and spore size measurements) were analyzed, but in the case of the rare cytotypes (diploids, hexaploids) only five and four plants were used, respectively. Length of stomata among the cytotypes was compared by one-way ANOVA with Tukey HSD test; each individual was represented by the mean value.
All spore and stomata measurements were done using Olympus CX31 microscope equipped with Lumenera Infinity 1 camera and QuickPhoto camera 3.2 software (Promicra Program 2020; www.promicra.com).
Results
Genome sizes and karyology
Flow cytometry screening of 1330 samples from 208 populations in central Europe revealed five different cytotypes (Fig. 1) with no overlapping values of the (relative) genome sizes between the cytotypes and only small variation within the cytotypes (Fig. 1, Table 1). The chromosome numbers of the lowest-ploidy cytotypes were 2n = 140 (mean of the two best chromosome figures being 137 see Fig. 1 and 143 see Supplementary data Fig. S3), ca 204 (only one figure available) and ca 262 (mean of five best figures (Fig. 1), range 253–265 see Supplementary data Fig. S4). These values approach the expected values based on the published x = 67 (Wagner 1992), 2n = 134, 201, 268 for diploids, triploids and tetraploids, respectively. The two other cytotypes were then identified as DNA-5 × and DNA-6 × based on the genome sizes. The monoploid genome size (1Cx value) was similar among all recorded ploidies, with the maximum value in diploid (5.02 pg) and minimum value in triploid (4.50 pg).

(1) Flow cytometric histogram showing a simultaneous analysis of the 5 detected ploidy levels of H. selago measured from the fresh plants coming from Central Europe. (2) Mitotic chromosome spreads of H. selago under fluorescent microscope: a diploid, 2n ⁓ 137 chromosomes, locality ID 108, plant 2 (Rasen-Antholz, the Alps, Italy); b triploid, 2n ⁓ 204 chromosomes, locality ID 102 (Tatranská Lomnica, Vysoké Tatry Mts, Slovakia); c tetraploid 2n ⁓ 262 chromosomes, locality ID 45 (Uricani, Retezat Mts, Romania); scale bar = 10 µm. For locality ID see Supplement SI1. (3) Illustrative pictures of H. selago cytotypes: a diploid, locality ID 159 (Rasen-Antholz, the Alps, Italy), b triploid, locality ID 30 (Hohentauern, Niedere Tauern Mts, Austria), c tetraploid, locality ID 160 (Anholtz-Mittertal, the Alps, Italy), d pentaploid, locality ID 117 (Matrei in Osttirol, Hohe Tauern Mts, Austria), e hexaploid, locality ID 84 (Andermatt, Glarner Alpen, Switzerland). All photos L. Ekrt. For locality ID see Supplement SI1
Cytogeography
In our Central European dataset the most common ploidy level was tetraploid (72.7%, Fig. 2a), the second most common one was triploid (21.3%). Other ploidies were very rare (2x: 0.8, 5x: 4.2 and 6x: 0.9%). Tetraploids were ubiquitous, they are distributed across the whole sampling area from lowlands to the highest mountains (195–2860 m a. s. l.; Fig. 2) and as a result are morphologically very polymorphic (Fig. S6). The other ploidies were rare and concentrated only in the Alps, except for triploids that were also found in the highest part of the Western Carpathians (Fig. 2b). Apart from the common tetraploids, all other ploidies were found only in higher elevations (1570–2860 m a. s. l.), but without any clear distribution pattern. Diploids were only found extremely rarely in one area (two near sites) of the mountain saddle Staller Sattel (2052 m) on the Austrian-Italian state border by Sankt Jakob in Defereggen, Tirol (Fig. 2c).

Cytotype proportion in the study area (a), altitude gradient (b), and spatial (c) distribution of cytotypes of H. selago in Central Europe. Mixed-ploidy populations are depicted as pie charts showing the local frequency of DNA ploidy levels (following relative genome size). Entirely tetraploid populations (green) are situated in the back of other cytotypes for improved clarity.
The sampled H. selago populations were mostly ploidy-uniform (72.7%), but approximately a quarter were mixed (27.3%; only populations with more than three samples were counted). The most common uniform populations were of tetraploids (57.6%), uniform populations of the other ploidies were less frequent (2x: 0%, 3x: 13.1%, 5x: 2.0%, 6x: 0%). Mixed populations consisted of two or three different ploidies in various combinations. Diploids were not observed in separated populations but only in mixed populations with tetraploids or with tetraploids and triploids. Similarly, hexaploids did not create separated populations, they were observed in populations together with either tetraploids, pentaploids, or both.
Most of the 208 populations were collected on silicate bedrock. Only 15 populations (7.2%) were collected on calcareous bedrock. Most of them were tetraploid, only one population from Slovakia comprised tetraploids and triploids together.
Genome size comparison of fresh and dried plants
All genome size measurements (Table 1) were obtained from living plants. However, to investigate the potential for the use of this method in further studies, we tested the fluorescence intensity and stability in comparison of fresh and dried plants. Age of most of dried tissues were similar (on the order of months), therefore this factor was not tested.
In the relative genome size measurements (DAPI stain), there was a significant effect of the material preservation method (Table 2). Especially the herbarium samples showed a slightly lower relative genome size for both ploidies tested (Table 2). However, no overlap of individual cytotypes was ever observed (Fig. 3). Both dried tissue types of triploids showed also somewhat wider range of values compared to living tissues. A different trend was observed in the absolute genome size measurements (PI stain). Results were also significantly influenced by type of material (Table 2), however, both types of dried tissues and especially in herbarium samples yielded higher values than living material (Fig. 3). The relative shift was higher in triploids than in tetraploids: + 15.2 and + 7.7%, respectively, for silica dried samples, and + 24.3 and + 15.4%, respectively, for herbarium samples.

Comparison of flow cytometry results with different type of material analyzed (fresh = fresh living tissue, silica = silica-dried tissue, herb = air dried tissue from herbarium vouchers) in triploid and tetraploid cytotype of H. selago: a relative genome size (DAPI staining), b genome size (PI staining), c sample peak coefficients of variation (CV) (DAPI staining), d sample peak CV (PI staining). Blue = results of relative genome size (RGS) measurements with DAPI staining, Red = results of genome size measurements with whole genome PI staining. Letters above boxes denote differences in Tukey contrasts at alpha = 0.05
The type of material influenced significantly CV of the sample peak, which is a measure of the analysis quality (Table 2). In both type of staining silica-gel dried tissues had the highest CV, while herbarium samples had CV similar to fresh material in DAPI staining and intermediate with PI staining (Fig. 3).
Spore size, spore abortion, and stomata size
Even though Huperzia is homosporous, we found that the size and shape of spores in sporangia were not homogeneous. Plants formed tricolpate spores (regular spores), unseparated tetrads, aborted regular spores, large spherical spores (diplospores), and aborted diplospores—see Supplementary data Fig. S1. Unseparated tetrads were not evaluated, because they were considered as a residuum of unripeness. Only the size of non-aborted spores was measured. When the length of regular spores and diplospores was compared, there was a marked difference, diplospores were 1.54 × (± 0.11) larger with no overlap in size. Additionally, there were significant differences among ploidies (one-way ANOVA; regular spores: F = 19.09, df = 4, 32, p < 0.001; diplospores: F = 21.37, df = 4, 28, p < 0.001). In both spore types, diploids had significantly smaller spores than all other ploidies. In polyploids, spore size differed significantly between triploids and pentaploids, while tetraploids and hexaploids were either intermediate or similar to triploids; diplospores in hexaploids were not evaluated due to lack of material (Table 1, Supplementary data Fig. S5).
The proportion of diplospores (Table 1) was variable (0–19.0%). The lowest proportion was observed in hexaploids (but note that only 2 plants were available) and the highest proportion was in pentaploids. The highest variability between individual plants was observed in pentaploids and diploids.
Spore abortion (SAI), counted from all spores except for unseparated tetrads (i.e., including both regular spores and diplospores), showed difference among ploidy levels (one-way ANOVA, F = 14.48, df = 4, 31, p < 0.001). Tetraploids and hexaploids had lower abortion rate than odd ploidy levels, which also differed from each other (abortion rate in pentaploids being higher than in triploids). Surprisingly, diploids had relatively high abortion rate intermediate between the two odd ploidy levels and significantly higher than both even ploidy levels (Table 1, Supplementary data Fig. S5).
As a last character, stoma size was measured (Table 1, Supplementary data Fig. S5). There were significant differences among ploidy levels (F = 14.85, df = 4, 34, p < 0.001). Diploids (42.38 ± 1.60 µm; mean and standard deviation) were different from all other ploidies. Among polyploids, triploids (51.46 ± 2.15 µm) differed from tetraploids (58.92 ± 3.04 µm), while pentaploids and hexaploids were intermediate between them (55.85 ± 6.31 µm and 55.80 ± 3.08 µm, respectively).
Discussion
Resolution of cytotypes from genome size and chromosome number
Five cytotypes (2x, 3x, 4x, 5x, 6x) were detected in Central European H. selago. Polyploidization is a common evolutionary process in many lycophytes (Bolin et al. 2018; Xia et al. 2021) with the exception of Selaginellaceae, in which polyploidy is very rare (Bainard et al. 2011a). However, ploidy series, such as the one observed in this study, are uncommon in this group of homosporous lycopods. They are usually known from the major spore-bearing land plant group, ferns (e.g., Liu et al. 2019; Hanušová et al. 2019), and in many angiosperms (e.g., Duchoslav et al. 2020) but only few examples were published in lycophytes—in Lycopodium (Takamiya and Tanaka 1982), Asian Huperzia (Beitel and Wagner 1982) and the heterosporous family Isoëtaceae (Suissa et al. 2022). The discovery of such an extensive polyploidy series in European H. selago is indeed unusual.
In this study, the genome size of Huperzia cytotypes was determined by flow cytometry. Within the genus, there are only several published values so far, all belonging to the North American-Asian diploid H. lucidula: 2C = 11.27 pg (Bainard et al. 2011a) and 11.40 pg (Wang et al. 2005). Our lowest record, also for a diploid cytotype (2C = 10.09 pg), corresponds to these values. Similar value was published recently for tetraploid H. asiatica 2C = 16.24 pg (Li et al. 2024) but this value was unfortunately based on the standard which had its genome size established molecularly, and thus this value is not comparable to the current data. The largest genome size known to date in heterosporous lycophytes 2C = 23.94 pg, was revealed in I. lacustris (Hanson and Leitch 2002). The recently published maxima of 2C values in homosporous lycophytes are 27.43 and 31.68 pg for Phlegmariurus squarrosus and P. carinatus, respectively (Wang et al. 2022). In this study the highest values of hexaploidy cytotype of H. selago, 2C = 28.39 pg is reaching the maximum lycophyte published genome size values.
Chromosome counting (especially in meiotic division) in the genus Huperzia was considered extremely difficult even by the European leading specialist I. Manton, due to high chromosome number, extreme irregularity of pairing, chromosome stickiness, long and thin chromatin, and formation of micronuclei (Manton 1950). We finally counted mitotic chromosomes in the somatic gemmae roots of three lowest cytotypes. However, since the chromosomes were not ideally condensed and because of high chromosome numbers and various lengths of chromosomes, the counts should be considered approximates.
The expected gametophytic chromosome number in diploid Huperzia, which is identical to the base chromosome number of the whole complex, is n = x = 67 or 68 (Wagner 1992). This number is very high in comparison with seed plant chromosome numbers (Rice et al. 2015). The high chromosome number of H. selago might have been caused by frequent neo- and palaeopolyploidy events, which are supported in lycophytes (e.g. Suissa et al. 2022). Similarly high chromosome counts are also common in ferns, in which these events are supported too (e.g., Schneider et al. 2017). However, genome multiplication resulting from palaeopolyploidization are not commonly accounted in ploidy multiplicity in spore-bearing plants. Thus, the lowest chromosome numbers found in Huperzia (2n = 134 or 136) are perceived as diploid, based on Wagner (1992). These numbers also correspond with other species, except of H. selago, such as H. lucidula and H. serrata (Beitel and Wagner 1982; Takamiya and Kurita 1983).
The value of the base chromosome number x = 67 results in chromosome numbers 2n = 201, 268, 335, 402 expected for triploids, tetraploids, pentaploids and hexaploids, respectively. The published counts are consistent with this series, although there are minor deviations. For example, chromosome count reported for H. selago from Europe is 2n = 264 (Manton 1950), instead of 268 (counted also in North America by Wagner 1992). Similarly, our counts for three cytotypes do not exactly match the theoretical values (being 2n ~ ca 140, ca 204, ca 262). Whether these deviations stem from dysploidy/aneuploidy or from technical issues (counting in Huperzia being extremely difficult, see above), remains an open question. However, the existence of a continuous ploidy series is indicated by the known chromosome numbers in the genus Huperzia: 2n = 134, 136, 204, 264, 272, 276, and 528 (Manton 1950; Ghatak 1965; Beitel and Wagner 1982; Takamiya and Kurita 1983; Takamiya 1984; Li et al. 2024), where 2n = 134 is stated also as 2 × value and 2n = 276 as tetraploid value. Apart from the values mentioned and discussed here, no other lower numbers were counted, and broader geographical sampling in Northern Europe and other northern regions is possibly needed. However, the reported approach of ploidy discrimination is also in line with discovered chromosome count variability in Asian species (Ghatak 1965; Takamiya and Kurita 1983; Takamiya 1984).
Ploidy distribution patterns
In our cytogeographical study focusing on broader Central Europe, surprisingly five cytotypes in H. selago were found. Each of them represents a different ploidy level, while there is virtually no genome size variation within ploidy level. The geographically most widespread cytotype (tetraploid) also occupies the widest amplitude of environmental conditions. It was almost ubiquitous across the whole sampled area reaching a wide altitudinal gradient ranging 195–2860 m a. s. l. It was dominant on silicate, but also found on calcareous bedrock. On the other hand, the other cytotypes (diploid, triploid, pentaploid and hexaploid) occurred only in the subalpine zone of the Alps and the Tatra Mts (the West Carpathians) at about 1680–2860 m a. s. l. The presence of rare ploidies in subalpine zones may be related with different aspects connected with their evolutionary history. While for diploids such habitat can be the remainder of their former distribution (Brochmann et al. 2004), so for polyploids it can be a free niche which they can colonize thanks to their higher tolerance to stress in harsher environments (Rice et al. 2019).
The distribution of rare Huperzia cytotypes can be explained from additional points of view. Firstly, they can represent relicts that survived in refugia similarly as some angiosperm plants (e.g. Schönswetter et al. 2005), that could have spread northward after deglaciation (e.g., Abbott et al. 2000). By contrast, they may have colonized the Alps from northern regions (Kruk et al. 2015). In the case of the genus Huperzia there could be a relation to the reported arctic species H. arctica. This hypothesis would be supported by the parallel distribution observed in the Arcto-alpine lycopod taxa—Lycopodium clavatum boreal type (L. c. subsp. monostachyon), which occurs also in the Alps in the alpine zone (Tribsch and Schönswetter 1999). Finally, the rare cytotypes could be formed uniquely or repeatedly in Central Europe. The hypotheses of independent origin or origins can be supported by existence of mixed populations in higher altitude and no pattern of their distribution. However, to understand properly the right scenario of their distribution and phylogeny, a molecular study is needed.
According to the resolved cytotype diversity, it is impossible to detect with certainty the phylogenetic context of the discovered cytotypes and their relationship to northern regions, where several taxa of genus Huperzia were reported (H. acicularis, H. arctica, H. continentalis, H. europaea, and H. selago). The main problem is the identification of Huperzia taxa in general. Its simple frond morphology without distinct identification characters makes it almost impossible to morphologically delimit individual taxa. The situation is probably strongly affected by very high morphological plasticity of plants growing in different environmental conditions (Löve and Löve 1965). From another point of view, even ploidy level or chromosome counts do not help to determine the taxa. They have not been clearly given for any of these taxa mentioned above, except H. selago, which was the only one to have its chromosomes validly counted, but ploidy has not been reported. In contrast to this condition, in the several European floras (Valentine and Moore 1993; Blockeel 2006) a lower chromosome count was assumed for H. arctica (2n = 90) and a higher chromosome count for H. selago (2n = 264), but it is not supported by primary data. Although the cytotype variation we found may theoretically be related to different numbers of chromosomes, sampling will need to be extended to northern regions (Vejvodová et al. in progress) to determine the actual distribution, morphology and relationships of the various types and their relationship to Huperzia taxa stated in Europe. For now, we consider the polyploid series discovered in the mountains of Central Europe under H. selago sensu lato. It can also be assumed that some revealed cytotypes may be part of cryptic taxa or complexes, either known from other regions or new ones. However, only a detailed molecular analysis of these types can answer this question (Vejvodová et al. in progress).
Ploidy estimation from of dried tissues
To further study the genus Huperzia, including obtaining samples from more distant areas, we tested the applicability of genome size estimation by flow cytometry from dried tissues. Based on our results, ploidy determination from silica-dried and herbarium samples is reliable using DAPI-stain, although lower quality of the analyses was observed, namely a reduction in signal intensity and a higher coefficient of variation. This is comparable to previous findings in flowering plants (Schönswetter et al. 2007), even in ferns (Wolf et al. 2015).
The analysis of herbarium and silica-dried tissues showed a notable increase in genome size due to higher CV, which was found to be a significant factor, particularly evident when using PI-stain. An increased CV has also been noted in another study (Tomaszewska et al. 2021) and some authors do not recommend using dried material for precise genome size measurements (Doležel and Bartoš 2005). Caution is in order here, because an increase in genome size using dried samples in our dataset was observed—similarly as in Bainard et al. (2011b). On the other hand, distinct outcomes were observed in lycophytes, in the genus Selaginella, where both herbarium and silica dried tissues aged 14.4 years were successfully used to measure the absolute genome size (Little et al. 2007). However, our data revealed a substantial disparity in the characteristics of dried materials. Herbarium specimens exhibited more pronounced shifts in genome size compared to silica-dried tissues. Rarely, an overlap of ploidies was found (in the case of the genome size of herbarium tetraploid and living triploid). The reduced efficiency in determining genome size in material from triploids may stem from the potential instability associated with the odd ploidy, likely due to limited chromosome pairing and the possibility of a hybrid origin. Therefore, genome size measurement obtained from dried Huperzia tissue should be utilized carefully, without direct comparison of fresh and dried plant values.
Developed and aborted spores in lycophytes
In ferns, well established species (usually having even ploidies) have well developed spores, while hybrids are generally almost completely aborted, which is a result of irregular meiosis after hybridization (Wagner et al. 1986). Similarly, in angiosperms the pollen (non)viability often reflects meiotic problems in hybrids or odd ploidy levels (e.g., Zhang et al. 2006). We therefore tested if spore viability correlates with ploidy level in the Huperzia cytotypes.
The assessment of the well-developed and aborted lycophyte spores is complicated by the thick spore wall (e.g., Ramos Giacosa et al. 2016). Therefore, there is almost no rule for recognizing abortive and viable spores. Examples of description of abortive spores were published only a few times for megaspores of Isoëtes (Wagner et al. 1986) and as histochemical difference of abortive and fertile spores in Lycopodium and Diphasiastrum (Alexander 1969). Abortive megaspores were detected in hybrids and odd ploidies in Isoëtes (Suissa et al. 2022), where an odd ploidies individually produced 99% of aborted megaspores (Taylor in Wagner et al. 1986), and in Huperzia (Beitel and Mickel 1992), where triploids aborted 95% of their spores (Takamiya 1984).
In general, our data indicated a trend similar to the general one, that odd ploidies have high spore abortion rates. Even ploidies (4x, 6x), except diploids aborted 31.8 and 11.8%, respectively, while odd ploidies (3x, 5x) had 55.0 and 75.4%, respectively, of aborted spores. However, spore abortion of odd ploidies was not as high as observed in Isoëtes and Huperzia. Additionally, ploidy resolution was also limited because diploids surprisingly had an extremely high spore abortion about 70.2%. This is comparable to odd ploidies. It is difficult to explain with our current knowledge and it may be probably related to undersampling of diploids (only 10 plants from one area) connected with the extremely rare occurrence and possible genetic drift within the small population of diploids (Spielman et al. 2004). The higher proportion of aborted spores could also be caused by abiotic stress (Arosa et al. 2009) or long-term isolation of a small population and possible inbreeding (Simiqueli et al. 2018). Both hypotheses are supported by the fact that diploids have only been found in Central Europe at one small site in the central Alps. We could speculate that the diploids survived in very limited population number whereas tetraploids were more successful and became widespread after last glacial maximum, as in some examples of ferns (Vogel et al. 1999).
Resolution of cytotypes via spores and stomata
In ferns, the spore and stoma size are usually correlated with ploidy level (Ekrt et al. 2021). In angiosperms a similar trend is known with pollen and stomata (e.g., McGoey et al. 2014). These correlations are also assumed in lycophytes (Bolin et al. 2018) but there is still a lack of data. Only a few studies focusing on stomata and spore size were published in genus Isoëtes. In general, ploidy did not correlate with stoma and megaspore size. On the other hand, there appeared a positive correlation of ploidy and microspore size (Troìa 2001). In contrast to these results, our data confirm significant differences in stoma size only among diploids, triploids and tetraploids and in spore size among diploids and the other ploidies. Surprisingly, samples with higher ploidy levels show no difference in these microcharacters.
Besides regular (haploid) spores, additional bigger spherical spores (in our study called “diplospores”) were also found in our study. On average, their size is about 51–60% larger than regular spore size. Such spore formations were observed in Huperzia in the past only very sporadically and, in this case, they were considered to be spore mother cells (Beitel and Mickel 1992), the diploid (2n) precursors to haploid spores. However, the bigger spherical spores were noted in lycophytes even in genera Diphasiastrum (Schnittler et al. 2019) and Lycopodium (Takamiya and Tanaka 1982), but their cytological character is still not clear. Schnittler et al. (2019) and Wagner (1986) discussed a possibility of their unreduced ploidy level and their role in life cycle. Such unreduced spores could represent spore mother cells or diplospores. In ferns, diplospory is the predominant process of spore formation in fern apomicts (Manton 1950; Grusz 2016), and, to a lesser extent, their hybrids with sexual species (apo-sex hybrids; Hornych et al. 2022). Diplospores also occur in regular sexually reproducing species (Ekrt et al. 2021), but the proportion of unreduced spores is likely very low (Nakato and Masuyama 2021). If the role of bigger spherical spores in lycophytes should be evaluated, special attention must be pai d to many aspects of lycophyte reproduction biology such as sporogenesis, ploidy level of spores, their viability and their gametophytes biology.
References
Abbott RJ, Smith LC, Milne RI et al (2000) Molecular analysis of plant migration and refugia in the Arctic. Science 289:1343–1346. https://doi.org/10.1126/science.289.5483.1343
Alexander MP (1969) Differential staining of aborted and nonaborted pollen. Stain Technol 44:117–122. https://doi.org/10.3109/10520296909063335
Arosa ML, Quintanilla LG, Ramos JA et al (2009) Spore maturation and release of two evergreen macaronesian ferns, Culcita macrocarpa and Woodwardia radicans, along an altitudinal gradient. Am Fern J 99:260–272. https://doi.org/10.1640/0002-8444-99.4.260
Bainard JD, Henry TA, Bainard LD, Newmaster SG (2011a) DNA content variation in monilophytes and lycophytes: large genomes that are not endopolyploid. Chromosome Res 19:763–775. https://doi.org/10.1007/s10577-011-9228-1
Bainard JD, Husband BC, Baldwin SJ et al (2011b) The effects of rapid desiccation on estimates of plant genome size. Chromosome Res 19:825–842. https://doi.org/10.1007/s10577-011-9232-5
Bainard JD, Newmaster SG, Budke JM (2020) Genome size and endopolyploidy evolution across the moss phylogeny. Ann Bot 125:543–555. https://doi.org/10.1093/aob/mcz194
te Beest M, Le Roux JJ, Richardson DM et al (2012) The more the better? The role of polyploidy in facilitating plant invasions. Ann Bot 109:19–45. https://doi.org/10.1093/aob/mcr277
Beitel JM, Mickel JT (1992) The Appalachian firmoss, a new species in the Huperzia selago (Lycopodiaceae) complex in Eastern North America, with a new combination for the western firmoss. Am Fern J 82:41–46. https://doi.org/10.2307/1547376
Beitel JM, Wagner FS (1982) The Chromosomes of Lycopodium lucidulum. Am Fern J 72:33–35. https://doi.org/10.2307/1547051
Björk CR (2020) Notes on the holarctic species of Huperzia (Lycopodiaceae), with emphasis on British Columbia, Canada. Ann Bot Fenn 57:255–278. https://doi.org/10.5735/085.057.0410
Blockeel TL (2006) The liverworts mosses ferns of Europe. Harley Books, Colchester
Bolin JF, Hartwig CL, Schafran P, Komarnytsky S (2018) Application of DNA flow cytometry to aid species delimitation in Isoëtes. Castanea 83:38–47. https://doi.org/10.2179/16-120
Boucher FC, Zimmermann NE, Conti E (2016) Allopatric speciation with little niche divergence is common among alpine Primulaceae. J Biogeogr 43:591–602. https://doi.org/10.1111/jbi.12652
Brochmann C, Brysting AK, Alsos IG et al (2004) Polyploidy in arctic plants. Biol J Linn Soc 82:521–536. https://doi.org/10.1111/j.1095-8312.2004.00337.x
Brunton DF, Sokoloff PC, Aymerich P (2020) The taxonomy, status and origin of Isoëtes × brochonii and I. creussensis (Isoëtaceae), two Pyrenean endemic taxa. Bot Lett 167:391–408. https://doi.org/10.1080/23818107.2020.1790034
Comai L (2005) The advantages and disadvantages of being polyploid. Nat Rev Genet 6:836–846. https://doi.org/10.1038/nrg1711
Doležel J, Bartoš J (2005) Plant DNA flow cytometry and estimation of nuclear genome size. Ann Bot 95:99–110. https://doi.org/10.1093/aob/mci005
Doležel J, Greilhuber J, Lucretti S et al (1998) Plant genome size estimation by flow cytometry: Inter-laboratory comparison. Ann Bot 82:17–26. https://doi.org/10.1093/oxfordjournals.aob.a010312
Doležel J, Sgorbati S, Lucretti S (1992) Comparison of three DNA fluorochromes for flow cytometric estimation of nuclear DNA content in plants. Physiol Plant 85:625–631. https://doi.org/10.1111/j.1399-3054.1992.tb04764.x
Duchoslav M, Jandová M, Kobrlová L et al (2020) Intricate distribution patterns of six cytotypes of Allium oleraceum at a continental scale: Niche expansion and innovation followed by niche contraction with increasing ploidy level. Front Plant Sci 11:591137. https://doi.org/10.3389/fpls.2020.591137
Ekrt L, Podroužek J, Hornych O et al (2021) Cytotypes of bracken (Pteridium aquilinum) in Europe: widespread diploids and scattered triploids of likely multiple origin. Flora 274:151725. https://doi.org/10.1016/j.flora.2020.151725
Ghatak J (1965) Some evidences of cytological evolution in Lycopodium L. s. l. The Nucleus 8:45–58
Grusz AL (2016) A current perspective on apomixis in ferns. J Syst Evol 54:656–665. https://doi.org/10.1111/jse.12228
Hanson L, Leitch IJ (2002) DNA amounts for five pteridophyte species fill phylogenetic gaps in C-value data. Bot J Linn Soc 140:169–173. https://doi.org/10.1046/j.1095-8339.2002.00083.x
Hanušová K, Čertner M, Urfus T et al (2019) Widespread co-occurrence of multiple ploidy levels in fragile ferns (Cystopteris fragilis complex; Cystopteridaceae) probably stems from similar ecology of cytotypes, their efficient dispersal and inter-ploidy hybridization. Ann Bot 123:845–855. https://doi.org/10.1093/aob/mcy219
Hagerup O, Petersson V (1960) A botanical atlas: mosses, ferns, conifers, horsetails, lycopods, phylogeny, 2. Ejnar Munksgaard, Kobenhavn
Hewitt GM (2004) Genetic consequence of climatic oscillations in the quaternary. Philos Trans R Soc B 359:183–195. https://doi.org/10.1098/rstb.2003.1388
Hornych O, Ekrt L (2017) Spore abortion index (SAI) as a promising tool of evaluation of spore fitness in ferns: an insight into sexual and apomictic species. Plant Syst Evol 303:497–507. https://doi.org/10.1007/s00606-016-1386-3
Hornych O, Férová A, Hori K et al (2022) Apomictic fern fathers: an experimental approach to the reproductive characteristics of sexual, apomict, and hybrid fern gametophytes. Am J Bot 109:628–644. https://doi.org/10.1002/ajb2.1817
Hothorn T, Bretz F, Westfall P (2008) Simultaneous inference in general parametric models. Biom J 50:346–363. https://doi.org/10.1002/bimj.200810425
Kaplan Z, Danihelka J, Chrtek J et al (2019) Distributions of vascular plants in the Czech Republic. Part 8. Preslia 91:257–368. https://doi.org/10.23855/preslia.2019.257
Kirchheimer B, Schinkel CCF, Dellinger AS et al (2016) A matter of scale: apparent niche differentiation of diploid and tetraploid plants may depend on extent and grain of analysis. J Biogeogr 43:716–726. https://doi.org/10.1111/jbi.12663
Kruk J, Sliwinska E, Grabowska-Joachimiak A et al (2015) Woodsia pulchella in the Western Carpathians: a relict species at the northern limit of its distribution. Ann Bot Fenn 52:193–201. https://doi.org/10.5735/085.052.0310
Li C, Wickell D, Kuo LY et al (2024) Extraordinary preservation of gene collinearity over three hundred million years revealed in homosporous lycophytes. Proc Natl Acad Sci USA 121:e2312607121. https://doi.org/10.1073/pnas.2312607121
Little DP, Moran RC, Brenner ED, Wm Stevenson D (2007) Nuclear genome size in Selaginella. Genome 50:351–356. https://doi.org/10.1139/G06-138
Liu H, Ekrt L, Koutecký P et al (2019) Polyploidy does not control all: Lineage-specific average chromosome length constrains genome size evolution in ferns. J Syst Evol 57:418–430. https://doi.org/10.1111/jse.12525
Liu X, Gituru WR, Wang Q-F (2004) Distribution of basic diploid and polyploid species of Isoëtes in East Asia. J Biogeogr 31:1239–1250. https://doi.org/10.1111/j.1365-2699.2004.01115.x
Löve A, Löve D (1948) Chromosome numbers of northern plant species, 3. Ingólfsprent, Reykjavík
Löve A, Löve D (1961) Chromosome numbers of central and northwest European plant species. Opera Bot 5:1–24
Löve Á, Löve D (1965) Taxonomic remarks on some American alpine plants. University of Colorado, Boulder
Mandáková T, Lysak MA (2016) Chromosome preparation for cytogenetic analyses in Arabidopsis. Curr Protoc Plant Biol 1:43–51. https://doi.org/10.1002/cppb.20009
Manton I (1950) Problems of cytology and evolution in the Pteridophyta. Cambridge University Press, London
McGoey BV, Chau K, Dickinson TA (2014) Stomata size in relation to ploidy level in North American hawthorns (Crataegus, Rosaceae). Madroño 61:177–193. https://doi.org/10.3120/0024-9637-61.2.177
Mittelsten Scheid O, Afsar K, Paszkowski J (2003) Formation of stable epialleles and their paramutation-like interaction in tetraploid Arabidopsis thaliana. Nat Genet 34:450–454. https://doi.org/10.1038/ng1210
Nakato N, Ootsuki R, Murakami N, Masuyama S (2012) Two types of partial fertility in a diploid population of the fern Thelypteris decursive-pinnata (Thelypteridaceae). J Plant Res 125:465–474. https://doi.org/10.1007/s10265-011-0461-7
Ostevik KL, Moyers BT, Owens GL, Rieseberg LH (2012) Parallel ecological speciation in plants? Int J Ecol 2012:939862. https://doi.org/10.1155/2012/939862
Otto F (1992) Preparation and stain of cells for high-resolution DNA analysis. In: Radbruch A (ed) Flow cytometry and cell sorting. Springer-Verlag, Berlin, pp 101–104
R Core Team (2021) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Ramos Giacosa JP, Morbelli MA, Giudice GE, Gorrer DA (2016) Spore morphology and wall ultrastructure of Lycopodiaceae from northwest Argentina. Rev Palaeobot Palynol 225:84–94. https://doi.org/10.1016/j.revpalbo.2015.11.009
Ramsey J (2011) Polyploidy and ecological adaptation in wild yarrow. Proc Natl Acad Sci USA 108:7096–7101. https://doi.org/10.1073/pnas.1016631108
Rice A, Glick L, Abadi S et al (2015) The chromosome counts database (ccdb) – a community resource of plant chromosome numbers. New Phytol 206:19–26. https://doi.org/10.1111/nph.13191
Rice A, Šmarda P, Novosolov M et al (2019) The global biogeography of polyploid plants. Nat Ecol Evol 3:265–273. https://doi.org/10.1038/s41559-018-0787-9
Schneider H, Liu H-M, Chang Y-F et al (2017) Neo- and paleopolyploidy contribute to the species diversity of Asplenium – the most species-rich genus of ferns. J Syst Evol 55:353–364. https://doi.org/10.1111/jse.12271
Schnittler M, Horn K, Kaufmann R et al (2019) Genetic diversity and hybrid formation in Central European club-mosses (Diphasiastrum, Lycopodiaceae) – new insights from cp microsatellites, two nuclear markers and AFLP. Mol Phylogenet Evol 131:181–192. https://doi.org/10.1016/j.ympev.2018.11.001
Schönswetter P, Stehlik I, Holderegger R, Tribsch A (2005) Molecular evidence for glacial refugia of mountain plants in the European Alps. Mol Ecol 14:3547–3555. https://doi.org/10.1111/j.1365-294X.2005.02683.x
Simiqueli GF, de Resende MDV, Motoike SY, Henriques E (2018) Inbreeding depression as a cause of fruit abortion in structured populations of macaw palm (Acrocomia aculeata): Implications for breeding programs. Ind Crops Prod 112:652–659. https://doi.org/10.1016/j.indcrop.2017.12.068
Sliwinska E, Loureiro J, Leitch IJ et al (2022) Application-based guidelines for best practices in plant flow cytometry. Cytom Part A 101:749–781. https://doi.org/10.1002/cyto.a.24499
Sliwinska E, Zielinska E, Jedrzejczyk I (2005) Are seeds suitable for flow cytometric estimation of plant genome size? Cytom Part A 64A:72–79. https://doi.org/10.1002/cyto.a.20122
Sonnleitner M, Weis B, Flatscher R et al (2013) Parental ploidy strongly affects offspring fitness in heteroploid crosses among three cytotypes of autopolyploid Jacobaea carniolica (Asteraceae). PLoS ONE 8:e78959. https://doi.org/10.1371/journal.pone.0078959
Sorsa V (1962) Chromosomenzahlen finnischer Kormophyten. I. Ann Acad Sci Fenn A IV Bilol 58:1–14
Spielman D, Brook BW, Frankham R (2004) Most species are not driven to extinction before genetic factors impact them. Proc Natl Acad Sci USA 101:15261–15264. https://doi.org/10.1073/pnas.0403809101
Suissa JS, Kinosian SP, Schafran PW et al (2022) Homoploid hybrids, allopolyploids, and high ploidy levels characterize the evolutionary history of a western North American quillwort (Isoëtes) complex. Mol Phylogenet Evol 166:107332. https://doi.org/10.1016/j.ympev.2021.107332
Takamiya M (1984) A triploid cytotype of Lycopodium serratum, pteridophyte. Chromosome Information Service 37:25–26
Takamiya M, Kurita S (1983) Cytotaxonomic studies on Japanese species of the genus Lycopodium sensu lato. Acta Phytotax Geobot 34:66–79
Takamiya M, Tanaka R (1982) Polyploid cytotypes and their habitat preferences in Lycopodium clavatum. Bot Mag Tokyo 95:419–434
Testo W, Haines A, Gilman AV (2016) Huperzia continentalis (Lycopodiaceae), a new species of gemmiferous firmoss separated from Huperzia haleakalae. Syst Bot 41:894–901. https://doi.org/10.1600/036364416X693982
Tomaszewska P, Pellny TK, Hernández LM et al (2021) Flow cytometry-based determination of ploidy from dried leaf specimens in genomically complex collections of the tropical forage grass Urochloa s.1. Genes 12:957. https://doi.org/10.3390/genes12070957
Tribsch A, Schönswetter P (1999) Lycopodium clavatum ssp. monostachyon (L. lagopus) in den Ostalpen. Verh Zool-Bot Ges Wien 136:235–248
Troìa A (2001) The genus Isoëtes L. (Lycophyta, Isoëtaceae): synthesis of karyological data. Webbia 56:201–218. https://doi.org/10.1080/00837792.2001.10670712
Valentine DH, Moore DM (1993) Lycopsida. In: Tutin TG, Burges NA, Chater AO et al (eds) Flora Europea, Psilotaceae to Platanaceae, 2nd edn. Cambridge University Press, Cambridge, UK
VogelRumsey JCFJ, Schneller JJ et al (1999) Where are the glacial refugia in Europe? Evidence from pteridophytes. Biol J Linn Soc 66:23–37
Wagner FS (1992) Cytological problems in Lycopodium sens. lat. Ann Missouri Bot Gard 79:718–729
Wagner WH, Wagner FS, Taylor WC (1986) Detecting abortive spores in herbarium specimens of sterile hybrids. Am Fern J 76:129–140. https://doi.org/10.2307/1547721
Wang F-G, Wang A-H, Bai C-K et al (2022) Genome size evolution of the extant lycophytes and ferns. Plant Divers 44:141–152. https://doi.org/10.1016/j.pld.2021.11.007
Wang W, Tanurdzic M, Luo M et al (2005) Construction of a bacterial artificial chromosome library from the spikemoss Selaginella moellendorffii: a new resource for plant comparative genomics. BMC Plant Biol 5:8. https://doi.org/10.1186/1471-2229-5-10
Wolf PG, Rowe CA, Der JP et al (2015) Origins and diversity of a cosmopolitan fern genus on an island archipelago. AoB Plants 7:plv118
Wood TE, Takebayashi N, Barker MS et al (2009) The frequency of polyploid speciation in vascular plants. Proc Natl Acad Sci USA 106:13875–13879. https://doi.org/10.1073/pnas.0811575106
Xia Z-Q, Wei Z-Y, Shen H et al (2021) Lycophyte transcriptomes reveal two whole-genome duplications in Lycopodiaceae: insights into the polyploidization of Phlegmariurus. Plant Divers 44:262–270. https://doi.org/10.1016/j.pld.2021.08.004
Zhang ZS, Lu YG, Liu XD et al (2006) Cytological mechanism of pollen abortion resulting from allelic interaction of F1 pollen sterility locus in rice (Oryza sativa L.). Genetica 127:295–302. https://doi.org/10.1007/s10709-005-4848-z
Acknowledgements
We are also grateful to numerous colleagues who helped with field material collection: A. Čejková, K. Hanušová, J. Harčarik, J. Prančl, A. Jelínek, S. Jessen, A. Jirsa, M. Kessler, F. Kolář, M. Kolář, M. Konečná, M. Štech, M. Krištof, J. Kučera, L. Lehmann, J. Lepš, D. Půbal, F. Riedel and L. Šternerová.
Funding
Open access publishing supported by the National Technical Library in Prague. Partial financial support was received from University of South Bohemia, Faculty of Science, Department of Botany.
Author information
Authors and Affiliations
Contributions
Conceptualization: LE, KV; methodology: LE, KV, ML; field- and lab-work: LE, KV, JK, PK, ML; formal analysis and investigation: LE, KV, PK; writing original draft preparation: LE, KV, PK, ML, OH; supervision: LE.
Corresponding author
Ethics declarations
Declarations
The authors have no relevant financial or non-financial interests to disclose. The authors have no competing interests to declare that are relevant to the content of this article. All authors certify that they have no affiliations with or involvement in any organization or entity with any financial interest or non-financial interest in the subject matter or materials discussed in this manuscript. The authors have no financial or proprietary interests in any material discussed in this article.
Competing interests
None.
Availability of data and materials
Yes, on request.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Vejvodová, K., Krejčí, J., Koutecký, P. et al. High mountains of central Europe as a refuge of surprising cytotype diversity of Huperzia selago (Lycopodiaceae). Alp Botany 134, 87–100 (2024). https://doi.org/10.1007/s00035-024-00312-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00035-024-00312-0