
Abstract
The morphogen Sonic Hedgehog (SHH) plays an important role in coordinating embryonic development. Short- and long-range SHH signalling occurs through a variety of membrane-associated and membrane-free forms. However, the molecular mechanisms that govern the early events of the trafficking of neosynthesised SHH in mammalian cells are still poorly understood. Here, we employed the retention using selective hooks (RUSH) system to show that newly-synthesised SHH is trafficked through the classical biosynthetic secretory pathway, using TMED10 as an endoplasmic reticulum (ER) cargo receptor for efficient ER-to-Golgi transport and Rab6 vesicles for Golgi-to-cell surface trafficking. TMED10 and SHH colocalized at ER exit sites (ERES), and TMED10 depletion significantly delays SHH loading onto ERES and subsequent exit leading to significant SHH release defects. Finally, we utilised the Drosophila wing imaginal disc model to demonstrate that the homologue of TMED10, Baiser (Bai), participates in Hedgehog (Hh) secretion and signalling in vivo. In conclusion, our work highlights the role of TMED10 in cargo-specific egress from the ER and sheds light on novel important partners of neosynthesised SHH secretion with potential impact on embryonic development.





Similar content being viewed by others

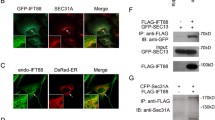

Data and materials availability
All data are available in the main text, the supplementary materials or will be made available on reasonable request.
References
Wolpert L (2016). Positional information and pattern formation. In: Current topics in developmental biology. Vol. 117. Elsevier Inc. https://doi.org/10.1016/bs.ctdb.2015.11.008.
Álvarez-Buylla A, Ihrie RA (2014) Sonic hedgehog signaling in the postnatal brain. Semin Cell Dev Biol 33:105–111. https://doi.org/10.1016/j.semcdb.2014.05.008
Briscoe J, Thérond PP (2013) The mechanisms of hedgehog signalling and its roles in development and disease. Nat Rev Mol Cell Biol 14(7):418–431. https://doi.org/10.1038/nrm3598
Dahmane N, Ruiz I Altaba A (1999) Sonic hedgehog regulates the growth and patterning of the cerebellum. Development (Cambridge, England) 126(14):3089–3100
Groves I, Placzek M, Fletcher AG (2020) Of mitogens and morphogens: modelling Sonic Hedgehog mechanisms in vertebrate development: modelling Shh mechanisms in development. Philos Trans R Soc B Biol Sci. https://doi.org/10.1098/rstb.2019.0660
Palma V, Lim DA, Dahmane N, Sánchez P, Brionne TC, Herzberg CD, Gitton Y, Carleton A, Álvarez-Buylla A, Ruiz I Altaba A (2005) Sonic hedgehog controls stem cell behavior in the postnatal and adult brain. Development 132(2):335–344. https://doi.org/10.1242/dev.01567
Straface G, Aprahamian T, Flex A, Gaetani E, Biscetti F, Smith RC, Pecorini G et al (2009) Sonic hedgehog regulates angiogenesis and myogenesis during post-natal skeletal muscle regeneration. J Cell Mol Med 13(8b):2424–2435. https://doi.org/10.1111/j.1582-4934.2008.00440.x
Corrales JMD, Blaess S, Mahoney EM, Joyner AL (2006) The Level of sonic hedgehog signaling regulates the complexity of cerebellar foliation. Development 133(9):1811–1821. https://doi.org/10.1242/dev.02351
Melamed JR, Morgan JT, Ioele SA, Gleghorn JP, Sims-Mourtada J, Day ES (2018) Investigating the role of hedgehog/GLI1 signaling in glioblastoma cell response to temozolomide. Oncotarget 9(43):27000–27015. https://doi.org/10.18632/oncotarget.25467
Rubin LL, de Sauvage FJ (2006) Targeting the hedgehog pathway in cancer. Nat Rev Drug Discov 5(12):1026–1033. https://doi.org/10.1038/nrd2086
Brady MV, Vaccarino FM (2021) Role of SHH in patterning human pluripotent cells towards ventral forebrain fates. Cells. https://doi.org/10.3390/cells10040914
Hill SA, Blaeser AS, Coley AA, Xie Y, Shepard KA, Harwell CC, Gao W-J, Denise A, Garcia R (2019) Sonic hedgehog signaling in astrocytes mediates cell type-specific synaptic organization. Elife 8:1–23. https://doi.org/10.7554/elife.45545
Yam PT, Charron F (2013) Signaling mechanisms of non-conventional axon guidance cues: the Shh, BMP and Wnt morphogens. Curr Opinion Neurobiol Dev Neurons Glia 23(6):965–973. https://doi.org/10.1016/j.conb.2013.09.002
Maity T, Fuse N, Beachy PA (2005) Molecular Mechanisms of sonic hedgehog mutant effects in holoprosencephaly. Proc Natl Acad Sci 102(47):17026–17031. https://doi.org/10.1073/pnas.0507848102
McClelland K, Li W, Rosenblum ND (2022) Pallister-Hall syndrome, GLI3, and kidney malformation. Am J Med Genet C Semin Med Genet 190(3):264–278. https://doi.org/10.1002/ajmg.c.31999
Hutchings J, Zanetti G (2019) Coat flexibility in the secretory pathway: a role in transport of bulky cargoes. Curr Opin Cell Biol 59(August):104–111. https://doi.org/10.1016/j.ceb.2019.04.002
Zanetti G, Pahuja KB, Studer S, Shim S, Schekman R (2012) COPII and the regulation of protein sorting in mammals. Nat Cell Biol 14(1):20–28. https://doi.org/10.1038/ncb2390
Peotter J, Kasberg W, Pustova I, Audhya A (2019) COPII-mediated trafficking at the ER/ERGIC interface. Traffic 20(7):491–503. https://doi.org/10.1111/tra.12654
Saraste J, Marie M (2018) Intermediate compartment (IC): from Pre-Golgi vacuoles to a semi-autonomous membrane system. Histochem Cell Biol 150(5):407–430. https://doi.org/10.1007/s00418-018-1717-2
Borchers A-C, Langemeyer L, Ungermann C (2021) Who’s in control? Principles of Rab GTPase activation in endolysosomal membrane trafficking and beyond. J Cell Biol 220(9):e202105120. https://doi.org/10.1083/jcb.202105120
Deffieu MS, Cesonyte I, Delalande F, Boncompain G, Dorobantu C, Song E, Lucansky V et al (2021) Rab7-harboring vesicles are carriers of the transferrin receptor through the biosynthetic secretory pathway. Sci Adv 7(2):eaba7803. https://doi.org/10.1126/sciadv.aba7803
Goud B, Liu S, Storrie B (2018) Rab proteins as major determinants of the Golgi complex structure. Small GTPases 9(1–2):66–75. https://doi.org/10.1080/21541248.2017.1384087
Buglino JA, Resh MD (2008) Hhat is a palmitoylacyltransferase with specificity for N-Palmitoylation of Sonic Hedgehog. J Biol Chem 283(32):22076–22088. https://doi.org/10.1074/jbc.M803901200
Chamoun Z, Mann RK, Nellen D, von Kessler DP, Bellotto M, Beachy PA, Basler K (2001) Skinny hedgehog, an acyltransferase required for palmitoylation and activity of the hedgehog signal. Science 293(5537):2080–2084. https://doi.org/10.1126/science.1064437
Porter JA, Young KE, Beachy PA (1996) Cholesterol modification of hedgehog signaling proteins in animal development. Science 274(5285):255–259. https://doi.org/10.1126/science.274.5285.255
Resh MD (2021) Palmitoylation of hedgehog proteins by hedgehog acyltransferase: roles in signalling and disease. Open Biol 11(3):rsob.200414. https://doi.org/10.1098/rsob.200414
D’Angelo G, Matusek T, Pizette S, Thérond PP, D’Angelo G, Matusek T, Pizette S et al (2015) Endocytosis of hedgehog through dispatched regulates long-range signaling. Dev Cell 32(3):290–303. https://doi.org/10.1016/j.devcel.2014.12.004
Gore T, Matusek T, D’Angelo G, Giordano C, Tognacci T, Lavenant-Staccini L, Rabouille C, Thérond PP (2021) The GTPase Rab8 differentially controls the long- and short-range activity of the hedgehog morphogen gradient by regulating hedgehog apico-basal distribution. Development 148(5):dev191791. https://doi.org/10.1242/dev.191791
Pizette S, Matusek T, Herpers B, Thérond PP, Rabouille C (2021) Hherisomes, hedgehog specialized recycling endosomes, are required for high level hedgehog signaling and tissue growth. J Cell Sci. https://doi.org/10.1242/jcs.258603
Sandoval Li, Labarca M, Retamal C, Sánchez P, Larraín J, González A (2022) Sonic hedgehog is basolaterally sorted from the TGN and transcytosed to the apical domain involving dispatched-1 at Rab11-ARE. Fronti Cell Dev Biol. https://doi.org/10.3389/fcell.2022.833175
Boncompain G, Divoux S, Gareil N, de Forges H, Lescure A, Latreche L, Mercanti V, Jollivet F, Raposo G, Perez F (2012) Synchronization of secretory protein traffic in populations of cells. Nat Methods 9(5):493–498. https://doi.org/10.1038/nmeth.1928
Thauvin M, Amblard I, Rampon C, Mourton A, Queguiner I, Li C, Gautier A, Joliot A, Volovitch M, Vriz S (2022) Reciprocal regulation of Shh trafficking and H2O2 levels via a noncanonical BOC-Rac1 pathway. Antioxidants (Basel, Switzerland) 11(4):841–856. https://doi.org/10.3390/antiox11040718
Chamberlain CE, Jeong J, Guo C, Allen BL, McMahon AP (2008) Notochord-derived Shh concentrates in close association with the apically positioned basal body in neural target cells and forms a dynamic gradient during neural patterning. Development 135(6):1097–1106. https://doi.org/10.1242/dev.013086
Aber R, Chan W, Mugisha S, Jerome-Majewska LA (2019) Transmembrane Emp24 domain proteins in development and disease. Genet Res 101:e14. https://doi.org/10.1017/S0016672319000090
Strating JRPM, Martens GJM (2009) The P24 family and selective transport processes at the ER-Golgi interface. Biol Cell 101(9):495–509. https://doi.org/10.1042/bc20080233
D’Arcangelo JG, Crissman J, Pagant S, Čopič A, Latham CF, Snapp EL, Miller EA (2015) Traffic of P24 proteins and COPII coat composition mutually influence membrane scaffolding. Curr Biol 25(10):1296–1305. https://doi.org/10.1016/j.cub.2015.03.029
Schuiki I, Volchuk A (2012) Diverse Roles for the P24 family of proteins in eukaryotic cells. Biomol Concepts 3(6):561–570. https://doi.org/10.1515/bmc-2012-0028
Pastor-Cantizano N, Montesinos JC, Bernat-Silvestre C, Marcote MJ, Aniento F (2016) P24 family proteins: key players in the regulation of trafficking along the secretory pathway. Protoplasma 253(4):967–985. https://doi.org/10.1007/s00709-015-0858-6
Petrov K, Wierbowski BM, Liu J, Salic A (2020) Distinct cation gradients power cholesterol transport at different key points in the hedgehog signaling pathway. Dev Cell 55(3):314-327.e7. https://doi.org/10.1016/j.devcel.2020.08.002
Tashima Y, Hirata T, Maeda Y, Murakami Y, Kinoshita T (2021) Differential Use of P24 family members as cargo receptors for the transport of glycosylphosphatidylinositol-anchored proteins and Wnt1. J Biochem 171(1):75–83. https://doi.org/10.1093/jb/mvab108
Blum R, Lepier A (2008) The luminal domain of P23 (Tmp21) plays a critical role in P23 cell surface trafficking. Traffic 9(9):1530–1550. https://doi.org/10.1111/j.1600-0854.2008.00784.x
Lopez S, Perez-Linero AM, Manzano-Lopez J, Sabido-Bozo S, Cortes-Gomez A, Rodriguez-Gallardo S, Aguilera-Romero A, Goder V, Muñiz M (2020) Dual independent roles of the P24 complex in selectivity of secretory cargo export from the endoplasmic reticulum. Cells 9(5):1295. https://doi.org/10.3390/cells9051295
Weigel AV, Chang C-L, Gleb Shtengel C, Shan Xu, Hoffman DP, Freeman M, Iyer N et al (2021) ER-to-golgi protein delivery through an interwoven, tubular network extending from ER. Cell 184(9):2412-2429.e16. https://doi.org/10.1016/j.cell.2021.03.035
Hatori R, Kornberg TB (2020) Hedgehog produced by the Drosophila wing imaginal disc induces distinct responses in three target tissues. Development (Cambridge, England) 147(22):dev195974. https://doi.org/10.1242/dev.195974
Tabata T, Kornberg TB (1994) Hedgehog is a signaling protein with a key role in patterning Drosophila imaginal discs. Cell 76(1):89–102. https://doi.org/10.1016/0092-8674(94)90175-9
Tanimoto H, Itoh S, ten Dijke P, Tabata T (2000) Hedgehog creates a gradient of DPP activity in Drosophila wing imaginal discs. Mol Cell 5(1):59–71. https://doi.org/10.1016/s1097-2765(00)80403-7
Torroja C, Gorfinkiel N, Guerrero I (2005) Mechanisms of Hedgehog Gradient Formation and Interpretation. J Neurobiol 64(4):334–356. https://doi.org/10.1002/neu.20168
Matusek T, Wendler F, Polès S, Pizette S, D’Angelo G, Fürthauer M, Therond PP (2014) The ESCRT machinery regulates the secretion and long-range activity of hedgehog. Nature 516(729):99–103. https://doi.org/10.1038/nature13847
Burke R, Nellen D, Bellotto M, Hafen E, Senti K-A, Dickson BJ, Basler K (1999) Dispatched, a novel sterol-sensing domain protein dedicated to the release of cholesterol-modified hedgehog from signaling cells. Cell 99(7):803–815. https://doi.org/10.1016/S0092-8674(00)81677-3
Balch WE, Michael McCaffery J, Plutner H, Farquhar MG (1994) Vesicular stomatitis virus glycoprotein is sorted and concentrated during export from the endoplasmic reticulum. Cell 76(5):841–852. https://doi.org/10.1016/0092-8674(94)90359-X
Barlowe C, Helenius A (2016) Cargo capture and bulk flow in the early secretory pathway. Annu Rev Cell Dev Biol 32:197–222. https://doi.org/10.1146/annurev-cellbio-111315-125016
Fourriere L, Kasri A, Gareil N, Bardin S, Bousquet H, Pereira D, Perez F, Goud B, Boncompain G, Miserey-Lenkei S (2019) RAB6 and microtubules restrict protein secretion to focal adhesions. J Cell Biol 218(7):2215–2231. https://doi.org/10.1083/jcb.201805002
Dickson LJ, Liu S, Storrie B (2020) Rab6 Is required for rapid, cisternal-specific, intra-Golgi cargo transport. Sci Rep 10(1):16604. https://doi.org/10.1038/s41598-020-73276-w
Huber ME, Kurapova R, Heisler CM, Karamooz E, Tafesse FG, Harriff MJ (2020) Rab6 regulates recycling and retrograde trafficking of MR1 molecules. Sci Rep 10(1):20778. https://doi.org/10.1038/s41598-020-77563-4
Patwardhan A, Bardin S, Miserey-Lenkei S, Larue L, Goud B, Raposo G, Delevoye C (2017) Routing of the RAB6 secretory pathway towards the lysosome related organelle of melanocytes. Nat Commun. https://doi.org/10.1038/ncomms15835
Coulter ME, Dorobantu CM, Lodewijk GA, Delalande F, Cianferani S, Ganesh VS, Smith RS et al (2018) The ESCRT-III protein CHMP1A mediates secretion of sonic hedgehog on a distinctive subtype of extracellular vesicles. Cell Rep 24(4):973-986.e8. https://doi.org/10.1016/j.celrep.2018.06.100
Bernat-Silvestre C, De Sousa V, Vieira J-S, Pastor-Cantizano N, Hawes C, Marcote MJ, Aniento F (2020) P24 family proteins are involved in transport to the plasma membrane of GPI-anchored proteins in plants. Plant Physiol 184(3):1333–1347. https://doi.org/10.1104/pp.20.00880
Mañuel M, Howard R (2016) Trafficking of glycosylphosphatidylinositol anchored proteins from the endoplasmic reticulum to the cell surface. J Lipid Res 57(3):352–360. https://doi.org/10.1194/jlr.R062760
Tang X, Chen R, St Dollente Mesias V, Wang T, Wang Y, Poljak K, Fan X et al (2022) A SURF4-to-proteoglycan relay mechanism that mediates the sorting and secretion of a tagged variant of Sonic Hedgehog. Proc Natl Acad Sci USA 119(11):e2113991119. https://doi.org/10.1073/pnas.2113991119
Mitrovic S, Ben-Tekaya H, Koegler E, Gruenberg J, Hauri H-P (2008) ‘The Cargo Receptors Surf4, Endoplasmic Reticulum-Golgi Intermediate Compartment (ERGIC)-53, and P25 Are Required to Maintain the Architecture of ERGIC and Golgi. Edited by Benjamin Glick. Mol Biol Cell 19(5):1976–1990. https://doi.org/10.1091/mbc.e07-10-0989
Di Minin G, Holzner M, Grison A, Dumeau CE, Chan W, Monfort A, Jerome-Majewska LA, Roelink H, Wutz A (2022) TMED2 binding restricts SMO to the ER and Golgi compartments. PLoS Biol. https://doi.org/10.1371/journal.pbio.3001596
Kim P, Scott MR, Meador-Woodruff JH (2019) Abnormal ER quality control of neural GPI-anchored proteins via dysfunction in ER export processing in the frontal cortex of elderly subjects with schizophrenia. Transl Psychiatry 9(1):1–9. https://doi.org/10.1038/s41398-018-0359-4
Qiu K, Zhang X, Wang S, Li C, Wang X, Li X, Wu Y (2019) TMP21 in Alzheimer’s disease: molecular mechanisms and a potential target. Front Cell Neurosci. https://doi.org/10.3389/fncel.2019.00328
Shin JH, Park SJ, Jo DS, Park NY, Kim JB, Bae JE, Jo YK et al (2019) Down-regulated TMED10 in Alzheimer disease induces autophagy via ATG4B activation. Autophagy 15(9):1495–1505. https://doi.org/10.1080/15548627.2019.1586249
Vetrivel KS, Kodam A, Gong P, Chen Y, Parent AT, Kar S, Thinakaran G (2008) Localization and regional distribution of P23/TMP21 in the brain. Neurobiol Dis 32(1):37–49. https://doi.org/10.1016/j.nbd.2008.06.012
Xie J, Yang Y, Li J, Hou J, Xia K, Song W, Liu S (2014) Expression of Tmp21 in normal adult human tissues. Int J Clin Exp Med 7(9):2976–2983
Li X, Yihui Wu, Shen C, Belenkaya TY, Ray L, Lin X (2015) Drosophila P24 and Sec22 regulate wingless trafficking in the early secretory pathway. Biochem Biophys Res Commun 463(4):483–489. https://doi.org/10.1016/j.bbrc.2015.04.151
Port F, Hausmann G, Basler K (2011) A Genome-wide RNA interference screen uncovers two P24 proteins as regulators of wingless secretion. EMBO Rep 12(11):1144–1152. https://doi.org/10.1038/embor.2011.165
Ayers KL, Gallet A, Staccini-Lavenant L, Thérond PP (2010) The Long-Range activity of hedgehog is regulated in the apical extracellular space by the glypican dally and the hydrolase notum. Dev Cell 18(4):605–620. https://doi.org/10.1016/j.devcel.2010.02.015
Glashauser J, Camelo C, Hollmann M, Backer W, Jacobs T, Sanchez JI, Schleutker R et al (2023) Acute manipulation and real-time visualization of membrane trafficking and exocytosis in Drosophila. bioRxiv. https://doi.org/10.1101/2022.03.25.483021
Major EO, Miller AE, Mourrain P, Traub RG, de Widt E, Sever J (1985) Establishment of a line of human fetal glial cells that supports JC virus multiplication. Proc Natl Acad Sci 82(4):1257–1261. https://doi.org/10.1073/pnas.82.4.1257
Bonsergent E, Grisard E, Buchrieser J, Schwartz O, Théry C, Lavieu G (2021) Quantitative characterization of extracellular vesicle uptake and content delivery within mammalian cells. Nat Commun 12(1):1864. https://doi.org/10.1038/s41467-021-22126-y
Acknowledgements
We acknowledge the MRI imaging facility, member of the national infrastructure France-BioImaging, for advice and training. We thank Drs. Adrian Salic, Franck Perez, Gregory Lavieu and Robert Blum for the gift of constructs used in this study
Funding
This work was supported by the Agence Nationale de la Recherche (ANR-18-CE13-0003–01) to RG and PT. YB obtained a post-doctoral fellowship from Fondation pour la Recherche Médicale (FRM, SPF202110014043).
Author information
Authors and Affiliations
Contributions
Conceptualisation: YB, RG. Methodology: YB, TM, SV, PT, MSD, RG. Investigation: YB, TM, PT, RG. Analysis: YB, TM, PT, RG. Supervision: PT, RG. Writing—original draft: YB, RG. Writing—review & editing: YB, TM, SV, PT, MSD, RG.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Video S1: Dual-colour live-cell imaging of SHH-RUSH trafficking through the Golgi complex, related to Figure 1. SVG-A cell co-transfected with SHH-RUSH (magenta) and the medial Golgi marker, Mannosidase-II-EGFP (green) and imaged using 3D spinning disk confocal microscopy. Acquisition started 5 mins after the addition of 40 µM biotin. Frames were taken every 5 mins for 90 mins. Scale bar – 10µm
Video S2: Zoomed inset of Video S1, related to Figure 1. SHH-RUSH (magenta) transported into and exits from the Golgi (green). Frames taken every 5 mins for 90 min. Scale bar – 5 µm
Video S3: Two colour TIRF imaging of SHH-RUSH and GFP-Rab6a, related to Figure 2. SVG-A cell co-transfected with SHH-RUSH (magenta) and GFP-Rab6a (green). Acquisition started 30 mins after the addition of 40 µM biotin. Frames were taken every 2 seconds for an additional 30 mins. Scale bar – 10 µM
Video S4: Zoomed inset of Video S3, related to Figure 2. Video corresponds to 1890s to 1920s post biotin addition and demonstrates a double-positive vesicle of SHH-RUSH (magenta) and GFP-Rab6a (green) arriving at the plasma membrane. Scale bar – 2 µm
Video S5: Dual-colour live-cell imaging of SHH-RUSH and GFP-TMED10, related to Figure 3. SVG-A cell co-transfected with SHH-RUSH (magenta) and GFP-TMED10 (green) and imaged using 3D spinning disk confocal microscopy. Acquisition started immediately after the addition of 40 µM biotin. Frames were taken every 500 ms for 10 mins. Scale bar – 10 µM
Video S6: Zoomed inset of Video S5, related to Figure 3. A double-positive vesicle of SHH-RUSH (magenta) and GFP-TMED10 (green) trafficking towards a perinuclear region. Video corresponds to 49s to 99s post biotin addition. Scale bar – 5 µm
Video S7: Dual colour live-cell imaging of SHH-RUSH and SEC24D-EGFP, related to Figure 4. SVG-A cells co-transfected with SHH-RUSH (magenta) and SEC24D-EGFP (green) and imaged using 3D spinning disk confocal microscopy. Acquisition started immediately after the addition of 40 µM biotin. Frames were taken every 2 s for 15 min. Scale bar – 10 µm
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Bare, Y., Matusek, T., Vriz, S. et al. TMED10 mediates the loading of neosynthesised Sonic Hedgehog in COPII vesicles for efficient secretion and signalling. Cell. Mol. Life Sci. 80, 266 (2023). https://doi.org/10.1007/s00018-023-04918-1
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00018-023-04918-1