
Summary
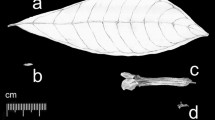
The genomic formula ACD of the hexaploid oat,Avena saliva, AC for the tetraploid speciesA. magna andA. murphyi, and the hypothesis that the A genome was contributed byA. strigosa group and C genome byA. ventricosa—A. eriantha group, have derived from karyotype studies. Cytogenetic evidence, however, does not support any of the known diploid oats as possible progenitors of the hexaploid oats.A. magna, and to a lesser extent,A. murphyi, are closer to the hexaploids than other tetraploids. Morphologically, these tetraploids resemble thesterilis type, which is apparently the genuine form of the hexaploid oats, with a few important differences. Spikelet morphology of these three wild oats indicates some features that must also occur in one or more of their diploid progenitors: 1. Ventricose diaspore containing the entire spikelet, 2. Long and broad callus with blunt tip, 3. Awn insertion at the lower one quarter of the lemma, 4. Close proximity of lemma tips of the two lower florets.A. magna andA. murphyi have narrow ecological ranges and are restricted to heavy alluvial soils which are now mostly under cultivation. It is likely that on similar soil type the diploid progenitors would grow as well and a search for them should be attempted there. Morocco is the most promising area for finding the missing diploids. Not onlyA. magna andA. murphyi grow there, but except one, all other annual oat species as well.
Similar content being viewed by others


References
Baum, B.R., 1977. Oats, Wild and Cultivated. Print. and Publ. Suppl. and Sev., Ottawa, Canada.
Ladizinsky, G., 1969. New evidence on the origin of the hexaploid oats. Evolution 23: 676–684.
Ladizinsky, G., 1971. Chromosome relationships between tetraploid (2n = 28)Avena murphyi and some diploid, tetraploid and hexaploid species. Can. J. Genet. Cytol. 13: 203–209.
Ladizinsky, G., 1993. The taxonomic status ofAvena magna: A reappraisal. Lagascalia (in press).
Ladizinsky, G. & D. Zohary, 1967.Avena ventricosa: Possible diploid contributor to hexaploid oats. Science 155: 1553–1554.
Ladizinsky, G. & D. Zohary, 1968. Genetic relationships between diploids and tetraploids in seriesEubarbata ofAvena. Can. J. Genet. Cytol. 10: 68–81.
Legget, J.M., G. Ladizinsky, P. Hagberg. & M. Obbanni, 1992. The distribution of nineAvena species in Spain and Morocco. Can. J. Bot. 70: 240–244.
Nishiyama, I., 1929. The genetics and cytology of certain cereals. 1. Morphological and cytological studies in triploid, pentaploid and hexaploidAvena hybrids. Jap. J. Genet. 5: 1–48.
Rajhathy, T., 1963. A standard karyotype forA. satina. Can. J. Genet. Cytol. 5: 127–132.
Rajhathy, T., 1966. Evidence and an hypothesis for the origin of the C genome of hexaploidAvena. Can. J. Genet. Cytol. 8: 774–779.
Rajhathy, T. & J.W. Morrison, 1959. Chromosome morphology in the genusAvena. Can. J. Bot. 37: 331–337.
Rajhathy, T. & R.S. Sadasivaiah, 1968. The chromosomes of Avena magna. Can. J. Genet. Cytol. 10: 385–389.
Rajhathy, T. & H. Thomas, 1974. Cytogenetics of Oats (Avena L.). pp 1–99, Misc. Publ. Genet. Soc. Ottawa, ON.
Thomas, H., 1992. Cytogenetics ofAvena./ In: H.G. Marshall & M.E. Sorrells (Eds), Oat Science and Technology. pp. 473–507. Sec. Agron. Madison, Wisconsin.
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Ladizinsky, G. Characterization of the missing diploid progenitors of the common oat. Genet Resour Crop Evol 42, 49–55 (1995). https://doi.org/10.1007/BF02310683
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1007/BF02310683