
Abstract
Dinosaur skin impressions are rare in the Upper Jurassic Morrison Formation, but different sites on the Howe Ranch in Wyoming (USA), comprising specimens from diplodocid, camarasaurid, allosaurid and stegosaurian dinosaurs, have proven to be a treasure-trove for these soft-tissue remains. Here we describe stegosaurian skin impressions from North America for the first time, as well as the first case of preservation of an impression of the integument that covered the dorsal plates of stegosaurian dinosaurs in life. Both have been found closely associated with bones of a specimen of the stegosaurian Hesperosaurus mjosi Carpenter, Miles and Cloward 2001. The scales of the skin impression of H. mjosi are very similar in shape and arrangement to those of Gigantspinosaurus sichuanensis Ouyang 1992, the only other stegosaurian dinosaur from which skin impressions have been described. Both taxa show a ground pattern of small polygonal scales, which in some places is interrupted by larger oval tubercles surrounded by the small scales, resulting in rosette-like structures. The respective phylogenetic positions of G. sichuanensis as a basal stegosaurian and H. mjosi as a derived form suggest that most stegosaurians had very similar skin structures, which also match the most common textures known in dinosaurs. The integumentary impression from the dorsal plate brings new data to the long-lasting debate concerning the function of dorsal plates in stegosaurian dinosaurs. Unlike usual dinosaur skin impressions, the integument covering the dorsal plates does not show any scale-like texture. It is smooth with long and parallel, shallow grooves, a structure that is interpreted as representing a keratinous covering of the plates. The presence of such a keratinous covering has affects on all the existing theories concerning the function of stegosaurian plates, including defense, thermoregulation, and display, but does not permit to rule out any of them.
Similar content being viewed by others


Introduction
Reports of skin impressions from the Upper Jurassic Morrison Formation are rare, although it represents one of the most dinosaur fossil-rich Mesozoic strata and despite the fact that it has been explored intensely for over a century (Dodson et al. 1980; Foster 2003; Turner and Peterson 2004). To our knowledge, the only published descriptions of dinosaur skin impressions from the Morrison Formation are associated with a juvenile allosaurid from the Meilyn Quarry in Southeastern Wyoming (Pinegar et al. 2003), with the skeletons of juvenile diplodocids from the Mother’s Day Quarry in South Central Montana (Myers and Storrs 2007), with Barosaurus lentus Marsh 1890 remains from Dinosaur National Monument in Northwestern Colorado (White 1967), with sauropod footprints from the Bighorn Basin of North Central Wyoming (Platt and Hasiotis 2006), as well as loose Dinosauria indet. skin pieces from the Mygatt-Moore Quarry in central western Colorado (Kirkland and Carpenter 1994).
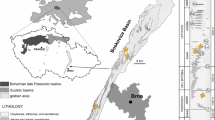
However, the quarries at the Howe Ranch near Shell, Wyoming, USA (Fig. 1), are an exception to this rule in being remarkably productive for integumentary impressions. Sauropod skin impressions from the Howe Ranch were first reported by Brown (1935) after his excavations at the Howe Quarry in 1934. During their annual field campaigns from 1990 to 2003 at the same locality, the excavation team of the Sauriermuseum Aathal (SMA) found a large number of dinosaur skin impressions in different quarries on the Howe Ranch. Most of these were isolated pieces without any connection to bones. However, some pieces were found associated with the caudal vertebrae of a diplodocid sauropod, revealing the presence of triangular integumentary appendages on the dorsal side of the animal, and thus influencing the way diplodocid sauropods are now reconstructed (Czerkas 1992). Other skin impressions found by the SMA include skin impressions from various parts of a Camarasaurus Cope 1877 (Tschopp 2008), as well as from an Allosaurus Marsh 1877 (which could not be recovered during excavation; H.-J. Siber, pers. comm. 2009) and stegosaurian skin impressions (described herein).

Location of the Howe Ranch quarries (star) in central northern Wyoming (USA). Scale bar 100 km
Reports of skin impressions from Thyreophora are very rare. As far as we know, the only thyreophoran specimens exhibiting such preservation are: Gigantspinosaurus sichuanensis ZDM 0019 (Xing et al. 2008), cf. Scelidosaurus Owen 1860 (BRSMG CF2781; Martill et al. 2000), an unpublished juvenile specimen cf. Scelidosaurus sp. (BRSM 12785), an unpublished find of a cf. Stegosaurus Marsh 1877 from Bone Cabin Quarry West in Southeastern Wyoming (H.-J. Siber, pers. comm. 2009; K. Carpenter, pers. comm. 2010), and the present specimen of Hesperosaurus mjosi. Other thyreophoran skin impressions have been found in connection with tracks of both stegosaurians (Mateus et al. 2010) and ankylosaurs (McCrea et al. 2001). Furthermore, osteoderms with a scale-like shape have been reported in a number of ankylosaurs (e.g., Penkalski 2001) and in Stegosaurus stenops Marsh 1887 (e.g., Gilmore 1914), but these do not represent the surface of the skin, although studies based on extant archosaurs have shown that they are generally overlaid by a scale of similar dimensions and morphology (Vickaryous and Sire 2009).
Geographical and geological context
Locality
The specimen with skin impressions (SMA 0018, nicknamed “Victoria”) was discovered in 1995 on the Howe Ranch, approximately 15 km north of Shell in central northern Wyoming, USA (Fig. 1). It was found at the Howe-Stephens Quarry (44°39′N, 107°49′W), 450 m southwest of the historic Howe Quarry, which was excavated during the Sinclair Dinosaur Expedition of the American Museum of Natural History in 1934 (Brown 1935; Ayer 2000).
Stratigraphy
The quarry is located in the middle part of the Upper Jurassic Morrison Formation below the so-called clay change. Like the historic Howe Quarry, which is approximately 10 m lower in the stratigraphical column, the Howe-Stephens Quarry is among the geologically oldest fossil sites known from the Morrison Formation (Turner and Peterson 1999; Foster 2003; Ikejiri 2005; Schwarz et al. 2007).
Sedimentology, palaeoenvironment and taphonomy
The Howe-Stephens Quarry has a horizontal extension of 18 × 24 m, and a maximal vertical extension of approximately 8 m (H.-J. Siber, pers. comm. 2003; pers. obs. 2005). The sediments are fine-grained fluvial sandstone exhibiting cross-bedding in some layers (Schwarz et al. 2007; J. Ayer, pers. comm. 2007). The quarry is an exception within the Morrison Formation as it contains abundant plant material, including two very large silicified tree logs, plenty of carbonized plant fragments, as well as numerous carbonized branches (Ayer 2000; pers. obs. 2003). Most of the specimens are preserved in an area of 10 × 12 m, in a layer that is only 1 m thick, and it has been hypothesized that the tree logs blocked the dinosaur carcasses during an interval of heavy flooding and thereby allowed for a rapid burial of the specimens (Ayer 2000). The latter is generally thought to be crucial for the preservation of articulated and almost complete skeletons, as well as skin impressions.
The reported specimen of Hesperosaurus mjosi was found lying on its right side (Fig. 2) and the preservation of that side of the specimen is far superior to that of the left side, as can be seen from the differential preservation of the mani, pes and ribcage (Siber and Möckli 2009). There was also variation in the degree of articulation within the skeleton, with some parts found in full articulation, whereas other parts were found disarticulated and some lighter elements were missing (Fig. 2). This seems to indicate that the specimen was only partly buried in a first time, while other parts were still exposed to the elements and that the complete burial of the specimen only happened in a second instant. Skin impressions where only found on the specimens underside, which is though to have been buried more rapidly.

Quarry map of Hesperosaurus mjosi (SMA 0018). The areas where integument impressions have been found are marked in grey. Drawing by Esther Premru, modified from Siber and Möckli (2009). Scale bar 1 m
The dinosaur faunal assemblage found in the Howe-Stephens Quarry includes all of the most common dinosaurs of the Morrison Formation. Besides Hesperosaurus mjosi, it includes a virtually complete articulated skeleton of Camarasaurus sp. (Tschopp 2008), an articulated post-cranial skeleton of a juvenile diplodocid (Schwarz et al. 2007), an almost complete skeleton of Allosaurus sp., a partial skeleton of Othnielosaurus Galton 2006 and several subadult to adult specimens of cf. Diplodocus Marsh 1878 (H.-J. Siber, pers. comm. 2003; pers. obs. 2005).
Material
Taxonomy
The reported specimen (SMA 0018, “Victoria”) is a well-preserved skeleton including the skull and most of the post-cranial skeleton (Fig. 2). SMA 0018 is identified as Hesperosaurus mjosi, based on the transverse processes on anterior caudal vertebrae projecting ventrally rather than laterally, the postzygapophyses on posterior cervical vertebrae being elongated posteriorly and overhanging the back of the centrum, the rectangular acromial process of the scapula, the neural arches of the dorsal vertebrae are not elongated above the neural canal, the cervical dermal plates that are longer anteroposteriorly than tall dorsoventrally, and the neural spines of the proximal caudal vertebrae that are enlarged transversely, but not bifurcated (Carpenter et al. 2001; Maidment et al. 2008; Carpenter 2010). Maidment et al. (2008) also referred the specimen to Hesperosaurus mjosi, but quoted a wrong institutional number (SMA V03 instead of SMA 0018).
Maidment et al. (2008) questioned the erection of the genus Hesperosaurus by Carpenter et al. (2001), arguing that H. mjosi is congeneric with Stegosaurus armatus Marsh 1877, based on a high degree of similarity between the two taxa. However, in the same paper, Maidment et al. (2008) report seven differences in character states between S. armatus and H. mjosi and three autapomorphies for H. mjosi. In our opinion, ten distinguishable differences in morphology are too numerous to consider the two taxa congeneric, even though they are sister groups in recent phylogenetic analyses (Maidment et al. 2008; Mateus et al. 2009). We therefore consider Hesperosaurus to be a valid genus, in agreement with Carpenter (2010).
Preservation of integument impressions
Integumentary impressions are preserved in several areas on the underside of SMA 0018 (Fig. 2). Several large pieces were found during preparation of a field jacket containing the anterior right side of the rib cage (Fig. 3). Some of them have been preserved in situ, while others have been removed in order to see the bones (Siber and Möckli 2009). Another smaller piece was found attached to a dorsal plate lying under the left ribs. Its preservation as a negative imprint reveals that it is from the skin covering the distal part of the left side ribs, which became dislocated onto the plate during the decomposition of the animal (Fig. 2). Below this skin impression, a second integument impression was found, this time pertaining to the structure that covered the dorsal plate.

View of the anterior part of the right side of the ribcage and the posterior end of the right scapula, showing the position of the skin impressions. Rectangles mark impressions of small polygonal scales forming the ground pattern; circles mark rosettes. Scale bar 10 cm
The integument impressions are preserved in three different modes: (1) as cross-sections at various angles, (2) as natural molds, and (3) as natural casts. The natural casts have the appearance of fossil skin, but they are made up of sedimentary rock and have no inner structure that could justify their identification as a permineralization of the actual skin. We therefore interpret them as infills of natural molds. The skin impressions are preserved in all the above-mentioned manners, whereas the plate impressions only exist as natural molds.
A thin and friable, dark brown to black, layer is found intercalated between the natural mold and the natural cast, both on the skin and the plate impressions. Such a dark layer is associated with various integument impression found in the Howe Ranch quarries (Brown 1935; Czerkas 1992; Tschopp 2008; pers. obs. 2007), but has never been observed in association with the bones or isolated within the sediment (pers. obs. 2007). Both Brown (1935) and Czerkas (1992) interpreted the layer as the possible remains of the actual epidermis, but did not study it thoroughly. However, different finds of similar thin layers covering dinosaur integumentary impressions have been interpreted as microbial mats (e.g., Keller 1992; Briggs et al. 1997) or as a result of authigenic mineralization, in which the replication of the tissue morphology in form of a layer of minerals is the product of decay bacteria (Briggs 2003). Identifying the origin of the dark layer through microstructural and chemical studies is not the scope of this paper, but would be a crucial step in understanding the process that created the fossil and in perceiving the exact nature of the structures that we are observing.
Description
Skin impressions
The skin impressions mainly consist of small, non-imbricating, polygonal (predominantly hexagonal) tuberculate scales, which make up the ground pattern of the integument on the anterior part of the rib cage. Their diameters range from 2 to 7 mm with the majority ranging from 4 to 5 mm in diameter. The single tubercles are closely spaced, with shallow and narrow grooves separating them. A relatively ordered, nearly linear pattern of the scales can be seen on the ventral part of the ribcage (Fig. 4), but on its dorsal part, this pattern is lost and the distribution of the scales is more random.

Skin impression from the ventral part of the body showing a nearly linear arrangement of the small polygonal scales of the ground pattern. Preserved as a natural mold. Scale bar 2 cm
A much rarer and larger second type of scale is present towards the dorsal side of the animal. These are higher, domed, five to ten times larger than the small scales, and ellipsoid in outline (Fig. 5). Each of them is surrounded by 9–13 small tubercles, which are not distinguishable from the scales forming the ground pattern. Only two such structures can be confidently identified, and thus a specific arrangement of these is therefore not discernable. The larger of the two scales is 20 × 15 mm, whereas the other is 10 × 8 mm. The surface of both types of scales does not appear to be sculptured.

Skin impressions from the dorsal part of the body showing a rosette. Preserved as a natural mold. Scale bar 1 cm
Plate cover impression
The plate cover impression is preserved only as a natural mold. The single largest preserved surface measures approximately 200 cm2. Together with the other pieces, it leaves little doubt that most of the upper part of the dorsal plate was covered by this structure. The plate cover impression is uniformly flat and smooth with low parallel ridges running ventrodorsally (Fig. 6). The length of the ridges is typically 2–3 cm and although they have variable heights, none of them exceed 0.5 mm. In some areas, the ridges are as close as 2 mm, whereas they are usually more separated from each other. No scaly structures are visible. Given that the plate cover impression is a mold, the ridges would in reality have been shallow grooves on the surface of the integument.

Detail of the impression of the presumed keratinous covering of the plate, showing low parallel ridges extending from the upper left to the lower right corner. Note the remains of the dark layer that cover the impression in the central lower part of the picture. Preserved as a natural cast. Scale bar 1 cm
Comparison and discussion
Scales of the ground pattern
The small polygonal scales that make up the ground pattern of the skin impressions in Hesperosaurus mjosi are common to all known ornithischian skin impressions and are usually pentagonal or hexagonal (Czerkas 1997). The size of these scales varies among different taxa, but in many taxa the small scales building up the ground pattern are 5–7 mm in diameter like those in stegosaurians. A notable variation from this is the ceratopsian Chasmosaurus belli Lambe 1914 (Sternberg 1925; Czerkas 1997) that has much larger scales with a diameter of approximately 20–25 mm.
Few skin impressions have been attributed to saurischian dinosaurs, but these finds indicate a larger variety of shapes of the scales that make up the ground pattern of the skin in this clade. In addition to the classic polygonal pattern known from brachiosaurids (Hooley 1917) and diplodocids (Czerkas 1992; Mateus and Milàn 2009), a ground pattern of round scales has been reported in both sauropods (Powell 1980; Chiappe et al. 1998) and theropods (Bonaparte et al. 1990; Gatesy et al. 2005). Furthermore, linked sub-rectangular scales forming lines (Chiappe et al. 1998), as well as large, 20–30 mm broad, polygonal scales forming a ground pattern in tracks, have also been reported in sauropods (e.g., Mateus and Milàn 2009).
Comparison of the skin structures of Hesperosaurus mjosi (SMA 0018) to the one described from Gigantspinosaurus sichuanensis ZDM 0019 (Xing et al. 2008) indicates only minor differences. The small polygonal scales exhibit maximum diameters of about 5.2–7 mm in the H. mjosi (SMA 0018), whereas Xing et al. (2008) report 5.7–9.2 mm scale diameters in G. sichuanensis (ZDM 0019). The grooves separating the single scales appear to be broader in some areas in G. sichuanensis (Xing et al. 2008: fig. 2), but this could be the result of local stretching of the skin, since other areas do not show this difference (Xing et al. 2008: fig. 1).
Rosettes
The second structure that can be recognized in the skin impression of Hesperosaurus mjosi are isolated larger ellipsoid scales surrounded by the small polygonal scales of the ground pattern, thus forming rosettes. Rosettes consisting of 13–14 smaller scales surrounding a larger one have also been reported in Gigantspinosaurus sichuanensis (Xing et al. 2008). This kind of skin pattern is widely distributed among dinosaurs (e.g., Ornithopoda: Brown 1916; Ceratopsia: Sternberg 1925; Theropoda: Bonaparte et al. 1990; and Sauropoda: Chiappe et al. 1998), and can therefore—together with the ground pattern of small, non-imbricating scales—be hypothesized to represent the plesiomorphic state of the structure of the dinosaurian epidermis. As with the size of the small scales, the dimension of the larger ones varies among different taxa. However, the size ratio between the small scales of the ground pattern and the large scales of the rosettes remains relatively constant, and in all the dinosaurian taxa for which rosettes are known, approximately 10–20 smaller scales pertaining to the ground pattern surround the large scales.
Due to the restricted areas of the skin impression surfaces that have been preserved in both H. mjosi and G. sichuanensis, a specific arrangement of the rosettes in stegosaurians cannot be described. However, the two rosettes identified in SMA 0018 are found at a similar height on the ribcage, and it is possible that they were arranged in irregular longitudinal rows as can be seen in Carnotaurus sastrei Bonaparte, 1985 (Bonaparte et al. 1990), Corythosaurus casuarius Brown 1914 (Brown 1916), Chasmosaurus belli (Sternberg 1925), and Styracosaurus albertensis Lambe 1913 (Brown 1917). In H. mjosi (SMA 0018), the rosettes are found on the dorsal part of the ribcage, and the same is the case for G. sichuanensis (ZDM 0019; Xing et al. 2008), whereas the skin impressions found on the lower part of the ribcage only contain small polygonal scales. This distribution is not unique to stegosaurians, but is according to Czerkas (1997) and Sternberg (1925) a general pattern found in many dinosaurs. Sternberg (1925) noted that rosettes are more common and larger on the dorsal region of the body of ceratopsian dinosaurs and decrease in size and frequency ventrally, with the belly skin being composed only of the small polygonal scales of the ground pattern.
Integumentary covering of the dorsal plates
Dinosaur skin impression typically consists of tuberculate scales. Although exceptions are known, no impressions of skin structures comparable to the impressions found on the dorsal plates of SMA 0018 have been reported in any dinosaur. A scale-less skin impression is known from Pelecanimimus polyodon Perez-Moreno, Sanz, Buscalioni, Moratalla, Ortega, & Rasskin-Gutman 1994 (Briggs et al. 1997), but it shows cross-hatching lines similar to mammalian skin, rather than long, straight, and parallel grooves (Fig. 6). Also, the uniformly smooth structure, the lack of folds or waves and the thin straight grooves in the integument covering the plates of SMA 0018 seem to indicate a flat covering with little or no plasticity.
Although we do not have direct proof that this integument was a keratinous covering, there are indirect lines of evidence, since a keratinous covering is the only hard integument that is known to cover such a large surface in vertebrates and that the neurovascular sulci on the surface of the bone are highly indicative of a keratinous covering (Hieronymus et al. 2009).
The function of the dorsal plates in stegosaurians
Previous works have almost exclusively concentrated on the genus Stegosaurus, but the dorsal plates of Hesperosaurus have a morphology and distribution that is so similar to those of Stegosaurus that hypotheses on their function are likely valid for both genera. These theories include the use of dorsal plates for display (Carpenter 1998; Main et al. 2005; Hayashi et al. 2009), thermoregulation (Farlow et al. 1976; Buffrénil et al. 1986; Farlow et al. 2010) and defence (Marsh 1877; Gilmore 1914; Bakker 1986, Mallison 2010). The fact that the plates had a keratinous covering has implications on all these hypotheses, and must therefore be considered when evaluating these suggestions.
Defence
The function of stegosaurian dorsal plates as defensive structures has been rejected by several authors, based on their internal trabecular structure, which is not considered crush resistant, and would therefore make a defensive use ineffective (Buffrénil et al. 1986; Main et al. 2005). The histological study of Buffrénil et al. (1986) also did not find an asymmetrical distribution of Sharpey’s fibres within the plates that would have indicated the possibility of moving the plates to a recumbent and therefore more protective position, as suggested by Bakker (1986). However, the keratinous covering could have increased their strength considerably, depending on its thickness, since beta-keratin, which makes up scales, claws, and other cornified structures in reptiles, is considered to be one of the strongest natural proteins (Baden et al. 1974). The keratinous covering could also have created much sharper edges on the plates than what is seen on the osteoderms and thereby increase their value as defensive structures, even in an upright position. Although the protection of the upright plates might not have been needed over the spinal cord in the pelvic and thoracic regions as pointed out by Bakker (1986), it would have been important protection for one of the most exposed regions of the animal, the neck. The cervical plates would have greatly increased the height of the neck making a lateral attack more difficult and could have been an effective defence against a dorsal attack on the neck, which a reported cervical plate with a possible bite mark (Carpenter et al. 2005) might prove.
Display
The theory that the dorsal plates of stegosaurians were used for inter- or intraspecific display is the most recognized theory amongst recent authors (Carpenter 1998; Main et al. 2005; Hayashi et al. 2009). The main evidence supporting this hypothesis are the size of the plates and their asymmetrical positioning in at least Stegosaurus and Hesperosaurus, which increases the outline of the animal considerably. The fact that the plates had a keratinous covering corroborates this theory, since it is likely to have increased the surface area of the osteoderms significantly, as seen on horns and claws, thereby enhancing the visual effect in lateral view. Moreover, keratinous covering in reptiles and birds, like the beaks of birds, are often coloured and interpreted as being used for display (e.g., Tattersall et al. 2009).
Thermoregulation
A thermoregulatory function for the plates has been discussed extensively (e.g., Farlow et al. 1976; Buffrénil et al. 1986; Main et al. 2005; Hayashi et al. 2009; Farlow et al. 2010), with recent studies accepting a possible exaptation of the plates for this use, although they do not consider it the main function (Main et al. 2005; Hayashi et al. 2009; Farlow et al. 2010). As noted by Farlow et al. (1976), a thermoregulatory use would not be optimized through the existence of a keratinous covering. The keratinous covering would reduce the effectiveness of the heat transfer between the blood and the environment, to an extent that would depend on its thickness. Moreover, if the use of forced convection is hypothesized (Farlow et al. 1976), the larger the surface is, the more efficient is the heat transfer. Thus, an irregular and pitted surface would be more optimal than the flat surface present in the integument impressions of H. mjosi. It must, however, be noted that a thermoregulatory function cannot be excluded by the existence of a keratinous covering, since keratin covered organs have been shown to have a thermoregulatory function in the case of bovid horns (Picard et al. 1996, 1999; Hoefs 2000) and bird beaks (ducks: Hagan and Heath 1980; toucans: Tattersall et al. 2009).
Conclusions
A ground pattern of small, polygonal, non-imbricating scales and rosettes with larger oval scales at their centre is observed in both Hesperosaurus mjosi and Gigantspinosaurus sichuanensis. Given that recent phylogenetic analyses (Maidment et al. 2008; Mateus et al. 2009) have shown G. sichuanensis to be one of the most basal and H. mjosi to be one of the most derived taxa within Stegosauria, a similar epidermal structure can be inferred for other stegosaurians by means of phylogenetic bracketing. This pattern exhibiting small non-imbricating scales and rosettes is also hypothesized to be the plesiomorphic state of the dinosaurian integument, due to its existence in all of the major higher-level dinosaurian taxa.
The dorsal plates of stegosaurians are shown to have had a keratinous covering over most of their surface. This discovery has implications for all the hypotheses that have been put forward concerning the function of the plates, but none of them can be rejected on this ground. However, future knowledge concerning the thickness of the keratinous covering might help to discriminate amongst them, as a thick covering may favour a defensive use, whereas a thin covering may favour a thermoregulatory function. Finally, multiple functions of stegosaurian dorsal plates should not be excluded nor the possibility that the importance of different functions could have varied amongst different stegosaurian species.
Abbreviations
- BRSM:
-
Bristol’s City Museum and Art Gallery, England
- SMA:
-
Sauriermuseum Aathal, Switzerland
- ZDM:
-
Zigong Dinosaur Museum, China
References
Ayer, J. (2000). The Howe Ranch Dinosaurs. Aathal, Switzerland: Sauriermuseum Aathal. 96 pp.
Baden, H. P., Sviokla, S., & Roth, I. (1974). The structural protein of reptilian scales. Journal of Experimental Zoology, 187, 287–294.
Bakker, R. T. (1986). The Dinosaur Heresies: New theories unlocking the mystery of the dinosaurs and their extinction. New York: William Morrow, 481 pp
Bonaparte, J. F., Novas, F. E., & Coria, R. A. (1990). Carnotaurus sastrei Bonaparte, the horned, lightly build Carnosaur from the Middle Cretaceous of Patagonia. Natural History Museum of Los Angeles County, Contributions in Science, 416, 1–42.
Briggs, D. E. G. (2003). The role of decay and mineralization in the preservation of soft-bodied fossils. Annual Review of Earth and Planetary Sciences, 31, 275–301.
Briggs, D. E. G., Wilby, P. R., Pérez-Moreno, B. P., Sanz, J. L., & Fregenal-Martínez, M. (1997). The mineralization of dinosaur soft tissue in the Lower Cretaceous of Las Hoyas, Spain. Journal of the Geological Society, 154, 587–588.
Brown, B. (1916). Corythosaurus casuarius: Skeleton, musculature and epidermis. Bulletin of the American Museum of Natural History, 35, 709–716.
Brown, B. (1917). A complete skeleton of the horned dinosaur Monoclonius, and description of a second skeleton showing skin impressions. Bulletin of the American Museum of Natural History, 37, 281–306.
Brown, B. (1935). Sinclair dinosaur expedition, 1934. Natural History, 36, 3–15.
Buffrénil, V. de., Farlow, J.O., & Ricqlès, A.de. (1986). Growth and function of Stegosaurus plates: Evidence from bone histology. Paleobiology, 12, 459–473.
Carpenter, K. (1998). Armor of Stegosaurus stenops, and the taphonomic history of a new specimen from Garden Park, Colorado. Modern Geology, 23, 127–144.
Carpenter, K. (2010). Species concept in North American stegosaurs . Swiss Journal of Geosciences 103 (this volume). doi:10.1007/s00015-010-0020-6
Carpenter, K., Miles, C., & Cloward, K. (2001). New primitive stegosaur from the Morrison Formation, Wyoming. In K. Carpenter (Ed.), The Armored Dinosaurs (pp. 55–75). Bloomington: Indiana University Press.
Carpenter, K., Sanders, F., McWhinney, L., & Wood, L. (2005). Evidence for predator–prey relationships: Example for Allosaurus and Stegosaurus. In K. Carpenter (Ed.), The Carnivorous Dinosaurs (pp. 325–350). Bloomington: Indiana University Press.
Chiappe, L. M., Coria, R. A., Dingus, L., Jackson, F., Chinsamy, A., & Fox, M. (1998). Sauropod dinosaur embryos from the Late Cretaceous of Patagonia. Nature, 396, 258–261.
Czerkas, S. (1992). Discovery of dermal spines reveals new look for sauropod dinosaurs. Geology, 20, 1068–1070.
Czerkas, S. (1997). Skin. In P. J. Currie & K. Padian (Eds.), Encyclopedia of dinosaurs (pp. 669–675). San Diego: Academic Press.
Dodson, P., Behrensmeyer, A. K., Bakker, R. T., & McIntosh, J. S. (1980). Taphonomy and paleoecology of the dinosaur beds of the Jurassic Morrison Formation. Paleobiology, 6, 208–232.
Farlow, J. O., Hayashi, S., & Tattersall, G. J. (2010). Internal vascularity of the dermal plates of Stegosaurus (Ornithischia, Thyreophora). Swiss Journal of Geosciences 103 (this volume). doi:10.1007/s00015-010-0021-5
Farlow, J. O., Thompson, C., & Rosner, D. (1976). Plates of the dinosaur Stegosaurus: Forced convection heat loss fins? Science, 192, 1123–1125.
Foster, J. R. (2003). Paleoecological analysis of the vertebrate fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain Region, USA. New Mexico Museum of Natural History & Science Bulletin, 23, 1–95.
Gatesy, S. M., Shubin, N. H., & Jenkins, F. A., Jr. (2005). Anaglyph stereo imaging of dinosaur track morphology and microtopography. Palaeontologia Electronica, 8, 1–12.
Gilmore, C. W. (1914). Osteology of the armored Dinosauria in the United States National Museum, with special reference to the genus Stegosaurus. United States National Museum Bulletin, 89, 1–143.
Hagan, A., & Heath, J. (1980). Regulation of heat loss in the duck by vasomotion in the bill. Journal of Thermal Biology, 5, 95–101.
Hayashi, S., Carpenter, K., & Suzuki, D. (2009). Different growth patterns between the skeleton and osteoderms of Stegosaurus (Ornithischia: Thyreophora). Journal of Vertebrate Paleontology, 29, 123–131.
Hieronymus, T. L., Witmer, L. M., Tanke, D. H., & Currie, P. J. (2009). The facial integument of Centrosaurine Ceratopsids: Morphological and histological correlates of novel skin structures. The Anatomical Record, 292, 1370–1396.
Hoefs, M. (2000). The thermoregulatory potential of Ovis horn cores. Canadian Journal of Zoology, 78, 1419–1426.
Hooley, R. W. (1917). On the integument of Iguanodon bernissartensis Boulenger, and of Morosaurus becklesii Mantell. Geological Magazine, 6, 148–150.
Ikejiri, T. (2005). Distribution and Biochronology of Camarasaurus (Dinosauria, Sauropoda) from the Jurassic Morrison Formation in the Rocky Mountain Region. In New Mexico Geological Society, 56th Field Conference Guidebook, Geology of the Chama Basin (pp. 367–379).
Keller, T. (1992). «Weichteil-Erhaltung» bei grösseren Vertebraten (Ichthyosauriern) des Posidonienschiefers Holzmadens (Oberer Lias, Mesozoikum Süddeutschlands). Kaupia, 1, 23–63.
Kirkland, J. I., & Carpenter, K. (1994). North America’s first pre-Cretaceous ankylosaur (Dinosauria) from the Upper Jurassic Morrison Formation of Western Colorado. Brigham Young University Geological Studies, 40, 25–42.
Maidment, S., Norman, D., Barrett, P., & Upchurch, P. (2008). Systematics and phylogeny of Stegosauria (Dinosauria: Ornithischia). Journal of Systematic Paleontology, 6, 367–407.
Main, R., Ricqlès, A. de, Horner, J., & Padian, K. (2005). The evolution and function of thyreophoran dinosaur scutes: Implications for plate function in stegosaurs. Paleobiology, 31, 291–314.
Mallison, H. (2010). CAD assessment of the posture and range of motion of Kentrosaurus aethiopicus HENNIG 1915. Swiss Journal of Geosciences, 103 (this volume). doi:10.1007/s00015-010-0024-2
Marsh, O. C. (1877). A new order of extinct Reptilia (Stegosauria) from the Jurassic of the Rocky Mountains. American Journal of Science, 14(3rd series), 513–514.
Marsh, O. C. (1890). Description of new dinosaurian reptiles. American Journal of Science 38(3rd series), 81–86.
Martill, D. M., Batten, D. J., & Loydell, D. K. (2000). A new specimen of the thyreophoran dinosaur cf. Scelidosaurus with soft tissue preservation. Palaeontology, 43, 549–559.
Mateus, O., Maidment, S. C. R., & Christiansen, N. A. (2009). A new long-necked ‘sauropod-mimic’ stegosaur and the evolution of the plated dinosaurs. Proceedings of the Royal Society B, 276, 1815–1821.
Mateus, O., & Milàn, J. (2009). A divers Upper Jurassic dinosaur ichnofauna from central-west Portugal. Lethaia, 43, 245–257.
Mateus, O., Milàn, J., Romano, M., & Whyte, M. A. (2010). New finds of stegosaurid tracks from the Upper Jurassic Lourinhã Group, central-west Portugal: Implications for the spatial, temporal and palaeoecological distribution of the morphotype Deltapodus. Acta Palaeontologica Polonica (in press).
McCrea, R. T., Lockley, M. G., & Meyer C. A. (2001). Global distribution of purported ankylosaur track occurrences. In K. Carpenter (Ed.), The Armored Dinosaurs (pp. 413–454). Bloomington: Indiana University Press.
Myers, T. S., & Storrs, G. W. (2007). Taphonomy of the Mother’s Day Quarry, Upper Jurassic Morrison Formation, South-Central Montana, USA. Palaios, 22, 5–123.
Penkalski, P. (2001). Variation in specimens referred to Euoplocephalus tutus. In K. Carpenter (Ed.), The Armored Dinosaurs (pp. 261–297). Bloomington: Indiana University Press.
Picard, K., Festa-Bianchet, M., & Thomas, D. (1996). The cost of horniness: Heat loss may counter sexual selection for large horns in temperate bovids. Ecoscience, 3, 280–284.
Picard, K., Thomas, D., Festa-Bianchet, M., Belleville, F., & Laneville, A. (1999). Differences in the thermal conductance of tropical and temperate bovid horns. Ecoscience, 6, 148–158.
Pinegar, R. T., Loewen, M. A., Cloward, K. C., Hunter, R. J., & Weege, C. J. (2003). A juvenile allosaur with preserved integument from the basal Morrison Formation of Central Wyoming. Journal of Vertebrate Paleontology, 23(Suppl. 3), 87A–88A.
Platt, B. F., & Hasiotis, S. T. (2006). Newly discovered sauropod dinosaur tracks with skin and foot-pad impressions from the Upper Jurassic Morrison Formation, Bighorn Basin, Wyoming, USA. Palaios, 21, 249–261.
Powell, J. E. (1980). Sobre la presencia de una armadura dermica en algunos dinosaurios titanosauridos. Acta Geologica Lilloana, 15, 41–47.
Schwarz, D., Ikejiri, T., Breithaupt, B. H., Sander, P. M., & Klein, N. (2007). A nearly complete skeleton of an early juvenile diplodocid (Dinosauria: Sauropoda) from the Lower Morrison Formation (Late Jurassic) of north central Wyoming and its implications for early ontogeny and pneumaticity in sauropods. Historical Biology, 19, 225–253.
Siber, H. J. & Möckli, U. (2009). The Stegosaurs of the Sauriermuseum Aathal. Aathal, 56 pp.
Sternberg, C. M. (1925). Integument of Chasmosaurus belli. Canadian Field Naturalist, 39, 108–110.
Tattersall, G., Andrade, D., & Abe, A. (2009). Heat exchange from the toucan bill reveals a controllable vascular thermal radiator. Science, 325, 468–470.
Tschopp, E. (2008). The complete set of autopodia of the Camarasaurus SMA 0002 and what it can tell us about systematics, taphonomy, ontogeny, and footprint shape. Unpublished Master Thesis, University of Zurich, Switzerland, 185 pp.
Turner, C. E. & Peterson, F. (1999). Biostratigraphy of dinosaurs in the Upper Jurassic Morrison Formation of the Western Interior, USA. In D. D. Gillette. (Ed.), Vertebrate Paleontology in Utah (pp. 77–114). Utah Geological Survey Miscellaneous Publication, no. 99-1, Utah Geological Survey, Salt Lake City.
Turner, C. E., & Peterson, F. (2004). Reconstruction of the Upper Jurassic Morrison Formation extinct ecosystem—a synthesis. Sedimentary Geology, 167, 309–355.
Vickaryous, M. K., & Sire, J.-Y. (2009). The integumentary skeleton of tetrapods: Origin, evolution, and development. Journal of Anatomy, 214, 441–464.
White, T. E. (1967). Dinosaurs at home. New York: Vantage Press, 232 pp
Xing, L. D., Peng, G. Z., & Shu, C. K. (2008). Stegosaurian skin impressions from the Upper Jurassic Shangshaximiao Formation, Zigong, Sichuan, China: A new observation. Geological Bulletin of China, 27, 1049–1053. (in Chinese with English abstract).
Acknowledgments
We thank Hans-Jakob Siber (Sauriermuseum Aathal) for having invited us and encouraged our study on the stegosaurian integument impressions of the SMA. We are also grateful to the field team of SMA for having found and preserved the integument impressions, notably to the preparators Ben Pabst and Esther Premru. The manuscript benefited greatly from the comments and suggestions of Octávio Mateus (Universidade Nova de Lisboa; Museu da Lourinhã, Portugal), Rui Castanhinha (Instituto Gulbenkian de Ciência; Museu da Lourinhã, Portugal), James Farlow (Indiana University, Fort Wayne, Indiana, USA), and Stephen Brusatte (American Museum of Natural History, New York, USA) who kindly reviewed earlier versions of the manuscript, as well as of those of the two reviewers Matt Vickaryous (Department of Biomedical Sciences, University of Guelph, Ontario, Canada) and Kenneth Carpenter (College of Eastern Utah Prehistoric Museum, Price, Utah, USA). We are grateful to James Farlow, Shoji Hayashi, Octávio Mateus, Jesper Milàn, Kenneth Carpenter as well as Jacques Ayer for sharing their unpublished manuscripts with us, and for letting us incorporate some of their results into our work. We also thank Hans-Jakob Siber and Esther Premru for the permission to use their quarry map of SMA 0018, and Urs Möckli for providing us with high quality photos of the skin impressions.
Author information
Authors and Affiliations
Corresponding author
Additional information
Editorial handling: Jean-Paul Billon-Bruyat & Daniel Marty.
Rights and permissions
About this article
Cite this article
Christiansen, N.A., Tschopp, E. Exceptional stegosaur integument impressions from the Upper Jurassic Morrison Formation of Wyoming. Swiss J Geosci 103, 163–171 (2010). https://doi.org/10.1007/s00015-010-0026-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00015-010-0026-0