
Abstract
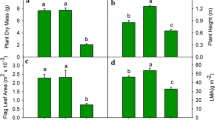
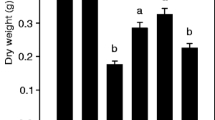
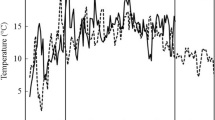
The plastochron model was used to evaluate the differences in the growth response of two Lycopersicon spp. grown under two temperature regimes (25/18 and 12/5°C). Two altitudinal accessions of L. hirsutum Homb. et Bnpl., from low and high altitude, a breeding line of L. esculentum (L.) Mill. and the hybrid between the latter and the high-altitude L. hirsutum were studied. The plastochron (P) values were estimated directly according to the formula of R.O. Erickson and F. Michelini (1957, Am. J. Bot. 44, 297–305), and indirectly through a linear model estimating the exponential rates of leaf elongation (r) and the ln of the plastochron ratios (q). The P values were obtained as P=q/r, and with one exception values obtained with both methods were comparable. Low temperature significantly decreased r in all genotypes, but the extent of this reduction depended on the genotype. The hybrid exhibited the least reduction, followed by the high-elevation L. hirsutum, L. esculentum and the lowelevation L. hirsutum. While the q values of the L. hirsutum accessions were significantly reduced by low temperature, those of L. esculentum and the hybrid were not. With the exception of the low-altitude L. hirsutum, low temperature significantly increased P, however the extent of the increase was significantly greater in L. esculentum. Analysis of temperature dependent changes of r, q and P indicate that L. esculentum extended its P by approximately the same factor its r was reduced. On the other hand, the L. hirsutum accessions increased P to a lesser extent, therefore having the ability to produce, comparatively, more leaves at lower temperatures than the cultivated tomato. The linear model of the plastochron is proposed as a tool for comparative studies of environmental growth responses of different genotypes. Plant size was reduced by low temperature. Considering plant size attained at high temperature as 100%, at low temperature sizes were reduced to 73% for the hybrid, 61% for the high-altitude L. hirsutum, 39% for L. esculentum and 30% for the low-altitude L. hirsutum. The low-temperature regime delayed flowering by two, three and nine plastochrons in the hybrid, the high-altitude L. hirsutum and L. esculentum, respectively, while the low-altitude L. hirsutum did not flower for the duration of the experiment. When artificially pollinated, L. esculentum yielded parthenocarpic fruits, while the high-altitude L. hirsutum and the hybrid produced fruits with viable seeds.
Similar content being viewed by others
Abbreviations
- a :
-
plastochron ratio
- CV:
-
coefficient of variability
- HTR:
-
high-temperature regime
- LPI:
-
leaf plastochron index
- LTR:
-
low-temperature regime
- P :
-
plastochron
- PI :
-
plastochron index
- q :
-
ln of the plastochron ratio
References
Coleman, W.K., Greyson, R.I. (1976) The growth and development of the leaf in tomato (Lycopersicon esculentum). I. The plastochron index, a suitable basis for description. Can. J. Bot. 54, 2421–2428
Erickson, R.O., Michelini, F.J. (1957) The plastochron index. Am. J. Bot. 44, 297–305
Foster, A.C., Tatman, E.C. (1937) Environmental conditions influencing the development of tomato pockets or puffs. Plant Physiol. 12, 875–880
Graham, D., Patterson, B.D. (1982) Responses of plants to low, non-freezing temperatures: proteins, metabolism, and acclimation. Annu. Rev. Plant Physiol. 33, 347–372
Gustafson, F.G. (1942) Parthenocarpy: natural and artificial. Bot. Rev. 8, 599–654
Horie, T., de Wit, C.T., Goudriaan, J., Bensink, J. (1979) A formal template for the development of cucumber in its vegetative state. I. Proc. K. Ned. Akad. Wet. Ser. C 82, 449–479
Lamoreaux, R.J., Chaney, W.R., Brown, K.M. (1978) The plastochron index: a review after two decades of use. Am. J. Bot. 65, 586–593
Lyons, J.M., Raison, J.K., Steponkus, P.L. (1979) The plant membrane in response to low temperature: an overview. In: Low temperature stress in crop plants. The role of the membrane, pp. 1–24, Lyons, J.M., Graham, D., Raison, J.K., eds. Academic Press, New York
Milthorpe, F.L. (1956) The relative importance of different stages of leaf growth in determining the resultant leaf area. In: The growth of leaves, pp. 141–150, Milthorpe, F.L., ed. Butterworths, London
Nitsch, J.P., Kurtz, E.B., Liverman, J.L., Went, F.W. (1952) The development of sex expression in cucurbit flowers. Am. J. Bot. 39, 32–43
Rick, C.M. (1973) Potential genetic resources in tomate species: clues from observations in native habitats. In: Genes, enzymes, and populations, pp. 255–269, Srb, A.M., ed. Plenum Press, New York
Rick, C.M. (1974) The tomato. In: Handbook of genetics, vol. 2, pp. 247–280, King, R.C., ed. Plenum Press, New York
Scheffé, H. (1970) The analysis of variance. Wiley, New York
Searle, S.K. (1971) Linear models. Wiley, New York
Vallejos, C.E., Tanksley, S.D. (1983) Segregation of isozyme markers and cold tolerance in an interspecific backcross of tomato. Theor. Appl. Genet. (in press)
Went, F.W. (1944) Plant growth under controlled conditions. II. Thermoperiodicity in growth and fruiting of the tomato. Am. J. Bot. 31, 135–150
Zamir, D., Tanksley, S.D., Jones, R.A. (1981) Low temperature effect on selective fertilization by pollen mixtures of wild and cultivated tomato species. Theor. Appl. Genet. 59, 235–238
Author information
Authors and Affiliations
Additional information
Taken in part from the doctoral dissertation submitted by C.E.V. in partial fulfillment of the Ph.D. requirements, University of California, Davis (1982)
Rights and permissions
About this article
Cite this article
Vallejos, C.E., Lyons, J.M., Breidenbach, R.W. et al. Characterization of a differential low-temperature growth response in two species of Lycopersicon: the plastochron as a tool. Planta 159, 487–496 (1983). https://doi.org/10.1007/BF00409137
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1007/BF00409137