
Abstract
Key message
Emerald ash borer-caused canopy gaps alter resource availability which affects woody plant community dynamics in eastern US forests. Our findings indicate that both canopy trees and seedlings are impacted by the disturbance. Seedlings of sugar maple and introduced woody plants benefited most from ash death, perhaps because they are adapted to ephemeral resource fluctuations.
Context
Ash, Fraxinus spp., mortality caused by emerald ash borer (EAB), Agrilus planipennis, results in canopy gaps which could increase resource availability and profoundly affect eastern US forests.
Aims
We tested two mechanisms: (1) EAB-caused ash decline releases growth of upper forest layers (non-ash canopy and subcanopy trees), constraining any release of lower forest strata; (2) EAB-caused ash decline increases canopy openness, releasing lower strata (shrub and seedling layer).
Methods
Sites representing a gradient of ash mortality were sampled throughout western Ohio to investigate how forest strata relative growth rates (RGRs) relate to EAB-caused ash mortality. Models including individual and additive effects of ash mortality and upper strata effects on woody seedlings were also investigated.
Results
Greater RGR of non-ash canopy trees, particularly maple (Acer spp.), was found in sites with more poor condition ash. Abundance of introduced seedlings was correlated with greater ash mortality and shrub cover. Sugar maple seedling height growth improved with ash loss and more subcanopy basal area (BA). Sites with greater subcanopy BA had greater introduced seedling recruitment and native seedling survival.
Conclusion
We found evidence to support both mechanisms. Our findings indicate species best adapted to ephemeral resource fluctuations, specifically sugar maple and introduced seedlings, benefited most from ash death.
Similar content being viewed by others

1 Introduction
Heterogeneity resulting from canopy tree mortality is important for forest regeneration (Sapkota et al. 2009). Canopy gaps cause dramatic shifts in light, temperature, soil moisture, and nutrient availability (Muscolo et al. 2014). As a consequence, gap dynamics may increase structural complexity, habitat diversity, and the species diversity of fauna and flora (Muscolo et al. 2014). However, gaps formed by tree mortality due to the activities of invasive species are predicted to have unique impacts on forests compared to natural mortality and abiotic disturbances (Ellison et al. 2005; Lovett et al. 2006; Lovett et al. 2013; Gandhi and Herms 2010; Lovett et al. 2016) owing to protracted host tree decline that results in widespread, selective mortality of specific tree taxa (Eschtruth et al. 2006; Runkle 2007). Often these organisms are insect herbivores, which cause a multitude of direct and indirect impacts to North American forests (Haack 2006; Flower et al. 2013a; Gandhi et al. 2014; Flower et al. 2014; Costilow et al. 2017; Perry and Herms 2017; Savage and Rieske 2018). Widespread defoliation and gap formation by invasive insects have been shown to lead to changes in forest structure, canopy composition, and ecosystem processes (Kenis et al. 2009; Gandhi and Herms 2010; Flower et al. 2013a). Since invasive herbivores often target a specific genus or small collection of species, the severity of the disturbance may depend on the mortality level, distribution, dominance, and density of the host trees (Lovett et al. 2006; Gandhi and Herms 2010). These events can result in increases in understory light (Eschtruth et al. 2006), release of other tree species (Ehrenfeld 1980; Muzika and Liebhold 1999; Jedlicka et al. 2004; Flower et al. 2013a: Costilow et al. 2017), increase in species diversity (Eschtruth et al. 2006), and release of invasive plants (Eschtruth et al. 2006; Hoven et al. 2017).
One insect herbivore that has recently invaded North America and has the potential to shape forest plant community composition and structure is emerald ash borer (Agrilus planipennis) (EAB). This wood-boring beetle native to eastern Asia already has and continues to kill tens of millions of ash (Fraxinus spp.) trees as it spreads across North America (Kovacs et al. 2010; Liebhold et al. 2013). Based upon both current and expected damage, EAB is considered the most economically costly invasive forest insect ever introduced to North America (Herms and McCullough 2014). All 16 native U.S. species of ash are viable hosts (Herms et al. 2004) for the phloem-boring, larval stage of EAB (Flower et al. 2013b). Recent studies of ash loss caused by EAB have revealed declines in forest net primary productivity and carbon sequestration (Flower et al. 2013a), increases in coarse woody debris (Higham et al. 2017; Perry et al. 2018), decreases in both species diversity and richness of forest floor invertebrate communities (Perry and Herms 2016), as well as impacts on plant communities.
In addition to the negative ecological impacts noted above, the loss of ash from North American forests is contributing to significant changes to forest vegetation. EAB-caused ash loss has been shown to alter seedling community composition by depleting the ash seedbank (Klooster et al. 2014); decreasing ash seed production (Kashian 2016); decreasing green ash (F. pennsylvanica), black ash (F. nigra), and white ash (F. americana) seedling abundance (Klooster et al. 2014; Spei and Kashian 2017); and increasing seedlings of an invasive shrub (Hoven et al. 2017). In addition to the seedling layer, the effects of EAB-caused ash mortality on the sapling layer (Klooster et al. 2014; Margulies et al. 2017; Dolan and Kilgore 2018), invasive plants (Klooster 2012; Hoven et al. 2017; Margulies et al. 2017; Dolan and Kilgore 2018), and canopy tree layer (Flower et al. 2013a; Costilow et al. 2017; Spei and Kashian 2017; Dolan and Kilgore 2018) have also been explored.
While the responses of individual forest strata to ash mortality have been relatively well studied, simultaneously evaluating growth across multiple vegetation layers enables assessment of how upper levels interact with the resources made available by ash mortality to influence the response of lower layers. Consideration of the full set of these responses is expected to provide better insights into the longer-term trajectory of forest stand community composition. Finally, observations along a gradient of EAB-caused ash mortality can supplement experiments of simulated ash loss.
We conducted a 3-year, multi-strata study on the effects of ash loss across a gradient of ash mortality. Forest strata responses evaluated included growth of all woody vegetation layers (i.e., non-ash canopy, subcanopy, native and introduced shrubs, and woody seedlings) and richness, abundance, survival, and recruitment of seedlings, incorporating subsequent effects of higher vegetation layers on lower layers (Fig. 1). We tested the following two mechanisms for how woody plant communities respond to EAB-caused ash mortality (Fig. 1): (1) EAB-caused ash decline releases the growth of upper forest layers (non-ash canopy trees and subcanopy trees), constraining any release of lower forest strata; (2) EAB-caused ash decline increases canopy openness which in turn releases lower vegetation layers. Based upon the first mechanism, we predict that (i) ash decline is positively correlated with non-ash canopy and/or subcanopy growth and that (ii) the additive effect of ash decline and upper forest strata (which is represented by the basal area (BA) for all layers occurring above the layer of interest) is correlated with negative effects on shrub and seedling layers. In contrast, mechanism 2 predicts that (iii) canopy openness is positively correlated with ash decline, (iv) shrub growth is positively correlated with ash decline, and (v) seedling parameters are positively correlated with ash decline. We distinguished responses of native vs. introduced shrubs, as invasive plants often benefit from disturbance (Hobbs and Huenneke 1992), Gandhi and Herms (2010) predicted EAB would facilitate plant invasion, and Hoven et al. (2017) found that growth of an invasive shrub was associated with ash decline. To predict impacts on future forest composition, we distinguished responses of three groups of woody seedlings: introduced, native shade intolerant, and native shade tolerant.

Flowcharts describing our two mechanisms. Direct effects of EAB-caused ash decline (solid arrows) and indirect effects (dashed arrows). Arrows with “+”represent a positive effect on the strata and with “−” indicate a negative effect. Relative growth rates (RGRs) represent forest layer growth (2012–2014) and predictor variables represent size (2012). M1 mechanism 1, M2 mechanism 2, pi-pv predictions, BA basal area, RGR relative growth rate
2 Materials and methods
2.1 Study area
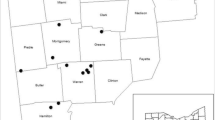
We conducted our study in 24 sites (Fig. 2) within three geographic regions of Ohio (Northwest, Central, and Southwest), selected from sites established by the US Forest Service to study impacts of EAB (Knight et al. 2013). Sites located in Northwest Ohio experience annual average temperatures of 11.9 °C and annual precipitation levels of 85.2 cm (www.usclimatedata.com), soils are loamy and clayey, level to gently sloping, very to somewhat poorly drained (websoilsurvey.nrcs.usda.gov). Annual average temperatures are 11.6 °C and annual precipitation levels are 142.5 cm for sites located in Central Ohio (www.usclimatedata.com), soils are silt loam, level to complex slopes, moderate to poorly drained (websoilsurvey.nrcs.usda.gov). Annual temperatures in Southwest Ohio are 11.6 °C with annual precipitation levels at 105.6 cm (www.usclimatedata.com), soils a silt loam, mainly level to moderately steep, moderate to well drained (websoilsurvey.nrcs.usda.gov). These sites represent a range of impacts from EAB infestation, based on first appearance of D-shaped exit holes ranging from 2006 to 2015 (Appendix 1). Ohio provided an ideal location to explore the effects of EAB-caused ash death because five species of ash are native to this state (Hausman et al. 2010), where they number approximately 279 million individuals, constituting approximately 6% of all trees in the state (Wildman 2008). Furthermore, the EAB infestation in Ohio represents a north to south gradient since colonization. The Northwest, where EAB was initially discovered in 2003, represents the longest time period since infestation (www.aphis.usda.gov 2018). Central Ohio represents an intermediate time period since EAB infestation and the most recent invasions were detected in the Southwest (Appendix 1). Sites were public or private, mostly secondary forests with minimal active management located within a matrix of suburban and agricultural lands.

Map of 24 study sites in Ohio, USA. Each dot denotes a site, nested within each site were three 400 m2 plots. Sites and plots were established by the US Forest Service for monitoring long-term EAB ecological impacts
Across the gradient, sites differed slightly in topography and presence of seasonal inundation. The Northwest Region contained two sites considered as floodplain, and four that were lowland. In the Central Region, five sites were classified as upland and one was classified as lowland. In the Southwest Region, there were ten sites classified as upland, and two sites that were lowland (Appendix 1). Despite topographical differences, there was no significant difference among the 24 sites in the plot-level proportion of ash in the canopy (one-way ANOVA, F23,48 = 1.04, p = 0.44; mean 40%; range 21–65%). However, sites did differ in which ash species dominated the plots (Appendix 1).
Although the year of initial infestation by EAB is unknown due to the difficulty of early detection, the presence of EAB was confirmed through yearly documentation of characteristic D-shaped exit holes on dying ash trees (Appendix 1) in all except two sites (CCSP4 and CLF—plots with no exit holes are distinguished with an NA) and yearly trapping of EAB adult beetles in a subset of sites. EAB was likely present at a site at low densities before these methods confirmed its presence. Using these methods, the gradient of known duration of EAB presence in the other 22 sites was 1 to 9 years at the time of final plant measurements (2014). Tree diameter, ash health, seedling measurements, canopy openness, as well as native and introduced shrub basal diameter and percent cover, were recorded annually from June through August, 2012–2014 (Appendices 0–0).
2.2 Study design
Three circular plots of 400 m2 were nested within each of the 24 sites (Hoven et al. 2017). All plots were located away from forest edges or trails, and spaced > 50 m apart. All plots included at least two ash trees > 10 cm diameter at breast height (DBH). For each plot, we determined the “prevalent” ash species as the one or two species that comprised ≥ 33% of the ash BA (see Appendix 1). Two species of ash, F. pennsylvanica and F. profunda, were pooled in all analyses because they were not consistently distinguished. Canopy openness was collected with a spherical concave densiometer, taking four measurements (open sky) at 6 m from the center of the plot in the four cardinal directions at a height of approximately 1.3 m, following the methods of Strickler (1959). The 16 measurements were averaged, and then multiplied by 1.04 to determine percent canopy openness (Lemmon 1956).
In each 400 m2 plot, we identified and annually measured DBH for all trees ≥ 10 cm DBH and assessed the health of each ash canopy tree on a scale of 1–5, where 1 is a healthy tree, 5 is a dead tree, and 2–4 are progressive stages of decline (Knight et al. 2013). All ash mortality observed in our sites was due to EAB attack rather than from “Ash Decline,” which is suggested to be caused by ash yellows, environmental conditions (e.g., spring drought) (Palik et al. 2011), or viral infection (Sinclair and Griffiths 1994) (Charles E. Flower, personal communication). BA for all canopy (≥ 10 cm DBH) trees, including dead and dying ash, was calculated using DBH, and is hereafter referred to as “total stand BA.” Basal area for all non-ash canopy trees was also calculated for each plot and is hereafter referred to as “non-ash stand BA.” Per-year relative growth rate (RGR) of non-ash stand BA, as well as all other RGR calculations were produced using measurements from 2012 (t = 0) and 2014 (t = 2) (Appendices 0–0) and calculated following (Feeley et al. 2007) as:
Because maple (Acer spp.) consistently constituted a sizeable portion of the non-ash canopy and subcanopy in nearly all plots and has been shown to respond EAB-caused ash decline (Flower et al. 2013a; Costilow et al. 2017), we also evaluated maple canopy and subcanopy tree RGR separately (Appendices 0–0).
To quantify EAB-caused ash decline, we calculated three metrics for each plot (Appendix 1). The first, ash decline index (ADI), was calculated as the total BA of ash trees that received a rating between 3 and 5 divided by the total stand BA. The second, ash mortality index (AMI), was the BA of dead ash (rated 5) divided by the total stand BA. These two indices were calculated to differentiate between the effects of thinning ash canopies (ADI) and the effects of ash death (AMI) on understory resource availability. Basal area measurements and ash condition ratings from 2012 were used in calculating both variables (Hoven et al. 2017). The third metric of ash decline was the first year that a D-shaped EAB exit hole was detected (1st Exit). Both ADI and AMI were negatively correlated with 1st Exit, where sites with earlier exit hole appearances had higher ADI and AMI values (ADI: Kendall’s rank correlation z = − 4.211, p < 0.001; AMI: z = − 5.321, p < 0.001).
Located at the center of each plot was a circular 200-m2 subplot where we measured subcanopy DBH as well as shrub basal diameter for each live stem which were used to calculate BA and shrub percent cover. In each subplot, we tagged all subcanopy trees (3.0–9.9 cm DBH) and measured DBH annually. To quantify growth in the subcanopy layer, we calculated BA of each tree based on its DBH and summed these to obtain subcanopy BA for each plot for 2012 and 2014 (Appendices 0–0). To quantify growth in the shrub layer, we tagged the two largest shrubs (criteria: ≥ 1 m tall and the largest basal diameter) of each species in each of the four quadrants of each subplot, for a total of up to eight individuals per species in each subplot (Appendix 2–3). Annually, we measured the basal diameter of each stem for each tagged shrub. Using these basal diameters, we calculated BA for each stem and then summed these for each individual shrub, following the methods of Elliott and Swank (1994).
Shrub species were classified into two categories, native or introduced (www.plants.usda.gov 2017; Appendix 5). The BAs for all shrubs in each category were summed for each subplot for 2012 and 2014 and used to calculate shrub RGR (Appendices 0–0).
Shrub percent cover was quantified using the line-point intercept method (Gordínez-Alvarez et al. 2009). Seven parallel transects were checked annually in each subplot. Transects were 2 m apart and points were sampled every 2 m. Five center transects (12 m each) had seven sample points each and the two transects (8 m each) which flanked the center transects had five sample points each. Transects sampled the entire extent of the subplot. At each sample point (N = 45/subplot), the presence or absence of a shrub was recorded. The number of points where a shrub was present was divided by 45 to determine shrub layer percent cover (Appendix 2).
Woody seedlings were sampled in each plot within four permanent circular 4 m2 micro-plots, located 6 m from plot center, one in each of the four cardinal directions. In 2012, we identified, measured the height, and tagged all tree and shrub seedlings (no vines) that were 20–100 cm tall in each of the micro-plots; these counts were pooled by plot to serve as the seedling abundance values in our analyses (Appendix 4). In 2013 and 2014, all tagged seedlings were re-measured. Seedlings which entered the 20–100 cm threshold in 2013 or 2014 were identified, tagged, and measured; these were pooled by plot and serve as the seedling recruits in the analyses (Appendix 4). Seedling species richness was calculated by pooling all species present within a plot (2012–2014) (Appendix 4). Survival was calculated as the proportion of 2012 seedlings in a plot that were still alive in 2014 (Appendix 4). Seedlings were divided into three categories: introduced, shade-intolerant natives, and shade-tolerant natives. We used shade tolerance to categorize the 46 native species (some of which were rare) because light limitation is often cited as the most limiting resource in closed canopy forests (Pacala et al. 1994). Following Hoven et al. (2017), we classified as shade intolerant those species with shade tolerance values of 1–2.99 in Niinemets and Valladares (2006), while those with values of 3–5 were classified as shade tolerant (Appendix 6).
We used sugar maple seedlings as a phytometer (Twolan-Strutt and Keddy 1996) to investigate seedling growth responses to EAB and avoid potential issues of different growth rates among multiple species, due to the importance of sugar maple response to ash mortality, as well as the widespread abundance of its seedlings in our plots. Heights for the 425 sugar maple seedlings that survived from 2012 to 2014 were used to calculate RGR for each seedling, then for each plot we determined the mean sugar maple seedling height RGR (Appendices 0–0).
2.3 Statistical analysis
We conducted statistical analyses using generalized linear mixed and linear mixed effects models with maximum likelihood estimation for fixed effects (Bolker et al. 2009) in the R packages lme4 and nlme in R version 3.0.2 (Bates et al. 2017) of the R programming language (R Development Core Team 2017). Linear mixed models were used when the response variable could be transformed to achieve normality of the residuals and generalized linear mixed models were used when the response variable followed a Poisson or binomial error distribution. Goodness of fit tests were used to determine appropriate model structure. Because analyses used observations from three replicate plots at each site, all models used “site” as a random variable (~1|Site), allowing us to account for the variation across plots nested within sites.
A candidate set of four models was used to understand the relationships between ash mortality and the following three response variables: canopy openness, non-ash stand BA RGR, and maple stand BA RGR. Each candidate model included a single predictor from among the following three variables: ADI, AMI, and 1st Exit. The fourth model in the candidate set was the null model and included only the random effect of site.
For all other response variables, there were multiple predictor variables and multiple combinations of predictors that we were interested in testing. To identify the most parsimonious models for each response, we conducted model selection of a candidate set of models. Response variables representing lower strata of the forest included: subcanopy BA RGR, maple subcanopy BA RGR, native shrub BA RGR, introduced shrub BA RGR, and sugar maple seedling height RGR. All RGR response variables were log transformed to approximate a normal error distribution. We also evaluated several seedling parameters: abundance, recruitment, and survival for each of our three seedling subsets (introduced, native shade-tolerant, and native shade-intolerant) as well as overall species richness. Seedling species richness was modeled with a Poisson error distribution and seedling survival was modeled with a binomial error distribution. All other response variables were log transformed and modeled as a normal distribution. Predictor variables for the candidate set of models included: ADI, AMI, subcanopy BA, shrub layer percent cover, and non-ash canopy BA to account for canopy shading differences among plots. Each predictor variable was calculated from measurements made in 2012. We also evaluated two-factor additive models composed of one ash decline variable (ADI or AMI) and a vegetation layer (non-ash canopy BA, subcanopy BA, or shrub layer) that occurred above the response layer of interest. We did not include the predictor 1st Exit in the candidate set, as this variable performed poorly compared to ADI and AMI during the initial hypothesis tests. Mean ADI and AMI were consistently lower for upland sites, 0.20 ± 0.03 and 0.13 ± 0.02, respectively, as compared to wet sites (floodplain and lowland combined), 0.32 ± 0.04 and 0.29 ± 0.05. Due to this, we decided to consider an additional predictor variable, topography (plots categorized as either upland or wet) when analyzing each response variable for which the best or competing models included ADI or AMI: introduced non-ash canopy RGR, maple canopy RGR, introduced seedling abundance, and sugar maple seedling height RGR.
The most parsimonious models included predictor variables that provided the lowest value of the Akaike information criteria corrected for small sample size (AICc), based on model comparisons using the R package AICcmodavg (Mazerolle 2015). Models with ΔAICc < 2 were considered competing models (Burnham and Anderson 2002). The relative strength of evidence for alternative models (most parsimonious vs competing models) was assessed using Akaike weights (reported as w). Employing AICc allowed us to evaluate and rank models to assess which most closely approximated reality based on the data that was collected during the study (Burnham and Anderson 2002). Most parsimonious and competing models are reported in Appendices 0–0, and only the most parsimonious models are reported in Table 1. When the null model (no fixed effects, site included a random effect) was included in the set of competing models (∆AICc < 2), the effects of predictor variables also included in the set were not considered further.
3 Results
In 2012, mean total stand basal area (BA) per plot was 39.1 ± 1.8 (SE) m2 ha−1, mean non-ash stand BA per plot was 23.0 ± 1.2 m2 ha−1, and mean maple stand BA per plot was 8.7 ± 1.0 m2 ha−1; maple constituted 37 ± 3% of the total stand BA in 2012 (Appendix 2). Our 24 sites represented a range of dates when the first exit hole was recorded (2006–2015) (Appendix 1). In 2012, mean ADI was 25 ± 0.02% and mean AMI was 19 ± 0.02% (Appendix 1). Subcanopy layer (trees 3.0–9.9 cm DBH) BA per plot averaged 1.8 ± 0.2 m2 ha−1 and maple subcanopy BA per plot was 0.4 ± 0.05 m2 ha−1; maple constituted 23 ± 0.02% of the total subcanopy layer in 2012 (Appendix 2). Per plot native shrub BA was 0.23 ± 0.04 m2 ha−1 and introduced shrub BA was 0.32 ± 0.07 m2 ha−1. Mean shrub layer (native and introduced combined) percent cover in 2012 was 34 ± 4% (Appendix 2).
3.1 Upper-forest strata relative growth rates
Non-ash stand BA RGR per plot from 2012 to 2014 was 0.04 ± 0.003 m2 m−2 year−1 (Appendices 0–0). We found that both ADI and AMI were competing models for predicting non-ash stand RGR, with the former comprising the most parsimonious model form our candidate set (Table 1, Appendix 7). There was greater radial growth of non-ash canopy trees in sites where more poor condition ash were present. When topography was included as a predictor variable there were four competing models, with the additive model of ADI and topography being the most parsimonious model (Fig. 3, Table 1, Appendix 7). Upland sites experienced less non-ash canopy radial growth than wet sites, and the positive association between growth and ash decline was only manifest in upland sites. Mean maple stand BA RGR per plot was 0.05 ± 0.005 m2 m−2 year−1. The most parsimonious model for predicting maple growth was AMI and a competing model was ADI (Fig. 4, Table 1, Appendix 7); this did not change when topography was added as a predictor variable (Table 1, Appendix 7). There was greater radial growth of maple canopy trees in sites where poor condition ash comprised more of the basal area. Mean tree BA RGR per plot for the subcanopy layer and maple subcanopy was 0.07 ± 0.01 m2 m−2 year−1and 0.4 ± 0.05 m2 m−2 year−1, respectively (Appendix 8). The null model (which included only site as a predictor) was the most parsimonious for both subcanopy and maple subcanopy radial growth (Appendix 8), indicating that the other predictors were not important.

Regression of non-ash canopy basal area (BA) annual relative growth rate (RGR) (2012–2014) on ash decline index (ADI) (2012). Study sites (the mean of 3 plots per site) are divided into two topographical categories, upland (solid line) and wet (dotted line). Lines illustrate the best fit to these site means, and the gray shaded regions represent the standard error around that line. However, the statistics reported are based on hypothesis testing of plot data using linear mixed models (χ2 = 4.76, p = 0.03) with “site” as the random factor

Regression of maple canopy basal area (BA) annual relative growth rate (RGR) (2012–2014) on ash mortality index (AMI) (2012). Each point represents one study site (the mean of 3 plots per site). The solid black line illustrates the line of the best fit to these site means, and the gray shaded region represents the standard error around that line. However, the statistics reported are based on hypothesis testing of plot data using linear mixed models (χ2 = 5.60, p = 0.01) with “site” as the random factor
3.2 Lower forest strata responses
We found that mean canopy openness per plot, as measured at 1.3 m, was 7.4 ± 0.6% SE in 2012. None of our ash decline indices were related to greater canopy openness (Appendix 7). Plots with poorer condition ash, or earlier attack by EAB, did not have greater canopy openness. Mean shrub BA RGR per plot from 2012 to 2014 was 0.1 ± 0.03 m2 m−2 year−1 for native shrubs and 0.13 ± 0.02 m2 m−2 year−1 for introduced shrubs (Appendices 0–0). The null model was the most parsimonious and only model for native shrub RGR; while there were three competing models for introduced shrub RGR, the null model was still the most parsimonious (Appendix 8). Mean sugar maple seedling height RGR per plot was 0.04 ± 0.01 m m−1 year−1 (Appendices 0–0). The most parsimonious model for A. saccharum seedling height RGR was the additive model of ADI and subcanopy BA (Table 1, Appendix 9). Sugar maple seedlings grew more in sites with poorer ash condition as well as greater subcanopy BA. One plot with very high mean sugar maple height growth appeared to be an outlier. When the plot was dropped from the analysis, the most parsimonious model remained the same. When the predictor variable topography was considered, all but one of these seedlings were on upland plots, so we did not reanalyze with topography as a predictor.
There were a total of 57 species identified in the woody seedling layer: 11 introduced, 25 shade-tolerant natives, and 21 shade-intolerant natives (Appendix 5). The average number of seedling species per plot was 7.4 ± 0.5 SE (all seedling counts are based on the total sampled area of the micro-plots, i.e., 16 m2 per plot). The null model was the most parsimonious predictor for overall seedling species richness (Appendices 0 and 0).
Introduced seedling abundance averaged 3020.8 ± 484.2 per ha in 2012, and the most parsimonious model included the effects of ADI and shrub layer percent cover in 2012 (Table 1, Appendices 0 and 0). There were two additional competing additive models for introduced seedling abundance; one was the additive effect of AMI and shrub layer percent cover and the other was solely shrub layer percent cover (Table 1, Appendices 0 and 0). More introduced seedlings were present in sites with greater shrub cover and with poorer ash condition. One plot with very high introduced seedling abundance appeared to be an outlier. When the plot was dropped from the analysis, the three competing models remained the same. When topography was included as a predictor variable, the most parsimonious model again included the additive effects of ADI and shrub cover as well as topography. The additive models of shrub cover and topography as well as the additive models including AMI, shrub cover, and topography were competing models (Appendices 0 and 0). Introduced seedlings were more abundant in upland than in wet sites, and the positive response of introduced seedling abundance to poorer ash condition as well as to shrub cover was only manifest in upland sites (Fig. 5a, b). Mean seedling abundances for shade-tolerant and shade-intolerant native seedlings were 9435.8 ± 1345.5 and 4088.5 ± 715.3 per ha, respectively. The null model was the most parsimonious model for shade-tolerant native seedling abundance and a competing model for shade-intolerant native seedling abundance (Appendices 0 and 0).

a, b Regressions of introduced seedling abundance (2012) on ash decline index (ADI) (2012) (a) and on shrub cover (2012) (b). Study sites (the mean of 3 plots per site) are divided into two topographical categories, upland (solid line) and wet (dotted line). Lines illustrate the best fit to these site means, and the gray shaded regions represent the standard error around that line
Introduced seedling recruitment averaged 1475.7 ± 212.8 seedlings per ha and was best approximated by only the effect of 2012 subcanopy BA (Table 1, Appendices 0 and 0). More seedlings of introduced species recruited in sites with greater subcanopy BA. Mean seedling recruitment for shade-tolerant and shade-intolerant native seedlings were 5590.3 ± 968.4 and 3125.0 ± 376.6 per ha, respectively. For both native seedling categories, the null model was the best predictor of seedling recruitment (Appendices 0 and 0).
Among plots, mean survival for seedlings from 2012 to 2014 was 80 ± 3%. Mean seedling survival was 76 ± 3% for shade-tolerant native seedlings, 91 ± 3% for introduced seedlings, and 77 ± 4% for shade-intolerant native seedlings. The most parsimonious model for shade-tolerant seedling survival included only the effect of 2012 subcanopy BA (Table 1, Appendices 0 and 0); survival was higher in plots with greater subcanopy BA. The most parsimonious model was not substantially better than the null model for introduced and intolerant native seedling survival (Appendices 0 and 0).
4 Discussion
Our results demonstrate the complexity of forest responses to diffuse disturbance from an invasive pest. Specifically, as summarized in Fig. 6, they indicate that EAB-caused ash decline enhanced non-ash canopy tree growth in upland sites which, combined with high shrub cover, was associated with greater abundance of introduced seedlings. Sugar maple seedling height growth in upland sites was greater in sites with greater ash decline and greater subcanopy BA. Finally, both recruitment of introduced seedlings and survival of shade-tolerant native seedlings were greater in sites with greater subcanopy BA. Therefore, in the short term, it seems that ash decline results in competitive release for other canopy trees and sugar maple seedlings, at least in upland sites. However, it may have a more insidious long-term effect by increasing the abundance of introduced seedlings. Furthermore, effects of ash decline on the seedling community are shaped by the initial density of other forest strata, particularly the subcanopy and shrub layers.

Flowchart illustrating effects of EAB-caused ash decline (solid arrows) and forest layers (dashed arrows) that were supported by this study. Relationships (arrows) shown in Fig. 1 that are not included here are those found not to be supported. Arrows with a “+” signify a positive effect. Black boxes represent predictor and response variables that are correlated with ash decline and/or other vegetation strata. Light gray boxes and text represent variables that were not included in the best model or competing model, and thus do not improve explanatory power. NSI native shade-intolerant seedlings, NST native shade-tolerant seedlings, INT introduced seedlings, U in upland plots only
4.1 Mechanism 1
Our findings were partially consistent with mechanism 1, whereby EAB-caused ash decline releases the growth of upper forest layers (non-ash canopy trees and subcanopy trees), constraining any release of lower forest strata. Specifically, prediction (i), was realized in part – growth of non-ash canopy trees, as well as just maples, were positively associated with EAB-caused ash mortality. For non-ash canopy trees, the effect of ash mortality on radial growth was apparent in upland sites compared to no growth response in wet sites. These results suggest increases in availability of belowground resources, specifically water, rather than light, may be driving canopy tree responses. Water availability potentially limits growth in the relatively dry upland sites, whereas in wetter sites, an influx of water would not be expected to affect growth. The canopy layer is both the largest by BA and also occupies the highest layer (overshadows all other strata); thus, resources should be attenuated by this layer first, likely causing the positive growth response. This pattern of non-host canopy trees experiencing a growth release has also been reported following outbreaks of gypsy moth, Lymantria dispar (Ehrenfeld 1980; Muzika and Liebhold 1999; Jedlicka et al. 2004).
We also found that the most parsimonious model for the RGR of canopy maples (Acer rubrum, A. saccharinum, A. nigrum, A. negundo, and A. saccharum) was a model that included only the effect of AMI. This finding was consistent with the positive radial growth responses in A. saccharinum and A. saccharum following EAB-caused ash mortality found by Flower et al. (2013a) as well as in A. rubrum and A. saccharinum shown by Costilow et al. (2017). Our data supports the premise that this pattern is maintained across the genus Acer, as well as reveals this pattern to hold across sites experiencing different stages of EAB-caused ash decline, and among sites which differ in topography and the presence of seasonal inundation. Interestingly, we did not find an effect of topography on maple canopy growth, indicating that perhaps an increase in belowground resources like water are not as responsible for increases in maple growth. This may indicate that other non-ash species are either more sensitive to water availability or maple is responding to other resource influxes.
Neither total subcanopy nor the subset of maples in the subcanopy responded to EAB-caused ash decline. Again, only 2 years of growth data and using only DBH measurements could also be responsible for why a relationship was not detected. Perhaps a measurement of height or leaf area index would have returned a positive relationship. Previous studies have found positive impacts of EAB on the subcanopy layer. Davis et al. (2016) found that non-ash sapling BA RGRs following girdling of canopy black ash (F. nigra) were significantly higher than those in control sites. However, the act of girdling ash trees may lead to quicker and simultaneous ash mortality and thus a greater spike in resources compared to EAB-caused ash mortality. Additionally, the larger percentage of the canopy consisting of ash BA (≥ 66%) prior to treatment (Davis et al. 2016), compared to our sites where ash averaged 40% of the canopy BA, could also result in greater resource availability for the subcanopy following girdling.
Instances of subcanopy responses following other invasive forest pests have produced mixed results. Ehrenfeld (1980) found a positive radial growth response of flowering dogwood, Cornus florida, following a gypsy moth defoliation event. Following the loss of American elm, Ulmus americana, to Dutch Elm Disease, Ophiostoma spp. (DED); a long-term investigation (Huenneke 1983) found no difference in subcanopy tree abundance between plots with single and multiple elm tree deaths. Differences in host breadth may explain these differences in responses. Unlike EAB and DED, gypsy moth is highly polyphagous (Wagner 2010) and thus may have a more profound impact on canopy openness by defoliating a greater percentage of canopy trees. Therefore, slower canopy die-back, as in the cases of EAB-caused ash decline and DED, may be insufficient to enhance growth in the subcanopy.
Based upon prediction (ii), we anticipated that that greater BA of upper vegetation layers would be associated with decreased growth of lower strata vegetation as well as suppressed seedling responses. Our prediction however was not supported; we actually found evidence of the opposite relationship. In the presence of greater subcanopy BA or shrub cover, sugar maple seedling height growth, shade-tolerant native seedling survival, and introduced seedling abundance and recruitment were all greater. Sugar maple seedling growth was greater in sites with poorer ash condition and greater subcanopy BA. Sugar maple seedling growth also did not show a consistent or statistically significant relationship with measured differences in white-tailed deer (Odocoileus virginianus) browse intensity (as measured by the sugar maple browse index (Rooney and Waller 2003) among the sites (Pearson’s product-moment correlation 0.23; 95% CI − 0.18–0.57). Furthermore, only 119 of the 424 sugar maple seedlings that survived had final heights shorter than initial heights, which may have been due to non-lethal browse. While greater growth of sugar maple in sites with poor ash condition will be discussed later, the association of sugar maple growth and shade-tolerant native seedling survival with greater subcanopy BA may be related to the adaptation of sugar maple to dense shade. A large proportion of our shade-tolerant native seedlings were sugar maple. Marks and Gardescu (1998) found over a 24-year period that shaded sugar maple seedlings experience high rates of survival while putting on very little height growth. Beaudet and Messier (1998) suggest that shade-tolerant species like sugar maple and American beech (Fagus grandifolia) have a tendency to allocate a larger proportion of their biomass to structures that promote long-term survival, like root growth, compared to less shade-tolerant competitors.
Introduced seedling recruitment was positively correlated with greater subcanopy BA, which could be explained by the “perch tree effect,” greater seed deposition under perch and roost structures (Holl 1998). This explanation is conceivable because all introduced seedling species recorded (Appendix 5) produce fleshy bird-dispersed fruits. Unfortunately, to our knowledge, this phenomenon has only been examined in open, early successional and patchy tree/shrub/open landscapes (Jordano and Schupp 2000). However, it seems plausible that abundant, small diameter trees may have a similar effect in forest interiors. Introduced seedling abundance was greater in sites that had poorer ash condition and more shrub cover. In a subset of these sites, we found Amur honeysuckle, Lonicera maackii, seedling abundance correlated with EAB-caused ash decline and greater L. maackii cover (Hoven et al. 2017). The relationship between ash mortality and introduced seedling abundance will be explored in the following section. The correlation between greater introduced seedling abundance and shrub cover could result from greater seedling establishment under parent shrubs, especially since most shrub cover was comprised of introduced shrubs.
4.2 Mechanism 2
We found mixed support for mechanism 2, where EAB-caused ash decline increases canopy openness which in turn releases lower vegetation layers including woody seedlings and native and introduced shrubs. For prediction (iii), we found no correlation between canopy openness (measured at 1.3 m) and ash decline. Similarly, Dolan and Kilgore (2018) found no relationship between canopy cover measured in the understory after EAB-caused ash mortality and relative ash BA before EAB. We think infilling by non-ash canopy species accounts for why ash death did not enhance canopy openness, thus preventing any increase in canopy openness we were capable of detecting with the methods used. This is supported by the relationship we found between improved non-ash canopy tree and specifically maple canopy tree RGR and plots with higher ADI and AMI, respectively. Infilling by upper strata vegetation has been previously documented by Beckage et al. (2000), who found that an intact shrub canopy prevented increases in understory light availability following gap formation from canopy tree girdling.
Mechanism 2 prediction (iv) which proposed that the growth of native and introduced shrubs are positively associated with ash decline was not realized. This was a surprising result since previous investigations have found correlations between native shrub responses and declines of American elm due to DED (Huenneke 1983; Dunn 1986) and of oak, Quercus spp., due to repeated gypsy moth defoliation events (Hicks and Hustin 1989). Dolan and Kilgore (2018) found shade-tolerant, but not shade-intolerant, shrubs and saplings showed greater increase in density in sites where EAB-impacted ash comprised a larger fraction of the canopy. Furthermore, it was unexpected that introduced shrubs in particular did not respond since Burnham and Lee (2010) found that the introduced glossy buckthorn, Frangula alnus, increased in stem density in canopy gaps. Luken et al. (1997) found that under high light environments, the non-native L. maackii was able to respond with higher growth rates than the native spicebush Lindera benzoin. In a subset of our sites where L. maackii was prevalent, we found its BA growth was positively correlated with ash decline (Hoven et al. 2017). One potential reason for this difference is that these previous studies focused on a single introduced species whereas in the present study, we grouped all nine introduced shrub species (Appendix 5), many of which were limited to a few sites. Dolan and Kilgore (2018) also grouped non-native species in the shrub and sapling layer, and found that their increase in density was not dependent on the basal area of ash in EAB-impacted stands Differences in growth habit and growth rate of some shrub species may have masked the positive response of others.
Some of our findings were consistent with mechanism 2, prediction (v), some seedling parameters were positively correlated with ash decline. Sugar maple seedling height growth was greater in sites with poorer ash condition. Sugar maple seedlings are likely responding to greater resource availability in sites with poor ash condition. Muscolo et al. (2014) reported that the physiology and morphological plasticity of shade-tolerant species like sugar maple, shade tolerance (4.76) rating (Appendix 6), enables them to respond quickly to rapid increases in light. Similarly, mean height RGR of tree seedlings (10 species pooled) planted to replace F. nigra was greater in clear-cut and girdling treatments (to simulate EAB-caused ash mortality) compared to partial overstory removal and control plots (Looney et al. 2017b). Several other studies have shown improved growth of woody seedlings in canopy gaps (Orwig and Foster 1998; Battles and Fahey 2000; Diaci et al. 2008; Caquet et al. 2010).
An additional finding consistent with prediction (v) was the positive association in upland sites between abundance of seedlings of introduced species and EAB-caused ash mortality. These results are consistent with the fluctuating resource hypothesis that a community is more easily invaded when there is an increase in unused resources (Davis et al. 2000), in this case resulting from death of canopy ash trees. Our finding that introduced seedling abundance was associated with ash decline in upland, but not wet, sites suggests that greater availability of water, rather than light, is responsible for this response. While elevated abundance of introduced plants following ash death was predicted by Hausman et al. (2010) and Gandhi and Herms (2010), those authors focused on an increase in light availability. Eschtruth and Battles (2014) demonstrated that even ephemeral forest disturbances can have long-term effects on plant community invasions.
However, we did not find the predicted positive correlation between EAB-caused ash decline and other seedling parameters. Specifically, neither ash decline nor amount of upper strata vegetation had any effect on woody seedling species richness, nor abundance and recruitment of native tree seedlings. Similarly, Davis et al. (2016) and Dolan and Kilgore (2018) found no relationship between relative ash BA and change in seedling density following ash decline. Looney et al. (2017a) found that only the clear-cut treatment resulted in a significant increase in seedling density when compared to girdling, partial overstory removal, and control plots. It is possible that EAB-caused canopy gaps, compared to anthropogenic girdling or cutting, may have too ephemeral of an effect on seedling demography, especially if infilling is also occurring by non-ash canopy trees. Seedling responses may also be a product of species-specific reproductive strategies (e.g., seed bank as compared to advanced regeneration), and exploring these dynamics could be a productive area for future research. Previous studies investigating native seedling recruitment following EAB-caused ash decline focused on Fraxinus seedlings. Kashian (2016) showed in Michigan that recruitment for F. pennsylvanica seedlings was high, but this correlated with a mast year rather than differences among stands in time-since-EAB attack. In Michigan and Ohio, Klooster et al. (2014) found a significant interaction between year and hydrological soil class, where seedling establishment was greatest in 2010 on xeric transects. However, their findings also indicated that Fraxinus spp. seedling increases can be ephemeral as sites where mature ash died prior to a mast year had nearly no seedlings. These Fraxinus spp. seedling recruitment studies indicate factors like seed production, soil hydrology, and overall year-to-year variation can complicate the effects of ash decline. In our analyses, Fraxinus spp. seedlings were grouped with shade-intolerant native seedlings, which did not respond to ash decline.
While the mechanisms proposed were motivated by an underlying assumption that light limits woody plant growth in forests, other resources such as nutrients and water may instead be limiting. The role of other limiting resources may be particularly important for non-ash canopy trees which likely already have access to full sun. Previous ash mortality studies have shown water table rise (Slesak et al. 2014), a decrease in sap flux during early of EAB infestation (Flower et al. 2018), and significant increases in maple tree radial growth without greater sun exposure each support belowground resource availability increasing with ash mortality. Furthermore, changes in belowground resources were attributed to higher fine root densities within red spruce, Picea rubens, decline-caused gaps (Battles and Fahey 2000). As we have shown, canopy openness was not related to EAB-caused ash mortality or to any ecological impacts on forest vegetation. Therefore, our ADI and AMI indices provide a more holistic measure of environmental conditions following EAB invasion. Rather than only measuring light availability, they likely also correlate with increases in water and nutrients that also become available for competing vegetation. Additionally, our findings that the positive associations of both non-ash canopy tree growth and abundance of introduced seedlings with ADI were manifest in upland sites but not wet sites adds evidence that availability of belowground resources, namely water, due to ash mortality is driving these responses.
Distinguishing between the direct effects of ash decline and indirect effects mediated by the various layers of vegetation would be improved by path analysis/structural equation modeling. Path analysis was not possible for our dataset, due to the limited number (24) of study sites, but should be explored if larger datasets become available.
Evaluating the effects of ash mortality on different forest strata enabled us to determine if EAB-caused ash mortality leads to a release in upper (mechanism 1) or lower forest layers (mechanism 2) as well as to assess the interaction between strata. We found evidence to support aspects of both mechanisms. Components of both the upper and lower strata responded positively to EAB-caused ash mortality. Surprisingly, when there was an interaction between layers, it was greater BA or shrub cover from an upper forest layer positively affecting lower layer. Mechanism 1 had strong support since the entire non-ash canopy, including maple, was shown to respond positively to ash loss. For those plants in the lower strata, we did not find as widespread a positive impact from ash death. Instead, positive responses were confined to maples and introduced species, groups that are likely adapted to ephemeral increases in resources. However, maples and introduced species within all layers did not respond similarly; subcanopy maples and introduced shrubs did not respond positively to ash mortality like those in the canopy and seedling layer. Therefore, both stratum (position in the vertical structure) and plant identity should be considered when predicting responses.
5 Conclusion
We found evidence to support both of our mechanisms, but greater evidence for mechanism 1. This indicates that in the short term, ash death releases other tree species in the canopy layer, particularly maples, resulting in rapid gap closure. In turn, the release of canopy trees may minimize the direct effect of EAB-caused ash mortality on lower strata vegetation in general, and dampen the importance of mechanism 2. However, those species that may be adapted to ephemeral resource fluctuations, specifically maple and introduced seedlings, responded positively, providing insight into future forest composition. Achieving greater measurement precision or conducting the study over a greater time span might improve the ability to estimate growth responses. It may also help to determine if differences among and within layer responses are derived from low-resolution data or could be a result from other factors unmeasured in our study (Davis et al. 2016).
Furthermore, focusing only on the short-term effects such as a release in the canopy layer may obscure the more insidious long-term effects of ash mortality; an increase in introduced seedling abundance. Moreover, our results suggest that long-term effects can also be influenced by more upper forest vegetation. Specifically, we found that the subcanopy and shrub layers influence seedling layer responses. Our conclusions support models of short- and long-term effects of forest pests mediated by pest and host tree characteristics (e.g., Lovett et al. 2006), and also demonstrate that the characteristics of the subcanopy and shrub layers are also important mediators of long-term effects on forest composition.
We also illustrated the importance of considering alternative measures to quantify forest pest impacts. Canopy openness alone did not account for the ecological effect of EAB on forest vegetation. Furthermore, we found what seems to be a difference in resource responses strengthening the argument that an inclusive index is superior to measuring a single resource (i.e., light). By adopting the ash mortality indices ADI and AMI, which account for relative abundance of host trees as well as the condition of those trees, we were able to show that EAB-caused ash mortality is associated with a release in non-ash and maple canopy growth. This suggests that in the short-term, ash death releases conspecifics in the canopy layer, resulting in rapid gap closure.
We suggest future investigations of forest community dynamics following the establishment of invasive forest pests should not only distinguish short- and long-term effects, as highlighted by Lovett et al. (2006), but also incorporate initial forest conditions and their ensuing interactions. Our findings also suggest that these structural differences may manifest into very dissimilar trajectories for these communities following an invasion and require very different management approaches.
Statement on data availability
The datasets generated during and/or analyzed during the current study are available in the appendices of this manuscript.
Abbreviations
- ADI:
-
Ash decline index
- AMI:
-
Ash mortality index
- BA:
-
Basal area
- DBH:
-
Diameter at breast height
- EAB:
-
Emerald ash borer
- RGR:
-
Relative growth rate
References
Bates D, Maechler M, Bolker B, Walker S, Christensen RHB, Singmann H, Dai B, Grothendieck G, Green P (2017) Linear mixed-effects models using ‘Eigen’ and S4, ver 1.1-8. http://lme4.r-forge.r-project.org/, Visited May 2017.
Battles JJ, Fahey TJ (2000) Gap dynamics following forest decline: a case study of red spruce forests. Ecol Appl 10:760–774
Beaudet M, Messier C (1998) Growth and morphological responses of yellow birch, sugar maple, and beech seedlings growing under a natural light gradient. Can J For Res 28:1007–1015
Beckage B, Clark JS, Clinton BD, Haines BL (2000) A long-term study of tree seedling recruitment in southern Appalachian forests: the effects of canopy gaps and shrub understories. Can J For Res 30:1617–1631
Bolker BM, Brooks ME, Clark CJ, Geange SW, Poulsen JR, Stevens MHH, White JS (2009) Generalized linear mixed models: a practical guide for ecology and evolution. Trends Ecol Evol 24:127–135
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach. Springer, New York
Burnham KM, Lee TD (2010) Canopy gaps facilitate establishment, growth, and reproduction of invasive Frangula alnus in a Tsuga canadensis dominated forest. Biol Invasions 12:1509–1520
Caquet B, Montpied P, Dreyer E, Epron D, Collet C (2010) Response to canopy opening does not act as a filter to Fagus sylvatica and Acer sp. Advance regeneration in a mixed temperate forest. Ann For Sci 67:105–116
Costilow KC, Knight KS, Flower CE (2017) Disturbance severity and canopy position control the radial growth response of maple trees (Acer spp.) in forests of northwest Ohio impacted by emerald ash borer (Agrilus planipennis). Ann For Sci 74:10
Davis MA, Grime JP, Thompson K (2000) Fluctuating resources in plant communities: a general theory of invasibility. J Ecol 88:528–534
Davis JC, Shannon JP, Bolton NW, Kolka RK, Pypker TG (2016) Vegetation responses to simulated emerald ash borer infestation in Fraxinus nigra dominated wetlands of Upper Michigan, USA. Can J For Res 47:319–330
Diaci J, Gyoerek N, Gliha J, Nagel TA (2008) Response of Quercus robur L. seedlings to north-south asymmetry of light within gaps in flood plain forests of Slovenia. Ann For Sci 65:105–110
Dolan B, Kilgore J (2018) Forest Regeneration Following Emerald Ash Borer (Agrilus planipennis Fairemaire) Enhances Mesophication in Eastern Hardwood Forests. Forests 9:353
Dunn CP (1986) Shrub layer response to death of Ulmus americana in southeastern Wisconsin lowland forests. B Torrey Bot Club 113:142–148
Ehrenfeld JG (1980) Understory response to canopy gaps of varying size in a mature oak forest. B Torrey Bot Club 1:29–41
Elliott KJ, Swank WT (1994) Impacts of drought on tree mortality and growth in a mixed hardwood forest. J Veg Sci 5:229–236
Ellison AM, Bank, M.S., Clinton BD, Colburn EA, Elliott K, Ford CR, Foster DR, Kloeppel BD, Knoepp JD, Lovett GM, Mohan J (2005) Loss of foundation species: consequences for the structure and dynamics of forested ecosystems. Front Ecol Environ 9:479–486
Eschtruth AK, Battles JJ (2014) Ephemeral disturbances have long-lasting impacts on forest invasion dynamics. Ecology 95:1770–1779
Eschtruth AK, Cleavitt NL, Battles JJ, Evans RA, Fahey TJ (2006) Vegetation dynamics in declining eastern hemlock stands: 9 years of forest response to hemlock woolly adelgid infestation. Can J For Res 36:1435–1450
Feeley KJ, Wright SJ, Supardi MN, Kassim AR, Davies SJ (2007) Decelerating growth in tropical forest trees. Ecol Lett 10:461–469
Flower CE, Knight KS, Gonzalez-Meler MA (2013a) Impacts of the emerald ash borer (Agrilus planipennis Fairmaire) induced ash (Fraxinus spp.) mortality on forest carbon cycling and successional dynamics in the eastern United States. Biol Invasions 15:931–944
Flower CE, Knight KS, Rebbeck J, Gonzalez-Meler MA (2013b) The relationship between the emerald ash borer (Agrilus planipennis) and ash (Fraxinus spp.) tree decline: using visual canopy condition assessments and leaf isotope measurements to assess pest damage. For Ecol Manage 303:143–147
Flower CE, Long LC, Knight KS, Rebbeck J, Brown JS, Gonzalez-Meler MA, Whelan CJ (2014) Native bark-foraging birds preferentially forage in infected ash (Fraxinus spp.) and prove effective predators of the invasive emerald ash borer (Agrilus planipennis Fairmaire). For Ecol Manage 313:300–306
Flower CE, Lynch DJ, Knight KS, Gonzalez-Meler MA (2018) Biotic and abiotic drivers of sap flux in mature green ash trees (Fraxinus pennsylvanica) experiencing varying levels of emerald ash borer (Agrilus planipennis) infestation. Forests 9:301
Gandhi KJK, Herms DA (2010) Direct and indirect effects of alien insect herbivores on ecological processes and interactions in forests of eastern North America. Biol Invasions 28:389–405
Gandhi KJ, Smith A, Hartzler DM, Herms DA (2014) Indirect effects of emerald ash borer-induced ash mortality and canopy gap formation on epigaeic beetles. Environ Entomol 43:546–555
Godínez-Alvarez H, Herrick JE, Mattocks M, Toledo D, Van Zee J (2009) Comparison of three vegetation monitoring methods: Their relative utility for ecological assessment and monitoring. Ecological Indicators 9 (5):1001-1008
Haack RA (2006) Exotic bark-and wood-boring Coleoptera in the United States: recent establishments and interceptions. Can J For Res 36:269–288
Hausman CE, Jaeger JF, Rocha OJ (2010) Impacts of the emerald ash borer (EAB) eradication and tree mortality: potential for a secondary spread of invasive plant species. Biol Invasions 12:2013–2023
Herms DA, McCullough DG (2014) Emerald ash borer invasion of North America: History, biology, ecology, impacts, and management. Annu Rev Entomol 59:13–30
Herms DA, Stone AK, Chatfield JA (2004) Emerald ash borer: the beginning of the end of ash in North America? In: Chatfield JA, Draper EA, Mathers HM, Dyke DE, Bennett PF, Boggs JF (eds) Ornamental plants: annual reports and research reviews 2003. OARDC/OSU Extension Special Circular 193:62–71
Hicks DJ, Hustin DL (1989) Response of Hamamelis virginiana L. to canopy gaps in a Pennsylvania oak forest. Am Midl Nat 1:200–204
Higham M, Hoven BM, Gorchov DL, Knight KS (2017) Patterns of Coarse Woody Debris in Hardwood Forests across a Chronosequence of Ash Mortality Due to the Emerald Ash Borer (Agrilus planipennis). Nat Area J 37:406–411
Hobbs RJ, Huenneke LF (1992) Disturbance, diversity, and invasion: implications for conservation. Conserv. Biol. 6:324–337
Holl KD (1998) Do perching structures elevate seed rain and seedling establishment in abandoned tropical pasture. Restor Ecol 6:253–261
Hoven BM, Gorchov DL, Knight KS, Peters VE (2017) The effect of emerald ash borer-caused tree mortality on the invasive shrub Amur honeysuckle and their combined effects on tree and shrub seedlings. Biol Invasions 19:2813–2836
Huenneke LF (1983) Understory response to gaps caused by the death of Ulmus americana in central New York. Bull Torrey Bot Club 110:170–175
Jedlicka J, Vandermeer J, Aviles-Vazquez K, Barros O, Perfecto I (2004) Gypsy moth defoliation of oak trees and a positive response of red maple and black cherry: an example of indirect interaction. Am Midl Nat 152:231–236
Jordano P, Schupp EW (2000) Seed disperser effectiveness: the quantity component and patterns of seed rain for Prunus mahaleb. Ecol Monogr 70:591–615
Kashian DM (2016) Sprouting and seed production may promote persistence of green ash in the presence of the emerald ash borer. Ecosphere 7:4
Kenis M, Auger-Rozenberg MA, Roques A, Timms L, Péré C, Cock MJW, Settele J, Augustin S, Lopez-Vaamonde C (2009) Ecological effects of invasive alien insects. Biol Invasions 11:21–45
Klooster WS (2012) Forest responses to emerald ash borer-induced ash mortality. The Ohio State University, PhD dissertation
Klooster WS, Herms DA, Knight KS, Herms CP, McCullough DG, Smith A, Gandhi KJK, Cardina J (2014) Ash (Fraxinus spp.) mortality, regeneration, and seed bank dynamics in mixed hardwood forests following invasion by emerald ash borer (Agrilus planipennis). Biol Invasions 16:859–873
Knight KS, Brown JP, Long RP (2013) Factors affecting the survival of ash (Fraxinus spp.) trees infested by emerald ash borer (Agrilus planipennis). Biol Invasions 15:371–383
Kovacs KF, Haight RG, McCullough DG, Mercader RJ, Siegert NW, Liebhold AM (2010) Cost of potential emerald ash borer damage in U.S. communities, 2009-2010. Ecol Econ 69:569–578
Lemmon PE (1956) A spherical densiometer for estimating forest overstory density. For Sci 2:314–320
Liebhold AM, McCullough DG, Blackburn LM, Frankel SJ, Von Holle B, Aukema JE (2013) A highly aggregated geographical distribution of forest pest invasions in the USA. Divers Distrib 19:1208–1216
Looney CE, D'Amato AW, Palik BJ, Slesak RA, Slater MA (2017a) The response of Fraxinus nigra forest ground-layer vegetation to emulated emerald ash borer mortality and management strategies in northern Minnesota, USA. For. Ecol Manage 389:352–363
Looney CE, D’Amato AW, Palik BJ, Slesak RA (2017b) Canopy treatment influences growth of replacement tree species in Fraxinus nigra forests threatened by the emerald ash borer in Minnesota, USA. Can J For Res 47:183–192
Lovett GM, Canham CD, Arthur MA, Weathers KC, Fitzhugh RD (2006) Forest ecosystem responses to exotic pests and pathogens in eastern North America. BioScience. 56:395–405
Lovett GM, Arthur MA, Weathers KC, Griffin JM (2013) Effects of introduced insects and diseases on forest ecosystems in the Catskill Mountains of New York. Ann NY Acad Sci 1298:66–77
Lovett GM, Weiss M, Liebhold AM, Holmes TP, Leung B, Lambert KF, Orwig DA, Campbell FT, Rosenthal J, McCullough DG, Wildova R (2016) Nonnative forest insects and pathogens in the United States: impacts and policy options. Ecol Appl 26:1437–1455
Luken JO, Kuddes LM, Tholemeier TC, Haller DM (1997) Comparative Responses of Lonicera maackii (Amur Honeysuckle) and Lindera benzoin (Spicebush) to Increased Light. American Midland Naturalist 138 (2):331
Margulies E, Bauer L, Ibáñez I (2017) Buying Time: Preliminary Assessment of Biocontrol in the Recovery of Native Forest Vegetation in the Aftermath of the Invasive Emerald Ash Borer. Forests 8:369
Marks PL, Gardescu S (1998) A case study of sugar maple (Acer saccharum) as a forest seedling bank species. J Torrey Bot Soc 1:287–296
Mazerolle M (2015) Model selection and multimodel inference base on (Q)AIC(c). ver 2.0-2. http://cran.r-project.org/web/packages/AICcmodavg, Visited (December 2017)
Muscolo A, Bagnato S, Sidari M, Mercurio R (2014) A review of the roles of forest canopy gaps. J Forest Res 25:725–736
Muzika RM, Liebhold AM (1999) Changes in radial increment of host and nonhost tree species with gypsy moth defoliation. Can J For Res 29:1365–1373
Niinemets U, Valladares F (2006) Tolerance to shade, drought, and waterlogging of temperate northern hemisphere trees and shrubs. Ecol Monogr 76:521–547
Orwig DA, Foster DR (1998) Forest response to the introduced hemlock woolly adelgid in southern New England, USA. J Torrey Bot Soc 1:60–73
Pacala SW, Canham CD, Silander JA Jr, Kobe RK (1994) Sapling growth as a function of resources in a north temperate forest. Can J For Res 24:2172–2183
Palik BJ, Ostry ME, Venette RC, Abdela E (2011) Fraxinus nigra (black ash) dieback in Minnesota: regional variation and potential contributing factors. Forest Ecol Manag 261:128–135
Perry KI, Herms DA (2016) Response of the forest floor invertebrate community to canopy gap formation caused by early stages of emerald ash borer-induced ash mortality. For Ecol Manage 375:259–267
Perry KI, Herms DA (2017) Effects of Late Stages of Emerald Ash Borer (Coleoptera: Buprestidae)-Induced Ash Mortality on Forest Floor Invertebrate Communities. J Insect Sci 17:119
Perry KI, Herms DA, Klooster WS, Smith A, Hartzler DM, Coyle DR, Gandhi KJ (2018) Downed Coarse Woody Debris Dynamics in Ash (Fraxinus spp.) Stands Invaded by Emerald Ash Borer (Agrilus planipennis Fairmaire). Forests 9:191
R Development Core Team (2017) The R Foundation for Statistical Computing, ver 3.2.1. Vienna, Austria: Vienna University of Technology. http://www.r-project.org/, Visited May
Rooney TP, Waller DM (2003) Direct and indirect effects of white-tailed deer in forest ecosystems. Forest Ecol Manag 181:165–176
Runkle JR (2007) Impacts of beech bark disease and deer browsing on the old-growth forest. Am Midl Nat 157:241–249
Sapkota IP, Tigabu M, Odén PC (2009) Species diversity and regeneration of old-growth seasonally dry Shorea robusta forests following gap formation. J For Res 20:7–14
Savage MB, Rieske LK (2018) Coleopteran Communities Associated with Forests Invaded by Emerald Ash Borer. Forests 9:69
Sinclair WA, Griffiths HM (1994) Ash yellows and its relationship to dieback and decline of ash. Annu Rev Phytopathol 32:49–60
Slesak RA, Lenhart CF, Brooks KN, D’Amato AW, Palik BJ (2014) Water table response to harvesting and simulated emerald ash borer mortality in black ash wetlands in Minnesota, USA. Can J For Res 44:961–968
Spei BA, Kashian DM (2017) Potential for persistence of blue ash in the presence of emerald ash borer in southeastern Michigan. Forest Ecol Manag 392:137–143
Strickler GS (1959) Use of the densiometer to estimate density of forest canopy on permanent sample plots
Twolan-Strutt L, Keddy PA (1996) Above-and belowground competition intensity in two contrasting wetland plant communities. Ecology 77:259–270
Wagner DL (2010) Caterpillars of eastern North America: a guide to identification and natural history. Princeton University Press, Princeton
Wildman RH (2008) Ohio’s forest resources, 2006. Res. Note. NRS-22 U.S. Department of Agriculture, Forest Service, Northern Research Station, Newton Square, PA
Acknowledgments
We thank the land managers of the study sites for permitting access and allowing this work to be conducted within the research plots. We thank the many field technicians that assisted in data collection and entry, particularly E. DeBurgomaster, M. Higham, D. Weeks, G. Hoven, J. Hoven, B. Flash, C. Flower, R. Ford, T. Fox, B. Gombash, R. Hefflinger, S. Jasani, J. Jolliff, P. Jones, S. Kelsey, T. Macy, M. Marshall, E. Monarch, Z. Morvay, R. Kappler, S. Starr, S. Stroebel, J. Throckmorton, and B. Wiggin. We would also like to thank R.M. Ward, T. Crist, H. Stevens, N. Money, and J. Maingi for their assistance in developing this study. Long-term monitoring plot data collection was supported by the USDA Forest Service and USDA APHIS. We thank Alejandro Royo De Sedas, Charlie Flower, and anonymous reviewers for valuable comments on earlier versions of this manuscript.
Funding
This study is financially supported by the following: Botanical Society of America Graduate Student Research Award, Sigma Xi, and the Academic Challenge programs of the Miami University Botany and Biology Departments.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Handling Editor:
Aurélien Sallé
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Appendices
Appendix 1
Location and summary data for 72 study plots (24 sites) in northwestern, central, and southwestern Ohio, USA. Forest type: based on topography and seasonal pressence of standing water; ash species: species with greatest canopy BA; ASH BA %: percentage of stand BA that is ash; non-ash canopy: two species that are largest percentage of stand BA (a single species indicates only one non-ash species present in the plot canopy); ADI: (ash decline index) total BA of ash trees rated 3–5 divided by stand BA; AMI: (ash mortality index) BA of dead ash trees rated 5 divided by stand BA. All data were obtained at the plot level. Values for canopy openness, ADI, and AMI are based on 2012 measurements. Plot abbreviations: CCG (Caesar Creek Gorge SNP), CCSP (Caesar Creek State Park), CLF (Clifton Gorge SNP), CLB (Culberson Woods SNP), DMP (Demsey Middle School), EDW (Edwards Furniture), ENG (Englewood Metropark), FT (Fallen Timbers), GHN (Gahanna Woods SNP), GRM (Germantown Metropark), GLN (Glenwood Gardens), HGH (Highbanks Metropark), HST (Hueston Woods SNP) MSF (Maumee State Forest), OO (Oak Openings), PM (Pearson Metropark), SYM (Seymour Woods SNP), SWCN (Sharon Woods Cincinnati), SWCL (Sharon Woods Columbus), STR (Stratford Ecological Center)
Plot | Latitude | Longitude | Region | Forest type | Ash species | Ash BA % | Non-ash canopy | ADI | AMI |
CCG 1 | 39.490 | − 84.098 | SW | Upland | F. quadrangulata; F. americana | 54% | Ulmus americana Juniperus virginiana | 3% | 0% |
CCG 2 | 39.487 | − 84.093 | SW | Upland | F. americana | 45% | Gymnocladus dioicus Celtis occidentalis | 32% | 0% |
CCG 3 | 39.487 | − 84.092 | SW | Upland | F. americana | 54% | Celtis occidentalis Quercus muehlenbergii | 9% | 0% |
CCSP 1_1 | 39.489 | − 84.037 | SW | Upland | F. americana | 18% | Juglans nigra Acer saccharum | 2% | 2% |
CCSP 1_2 | 39.490 | − 84.037 | SW | Upland | F. americana | 28% | Fagus grandifolia Carya cordiformis | 0% | 0% |
CCSP 1_3 | 39.490 | − 84.038 | SW | Upland | F. americana | 18% | Liriodendron tulipifera Juglans nigra | 9% | 5% |
CCSP 4_8 | 39.507 | − 84.050 | SW | Upland | F. americana | 42% | Acer saccharum | 0% | 0% |
CCSP 4_9 | 39.507 | − 84.051 | SW | Upland | F. americana | 36% | Acer saccharum | 12% | 0% |
CCSP 4_9B | 39.506 | − 84.051 | SW | Upland | F. americana | 36% | Acer saccharum | 9% | 9% |
CCSP 5_10 | 39.536 | − 84.003 | SW | Upland | F. americana | 25% | Quercus rubra Carya cordiformis | 0% | 0% |
CCSP 5_11 | 39.537 | − 84.003 | SW | Upland | F. americana | 23% | Quercus muehlenbergii Celtis occidentalis | 3% | 3% |
CCSP 5_12 | 39.537 | − 84.003 | SW | Upland | F. americana | 31% | Liriodendron tulipifera Quercus rubra | 13% | 0% |
CLB 1 | 39.369 | − 83.935 | SW | Lowland | F. pennsylvanica/profunda | 36% | Acer rubrum | 4% | 4% |
CLB 2 | 39.369 | − 83.934 | SW | Lowland | F. pennsylvanica/profunda | 25% | Acer rubrum Carya ovata | 7% | 0% |
CLB 3 | 39.370 | − 83.934 | SW | Lowland | F. pennsylvanica/profunda | 35% | Acer rubrum Quercus bicolor | 0% | 0% |
CLF 1 | 39.792 | − 83.841 | SW | Upland | F. quadrangulata | 16% | Juglans nigra Acer saccharum | 3% | 3% |
CLF 2 | 39.792 | − 83.841 | SW | Upland | F. quadrangulata | 41% | Acer saccharum Celtis occidentalis | 2% | 0% |
CLF 3 | 39.794 | − 83.840 | SW | Upland | F. quadrangulata; F. pennsylvanica/profunda | 27% | Liriodendron tulipifera Quercus rubra | 14% | 1% |
DMP 1 | 40.308 | − 83.085 | C | Upland | F. americana | 43% | Ulmus americana Ulmus sp. | 30% | 23% |
DMP 2 | 40.308 | − 83.085 | C | Upland | F. americana | 19% | Populus deltoides Quercus palustris | 18% | 18% |
DMP 3 | 40.308 | − 83.085 | C | Upland | F. americana | 26% | Acer negundo Ulmus sp. | 25% | 25% |
EDW 1 | 39.567 | − 84.260 | SW | Upland | F. americana; F. quadrangulata | 62% | Acer saccharum Ulmus rubra | 39% | 39% |
EDW 2 | 39.567 | − 84.260 | SW | Upland | F. americana | 40% | Prunus serotina Acer saccharum | 40% | 40% |
EDW 3 | 39.567 | − 84.261 | SW | Upland | F. americana | 20% | Acer saccharum Ulmus rubra | 19% | 16% |
ENG 1 | 39.888 | − 84.283 | SW | Lowland | F. pennsylvanica/profunda | 95% | Celtis occidentalis Carya ovata | 22% | 4% |
ENG 2 | 39.888 | − 84.283 | SW | Lowland | F. nigra; F. pennsylvanica/profunda | 56% | Gleditsia triacanthos Carya ovata | 35% | 30% |
ENG 3 | 39.889 | − 84.284 | SW | Lowland | F. pennsylvanica/profunda | 44% | Quercus ruba Gleditsia triacanthos | 15% | 3% |
FT 1_1 | 41.550 | − 83.694 | NW | Lowland | F. pennsylvanica/profunda | 41% | Populus deltoides Quercus rubra | 41% | 41% |
FT 1_2 | 41.550 | − 83.694 | NW | Lowland | F. pennsylvanica/profunda | 41% | Quercus rubra Populus deltoides | 41% | 41% |
FT 1_3 | 41.551 | − 83.696 | NW | Lowland | F. pennsylvanica/profunda | 29% | Acer rubrum Quercus rubra | 29% | 29% |
FT 2_1 | 41.554 | − 83.693 | NW | Lowland | F. pennsylvanica/profunda | 33% | Acer saccharinum Acer rubrum | 33% | 33% |
FT 2_2 | 41.554 | − 83.694 | NW | Lowland | F. pennsylvanica/profunda | 28% | Acer saccharinum Acer rubrum | 28% | 28% |
FT 2_3 | 41.553 | − 83.695 | NW | Lowland | F. pennsylvanica/profunda | 27% | Acer saccharinum Acer rubrum | 27% | 27% |
GHN 1 | 40.010 | − 82.836 | C | Lowland | F. pennsylvanica/profunda | 18% | Acer saccharinum Populus deltoides | 2% | 0% |
GHN 2 | 40.010 | − 82.835 | C | Lowland | F. pennsylvanica/profunda | 48% | Acer saccharinum Salix sp. | 0% | 0% |
GHN 3 | 40.010 | − 82.835 | C | Lowland | F. pennsylvanica/profunda | 57% | Ulmus sp. Populus deltoides | 14% | 0% |
GLN 1 | 39.257 | − 84.486 | SW | Upland | F. quadrangulata; F. americana | 90% | Acer saccharum Acer nigrum | 12% | 0% |
GLN 2 | 39.257 | − 84.486 | SW | Upland | F. quadrangulata; F. americana | 58% | Quercus rubra Acer saccharum | 50% | 20% |
GLN 3 | 39.257 | − 84.485 | SW | Upland | F. quadrangulata | 28% | Acer saccharum Acer nigrum | 0% | 0% |
GRM 1 | 39.639 | − 84.397 | SW | Upland | F. americana | 19% | Liriodendron tulipifera Quercus rubra | 0% | 0% |
GRM 2 | 39.639 | − 84.397 | SW | Upland | F. americana | 47% | Juglans nigra Ulmus americana | 10% | 5% |
GRM 3 | 39.639 | − 84.399 | SW | Upland | F. americana | 44% | Acer saccharum Robinia pseudoacacia | 4% | 0% |
HGH 1 | 40.145 | − 83.027 | C | Upland | F. americana | 49% | Acer rubrum Sassafras albidum | 46% | 41% |
HGH 2 | 40.145 | − 83.027 | C | Upland | F. americana | 51% | Acer rubrum Acer saccharum | 45% | 42% |
HGH 3 | 40.145 | − 83.026 | C | Upland | F. americana | 52% | Acer rubrum Prunus serotina | 44% | 36% |
HST 1 | 39.570 | − 84.754 | SW | Upland | F. americana | 21% | Fagus grandifolia Liriodendron tulipifera | 8% | 0% |
HST 2 | 39.570 | − 84.753 | SW | Upland | F. americana | 60% | Acer saccharum Ulmussp. | 60% | 25% |
HST 3 | 39.570 | − 84.753 | SW | Upland | F. americana | 52% | Fagus grandifolia Acer saccharum | 52% | 0% |
MSF 1_1 | 41.537 | − 83.930 | NW | Lowland | F. pennsylvanica/profunda | 32% | Quercus bicolor Acer rubrum | 31% | 31% |
MSF 1_2 | 41.537 | − 83.931 | NW | Lowland | F. pennsylvanica/profunda | 30% | Acer rubrum Quercus alba | 30% | 30% |
MSF 1_3 | 41.536 | − 83.931 | NW | Lowland | F. pennsylvanica/profunda | 63% | Quercus alba Acer rubrum | 63% | 63% |
OO 2_1 | 41.561 | − 83.868 | NW | Floodplain | F. pennsylvanica/profunda | 55% | Populus deltoides Acer saccharinum | 55% | 55% |
OO 2_2 | 41.562 | − 83.868 | NW | Floodplain | F. pennsylvanica/profunda | 47% | Acer rubrum Acer saccharinum | 47% | 47% |
OO 2_3 | 41.561 | − 83.868 | NW | Floodplain | F. pennsylvanica/profunda | 45% | Populus deltoides Ulmus sp. | 45% | 45% |
OO 3_1 | 41.542 | − 83.848 | NW | Floodplain | F. pennsylvanica/profunda | 68% | Quercus bicolor Acer negundo | 68% | 68% |
OO 3_2 | 41.543 | − 83.848 | NW | Floodplain | F. pennsylvanica/profunda | 16% | Gleditsia triacanthos Quercus rubra | 16% | 16% |
OO 3_3 | 41.543 | − 83.849 | NW | Floodplain | F. pennsylvanica/profunda | 45% | Quercus bicolor Gleditsia triacanthos | 45% | 45% |
PM 1_4 | 41.639 | − 83.434 | NW | Lowland | F. pennsylvanica/profunda | 59% | Acer saccharinum Acer rubrum | 59% | 59% |
PM 1_5 | 41.641 | − 83.435 | NW | Lowland | F. pennsylvanica/profunda | 81% | Acer negundo Juglans nigra | 81% | 81% |
PM 1_6 | 41.643 | − 83.436 | NW | Lowland | F. pennsylvanica/profunda | 15% | Populus deltoides Acer rubrum | 15% | 15% |
STR 1 | 40.255 | − 83.071 | C | Upland | F. americana | 27% | Acer saccharum Quercus alba | 27% | 27% |
STR 2 | 40.255 | − 83.072 | C | Upland | F. americana | 16% | Acer saccharum Prunus serotina | 16% | 16% |
STR 3 | 40.255 | − 83.073 | C | Upland | F. americana | 58% | Quercus velutina Quercus alba | 58% | 58% |
SWCL 2_4 | 40.120 | − 82.971 | C | Upland | F. americana; F. quadrangulata | 71% | Aesculus sp. Acer saccharum | 46% | 46% |
SWCL 2_5 | 40.120 | − 82.971 | C | Upland | F. americana; F. quadrangulata | 29% | Juglans nigra Acer saccharum | 24% | 18% |
SWCL 2_6 | 40.121 | − 82.969 | C | Upland | F. quadrangulata | 9% | Ulmus americana Acer negundo | 3% | 3% |
SWCN 1 | 39.281 | − 84.397 | SW | Upland | F. americana; F. quadrangulata | 30% | Acer saccharum Tilia americana | 0% | 0% |
SWCN 2 | 39.281 | − 84.397 | SW | Upland | F. americana | 52% | Quercus rubra Acer saccharum | 20% | 4% |
SWCN 3 | 39.281 | − 84.396 | SW | Upland | F. americana | 50% | Quercus muehlenbergii Acer saccharum | 20% | 0% |
SYM 1_1 | 40.228 | − 83.055 | C | Upland | F. americana | 39% | Quercus palustris Juglans nigra | 39% | 32% |
SYM 1_2 | 40.228 | − 83.056 | C | Upland | F. americana | 41% | Acer saccharum Prunus serotina | 37% | 28% |
SYM 1_3 | 40.228 | − 83.056 | C | Upland | F. americana | 16% | Prunus serotina Robinia pseudoacacia | 16% | 8% |
Mean | NA | NA | NA | NA | NA | 40% | NA | 25% | 19% |
Appendix 2
Woody plant initial size (2012) and vegetation strata predictor variable measurements. Measurements collected in northwestern, central, and southwestern Ohio, USA, within 72 plots (24 sites). Stand BAs were measured at the plot level (400 m2). Subcanopy BAs, shrub cover, and shrub BAs were each measured at the sub-plot level (200 m2). Canopy openness and sugar maple height were measured within four micro-plots (4 m2) per plot
Plot | Total stand BA (m2/ha) | Non-ash stand BA (m2/ha) | Maple stand BA (m2/ha) | Subcanopy BA (m2/ha) | Maple subcanopy BA (m2/ha) | Shrub cover percent | Native shrub BA (m2/ha) | Introduced shrub BA (m2/ha) | Canopy openness | Mean sugar maple seedling height |
CCG 1 | 29.8 | 14.6 | 1.6 | 0.8 | 0.0 | 93 | 0.39 | 2.86 | 3.6 | NA |
CCG 2 | 26.3 | 14.9 | 1.0 | 2.4 | 0.2 | 87 | 0.01 | 1.42 | 3.1 | NA |
CCG 3 | 29.9 | 14.1 | 0.3 | 3.2 | 0.0 | 78 | 0.52 | 2.62 | 3.5 | NA |
CCSP 1_1 | 33.1 | 27.1 | 6.9 | 2.0 | 0.8 | 58 | NA | 0.58 | 3.6 | NA |
CCSP 1_2 | 29.0 | 20.9 | 0.8 | 2.3 | 0.8 | 2 | 0.00 | 0.06 | 3.7 | 64.0 |
CCSP 1_3 | 28.6 | 23.4 | 2.0 | 1.0 | 0.4 | 76 | 0.26 | 0.53 | 6.4 | 49.5 |
CCSP 4_8 | 31.8 | 18.3 | 10.4 | 0.9 | 0.2 | 13 | 0.11 | 0.16 | 12 | 40.2 |
CCSP 4_9 | 78.7 | 43.8 | 12.5 | 2.0 | 0.9 | 18 | NA | 0.02 | 8.1 | 47.2 |
CCSP 4_9B | 36.5 | 23.4 | 7.9 | 2.8 | 1.4 | 20 | 0.07 | 0.18 | 7.1 | 32.0 |
CCSP 5_10 | 40.1 | 30.1 | NA | 2.9 | 0.1 | 93 | 0.35 | 0.73 | 3.2 | NA |
CCSP 5_11 | 28.8 | 22.2 | NA | 2.3 | 0.0 | 98 | 0.22 | 0.54 | 3.4 | NA |
CCSP 5_12 | 59.1 | 41.1 | 2.4 | 0.6 | 0.3 | 67 | 0.34 | 0.27 | 5 | NA |
CLB 1 | 38.1 | 24.3 | 24.3 | 0.2 | 0.0 | 73 | 0.40 | 0.00 | 4.4 | NA |
CLB 2 | 41.6 | 26.7 | 22.7 | 0.2 | 0.0 | 73 | 0.32 | 0.00 | 4.5 | NA |
CLB 3 | 61.5 | 40.2 | 14.4 | 0.5 | 0.1 | 76 | 1.05 | 0.00 | 3.6 | NA |
CLF 1 | 27.4 | 23.0 | 8.1 | 0.2 | 0.1 | 60 | 0.06 | 1.23 | 4.3 | NA |
CLF 2 | 31.8 | 18.6 | 12.7 | 1.5 | 0.2 | 58 | 0.35 | 0.41 | 4 | 22.0 |
CLF 3 | 49.8 | 36.4 | 8.4 | 0.3 | 0.2 | 47 | 0.34 | 0.08 | 5.7 | NA |
DMP 1 | 14.0 | 8.2 | 0.9 | 5.2 | 0.4 | 67 | 0.02 | 0.47 | 3.7 | NA |
DMP 2 | 41.6 | 34.0 | 1.5 | 2.5 | 1.1 | 96 | NA | 1.69 | 3.8 | NA |
DMP 3 | 15.9 | 12.1 | 6.4 | 1.8 | 0.2 | 98 | 0.20 | 2.17 | 1.8 | NA |
EDW 1 | 51.9 | 20.3 | 3.7 | 3.2 | 1.4 | 73 | NA | 0.86 | 6.8 | NA |
EDW 2 | 39.2 | 23.9 | 5.7 | 1.8 | 0.6 | 36 | NA | 0.15 | 8.8 | 27.2 |
EDW 3 | 20.3 | 16.2 | 7.4 | 1.6 | 0.8 | 20 | NA | 0.15 | 7.9 | 27.8 |
ENG 1 | 29.0 | 2.4 | 0.4 | 4.0 | 0.2 | 18 | 0.21 | 0.02 | 12.2 | NA |
ENG 2 | 34.0 | 15.5 | 2.8 | 1.8 | 0.0 | 31 | 0.20 | 0.04 | 12.7 | NA |
ENG 3 | 33.7 | 18.9 | 1.3 | 1.5 | 0.1 | 40 | 0.15 | 0.05 | 12.7 | 36.0 |
FT 1_1 | 39.6 | 23.3 | 2.6 | 0.8 | 0.1 | 4 | 0.06 | 0.02 | 10.3 | NA |
FT 1_2 | 43.0 | 25.3 | 2.6 | 0.9 | 0.0 | 4 | 0.00 | 0.27 | 7.2 | NA |
FT 1_3 | 42.6 | 30.4 | 12.1 | 1.1 | 0.0 | 4 | 0.01 | 0.32 | 6.3 | NA |
FT 2_1 | 50.1 | 33.7 | 31.9 | NA | 0.0 | 7 | 0.14 | 0.07 | 6.8 | NA |
FT 2_2 | 32.9 | 23.5 | 22.8 | 0.5 | 0.2 | 2 | NA | 0.01 | 5.1 | NA |
FT 2_3 | 61.8 | 45.2 | 44.3 | 0.6 | 0.3 | 2 | NA | 0.02 | 6.4 | NA |
GHN 1 | 12.3 | 10.7 | 4.5 | 5.3 | 2.0 | 16 | 1.28 | 0.00 | 4 | NA |
GHN 2 | 19.6 | 10.8 | 5.4 | 7.8 | 0.6 | 4 | 0.10 | 0.10 | 5 | NA |
GHN 3 | 15.2 | 6.6 | 2.1 | 5.5 | 0.6 | 2 | NA | NA | 3.1 | NA |
GLN 1 | 49.7 | 5.0 | 3.6 | 3.0 | 0.2 | 9 | 1.28 | 0.02 | 10.5 | 27.0 |
GLN 2 | 49.3 | 20.9 | 5.8 | 2.7 | 0.0 | 11 | NA | 0.05 | 4.7 | 46.3 |
GLN 3 | 26.7 | 13.1 | 12.7 | NA | 0.0 | 7 | 0.04 | NA | 7.7 | 29.5 |
GRM 1 | 41.0 | 33.5 | 3.3 | 0.1 | 0.0 | 16 | 0.13 | 0.07 | 19 | 37.0 |
GRM 2 | 27.0 | 14.2 | 1.8 | 2.9 | 0.3 | 13 | 0.01 | 0.08 | 15.9 | 38.4 |
GRM 3 | 42.0 | 23.5 | 9.2 | 1.4 | 0.1 | 29 | 0.00 | 0.39 | 11.4 | 28.3 |
HGH 1 | 30.0 | 16.2 | 7.6 | 0.8 | 0.3 | 44 | 0.31 | 0.19 | 8.9 | NA |
HGH 2 | 32.1 | 16.9 | 16.2 | 1.1 | 0.2 | 62 | 0.12 | 0.17 | 7.2 | NA |
HGH 3 | 26.6 | 14.1 | 7.6 | 2.5 | 0.7 | 49 | 0.08 | 0.06 | 5.5 | NA |
HST 1 | 43.2 | 34.1 | 2.7 | 0.5 | 0.2 | 11 | 0.06 | 0.02 | 6.6 | 23.7 |
HST 2 | 60.7 | 24.1 | 13.0 | 0.5 | 0.2 | 29 | 0.11 | 0.01 | 4.4 | 21.8 |
HST 3 | 41.8 | 20.7 | 3.5 | 0.5 | 0.3 | 36 | 0.19 | 0.01 | 4.7 | 27.9 |
MSF 1_1 | 32.8 | 22.5 | 4.8 | 1.3 | 0.6 | 7 | 0.09 | 0.01 | 8.5 | NA |
MSF 1_2 | 48.3 | 33.8 | 20.5 | 0.1 | 0.0 | 11 | NA | 0.07 | 9 | NA |
MSF 1_3 | 35.6 | 13.4 | 3.2 | 2.5 | 1.1 | 7 | NA | 0.01 | 9.1 | NA |
OO 2_1 | 79.5 | 36.0 | 15.2 | NA | 0.0 | 0 | NA | NA | 5.7 | NA |
OO 2_2 | 61.4 | 32.4 | 27.7 | 1.8 | 0.0 | 0 | NA | NA | 12.6 | NA |
OO 2_3 | 53.3 | 29.5 | 4.7 | 0.4 | 0.0 | 0 | NA | NA | 37.4 | NA |
OO 3_1 | 50.3 | 16.1 | 5.0 | 1.8 | 0.6 | 0 | NA | 0.01 | 25.1 | NA |
OO 3_2 | 49.0 | 41.3 | 0.8 | 1.5 | 0.0 | 0 | NA | NA | 9.8 | NA |
OO 3_3 | 55.1 | 31.3 | 4.3 | 1.3 | 0.5 | 11 | 0.24 | 0.03 | 11.4 | NA |
PM 1_4 | 73.0 | 30.3 | 29.3 | 1.3 | 0.4 | 22 | 0.28 | 0.07 | 7.7 | NA |
PM 1_5 | 44.2 | 8.5 | 2.5 | 0.5 | 0.2 | 64 | 0.52 | 0.04 | 7.9 | NA |
PM 1_6 | 60.4 | 51.1 | 14.4 | 0.7 | 0.1 | 24 | 0.18 | 0.00 | 6.4 | NA |
STR 1 | 49.7 | 36.1 | 18.0 | 1.7 | 0.3 | 0 | NA | NA | 5.7 | 29.3 |
STR 2 | 23.8 | 21.5 | 14.5 | 2.6 | 0.9 | 9 | NA | 0.01 | 6 | 29.5 |
STR 3 | 52.3 | 22.2 | 2.3 | 3.4 | 1.1 | 0 | NA | 0.01 | 4.5 | 27.7 |
SWCL 2_4 | 18.6 | 5.8 | 1.6 | 7.2 | 0.3 | 42 | 0.01 | 0.15 | 9.9 | 20.0 |
SWCL 2_5 | 20.4 | 14.9 | 5.7 | 4.4 | 1.5 | 44 | 0.10 | 0.30 | 8 | 35.5 |
SWCL 2_6 | 17.9 | 16.4 | 8.4 | 2.3 | 0.2 | 24 | 0.06 | 0.20 | 6.7 | NA |
SWCN 1 | 26.5 | 19.6 | 8.8 | 0.1 | 0.1 | 62 | 0.01 | 0.05 | 3.4 | 39.3 |
SWCN 2 | 49.5 | 23.6 | 5.8 | 0.1 | 0.0 | 96 | NA | 0.06 | 1.4 | NA |
SWCN 3 | 41.6 | 16.3 | 6.1 | 0.3 | 0.1 | 64 | 0.01 | 0.45 | 3.6 | 31.8 |
SYM 1_1 | 35.6 | 21.9 | 4.6 | 0.8 | 0.2 | 2 | NA | 0.08 | 3.8 | NA |
SYM 1_2 | 27.4 | 16.3 | 10.6 | 0.8 | 0.3 | 4 | NA | 0.06 | 4 | NA |
SYM 1_3 | 37.9 | 32.1 | 9.8 | 1.0 | 0.4 | 13 | NA | 0.05 | 4.4 | NA |
Mean | 39.1 | 23.0 | 8.7 | 1.8 | 0.4 | 34 | 0.23 | 0.32 | 7.4 | 33.6 |
Appendix 3
Woody plant strata final size (2014). Measurements collected in northwestern, central, and southwestern Ohio, USA, within 72 plots (24 sites). Stand BAs were measured at the plot level (400 m2). Subcanopy BAs and shrub BAs were measured at the sub-plot level (200 m2). Sugar maple height was measured within four micro-plots (4 m2) per plot
Plot | Non-ash stand BA (m2/ha) | Maple stand BA (m2/ha) | Subcanopy BA (m2/ha) | Maple subcanopy BA (m2/ha) | Native shrub BA (m2/ha) | Introduced shrub BA (m2/ha) | Mean sugar maple seedling height |
CCG 1 | 14.8 | 1.7 | 0.8 | 0.0 | 0.4 | 3.1 | NA |
CCG 2 | 15.7 | 1.2 | 2.6 | 0.3 | 0.0 | 1.6 | NA |
CCG 3 | 14.9 | 0.4 | 3.5 | 0.0 | 0.6 | 2.8 | NA |
CCSP 1_1 | 28.4 | 7.7 | 2.1 | 0.9 | NA | 0.6 | NA |
CCSP 1_2 | 22.0 | 1.0 | 2.8 | 0.9 | 0.0 | 0.1 | 48.0 |
CCSP 1_3 | 24.6 | 2.3 | 1.1 | 0.4 | 0.3 | 0.7 | 54.5 |
CCSP 4_8 | 18.6 | 10.6 | 1.1 | 0.3 | 0.2 | 0.3 | 38.8 |
CCSP 4_9 | 44.5 | 13.1 | 2.6 | 1.1 | NA | 0.0 | 57.2 |
CCSP 4_9B | 24.2 | 8.3 | 3.0 | 1.5 | 0.1 | 0.2 | 33.2 |
CCSP 5_10 | 32.1 | NA | 3.1 | 0.1 | 0.3 | 0.8 | NA |
CCSP 5_11 | 21.9 | NA | 2.6 | 0.0 | 0.3 | 0.6 | NA |
CCSP 5_12 | 43.0 | 2.6 | 0.7 | 0.3 | 0.3 | 0.4 | NA |
CLB 1 | 26.7 | 26.7 | 0.3 | 0.0 | 0.5 | 0.0 | NA |
CLB 2 | 28.0 | 23.9 | 0.2 | 0.0 | 0.4 | 0.0 | NA |
CLB 3 | 42.0 | 15.0 | 0.6 | 0.1 | 0.7 | 0.0 | NA |
CLF 1 | 23.8 | 8.4 | 0.2 | 0.1 | 0.0 | 1.5 | NA |
CLF 2 | 19.1 | 13.1 | 1.5 | 0.2 | 0.4 | 0.5 | 18.0 |
CLF 3 | 37.0 | 8.7 | 0.3 | 0.2 | 0.4 | 0.1 | NA |
DMP 1 | 9.0 | 0.9 | 5.8 | 0.5 | 0.0 | 0.5 | NA |
DMP 2 | 36.3 | 1.7 | 2.9 | 1.3 | NA | 1.9 | NA |
DMP 3 | 13.3 | 6.8 | 2.4 | 0.2 | 0.3 | 2.3 | NA |
EDW 1 | 22.0 | 5.1 | 3.7 | 1.6 | NA | 0.9 | NA |
EDW 2 | 26.9 | 7.1 | 2.1 | 0.7 | NA | 0.3 | 30.6 |
EDW 3 | 17.6 | 8.6 | 1.7 | 0.9 | NA | 0.2 | 28.2 |
ENG 1 | 2.6 | 0.4 | 4.4 | 0.2 | 0.2 | 0.0 | NA |
ENG 2 | 16.0 | 3.1 | 1.9 | 0.0 | 0.2 | 0.1 | NA |
ENG 3 | 19.9 | 1.3 | 1.6 | 0.2 | 0.2 | 0.1 | 45.0 |
FT 1_1 | 28.2 | 3.0 | 0.8 | 0.1 | 0.1 | 0.0 | NA |
FT 1_2 | 26.7 | 2.7 | 1.0 | 0.0 | 0.0 | 0.2 | NA |
FT 1_3 | 31.6 | 12.3 | 1.2 | 0.0 | 0.0 | 0.3 | NA |
FT 2_1 | 36.6 | 34.8 | NA | 0.0 | 0.1 | 0.1 | NA |
FT 2_2 | 25.3 | 24.5 | 0.6 | 0.2 | NA | 0.0 | NA |
FT 2_3 | 48.2 | 47.3 | 0.6 | 0.3 | NA | 0.0 | NA |
GHN 1 | 14.3 | 6.6 | 6.4 | 2.4 | 0.7 | 0.0 | NA |
GHN 2 | 12.5 | 6.7 | 8.5 | 0.0 | 0.1 | 0.1 | NA |
GHN 3 | 8.5 | 2.7 | 6.5 | 0.7 | NA | NA | NA |
GLN 1 | 5.7 | 4.0 | 3.4 | 0.7 | 0.7 | 0.0 | 29.0 |
GLN 2 | 22.8 | 6.2 | 3.4 | 0.2 | NA | 0.2 | 55.8 |
GLN 3 | 14.2 | 13.7 | NA | 0.0 | 0.0 | NA | 37.9 |
GRM 1 | 35.1 | 3.5 | 0.1 | 0.0 | 0.1 | 0.1 | 34.7 |
GRM 2 | 15.0 | 1.9 | 3.3 | 0.0 | 0.0 | 0.1 | 44.2 |
GRM 3 | 24.4 | 9.7 | 1.6 | 0.4 | 0.0 | 0.7 | 31.1 |
HGH 1 | 18.3 | 8.5 | 0.9 | 0.2 | 0.4 | 0.2 | NA |
HGH 2 | 18.7 | 18.0 | 1.2 | 0.3 | 0.2 | 0.2 | NA |
HGH 3 | 16.1 | 8.7 | 2.8 | 0.3 | 0.1 | 0.1 | NA |
HST 1 | 34.7 | 3.0 | 0.5 | 0.7 | 0.1 | 0.0 | 22.0 |
HST 2 | 24.7 | 13.3 | 0.6 | 0.3 | 0.1 | 0.0 | 25.6 |
HST 3 | 22.4 | 3.7 | 0.6 | 0.3 | 0.2 | 0.0 | 30.6 |
MSF 1_1 | 24.0 | 5.0 | 1.4 | 0.3 | 0.1 | 0.0 | NA |
MSF 1_2 | 43.2 | 28.9 | 0.1 | 0.7 | NA | 0.1 | NA |
MSF 1_3 | 15.8 | 4.1 | 2.7 | 0.0 | NA | 0.0 | NA |
OO 2_1 | 40.7 | 17.8 | NA | 1.2 | NA | NA | NA |
OO 2_2 | 37.2 | 31.3 | 1.9 | 0.0 | NA | NA | NA |
OO 2_3 | 31.8 | 5.3 | 0.4 | 0.0 | NA | NA | NA |
OO 3_1 | 17.8 | 5.8 | 2.3 | 0.0 | NA | 0.0 | NA |
OO 3_2 | 43.1 | 0.9 | 1.6 | 0.8 | NA | NA | NA |
OO 3_3 | 32.7 | 4.4 | 2.5 | 0.0 | 0.1 | 0.1 | NA |
PM 1_4 | 31.0 | 30.0 | 1.7 | 1.1 | 0.3 | 0.1 | NA |
PM 1_5 | 9.4 | 2.6 | 0.5 | 0.6 | 0.6 | 0.0 | NA |
PM 1_6 | 51.9 | 15.3 | 0.8 | 0.2 | 0.3 | 0.0 | NA |
STR 1 | 37.5 | 18.6 | 1.9 | 0.1 | NA | NA | 30.0 |
STR 2 | 22.4 | 14.9 | 2.8 | 0.4 | NA | 0.0 | 32.5 |
STR 3 | 21.7 | 2.5 | 3.6 | 1.0 | NA | 0.0 | 32.4 |
SWCL 2_4 | 6.3 | 2.0 | 8.0 | 1.2 | 0.0 | 0.3 | 21.0 |
SWCL 2_5 | 15.7 | 6.2 | 4.9 | 0.4 | 0.1 | 0.3 | 29.0 |
SWCL 2_6 | 16.8 | 8.3 | 2.6 | 1.6 | 0.1 | 0.2 | NA |
SWCN 1 | 20.1 | 9.2 | 0.2 | 0.2 | 0.0 | 0.1 | 38.1 |
SWCN 2 | 26.0 | 6.2 | 0.2 | 0.1 | NA | 0.1 | NA |
SWCN 3 | 17.4 | 6.7 | 0.3 | 0.0 | 0.0 | 0.7 | 36.6 |
SYM 1_1 | 23.4 | 5.3 | 0.9 | 0.1 | NA | 0.1 | NA |
SYM 1_2 | 16.8 | 11.1 | 0.9 | 0.3 | NA | 0.1 | NA |
SYM 1_3 | 32.9 | 10.3 | 1.1 | 0.4 | NA | 0.1 | NA |
Mean | 24.5 | 9.5 | 2.07 | 0.4 | 0.2 | 0.4 | 35.4 |
Appendix 4
Woody seedling data (2012–2014). Measurements collected in northwestern, central, and southwestern Ohio, USA, within 288 micro-plots (4 m2) nested within 72 plots (400 m2) and 24 sites. Seedling abundance was based on initial sample (2012) of seedlings 20–100 cm tall, survial was based on percentage of seedling survival between initial sample and final sample (2014), and recruitment was based on seedlings added to the cohort during 2013 and 2014. Species richness was calculated with all species recorded within all micro-plots per plot during the sampling period (2012–2014). Seedling categories include introduced (not native to North America), shade-intolerant natives, and shade-tolerant natives based upon shade tolerance values of 1–2.99 and 3–5, respectively (Niinemets and Valladares 2006)
Plot | Introduced abundance | Introduced survival | Introduced recruitment | Shade-tolerant abundance | Shade-tolerant survial % | Shade-tolerant recruitment | Shade-intolerant abundance | Shade-intolerant survival % | Shade-intolerant recruitment | Species richness |
CCG1 | 9 | 100 | 0 | 1 | 100 | 0 | 0 | 0 | 0 | 2 |
CCG2 | 4 | 100 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
CCG3 | 5 | 100 | 3 | 6 | 83.33 | 2 | 2 | 50 | 0 | 6 |
CCSP1_1 | 6 | 100 | 1 | 2 | 50 | 2 | 3 | 66.67 | 1 | 6 |
CCSP1_2 | 0 | 0 | 4 | 4 | 100 | 7 | 2 | 100 | 6 | 11 |
CCSP1_3 | 7 | 85.71 | 0 | 8 | 62.5 | 6 | 13 | 76.92 | 9 | 10 |
CCSP4_8 | 0 | 0 | 1 | 39 | 87.18 | 17 | 7 | 85.71 | 3 | 9 |
CCSP4_9 | 1 | 100 | 0 | 21 | 80.95 | 17 | 4 | 75 | 13 | 10 |
CCSP4_9B | 2 | 100 | 3 | 56 | 62.5 | 29 | 8 | 87.5 | 5 | 13 |
CCSP5_10 | 10 | 100 | 5 | 14 | 85.71 | 5 | 5 | 40 | 1 | 11 |
CCSP5_11 | 3 | 100 | 4 | 4 | 100 | 2 | 0 | 0 | 0 | 6 |
CCSP5_12 | 6 | 100 | 3 | 17 | 64.71 | 6 | 2 | 50 | 1 | 10 |
CLB_1 | 0 | 0 | 0 | 19 | 84.21 | 11 | 1 | 0 | 4 | 4 |
CLB_2 | 0 | 0 | 0 | 27 | 70.37 | 11 | 0 | 0 | 1 | 2 |
CLB_3 | 5 | 60 | 1 | 22 | 90.91 | 13 | 0 | 0 | 5 | 4 |
CLF_1 | 1 | 0 | 0 | 5 | 100 | 0 | 2 | 50 | 3 | 6 |
CLF_2 | 2 | 50 | 5 | 2 | 50 | 2 | 18 | 66.67 | 8 | 4 |
CLF_3 | 0 | 0 | 3 | 8 | 87.5 | 5 | 14 | 85.71 | 8 | 11 |
DMP_1 | 14 | 85.71 | 12 | 27 | 77.78 | 24 | 4 | 100 | 6 | 8 |
DMP_2 | 17 | 88.24 | 10 | 12 | 66.67 | 3 | 1 | 100 | 2 | 7 |
DMP_3 | 17 | 88.24 | 2 | 4 | 25 | 1 | 4 | 100 | 2 | 8 |
EDW1 | 5 | 80 | 3 | 4 | 25 | 4 | 0 | 0 | 0 | 4 |
EDW2 | 6 | 100 | 8 | 26 | 53.85 | 20 | 1 | 100 | 3 | 10 |
EDW3 | 2 | 100 | 1 | 67 | 53.73 | 58 | 0 | 0 | 3 | 8 |
ENG_1 | 4 | 100 | 4 | 4 | 75 | 3 | 0 | 0 | 1 | 7 |
ENG_2 | 2 | 100 | 3 | 12 | 100 | 5 | 5 | 80 | 6 | 10 |
ENG_3 | 3 | 100 | 4 | 12 | 91.67 | 9 | 2 | 100 | 8 | 12 |
FT1_1 | 2 | 100 | 0 | 0 | 0 | 0 | 1 | 100 | 0 | 2 |
FT1_2 | 4 | 100 | 0 | 2 | 100 | 3 | 1 | 100 | 3 | 7 |
FT1_3 | 3 | 33.33 | 0 | 0 | 0 | 2 | 6 | 83.33 | 6 | 6 |
FT2_1 | 0 | 0 | 3 | 8 | 75 | 9 | 61 | 91.8 | 13 | 8 |
FT2_2 | 0 | 0 | 0 | 0 | 0 | 0 | 9 | 100 | 7 | 1 |
FT2_3 | 0 | 0 | 0 | 5 | 60 | 1 | 26 | 84.62 | 9 | 5 |
GHN_1 | 0 | 0 | 0 | 0 | 0 | 0 | 12 | 66.67 | 14 | 2 |
GHN_2 | 4 | 100 | 1 | 2 | 100 | 1 | 1 | 0 | 4 | 8 |
GHN_3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
GLN1 | 5 | 100 | 4 | 6 | 33.33 | 9 | 1 | 100 | 0 | 7 |
GLN2 | 5 | 100 | 3 | 25 | 76 | 16 | 15 | 60 | 9 | 9 |
GLN3 | 1 | 100 | 2 | 45 | 77.78 | 38 | 8 | 87.5 | 5 | 11 |
GRM_1 | 4 | 100 | 0 | 13 | 92.31 | 7 | 11 | 72.73 | 1 | 12 |
GRM_2 | 10 | 100 | 3 | 9 | 100 | 3 | 5 | 100 | 1 | 10 |
GRM_3 | 10 | 100 | 1 | 22 | 95.45 | 7 | 6 | 100 | 5 | 7 |
HGH_1 | 13 | 92.31 | 3 | 23 | 95.65 | 9 | 25 | 80 | 9 | 18 |
HGH_2 | 19 | 100 | 4 | 20 | 75 | 10 | 15 | 86.67 | 5 | 19 |
HGH_3 | 8 | 100 | 0 | 19 | 94.74 | 11 | 9 | 100 | 13 | 17 |
HST1 | 0 | 0 | 0 | 9 | 88.89 | 11 | 11 | 18.18 | 1 | 7 |
HST2 | 4 | 100 | 0 | 28 | 71.43 | 14 | 2 | 50 | 3 | 9 |
HST3 | 0 | 0 | 0 | 24 | 87.5 | 34 | 15 | 46.67 | 5 | 9 |
MSF1_1 | 2 | 100 | 0 | 14 | 78.57 | 1 | 19 | 89.47 | 9 | 7 |
MSF1_2 | 0 | 0 | 0 | 4 | 75 | 1 | 27 | 88.89 | 14 | 6 |
MSF1_3 | 0 | 0 | 0 | 1 | 100 | 2 | 2 | 50 | 3 | 4 |
OO2_1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 |
OO2_2 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 100 | 0 | 1 |
OO2_3 | 0 | 0 | 0 | 5 | 20 | 2 | 4 | 25 | 1 | 4 |
OO3_2 | 0 | 0 | 0 | 5 | 100 | 5 | 1 | 100 | 1 | 3 |
OO3_3 | 0 | 0 | 1 | 7 | 71.43 | 2 | 0 | 0 | 0 | 3 |
PM1_4 | 2 | 100 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 3 |
PM1_5 | 3 | 100 | 2 | 19 | 73.68 | 3 | 1 | 100 | 3 | 11 |
PM1_6 | 0 | 0 | 0 | 3 | 100 | 1 | 1 | 100 | 0 | 2 |
STR_1 | 0 | 0 | 0 | 23 | 86.96 | 10 | 5 | 80 | 1 | 9 |
STR_2 | 1 | 100 | 2 | 99 | 78.79 | 80 | 2 | 100 | 22 | 11 |
STR_3 | 2 | 100 | 1 | 48 | 70.83 | 17 | 7 | 71.43 | 9 | 9 |
SWCL2_4 | 17 | 94.12 | 6 | 7 | 71.43 | 10 | 7 | 85.71 | 17 | 11 |
SWCL2_5 | 41 | 78.05 | 6 | 12 | 66.67 | 2 | 7 | 100 | 10 | 10 |
SWCL2_6 | 5 | 100 | 2 | 17 | 88.24 | 3 | 33 | 84.85 | 18 | 11 |
SWCN1 | 6 | 100 | 6 | 54 | 77.78 | 12 | 3 | 66.67 | 1 | 7 |
SWCN2 | 14 | 92.86 | 11 | 31 | 77.42 | 9 | 1 | 100 | 1 | 10 |
SWCN3 | 13 | 92.31 | 5 | 49 | 77.55 | 23 | 5 | 100 | 3 | 11 |
SYM1_1 | 3 | 66.67 | 6 | 4 | 50 | 6 | 0 | 0 | 8 | 9 |
SYM1_2 | 11 | 81.82 | 7 | 1 | 0 | 5 | 3 | 66.67 | 12 | 8 |
SYM1_3 | 5 | 80 | 9 | 1 | 100 | 3 | 2 | 50 | 14 | 10 |
Mean | 5 | 64.08 | 2 | 15 | 66.42 | 9 | 7 | 63.39 | 5 | 8 |
Appendix 5
All shrub species encountered in subplots in 24 sites located in northwestern, central, and southwestern Ohio, USA (2012–2014). Species identity and classification, i.e., is the shrub native or introduced to North America based on the USDA PLANTS database (https://plants.usda.gov)
Species | Classification |
Amelanchier sp. | Native |
Berberis thunbergii | Introduced |
Cephalanthus occidentalis | Native |
Cornus sp. | Native |
Elaeagnus umbellata | Introduced |
Euonymus alatus | Introduced |
Euonymus atropurpureus | Native |
Hamamelis virginiana | Native |
Hydrangea arborescens | Native |
Ligustrum vulgare | Introduced |
Lindera benzoin | Native |
Lonicera maackii | Introduced |
Lonicera morrowii | Introduced |
Lonicera tatarica | Introduced |
Rhamnus cathartica | Introduced |
Rosa multiflora | Introduced |
Spirea sp. | Native |
Staphylea trifolia | Native |
Viburnum acerifolium | Native |
Viburnum dentatum | Native |
Viburnum prunifolium | Native |
Zanthoxylum americanum | Native |
Appendix 6
All seedling species encountered in micro-plots in 24 sites located in northwestern, central, and southwestern Ohio, USA (2012–2014). Species identity, shade tolerance score (Niinemets and Valladares 2006), and analysis group are included for each species. Introduced refers to species not native to North America based on the USDA PLANTS database (https://plants.usda.gov)
Species | Shade tolerance ranking | Group |
Acer negundo | 3.47 | Shade-tolerant natives |
Acer nigrum | 3.00 | Shade-tolerant natives |
Acer rubrum | 3.44 | Shade-tolerant natives |
Acer saccharinum | 3.60 | Shade-tolerant natives |
Acer saccharum | 4.76 | Shade-tolerant natives |
Aesculus sp. | 3.81 | Shade-tolerant natives |
Asimina triloba | 3.95 | Shade-tolerant natives |
Berberis thunbergii | 1.50 | Introduced |
Carpinus caroliniana | 4.58 | Shade-tolerant natives |
Carya cordiformis | 2.07 | Shade-intolerant natives |
Carya laciniosa | 4.42 | Shade-tolerant natives |
Carya ovata | 3.40 | Shade-tolerant natives |
Carya sp. | 2.96 | Shade-intolerant natives |
Celtis occidentalis | 3.17 | Shade-tolerant natives |
Cercis canadensis | 3.00 | Shade-tolerant natives |
Cornus amomum | 3.00 | Shade-tolerant natives |
Cornus florida | 4.87 | Shade-tolerant natives |
Cornus sp. | 3.75 | Shade-tolerant natives |
Crataegus sp. | 2.13 | Shade-intolerant natives |
Elaeagnus umbellata | 1.35 | Introduced |
Euonymus alatus | 4.33 | Introduced |
Euonymus atropurpureus | 4.00 | Shade-tolerant natives |
Fagus grandifolia | 4.75 | Shade-tolerant natives |
Fraxinus quadrangulata | 1.84 | Shade-intolerant natives |
Fraxinus spp. | 2.67 | Shade-intolerant natives |
Ligustrum vulgare | 2.57 | Introduced |
Lindera benzoin | 3.00 | Shade-tolerant natives |
Liriodendron tulipifera | 2.07 | Shade-intolerant natives |
Lonicera maackii | 3.57 | Introduced |
Lonicera morrowii | 3.57 | Introduced |
Lonicera tatarica | 3.57 | Introduced |
Morus alba | 1.35 | Introduced |
Morus rubra | 2.34 | Shade-intolerant natives |
Ostrya virginiana | 4.58 | Shade-tolerant natives |
Prunus serotina | 2.46 | Shade-intolerant natives |
Pyrus calleryana | 1.35 | Introduced |
Quercus alba | 2.85 | Shade-intolerant natives |
Quercus bicolor | 2.98 | Shade-intolerant natives |
Quercus macrocarpa | 2.71 | Shade-intolerant natives |
Quercus muehlenbergii | 2.22 | Shade-intolerant natives |
Quercus palustris | 2.49 | Shade-intolerant natives |
Quercus rubra | 2.75 | Shade-intolerant natives |
Quercus sp. | 2.52 | Shade-intolerant natives |
Rhamnus cathartica | 1.93 | Shade-intolerant natives |
Rhamnus frangula | 1.93 | Shade-intolerant natives |
Rhamnus sp. | 1.93 | Introduced |
Ribes sp. | 3.36 | Shade-tolerant natives |
Rosa multiflora | 1.73 | Introduced |
Sambucus sp. | 1.35 | Shade-intolerant natives |
Sassafras albidum | 1.68 | Shade-intolerant natives |
Staphylea trifolia | 1.93 | Shade-intolerant natives |
Tilia americana | 3.98 | Shade-tolerant natives |
Ulmus sp. | 3.22 | Shade-tolerant natives |
Viburnum acerifolium | 4.00 | Shade-tolerant natives |
Viburnum dentatum | 4.00 | Shade-tolerant natives |
Viburnum prunifolium | 4.00 | Shade-tolerant natives |
Zanthoxylum americanum | 2.50 | Shade-intolerant natives |
Appendix 7
Model comparison results for models testing the effects of four different measures of EAB-caused ash decline on the responses of canopy openness, maple (Acer spp.) stand tree basal area (BA) relative growth rate (RGR), and non-ash stand tree BA RGR. Models in italics are the most parsimonious models (ΔAIC = 0) and any competing models with ΔAIC ≤ 2 indicating substantial support (Burnham and Anderson 2002). AICc is the small sample Akaike’s information criterion; ∆AIC is the difference between the best model and every other evaluated model; and w is Akaike’s weight. Predictor variable abbreviations are as defined in Appendix 1. NA indicates that those models were not evaluated for the specified response variable. P-values reported are from likelihood ratio tests.
ADI | AMI | 1st Exit | Topography | ADI + topography | AMI + topography | Null model | |
Canopy openness 2012 | |||||||
X2 | 0.39 | 2.44 | 2.52 | NA | NA | NA | NA |
p | 0.53 | 0.12 | NA | NA | NA | NA | |
Non-ash stand RGR | |||||||
AICc | − 240.25 | − 238.70 | − 193.36 | NA | NA | NA | − 237.73 |
ΔAICc | 0.00 | 1.54 | 46.89 | NA | NA | NA | 2.52 |
w | 0.57 | 0.26 | 0.00 | NA | NA | NA | 0.16 |
Non-ash stand RGR (with topography) | |||||||
AICc | − 240.25 | − 238.70 | − 234.36 | − 240.99 | − 241.86 | − 240.10 | − 237.73 |
ΔAICc | 1.61 | 3.15 | 7.50 | 0.87 | 0.00 | 1.76 | 4.13 |
w | 0.16 | 0.07 | 0.01 | 0.23 | 0.35 | 0.14 | 0.04 |
Maple stand RGR | |||||||
AICc | − 187.01 | − 188.81 | − 149.42 | NA | NA | NA | − 185.45 |
ΔAICc | 1.80 | 0.00 | 39.38 | NA | NA | NA | 3.35 |
w | 0.26 | 0.63 | 0.00 | NA | NA | NA | 0.12 |
Maple stand RGR (with topography) | |||||||
AICc | − 187.01 | − 188.81 | − 186.33 | − 184.30 | − 185.17 | − 186.62 | − 185.45 |
ΔAICc | 1.80 | 0.00 | 2.48 | 4.51 | 3.64 | 2.19 | 3.35 |
w | 0.16 | 0.40 | 0.12 | 0.04 | 0.07 | 0.13 | 0.08 |
Appendix 8
Model comparison results for models testing the effect of EAB-caused ash decline (ADI and AMI), non-ash stand BA, and subcanopy BA on subcanopy, native and introduced shrub RGR responses (2012–2014). Most parsimonious and competing models are presented in italics as in Appendix 4. Analyses were conducted using AICc model selection; response variable rows and abbreviations are as per Appendix 4
ADI | AMI | Non-ash stand | ADI + non-ash stand | AMI + non-ash stand | Subcanopy | ADI + subcanopy | AMI + subcanopy | Null model | |
Subcanopy RGR | |||||||||
AICc | − 159.45 | − 159.41 | − 159.71 | − 158.00 | − 157.84 | NA | NA | NA | − 161.29 |
ΔAICc | 1.84 | 1.89 | 1.58 | 3.29 | 3.42 | NA | NA | NA | 0.00 |
w | 0.15 | 0.15 | 0.17 | 0.07 | 0.07 | NA | NA | NA | 0.38 |
Maple subcanopy RGR | |||||||||
AICc | − 136.78 | − 136.85 | − 136.48 | − 134.56 | − 134.61 | NA | NA | NA | − 138.68 |
ΔAICc | 1.90 | 1.83 | 2.20 | 4.12 | 4.07 | NA | NA | NA | 0.00 |
w | 0.16 | 0.17 | 0.14 | 0.05 | 0.06 | NA | NA | NA | 0.42 |
Native shrub RGR | |||||||||
AICc | 7.36 | 7.28 | 7.11 | 9.61 | 9.56 | 8.36 | 10.88 | 10.78 | 4.98 |
ΔAICc | 2.38 | 2.30 | 2.13 | 4.63 | 4.58 | 3.38 | 5.91 | 5.80 | 0.00 |
w | 0.12 | 0.13 | 0.14 | 0.04 | 0.04 | 0.08 | 0.02 | 0.02 | 0.41 |
Introduced shrub RGR | |||||||||
AICc | − 7.49 | − 5.39 | − 5.39 | − 5.91 | − 3.06 | − 5.36 | − 4.88 | − 2.98 | − 7.66 |
ΔAICc | 0.17 | 2.27 | 2.27 | 1.75 | 4.59 | 2.30 | 2.77 | 4.68 | 0.00 |
w | 0.25 | 0.09 | 0.09 | 0.11 | 0.03 | 0.08 | 0.07 | 0.03 | 0.27 |
Appendix 9
Model comparison results for models testing the effect of EAB-caused ash decline (ADI and AMI), non-ash stand BA, subcanopy BA, and shrub cover on sugar maple (Acer saccharum) seedling height RGR responses (2012–2014). Most parsimonious and competing models are presented in italics as in Appendix 4. Analyses were conducted using AICc model selection; response variable rows and abbreviations are as per Appendix 4
ADI | AMI | Non-ash stand | Subcanopy | Shrub cover | ADI + non-ash stand | AMI + non-ash stand | ADI + subcanopy | AMI + subcanopy | ADI + shrub cover | AMI + shrub cover | Null model | |
Sugar maple seedling RGR | ||||||||||||
AICc | 34.08 | 37.51 | 38.19 | 35.01 | 38.17 | 37.23 | 40.65 | 30.82 | 36.63 | 37.20 | 40.65 | 35.39 |
ΔAICc | 3.27 | 6.70 | 7.38 | 4.19 | 7.36 | 6.42 | 9.83 | 0.00 | 5.82 | 6.39 | 9.83 | 4.57 |
w | 0.12 | 0.02 | 0.02 | 0.07 | 0.02 | 0.02 | 0.00 | 0.60 | 0.03 | 0.02 | 0.00 | 0.06 |
Appendix 10
Model comparison results for models testing the effect of EAB-caused ash decline, non-ash stand BA, subcanopy BA, and shrub layer cover on woody seedling species richness (2012-2014) of all seedlings and abundance of three subsets of seedlings. Most parsimonious and competing models are presented in italics as in Appendix 4. NA = model not included in an analysis. Analyses were conducted using AICc model selection; response variable rows and abbreviations are as per Appendix 4
ADI | AMI | Non-ash stand | Subcanopy | Shrub cover | ADI + non-ash stand | AMI + non-ash stand | ADI + subcanopy | AMI + subcanopy | ADI + shrub cover | AMI + shrub cover | ADI + topography | AMI + topography | Shrub cover + topography | ADI + shrub cover + topography | AMI + shrub cover + topography | Null model | |
Species richness | |||||||||||||||||
AICc | 387.78 | 387.90 | 387.66 | 387.92 | 387.81 | 389.87 | 389.87 | 389.84 | 390.00 | 389.80 | 389.92 | NA | NA | NA | NA | NA | 385.84 |
ΔAICc | 1.94 | 2.06 | 1.82 | 2.08 | 1.96 | 4.03 | 4.03 | 4.00 | 4.16 | 0.04 | 4.08 | NA | NA | NA | NA | NA | 0.00 |
w | 0.10 | 0.10 | 0.11 | 0.10 | 0.10 | 0.04 | 0.04 | 0.04 | 0.03 | 0.82 | 0.04 | NA | NA | NA | NA | NA | 0.27 |
Abundance: shade-tolerant native seedlings | |||||||||||||||||
AICc | 528.15 | 528.15 | 526.57 | 526.55 | 527.85 | 528.82 | 528.62 | 528.73 | 528.86 | 530.16 | 530.16 | NA | NA | NA | NA | NA | 525.92 |
ΔAICc | 2.23 | 2.23 | 0.65 | 0.64 | 1.94 | 2.90 | 2.70 | 2.81 | 2.94 | 4.24 | 4.24 | NA | NA | NA | NA | NA | 0.00 |
w | 0.07 | 0.07 | 0.15 | 0.16 | 0.08 | 0.05 | 0.06 | 0.05 | 0.05 | 0.03 | 0.03 | NA | NA | NA | NA | NA | 0.21 |
Abundance: shade-intolerant native seedlings | |||||||||||||||||
AICc | 412.02 | 411.77 | 411.51 | 409.89 | 0.28 | 413.64 | 413.45 | 412.11 | 411.93 | 413.39 | 412.95 | NA | NA | NA | NA | NA | 410.16 |
ΔAICc | 2.14 | 1.88 | 1.63 | 0.00 | 0.18 | 3.76 | 3.56 | 2.22 | 2.04 | 3.50 | 3.06 | NA | NA | NA | NA | NA | 0.28 |
w | 0.07 | 0.08 | 0.09 | 0.20 | 0.09 | 0.03 | 0.03 | 0.07 | 0.07 | 0.04 | 0.04 | NA | NA | NA | NA | NA | 0.18 |
Abundance: introduced seedlings | |||||||||||||||||
AICc | 349.86 | 350.68 | 346.58 | 339.48 | 335.38 | 348.08 | 352.28 | 340.96 | 342.79 | 333.72 | 334.58 | NA | NA | NA | NA | NA | 347.44 |
ΔAICc | 16.14 | 16.95 | 12.85 | 5.75 | 1.66 | 14.35 | 18.55 | 7.24 | 9.07 | 0.00 | 0.85 | NA | NA | NA | NA | NA | 13.72 |
w | 0.00 | 0.00 | 0.00 | 0.03 | 0.20 | 0.00 | 0.00 | 0.01 | 0.00 | 0.46 | 0.30 | NA | NA | NA | NA | NA | 0.00 |
Abundance: introduced seedlings (with topography) | |||||||||||||||||
AICc | 362.56 | 363.18 | 361.44 | NA | 346.88 | 363.41 | 363.72 | NA | NA | 347.65 | 348.54 | 352.14 | 352.18 | 341.59 | 341.28 | 341.69 | 361.56 |
ΔAICc | 21.28 | 21.90 | 20.15 | NA | 5.59 | 22.12 | 22.43 | NA | NA | 6.36 | 7.26 | 10.86 | 10.89 | 0.31 | 0.00 | 0.41 | 20.27 |
w | 0.00 | 0.00 | 0.00 | NA | 0.02 | 0.00 | 0.00 | NA | NA | 0.01 | 0.01 | 0.00 | 0.00 | 0.30 | 0.36 | 0.29 | 0.00 |
Appendix 11
Model comparison results for models testing the effect of EAB-caused ash decline, non-ash stand BA, subcanopy BA, and shrub layer cover on woody seedling recruitment (2013 and 2014) of three subsets of seedlings. Most parsimonious and competing models are presented in italics as in Appendix 4. Analyses were conducted using AICc model selection; response variable rows and abbreviations are as per Appendix 4
ADI | AMI | Non-ash stand | Subcanopy | Shrub cover | ADI + non-ash stand | AMI + non-ash stand | ADI + subcanopy | AMI + subcanopy | ADI + shrub cover | AMI + shrub cover | Null model | |
Recruitment: shade-tolerant native seedlings | ||||||||||||
AICc | 449.28 | 448.02 | 447.91 | 449.43 | 448.92 | 449.63 | 447.61 | 451.55 | 450.21 | 451.10 | 449.74 | 447.20 |
ΔAICc | 2.08 | 0.82 | 0.71 | 2.23 | 1.73 | 2.44 | 0.41 | 4.35 | 3.01 | 3.90 | 2.55 | 0.00 |
w | 0.07 | 0.12 | 0.13 | 0.06 | 0.08 | 0.06 | 0.15 | 0.02 | 0.04 | 0.03 | 0.05 | 0.19 |
Recruitment: shade-intolerant native seedlings | ||||||||||||
AICc | 389.37 | 389.29 | 389.38 | 389.27 | 388.05 | 391.67 | 391.60 | 391.58 | 391.44 | 390.36 | 390.17 | 387.13 |
ΔAICc | 2.23 | 2.15 | 2.24 | 2.13 | 0.92 | 4.54 | 4.46 | 4.44 | 4.31 | 3.23 | 3.04 | 0.00 |
w | 0.09 | 0.09 | 0.09 | 0.09 | 0.17 | 0.03 | 0.03 | 0.03 | 0.03 | 0.05 | 0.06 | 0.26 |
Recruitment: introduced seedlings | ||||||||||||
AICc | 285.20 | 285.22 | 282.81 | 278.81 | 282.64 | 285.09 | 285.05 | 281.01 | 281.04 | 284.92 | 284.94 | 283.03 |
ΔAICc | 6.39 | 6.42 | 4.01 | 0.00 | 3.83 | 6.28 | 6.24 | 2.20 | 2.24 | 6.11 | 6.13 | 4.23 |
w | 0.02 | 0.02 | 0.06 | 0.43 | 0.06 | 0.02 | 0.02 | 0.14 | 0.14 | 0.02 | 0.02 | 0.05 |
Appendix 12
Model comparison results for models testing the effect of EAB-caused ash decline, non-ash stand BA, subcanopy BA, and shrub layer percent cover on woody seedling survival (2012–2014) of three subsets of seedlings. Most parsimonious and competing models are presented in italics as in Appendix 4. Analyses were conducted using AICc model selection; response variable rows and abbreviations are as per Appendix 4
ADI | AMI | Non-ash stand | Subcanopy | Shrub cover | ADI + non-ash stand | AMI + Non- ash stand | ADI + subcanopy | AMI + subcanopy | ADI + shrub cover | AMI + shrub cover | Null model | |
Survival: shade-tolerant native seedlings | ||||||||||||
AICc | 235.66 | 235.55 | 235.66 | 227.64 | 235.77 | 237.88 | 237.78 | 229.87 | 229.81 | 237.95 | 237.82 | 233.56 |
ΔAICc | 8.02 | 7.90 | 8.02 | 0.00 | 8.13 | 10.23 | 10.13 | 2.23 | 2.17 | 10.30 | 10.18 | 5.92 |
w | 0.01 | 0.01 | 0.01 | 0.55 | 0.01 | 0.00 | 0.00 | 0.18 | 0.19 | 0.00 | 0.00 | 0.03 |
Survival: shade-intolerant native seedlings | ||||||||||||
AICc | 174.45 | 173.29 | 174.50 | 174.49 | 174.42 | 176.69 | 175.52 | 176.68 | 175.54 | 176.62 | 175.54 | 172.33 |
ΔAICc | 2.12 | 0.97 | 2.18 | 2.16 | 2.09 | 4.36 | 3.19 | 4.35 | 3.21 | 4.30 | 3.21 | 0.00 |
w | 0.09 | 0.16 | 0.09 | 0.09 | 0.09 | 0.03 | 0.05 | 0.03 | 0.05 | 0.03 | 0.05 | 0.25 |
Survival: introduced seedlings | ||||||||||||
AICc | 146.74 | 146.75 | 146.72 | 145.35 | 146.75 | 149.09 | 149.08 | 147.72 | 147.73 | 149.11 | 149.11 | 144.48 |
ΔAICc | 2.26 | 2.27 | 2.24 | 0.87 | 2.27 | 4.61 | 4.60 | 3.24 | 3.25 | 4.63 | 4.63 | 0.00 |
w | 0.09 | 0.09 | 0.09 | 0.17 | 0.09 | 0.03 | 0.03 | 0.05 | 0.05 | 0.03 | 0.03 | 0.27 |
Rights and permissions
About this article

Cite this article
Hoven, B.M., Knight, K.S., Peters, V.E. et al. Release and suppression: forest layer responses to emerald ash borer (Agrilus planipennis)-caused ash death. Annals of Forest Science 77, 10 (2020). https://doi.org/10.1007/s13595-019-0895-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13595-019-0895-y