
ABSTRACT
NEDDylation has been shown to participate in the DNA damage pathway, but the substrates of neural precursor cell expressed developmentally downregulated 8 (NEDD8) and the roles of NEDDylation involved in the DNA damage response (DDR) are largely unknown. Translesion synthesis (TLS) is a damage-tolerance mechanism, in which RAD18/RAD6-mediated monoubiquitinated proliferating cell nuclear antigen (PCNA) promotes recruitment of polymerase η (polη) to bypass lesions. Here we identify PCNA as a substrate of NEDD8, and show that E3 ligase RAD18-catalyzed PCNA NEDDylation antagonizes its ubiquitination. In addition, NEDP1 acts as the deNEDDylase of PCNA, and NEDP1 deletion enhances PCNA NEDDylation but reduces its ubiquitination. In response to H2O2 stimulation, NEDP1 disassociates from PCNA and RAD18-dependent PCNA NEDDylation increases markedly after its ubiquitination. Impairment of NEDDylation by Ubc12 knockout enhances PCNA ubiquitination and promotes PCNA-polη interaction, while up-regulation of NEDDylation by NEDD8 overexpression or NEDP1 deletion reduces the excessive accumulation of ubiquitinated PCNA, thus inhibits PCNA-polη interaction and blocks polη foci formation. Moreover, Ubc12 knockout decreases cell sensitivity to H2O2-induced oxidative stress, but NEDP1 deletion aggravates this sensitivity. Collectively, our study elucidates the important role of NEDDylation in the DDR as a modulator of PCNA monoubiquitination and polη recruitment.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
INTRODUCTION
Genome integrity is a basic requirement for cell growth, proliferation, and development. Under different exogenous or endogenous stresses, such as those caused by ionizing radiation, ultraviolet (UV) radiation, DNA-damaging chemical agents, and other oxidative chemicals, genome integrity and cell viability are greatly threatened (Jackson and Durocher, 2013). In response to these stresses, various signaling pathways involved in multiple repair mechanisms are induced, including the DNA damage response (DDR) that recruits a series of proteins to damaged DNA sites and triggers checkpoint signaling or essential repair steps (Hoeijmakers, 2001; Harrison and Haber, 2006; Branzei and Foiani, 2008; Bergink and Jentsch, 2009).
Recent studies have reported that histones H4 and H2A are substrates of NEDD8, and histone NEDDylation is involved in the DDR pathway (Ma et al., 2013; Li et al., 2014; Brown et al., 2015). NEDDylation is reported to regulate the decision between the non-homologous end joining (NHEJ) pathway and different homologous recombination (HR) subpathways by controlling CtBP interacting protein (CtIP)-mediated DNA resection (Jimeno et al., 2015). NEDD8 has the highest similarity with ubiquitin among all the ubiquitin-like proteins (Kamitani et al., 1997), and NEDDylation occurs via a similar process with different enzymatic ligases conjugating NEDD8 to the lysines of substrates (Rabut and Peter, 2008; Hochstrasser, 2009; Huang et al., 2009a; Wei et al., 2012; Zhou et al., 2016). The proteins of the Cullin family, p53, p73, BCA3, TβRII, E2F-1, ML3, AXR1 and ribosomal proteins have been identified as NEDD8 substrates (Ohh et al., 2002; Xirodimas et al., 2004; Gao et al., 2006; Watson et al., 2006; Xirodimas et al., 2008; Loftus et al., 2012; Mahata et al., 2012; Aoki et al., 2013; Hakenjos et al., 2013; Zuo et al., 2013; Zhang et al., 2014; Enchev et al., 2015; Lan et al., 2016; Mergner et al., 2017). However, compared to those of ubiquitination, the substrates of NEDD8 remain largely unknown, especially those in the DDR pathway.
Mounting evidence indicates that the ubiquitin system plays a critical role in major DDR pathways (Kirkin and Dikic, 2007; Jackson and Durocher, 2013), and one of the classic DDR pathways controlled by ubiquitination is dependent on PCNA (Hoege et al., 2002; Moldovan et al., 2007; Bergink and Jentsch, 2009). PCNA can form a homotrimeric ring-shaped structure that encircles DNA and acts as a clamp to assist DNA polymerase in mediating DNA replication (Moldovan et al., 2007). Strikingly, PCNA can be modified by ubiquitin on the conserved lysine residue (K164) and induce post-replication repair (PRR) when DNA is damaged by UV irradiation, some chemical agents, or oxidative stress (Hoege et al., 2002; Zlatanou et al., 2011). PRR is a type of DNA repair mechanism that uses translesion synthesis (TLS) or the error-free template switching pathway to resolve the DNA lesions encountered by DNA polymerase (Bergink and Jentsch, 2009; Sale et al., 2012). When PCNA is monoubiquinated by RAD6B-RAD18 E3 ligase, it can promote the recruitment of TLS polymerases like polη and mediate DNA synthesis across the bulky lesions (Kannouche et al., 2004; Bienko et al., 2005). Because many TLS polymerases have an ubiquitin-binding domain and PCNA-interacting-protein box, they can bind to monoubiquitinated PCNA specifically and ensure TLS (Kannouche et al., 2001; Kannouche et al., 2004; Bienko et al., 2005). Besides, a recent study showed that PCNA monoubiquitination has no effects on the binding affinity of polη, raising a debate on the function of PCNA monoubiquitination (Hedglin et al., 2016). Moreover, other studies have shown that yeast PCNA is modified by small ubiquitin-like modifier (SUMO) on Lys164 and Lys127, and SUMOylated PCNA can recruit a helicase Srs2 to prevent undesired recombination (Pfander et al., 2005). The crosstalk between ubiquitin and SUMO indicates that these two modifiers can compete for the substrate (Papouli et al., 2005). Recent research has shown that PCNA is also modified by ISG15, and ISGylation of PCNA recruits USP10 to deubiquitinate PCNA, thereby regulating PCNA ubiquitination under UV irradiation-induced stress (Park et al., 2014). However, whether other ubiquitin-like modifiers are involved in the switchboard among different modifications of PCNA remains incompletely understood.
In this study, we report that PCNA is a substrate of NEDD8. RAD18 functions as the E3 ligase and NEDP1 acts as its deNEDDylase. In response to hydrogen peroxide (H2O2)-induced oxidative stress, NEDDylation plays a pivotal role in the recruitment of polη by regulating PCNA monoubiquitination.
RESULTS
Modification of PCNA by NEDD8
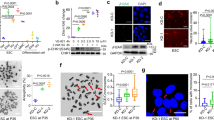
The mass spectrum data of NEDD8 substrates screening in the previous study showed that PCNA might be a substrate of NEDD8 (Xirodimas et al., 2008; Coleman et al., 2017). As PCNA is modified by ubiquitin, SUMO and ISG15 (Hoege et al., 2002; Park et al., 2014), we thus assessed whether PCNA was NEDDylated by NEDD8 using Ni2+ pull-down assay, in which His-tagged NEDD8 was expressed and NEDD8 conjugates were enriched by Ni2+ resin. As shown in Fig. 1A, we identified a specific band similar to PCNA ubiquitination that migrated slower than the unmodified PCNA, representing PCNA NEDDylation. By contrast, we did not observe modified PCNA in cells expressing a mutant NEDD8-ΔGG that lacked the last two-glycine residues (Figs. 1B and S1A). Moreover, we investigated whether NEDD8-activating enzyme inhibitor MLN4924 affected PCNA NEDDylation. As shown in Fig. 1B, MLN4924 abolished PCNA NEDDylation compared to that in untreated cells, indicating that the NEDD8 activating E1 is utilized for PCNA NEDDylation.

Modification of PCNA by NEDD8. (A) Immunoblotting analysis of PCNA NEDDylation in His-tagged NEDD8 conjugates or ubiquitin conjugates enriched by Ni2+ pull-down (Ni-PD). (B–D) Analysis of the effect of MLN4924 (B), PCNA-K164R (C), and NEDP1 (D) on PCNA NEDDylation. To obtain the results shown in (A–D), HEK293T cells were transfected with indicated plasmids, and then the whole cell lysates (input) and His-tagged NEDD8 conjugates or ubiquitin conjugates enriched by Ni-PD were subjected to SDS-PAGE and detected by Western blotting with the respective antibodies. (E) Analysis of the protein level of NEDP1 and NEDD8 in Nedp1 −/− HEK293T cells. Nedp1 −/− HEK293T cells were generated using CRISPR/CAS9 method. NEDP1 and NEDD8 were detected by Western blotting using the indicated antibodies. (F) Immunoblotting analysis of the effect of Nedp1 knockout on PCNA NEDDylation in His-tagged NEDD8 conjugates. HEK293T WT or Nedp1 −/− cells were transfected with indicated plasmids, and then His-tagged NEDD8 conjugates were enriched by Ni-PD and detected by Western blotting with the respective antibodies. (G) Immunoblotting analysis of endogenous PCNA NEDDylation in Nedp1 −/− HEK293T cells. Nedp1 −/− HEK293T cells were lysed in NP-40 lysis buffer and sonicated for 15 times, then immunoprecipitation was performed using anti-PCNA antibody
As the lysine 164 of PCNA is the modification site for both ubiquitin and SUMO (Bergink and Jentsch, 2009), to investigate whether NEDD8 also targets PCNA at Lys164, we constructed a mutant, PCNA-K164R, and examined its NEDDylation. No band corresponding to NEDDylated PCNA-K164R was detected (Fig. 1C), indicating that PCNA NEDDylation, ubiquitination, and SUMOylation occur on the same lysine residue.
Furthermore, we examined whether NEDP1 acted as the deNEDDylase of PCNA. As shown in Figs. 1D and S1B, NEDP1 significantly inhibited PCNA NEDDylation, but the mutant of NEDP1, NEDP1-C163S, in which the deNEDDylase activity was lost, could not deNEDDylate PCNA. Next, we constructed Nedp1 −/− cells and found that NEDD8-conjugated targets were enhanced significantly (Fig. 1E), explaining why only few conjugates were formed by endogenous NEDD8 (due to the presence of NEDP1). Compared to wild type (WT) cells, PCNA NEDDylation also increased significantly in Nedp1 −/− cells, which was eliminated by NEDP1 expression (Fig. 1F). These observations suggest that NEDP1 is the specific deNEDDylase for PCNA NEDDylation.
As both PCNA ubiquitination and NEDDylation could not be detected in normal cells, we performed immune precipitation (IP) to detect endogenous NEDDylation of PCNA in Nedp1 −/− cells. As shown in Fig. 1G, we succeeded to detect endogenous PCNA NEDDylation, which was abolished by MLN4924 treatment, confirming that PCNA is NEDDylated in vivo.
RAD18 serves as an E3 ligase of PCNA NEDDylation
It has been reported that some substrates of NEDD8 and ubiquitin share the same E3 ligase (Enchev et al., 2015), we therefore explored whether RAD18 also functions as the E3 ligase conjugating NEDD8 to PCNA. First, we detected the interaction between RAD18 and UBC12, an E2 of the NEDDylation pathway. Co-IP assay showed that both exogenous and endogenous RAD18 interacted with UBC12 (Figs. 2A and S1C). Next, we examined whether RAD18 promotes PCNA NEDDylation by Ni2+ pull-down assay. Indeed, RAD18 strongly enhanced PCNA NEDDylation (Fig. 2B). In addition, we constructed Ubc12 −/− HEK293T cell (Fig. S1D) and found that RAD18 could not promote PCNA NEDDylation without UBC12 (Fig. 2C). We further confirmed the effect of RAD18 on PCNA NEDDylation in vitro. As shown in Fig. 2D, UBC12/RAD18 but not RAD6B/RAD18 could NEDDylate PCNA, suggesting that RAD18 catalyzes PCNA NEDDylation using NEDD8 specific enzymes in vivo and vitro.

RAD18 acts as an E3 ligase of PCNA NEDDylation. (A) Analysis of the interaction between RAD18 and UBC12. HEK293T cells were lysed in NP-40 lysis buffer, and sonicated for 15 times, then immunoprecipitation was performed using antibody against RAD18. (B and C) Immunoblotting analysis of the effect of RAD18 on PCNA NEDDylation in His-tagged NEDD8 conjugates from HEK293T WT (B) or Ubc12 −/− cells (C). HEK293T WT cells or Ubc12 −/− cells were transfected with the corresponding plasmids, and PCNA NEDDylation or ubiquitination was analyzed by Ni-PD assay and Western blotting with indicated antibodies. (D) Analysis of PCNA NEDDylation by in vitro NEDDylation assay. GST-PCNA and His-SUMO-RAD18 were incubated with E1, UBC12 or RAD6B, and NEDD8 at 37°C for 1 h, and then the reactions were terminated by adding SDS buffer and detected by Western blotting with indicated antibody. (E) Immunoblotting analysis of PCNA NEDDylation in Rad18 −/− cells. HEK293T WT or Rad18 −/− cells were transfected with indicated plasmids, and PCNA NEDDylation was detected as described above. Fold change of PCNA modification intensity was analyzed using odyssey imaging systems (LI-COR, Lincoln, Nebraska USA)
The Cys28 in the RING domain of RAD18 is the key site essential for its E3 ligase activity (Huang et al., 2009b), we constructed two RAD18 mutants, C28F and ΔRING, and detected their effects on PCNA NEDDylation. The mutants diminished the activity of RAD18 in mediating PCNA NEDDylation (Fig. 2E, lines 1–4), demonstrating that Cys28 is essential for RAD18 to catalyze NEDDylation pathway. To investigate whether PCNA NEDDylation is dependent on RAD18, we constructed a Rad18-deficient stable HEK293T cell line using the Fast TALETM TALEN Assembly Kit and confirmed Rad18 knockout by Western blotting using an antibody against RAD18 (Fig. S1E). Then we examined the level of PCNA NEDDylation in Rad18 −/− cells. With Rad18 depletion, PCNA NEDDylation was abolished totally compared to that in WT cells, and only WT RAD18 but not the mutants could recover PCNA NEDDylation (Fig. 2E, lines 5–8). In summary, these data indicate that RAD18 acts as an E3 ligase of PCNA NEDDylation, which depends on UBC12.
NEDDylated PCNA accumulates under H2O2-induced oxidative stress
Previous research revealed that PCNA is monoubiquitinated under H2O2-induced oxidative stress, and monoubiquitinated PCNA helps recruiting polη to bypass DNA lesions (Zlatanou et al., 2011). We first detected whether NEDD8 accumulated at DNA damage sites after H2O2 treatment. As shown in Fig. 3A, NEDD8 formed foci in HeLa cells treated with H2O2, suggesting that NEDD8 participates in the H2O2-induced DDR pathway. To further assess whether PCNA NEDDylation also involves in the DDR pathway, we treated cells with 800 µmol/L H2O2 for various times and performed Ni2+ pull-down assay to examine PCNA NEDDylation. As shown in Fig. 3B, NEDDylation of PCNA occurred at 5 min, reached a peak at 15 min, decreased at 60 min, and finally disappeared after 90 min of H2O2 treatment. Meanwhile, in cells with PCNA-K164R, accumulation of NEDDylated PCNA did not occur after H2O2 treatment (Fig. 3C), indicating that the increased NEDDylation of PCNA is dependent on the conserved modification residue Lys164. We also observed an increase of endogenous PCNA NEDDylation in response to H2O2 treatment (Fig. 3D). In addition, NEDDylated PCNA also accumulated under DNA damage induced by UV irradiation (Fig. S2A and S2D). These data strongly suggest that PCNA NEDDylation is involved in the DDR pathway induced by various stimulations.

Hydrogen peroxide promotes PCNA NEDDylation. (A) Immunofluorescence analysis of NEDD8 localization after H2O2 treatment. HeLa cells were treated with or without 800 μmol/L H2O2 for 30 min, and then endogenous NEDD8 was examined by immunofluorescence staining using anti-NEDD8 antibody. The scale bar is 10 μm. (B) A time course analysis of PCNA NEDDylation in His-tagged NEDD8 conjugates enriched by Ni-PD in response to H2O2-induced oxidative stress. In (B), (C), and (E), HEK293T WT or Ubc12 −/− cells were transfected with the indicated plasmids for 36 h and harvested after treatment with 800 μmol/L H2O2 for the indicated time. The NEDD8 conjugates were enriched through Ni-PD and detected by Western blotting using anti-PCNA antibody. (C) Analysis of the effect of K164R on PCNA NEDDylation. (D) Analysis of endogenous PCNA NEDDylation in response to H2O2-induced oxidative stress. HEK293T cells treated with 800 μmol/L H2O2 for the indicated time were lysed in NP-40 lysis buffer and sonicated for 15 times. Immunoprecipitation was performed using anti-PCNA antibody. (E) Immunoblotting analysis of the effect of Ubc12 knockout on PCNA NEDDylation in His-tagged NEDD8 conjugates. (F) Analysis of the abundance of NEDP1 and PCNA modification in the Trixon-X100 insoluble fraction (TIF) in HEK293T WT or Nedp1 −/− cells under oxidative stress. HEK293T WT or Nedp1 −/− cells were treated with 800 μmol/L H2O2 for the indicated time. Then the TIF was isolated and subjected to Western blotting analysis with anti-PCNA and anti-NEDP1 antibodies. Fold change of PCNA modification intensity was analyzed using odyssey imaging systems. (G) Analysis of the interaction between PCNA and NEDP1 in response to H2O2 treatment. HEK293T cells were treated with H2O2 for indicated time, and then lysed in Triton-X100 lysis buffer and sonicated for 15 times. Immunoprecipitation was performed using antibody against PCNA. (H and I) Analysis of the effect of NEDP1 expression (H) or Nedp1 knockout (I) on accumulation of NEDDylated PCNA in response to H2O2 treatment. HEK293T WT or Nedp1 −/− cells were transfected with His-NEDD8, and then the cells were treated with or without 800 μmol/L H2O2 for 30 min before NEDDylated PCNA was enriched by Ni-PD and detected by Western blotting using anti-PCNA antibody
Previous study reported that oxidative stress causes a general increase in NEDDylation through ubiquitin E1 enzyme UBE1 but independent of the NEDD8 E1 enzyme (Leidecker et al., 2012), we questioned whether the increase in PCNA NEDDylation required the NEDD8 E1 APPBP1-UBA3. As shown in Fig. S2C, in MLN4924-treated 293T cells, we performed Ni2+ pull-down assay and observed a significant inhibition of MLN4924 on PCNA NEDDylation but not its ubiqitination, suggesting that PCNA NEDDylation utilizes the NEDD8 E1 under oxidative stress. In addition, UBC12-C111S, an activity-deficient mutant of UBC12 and a dominant blocker of the NEDDylation pathway by reducing the pool of free NEDD8, also impeded the increase in PCNA NEDDylation (Fig. S2B). Moreover, in Ubc12 −/− cells, we did not detect the increase of PCNA NEDDylation, while expression of UBC12 recovered the accumulation of NEDDylated PCNA (Fig. 3E).
Next, we investigated whether NEDP1 involved in the H2O2 treatment-induced DNA damage response. As the modified PCNA accumulated on chromatin during S-phase and became resistant to Triton-X100 extraction (Kannouche et al., 2004), we treated HEK293T cells with Triton-X100 and then detected whether NEDP1 affected PCNA modification. Consistent with the reduction of NEDP1 in the Triton-X100 insoluble fraction (TIF), we observed an obvious increase in modified PCNA (Fig. 3F, lanes 2–3 vs. lane 1), and in Nedp1 −/− cells, modified PCNA increased significantly compared to WT cells under oxidative stress (Fig. 3F, lanes 5–6 vs. lanes 2–3). Meanwhile, the result of co-IP showed that the interaction between PCNA and NEDP1 decreased after H2O2 treatment (Fig. 3G). Moreover, expression of NEDP1 abolished PCNA NEDDylation and inhibited NEDDylated PCNA under oxidative stress (Fig. 3H), but Nedp1 knockout enhanced PCNA NEDDylation significantly (Fig. 3I). These findings indicate that NEDP1 is involved in the DDR, and the increase in PCNA NEDDylation under H2O2-induced oxidative stress is associated with dissociation of NEDP1 from PCNA. Together, these results demonstrate that PCNA NEDDylation participates in oxidative stress-induced DDR.
RAD18 is essential for the accumulation of NEDDylated PCNA under oxidative stress
Because RAD18 acted as an E3 ligase of PCNA NEDDylation, we questioned whether oxidative stress-induced accumulation of NEDDylated PCNA is dependent on RAD18. In H2O2 treated WT cells, we observed an obvious increase in modified PCNA in the TIF, while in Rad18 −/− cells, we did not detect modified bands of PCNA under oxidative stress (Fig. 4A). As we could not directly distinguish PCNA NEDDylation from ubiquitination in Fig. 4A, we thus performed Ni2+ pull-down assay to detect NEDDylated PCNA. As shown in Fig. 4B, no NEDDylated PCNA could be detected in Rad18 −/− cells, even with H2O2 treatment. In addition, WT RAD18 but not the inactive enzyme mutants of RAD18, C28F or ΔRING could rescue PCNA NEDDylation (Fig. 4B and 4C). These data indicate that RAD18 is required for PCNA NEDDylation to participate in the DDR pathway.

RAD18 is essential for the accumulation of NEDDylated PCNA under oxidative stress. (A) Immunoblotting analysis of the effect of RAD18 on oxidative stress-induced PCNA ubiquitination and NEDDylation in the TIF from WT or Rad18 −/− HEK293T cells. Cells were treated with 800 μmol/L H2O2 for the indicated time, and then the TIF was isolated and subjected to staining with anti-PCNA antibody. (B and C) Immunoblotting analysis of the effect of RAD18 on PCNA NEDDylation under H2O2-induced oxidative stress. WT and Rad18 −/− HEK293T cells were transfected with the indicated plasmids, and at 36 h after transfection, the cells were treated with or without 800 μmol/L H2O2 for 30 min before PCNA NEDDylation was detected as described above. (D) Immunofluorescence analysis of RAD18 localization after H2O2 treatment. HeLa cells were treated with or without 800 μmol/L H2O2 for 30 min, and then endogenous RAD18 was examined by immunofluorescence staining using anti-RAD18 antibody. The scale bar is 10 μm. (E) Analysis of the effect of RAD18 on cell survival against oxidative stress-induced DNA damage. HeLa WT and Rad18 −/− cells were cultured and treated with the indicated doses of H2O2, and then the cells were cultured for 12 days before colony counting. The error bars represented the standard error of the mean (SEM). Anti-RAD18 antibody was used for Western blotting analysis
As RAD18 was essential in the cellular response to UV irradiation-induced stress (Watanabe et al., 2004), we next investigated whether RAD18 accumulated at DNA damage sites induced by H2O2. As shown in Fig. 4D, RAD18 formed foci in HeLa cells treated with H2O2, suggesting that RAD18 indeed participates in the H2O2-induced DDR pathway. To further investigate whether RAD18 is required for cell survival under H2O2-induced oxidative stress, we performed colony-forming assay and found that the survival ability of Rad18-deficient cells under oxidative stress reduced obviously (Fig. 4E). This finding suggests that RAD18 is essential for the cellular response to H2O2-induced oxidative stress and PCNA NEDDylation may play an important role in this process.
NEDDylation antagonizes PCNA ubiquitination in response to oxidative DNA damage
Because oxidative stress induces both PCNA NEDDylation and ubiquitination, we sought to clarify the relationship between these two modifications. First, we detected whether the change of PCNA-NEDD8 signal was different from that of ubiquitinated PCNA in response to DNA damage. As shown in Fig. 5A, PCNA ubiquitination increased significantly and reached a peak at 2 min, whereas PCNA NEDDylation got to the peak at 15 min, later than its ubiquitination (Figs. 3A, 5A, right panel). In UV-treated cells, we also found that PCNA ubiquitination reached a peak at 6 h after UV treatment, but decreased at 12 h, while its NEDDylation got to the peak at 12 h and lasted for 24 h (Fig. S2D). As PCNA NEDDylation and ubiquitination shared the same modified lysine K164, we want to know whether PCNA NEDDylation and ubiquitination compete with each other. As shown in Fig. 5B, expression of NEDD8 inhibited PCNA ubiquitination (line 2 vs. line 1, left panel), and vice versa (line 2 vs. line 1, right panel), suggesting that PCNA NEDDylation antagonizes its ubiquitination. In addition, in Nedp1 −/− cells, we examined whether Nedp1 knockout affected PCNA ubiquitination. As shown in Fig. 5B, PCNA ubiquitination decreased (line 3 vs. line 1, left panel) but its NEDDylation increased (line 3 vs. line 1, right panel) in Nedp1 −/− cells, and exogenous NEDD8 inhibited PCNA ubiquitination (line 4 vs. line 3, left panel), and vice versa (line 4 vs. line 3, right panel). Moreover, in Ubc12 −/− cells, we found that PCNA ubiquitination increased, which was reduced by UBC12 re-expression (Fig. S3A). This data also indicates that NEDD8 antagonizes PCNA ubiquitination.

NEDDylation regulates oxidative DNA damage-induced PCNA ubiquitination. (A) A time course analysis of NEDDylated PCNA and ubiquitinated PCNA in response to H2O2 treatment. HEK293T cells were transfected with indicated plasmids for 36 h and treated with or without 800 μmol/L H2O2 for different time. And then PCNA ubiquitination and NEDDylation were analyzed by Ni-PD and Western blotting with anti-PCNA antibody. (B) Immunoblotting analysis of the effect of NEDD8 expression and Nedp1 deletion on PCNA ubiquitination. HEK293T WT or Nedp1 −/− cells were transfected with the indicated plasmids for 36 h and then Ni-PD assay was performed to analyze PCNA ubiquitination and NEDDylation. (C) Analysis of the effect of NEDD8 expression on the accumulation of modified PCNA in TIF. Empty vector or HA-NEDD8 was transfected into HEK293T cells, and then the TIF was separated and analyzed as described above. (D) Analysis of the effect of NEDD8 on PCNA ubiquitination in response to H2O2 treatment. In (D) and (E), HEK293T WT or Nedp1 −/− cells were transfected with indicated plasmids for 36 h and then treated with or without 800 μmol/L H2O2 for different time. Then PCNA ubiquitination and NEDDylation were analyzed as described above. (E) Analysis of the effect of Nedp1 deletion on accumulation of ubiquitinated PCNA in response to H2O2 treatment
To investigate the effect of NEDDylation on PCNA ubiquitination in response to DNA damage, we treated cells with H2O2 and performed chromatin fractionation to detect PCNA modification. We found that NEDD8 blocked the increase of PCNA modification in the TIF (Fig. 5C). Furthermore, Ni2+ pull-down assay also showed that NEDD8 antagonized the accumulation of ubiquitinated PCNA (Fig. 5D, left panel), and vice versa (Fig. 5D, right panel). Moreover, we assessed whether Nedp1 knockout and Ubc12 knockout affected PCNA ubiquitination induced by H2O2 treatment. As shown in Fig. 5E, compared to WT cells, the accumulation of ubiquitinated PCNA was significantly inhibited in Nedp1 −/− cells. And in Ubc12 −/− cells, PCNA ubiquitination increased more significantly, while UBC12 re-expression reduced this increase (Fig. S3B). These results indicate that enhanced NEDDylation antagonizes PCNA ubiquitination in response to oxidative stress.
USP1 is a deubiquitinase of PCNA, which decreases under UV-induced stress (Huang et al., 2006). Here, we also examined whether USP1 affected PCNA NEDDylation. As shown in Fig. S4A, USP1 inhibited PCNA ubiquitination but not affected PCNA NEDDylation. In addition, in response to H2O2 treatment, we found that USP1 reduced the accumulation of ubiquitinated PCNA (Fig. S4B) but not inhibited NEDDylated PCNA (Fig. S4C), suggesting that USP1 does not affect PCNA NEDDylation.
NEDDylation pathway regulates polη recruitment
As NEDD8 antagonizes PCNA ubiquitination, we wondered whether promotion of NEDDylation affects PCNA-polη interaction. Notably, immunoprecipitation of PCNA by anti-Flag antibody enriched polη under oxidative stress (Fig. 6A, lane 3). Moreover, in cells not treated with H2O2, expression of ubiquitin but not NEDD8 promoted the recruitment of polη to PCNA (Fig. 6A, lanes 4 and 5). These results indicate that PCNA ubiquitination is required for the recruitment of polη and PCNA NEDDylation does not directly promote this recruitment. Next, we examined the PCNA-polη interaction in Nedp1 −/− cells and Ubc12 −/− cells, and detected the effect of NEDD8 on polη foci in response to H2O2 treatment. As shown in Fig. 6B, Nedp1 knockout reduced the interaction between PCNA and polη, suggesting that enhanced NEDDylation reduced PCNA ubiquitination, thus impeded polη recruitment. However, in Ubc12 −/− cells, PCNA-polη interaction increased (Fig. 6C), indicating that loss of PCNA NEDDylation promotes polη recruitment. In addition, Immunofluorescence assay showed that NEDD8 expression blocked polη foci formation under oxidative stress (Fig. 6D and 6E). Together, these results demonstrate that up-regulation of NEDDylation by NEDD8 overexpression or Nedp1 knockout antagonizes PCNA ubiquitination and thus inhibits polη recruitment, while Ubc12 knockout promotes PCNA ubiquitination and enhances polη recruitment.

NEDDylation affects polη recruitment under oxidative stress. (A) Analysis of the effect of ubiquitin and NEDD8 on PCNA-polη interaction in response to H2O2 treatment. In (A–C), HEK293T WT, Nedp1 −/− or Ubc12 −/− cells were transfected with the indicated plasmids. At 36 h after transfection, cells were treated with 800 μmol/L H2O2 for the indicated time, and then cells were lysed in Triton-X100 lysis buffer and sonicated for 15 times. Co-IP was performed using antibody against anti-Flag or anti-Myc antibody. (B and C) Analysis of the effect of Nedp1 knockout (B) or Ubc12 knockout (C) on PCNA-polη interaction in response to H2O2 treatment. Fold change of PCNA modification intensity was analyzed using odyssey imaging systems. (D) Immunofluorescence analysis of the effect of NEDD8 on H2O2-induced polη foci. HeLa cells were transfected with His-NEDD8 for 24 h, followed by 800 μmol/L H2O2 treatment for 30 min. Then immunofluorescence microscopy was performed to analyze the formation of polη foci. The scale bar is 10 μm. (E) Quantitative analysis of polη foci under oxidative stress. Cells (100 in total) with polη foci ≥10 were counted, and quantitative results are provided as mean ± SEM
NEDDylation regulates cell sensitivity to H2O2-induced oxidative stress
As polη-mediated TLS is essential for genome integrity maintenance and cell survival under UV treatment (Han et al., 2014), we further detected whether NEDDylation regulated cell sensitivity to H2O2 treatment. As shown in Fig. 7A, HEK293T WT or Nedp1 −/− cells were treated with H2O2 at different doses, followed by incubating with CCK8 (cell counting kit 8) and then cell density was examined. In comparison to WT cells, Nedp1 −/− cells were less resistant against H2O2 treatment, while NEDP1 re-expression increased cell survival (Fig. 7A). This result indicates that enhanced NEDDylation increases cell sensitivity to H2O2 treatment.

NEDDylation regulates cell sensitivity to H 2 O 2 -induced oxidative stress. (A and B) Analysis of the effect of NEDP1 deletion (A) or UBC12 deletion on cell sensitivity in response to H2O2 treatment. In (A and B), HEK293T WT, Nedp1 −/− or Ubc12 −/− cells were cultured in 96-well plates. After 24 h, cells were treated with H2O2 at indicated dose for 1 h, and then incubated with CCK8 for 4 h. OD450 was measured using BioTek Cytation 5, triplicates were performed. Results are provided as mean ± SD. (C and D) Analysis of the effect of UBC12 deletion on cell sensitivity in response to cisplatin (C) or HU (D) treatment. In (C and D), HEK293T WT or Ubc12 −/− cells were cultured and cell cytotoxicity assay was performed as described above
While in Ubc12 −/− cells, we found that UBC12 deletion increased cell survival while UBC12 re-expression increased cell sensitivity to oxidative stress induced by H2O2 treatment (Fig. 7B). In addition, Ubc12 −/− cells were more resistant against cisplatin or HU (hydroxyurea) treatment in comparison to WT cells (Fig. 7C and 7D). These data indicate that in Ubc12 −/− cells, impaired NEDDylation increases cell survival in response to DNA damaging agents.
DISCUSSION
In our study, we identify an important role for NEDD8 in regulating polη recruitment in the DDR pathway. As shown in Fig. 8A, we demonstrate that PCNA is modified by NEDD8 at the conserved Lys164. PCNA NEDDylation utilizes the conserved NEDD8 system, which is catalyzed by an E3 ligase RAD18. NEDP1 acts as the deNEDDylase of PCNA, and in response to oxidative damage, NEDP1 disassociates from PCNA and results in the accumulation of NEDDylated PCNA (Fig. 8B). More importantly, Ubc12 knockout promotes PCNA ubiquitination while Nedp1 knockout or NEDD8 expression inhibits PCNA ubiquitination and further reduces polη recruitment.

Model of PCNA NEDDylation. (A) Illustration of PCNA NEDDylation cascade. In an ATP-dependent manner, a NEDD8-adenylate intermediate is produced to form a NEDD8-E1 (APPBP1-UBA3) complex. Then NEDD8 is transferred from the E1 to the E2 (UBC12), which combines with the E3 ligase RAD18. NEDD8 is subsequently conjugated to the Lys164 of PCNA. In this process, MLN4924 can inhibit the function of E1, thus impeding PCNA NEDDylation, and NEDP1 acts as the deNEDDylase of PCNA. (B) The molecular function of PCNA NEDDylation in regulating its monoubiquitination and TLS. In cells without stress, PCNA acts as a clamp to recruiting polε for the normal replication process. When cells were under oxidative DNA damage, monoubiquitinated PCNA recruits TLS polymerase polη to bypass the lesions. Meanwhile, NEDP1 disassociates from PCNA and PCNA is NEDDylated. PCNA NEDDylation antagonizes its monoubiquitination and leads to defective PCNA-polη interaction and reduced TLS activity
Recent studies have revealed that PCNA is targeted by ubiquitin and SUMO at the same Lys164, and the ubiquitination and SUMOylation of PCNA induced by DNA damage promotes different branches of DNA damage bypass (Hoege et al., 2002; Papouli et al., 2005). Interestingly, our results show that PCNA NEDDylation also occurs at Lys164 and utilizes the same E3 ligase RAD18, suggesting a close relationship between PCNA ubiquitination and NEDDylation. Thus, it is interesting to clarify the functional correlation between these two posttranslational modifications of PCNA. Interestingly, in response to H2O2 treatment, we found that the peak of NEDDylated PCNA occurred later than that of ubiquitinated PCNA. Moreover, NEDD8 expression inhibits PCNA ubiquitination, and increased PCNA NEDDylation in Nedp1 −/− cells suppressed its ubiquitination, but impaired PCNA NEDDylation in Ubc12 −/− cells promoted its ubiquitination (Figs. 5 and S3). These data indicate that PCNA NEDDylation antagonizes its monoubiquitination in response to DNA damage and provides a new regulatory mechanism for the involvement of PCNA ubiquitination in the process of TLS. As RAD18 catalyzes both PCNA NEDDylation and ubiquitination, it is important to explain how RAD18 could recognize ubiquitin and NEDD8. Our results show that RAD18 interacts with UBC12 and catalyzes PCNA NEDDylation in a UBC12-dependent manner (Fig. 2). Meanwhile, RAD18 also forms a complex with RAD6B, an E2 of ubiquitin, to mono-ubiquitinate PCNA (Bailly et al., 1994). These observations suggest that RAD18 catalyzes both NEDDylation and ubiquitination of PCNA through interacting with their respective E2s.
It is well known that the monoubiquitination of PCNA plays a key role in the regulation of TLS. In response to DNA damage, PCNA is monoubiquitinated by the RAD18-RAD6 complex, which provides a platform for recruiting certain repair factors and specific translesion DNA polymerase such as polη (Hoege et al., 2002; Kannouche et al., 2004). The specialized TLS polymerases then bypass the DNA damage sites without disturbing the continuity of the replication fork but results in a high level of mutagenesis. Our data revealed that PCNA NEDDylation occurred later than its ubiquitination, ubiquitin but not NEDD8 could promote the interaction between PCNA and polη. In addition, Ubc12 knockout enhanced PCNA-polη interaction, but NEDD8 expression or NEDP1 deletion reduced the recruitment of polη (Fig. 6), indicating that enhanced NEDDylation reduces excessive polη staying on the replication fork to avoid high rate of mutagenesis. We therefore conclude that under H2O2 or UV-induced stress, PCNA NEDDylation occurs and antagonizes PCNA monoubiquitination and controls polη recruitment to avoid the excessive error-prone bypass.
Furthermore, the discovery of the detailed mechanism underlying how NEDP1 regulates TLS is important. In response to H2O2-induced oxidative stress, we found that NEDP1 disassociated from PCNA, consistent with the increase of PCNA NEDDylation (Fig. 3F and 3G). In addition, in the Nedp1 −/− cells, NEDD8-conjugates including NEDDylated PCNA increased obviously, but PCNA ubiquitination reduced significantly (Fig. 5B). This result indicates that the dissociation of NEDP1 from PCNA contributes to the increase of NEDDylated PCNA. Moreover, NEDP1 deletion reduced PCNA-polη interaction, thus prevented TLS pathway and increased cell sensitivity to H2O2-induced oxidative stress (Figs. 6B and 7A). Together, we demonstrate that NEDP1 regulates TLS and cell survival through regulating PCNA NEDDylation and its ubiquitination.
As both the increased ratio of free NEDD8 over ubiquitin and oxidative stress induce global atypical NEDDylation using ubiquitin enzyme UBE1, which could transfer NEDD8 to RAD6B (also called UBE2B), the E2 of PCNA ubiquitination (Hjerpe et al., 2012; Leidecker et al., 2012), it is possible that the ubiquitin enzymes cascade UBE1-RAD6B-RAD18 can also be utilized to mediate oxidative stress-induced PCNA NEDDylation. Our in vivo and in vitro NEDDylation assay showed that UBC12 is an E2 of PCNA NEDDylation (Fig. 2C and 2D) and MLN4924 could inhibit PCNA NEDDylation significantly when NEDD8 was overexpressed or under oxidative stress (Figs. 1B and S2C); therefore, NEDD8 should utilize common NEDD8 enzymes to target PCNA. Consideration of the criteria for identifing NEDD8 substrates (Enchev et al., 2015), in future, more experiments such as constructing cells with endogenously expressed His-tagged NEDD8 using CRISPR-CAS9 technology should be performed to validate the endogenous NEDDylation and investigate the functional significance of PCNA NEDDylation.
As mutation and inactivation of polη cause xeroderma pigmentosum variant (XPV), a type of autosomal recessive disease, which is prone to developing malignant skin cancer (Johnson et al., 1999; Han et al., 2014), we evaluated the clinical relevance of NEDDylation in skin cancer by bioinformatics analysis using SKCM (Skin Cutaneous Melanoma) dataset from GEPIA (gene expression profiling interactive analysis) (http://gepia.cancer-pku.cn/). As shown in Fig. S5A, the expression of polη was significantly higher in SKCM tissues in comparison to that of normal tissues. Meanwhile, correlative analysis revealed that low level of polη is associated with higher survival of SKCM patients (Fig. S5B), indicating that the highly expressed polη is associated with SKCM. In addition, we found that UBC12 was also expressed highly in the SKCM samples (Fig. S5C), suggesting that defective TLS activity caused by abnormal NEDDylation is also correlated with SKCM. In addition, in response to H2O2 or cisplatin treatment, as UBC12 deletion reduced cell sensitivity while UBC12 re-expression reduced cell survival (Fig. 7B and 7C), it is possible that enhanced NEDDylation of PCNA could reduce TLS activity and the sensitivity of tumor cells to cisplatin treatment, which needs further study in the future.
In summary, we identify PCNA as a new substrate of NEDD8 that has an important role in the DDR pathway. In response to oxidative DNA damage, NEDDylation regulates polη recruitment and cell survival by antagonizing PCNA ubiquitination. Overall, the present study provides new insights into the mechanism through which NEDDylation regulates the oxidative stress-induced DDR.
MATERIALS AND METHODS
Cell culture and transfection
HEK293T and HeLa cells (ATCC) were grown in Dulbecco’s Modified Eagle medium (Gibco, Carlsbad, CA, USA) supplemented with 10% fetal bovine serum in 5% CO2 at 37°C, and after 24 h in culture, cells were transfected with the indicated plasmids using the transfection reagent PEI (Polyscience, Warrington, PA, USA) following the manufacturer’s protocol.
Plasmids constructions and antibodies
3Flag-PCNA, 3Flag-PCNA-K164R, 3Flag-RAD18, 3Flag-RAD18-C28F, 3Flag-RAD18-ΔRING, 3Flag-NEDP1, and 3Flag-NEDP1-C163S were cloned into the pCDNA-3Flag vector through restriction sites EcoRI and XhoI. HA-PCNA was constructed by PCR and cloned into the pRK-HA vector. 3HA-polη was cloned into the pCMV-3HA vector. To construct His-Ubiquitin, His-NEDD8 and His-PCNA, the full-length cDNAs of ubiquitin, NEDD8 and PCNA were digested with EcoRI and NotI and inserted into the pEF1-c-6His vector. Myc-PCNA and Myc-UBC12 were cloned into pCDNA-Myc vector.
PCNA mouse monoclonal antibody (BE0029) and histone H3 monoclonal antibody (BE3015) were purchased from EASYBIO (Seoul, Korea); RAD18 rabbit monoclonal antibody (D2B8) was purchased from CST (Danvers, MA, USA); polη rabbit monoclonal antibody (BS6695) was purchased from Bioworld (St. Louis Park, MN, USA); NEDD8 rabbit monoclonal antibody (Y297) was purchased from Epitomics (Cambridge, MA, USA); HA mouse monoclonal antibody (H9658) and Flag mouse monoclonal antibody (F3165) were purchased from Sigma-Aldrich (St. Louis, MO, USA); His mouse monoclonal antibody (D291-3) and actin rabbit monoclonal antibody (PM053) were purchased from MBL (Nagano, Japan); and NEDP1 monoclonal antibody (F1512) was purchased from Santa Cruz Biotechnology (Dallas, TX, USA).
Nedp1 and Ubc12 knockout cell line
The Nedp1 −/− and Ubc12 −/− cell line were generated using the CRISPR/CAS9 method. The sgRNA sequences targeting NEDP1 were tacatggacagtctactg-NGG and gccatgttccttgaaccac-NGG. The sgRNA sequences targeting UBC12 were gagtcggcgggcggcaccaa-NGG and ctgcggatccagaagggtag-NGG. HEK293T cells were transfected with sgRNA vector and CAS9 vector, 48 h later, cells expressing sgRNAs were divided by FACS for green fluorescence and single cells were plated in a 96-well dish to screen for positive monoclonal cells.
Rad18 knockout cell line
The Rad18 −/− cell line was generated using the transcription activator-like effector nuclease (TALEN) method. The Fast TALETM TALEN Assembly Kit (SIDANSAI, Shanghai, China) was used to construct TALEN vectors targeting RAD18 by one-step ligation. The target sequences were L-ttcttgatcagagaaat and R-tttcttttatcaacaact. HEK293T cells were transfected with TALEN vectors and selected in the presence of puromycin for 3 days until all control cells transfected with empty vectors had died. The remained cells were diluted and plated in a 96-well dish to screen for positive monoclonal cells.
Ni2+ Pull-down assay (Ni-PD)
His-tagged NEDD8 conjugates and ubiquitin conjugates were maintained as previously described in denaturing conditions (Li et al., 2014). HEK293T cells transfected with the indicated plasmids were lysed in 6 mL lysis buffer for 30 min and then incubated with 80 μL Ni2+ resin for 4 h. The lysates were washed with buffers 1, 2, 3, and 4 in turn, and then the modified proteins were eluted in 30 μL elution buffer and boiled in 30 μL 2× SDS loading buffer. The samples were subjected to Western blotting analysis.
Subcellular fractionation and immunoprecipitation
HEK293T cells were cultured in 10-cm dishes and transfected with the indicated plasmids. Cells were harvested in 10 mL pre-cooled PBS and lysed for 30 min in 0.2% Triton lysis buffer with 5 μL cocktail (1 mmol/L phenylmethylsulfonyl fluoride, 1 μg/mL leupeptin, 1 μg/mL aprotinin, and 1 μg/mL pepstatin). Then the lysates were isolated as the Triton-soluble fraction, and the pellets were resuspended in 2× SDS loading buffer as the Triton-insoluble fraction or in buffer appropriate for other assays.
For immunoprecipitation assays, cells were lysed in 0.2% Triton lysis buffer, and the resulting pellets were resuspended in NP-40 lysis buffer with 0.1% SDS before being subjected to sonication fifteen times (5 s each) at 200 W. Next, the lysates were incubated with the indicated antibodies overnight at 4°C and precipitated by protein G for 5 h. After three washes with NP-40 lysis buffer, the samples were boiled with 2× SDS loading buffer and subjected to SDS-polyacrylamide gel electrophoresis (PAGE) and Western blotting analysis.
In vitro NEDDylation assay
In vitro NEDDylation analysis was performed using the NEDDylation kit (Enzo, Farmingdale, NY, USA). GST-PCNA and His-SUMO-RAD18 recombinant proteins were purified from E. coli. For the NEDDylation assay, 1 μg GST-PCNA and 1 μg His-SUMO-RAD18 were incubated at 37°C for 1 h with E1, E2, NEDD8 in a total reaction volume of 20 μL following the product manual. Reaction was terminated by adding 20 μL 2× SDS loading buffer and detected by Western blotting.
Immunofluorescence
HeLa cells were cultured on cover-glass in 6-well plates and transfected with the indicated plasmids for 24 h. Cells were then treated with 800 μmol/L H2O2 for 30 min, washed with 5 mL cold PBS, and lysed for 30 min in 0.2% Triton or NP-40 lysis buffer. Immunofluorescence staining was performed as described previously (Li et al., 2014). Cells were mounted with medium containing DAPI, and fluorescence images were obtained using a DuoScan microscope and LSM 710 NLO (Carl Zeiss, Oberkochen, Germany).
Colony formation assay
Aliquots of 500 HeLa WT or Rad18 −/− cells were treated with the indicated dose of H2O2 for 1 h and then cultured in 6-well plates for 12 days. Colonies were fixed and stained with crystal violet (Amresco, Solon, OH, USA), and the numbers of colonies were counted.
Cell cytotoxicity assay
Aliquots of 10,000 HEK293T WT, Nedp1 −/− or Ubc12 −/− cells were cultured for 24 h in 96-well plates, then cells were treated with indicated dose of H2O2 for 1 h, or cisplatin and HU for 24 h. Next, cells were incubated with 90 μL medium and 10 μL CCK8 (Cell Counting Kit-8, Dojindo, Japan) for 4 h, then OD450 was measured using BioTek Cytation 5 (BioTek Instruments, USA).
Bioinformatics analysis
Bioinformatics analyses were performed to analyze protein expressions of UBC12 and polη in patients of SKCM (skin cutaneous melanoma) using dataset of SKCM from GEPIA (gene expression profiling interactive analysis) (http://gepia.cancer-pku.cn/). And the correlation between the expression level of polη and SKCM prognosis was also analyzed.
References
Aoki I, Higuchi M, Gotoh Y (2013) NEDDylation controls the target specificity of E2F1 and apoptosis induction. Oncogene 32:3954–3964
Bailly V, Lamb J, Sung P, Prakash S, Prakash L (1994) Specific complex formation between yeast RAD6 and RAD18 proteins: a potential mechanism for targeting RAD6 ubiquitin-conjugating activity to DNA damage sites. Genes Dev 8:811–820
Bergink S, Jentsch S (2009) Principles of ubiquitin and SUMO modifications in DNA repair. Nature 458:461–467
Bienko M, Green CM, Crosetto N, Rudolf F, Zapart G, Coull B, Kannouche P, Wider G, Peter M, Lehmann AR et al (2005) Ubiquitin-binding domains in Y-family polymerases regulate translesion synthesis. Science 310:1821–1824
Branzei D, Foiani M (2008) Regulation of DNA repair throughout the cell cycle. Nat Rev Mol Cell Biol 9:297–308
Brown JS, Lukashchuk N, Sczaniecka-Clift M, Britton S, le Sage C, Calsou P, Beli P, Galanty Y, Jackson SP (2015) Neddylation promotes ubiquitylation and release of Ku from DNA-damage sites. Cell Rep. 11(5):704–714
Coleman KE, Bekes M, Chapman JR, Crist SB, Jones MJ, Ueberheide BM, Huang TT (2017) SENP8 limits aberrant neddylation of NEDD8 pathway components to promote cullin-RING ubiquitin ligase function. Elife. doi:10.7554/eLife.24325
Enchev RI, Schulman BA, Peter M (2015) Protein neddylation: beyond cullin-RING ligases. Nat Rev Mol Cell Biol 16:30–44
Gao F, Cheng J, Shi T, Yeh ET (2006) Neddylation of a breast cancer-associated protein recruits a class III histone deacetylase that represses NFkappaB-dependent transcription. Nat Cell Biol 8:1171–1177
Hakenjos JP, Bejai S, Ranftl Q, Behringer C, Vlot AC, Absmanner B, Hammes U, Heinzlmeir S, Kuster B, Schwechheimer C (2013) ML3 is a NEDD8- and ubiquitin-modified protein. Plant Physiol 163:135–149
Han J, Liu T, Huen MS, Hu L, Chen Z, Huang J (2014) SIVA1 directs the E3 ubiquitin ligase RAD18 for PCNA monoubiquitination. J Cell Biol 205:811–827
Harrison JC, Haber JE (2006) Surviving the breakup: the DNA damage checkpoint. Annu Rev Genet 40:209–235
Hedglin M, Pandey B, Benkovic SJ (2016) Characterization of human translesion DNA synthesis across a UV-induced DNA lesion. Elife. doi:10.7554/eLife.19788
Hjerpe R, Thomas Y, Chen J, Zemla A, Curran S, Shpiro N, Dick LR, Kurz T (2012) Changes in the ratio of free NEDD8 to ubiquitin triggers NEDDylation by ubiquitin enzymes. Biochem J 441:927–936
Hochstrasser M (2009) Origin and function of ubiquitin-like proteins. Nature 458:422–429
Hoege C, Pfander B, Moldovan GL, Pyrowolakis G, Jentsch S (2002) RAD6-dependent DNA repair is linked to modification of PCNA by ubiquitin and SUMO. Nature 419:135–141
Hoeijmakers JH (2001) Genome maintenance mechanisms for preventing cancer. Nature 411:366–374
Huang TT, Nijman SM, Mirchandani KD, Galardy PJ, Cohn MA, Haas W, Gygi SP, Ploegh HL, Bernards R, D’Andrea AD (2006) Regulation of monoubiquitinated PCNA by DUB autocleavage. Nat Cell Biol 8:339–347
Huang DT, Ayrault O, Hunt HW, Taherbhoy AM, Duda DM, Scott DC, Borg LA, Neale G, Murray PJ, Roussel MF et al (2009a) E2-RING expansion of the NEDD8 cascade confers specificity to cullin modification. Mol Cell 33:483–495
Huang J, Huen MS, Kim H, Leung CC, Glover JN, Yu X, Chen J (2009b) RAD18 transmits DNA damage signalling to elicit homologous recombination repair. Nat Cell Biol 11:592–603
Jackson SP, Durocher D (2013) Regulation of DNA damage responses by ubiquitin and SUMO. Mol Cell 49:795–807
Jimeno S, Fernandez-Avila MJ, Cruz-Garcia A, Cepeda-Garcia C, Gomez-Cabello D, Huertas P (2015) Neddylation inhibits CtIP-mediated resection and regulates DNA double strand break repair pathway choice. Nucleic Acids Res 43:987–999
Johnson RE, Kondratick CM, Prakash S, Prakash L (1999) hRAD30 mutations in the variant form of xeroderma pigmentosum. Science 285:263–265
Kamitani T, Kito K, Nguyen HP, Yeh ET (1997) Characterization of NEDD8, a developmentally down-regulated ubiquitin-like protein. J Biol Chem 272:28557–28562
Kannouche P, Broughton BC, Volker M, Hanaoka F, Mullenders LH, Lehmann AR (2001) Domain structure, localization, and function of DNA polymerase eta, defective in xeroderma pigmentosum variant cells. Genes Dev 15:158–172
Kannouche PL, Wing J, Lehmann AR (2004) Interaction of human DNA polymerase eta with monoubiquitinated PCNA: a possible mechanism for the polymerase switch in response to DNA damage. Mol Cell 14:491–500
Kirkin V, Dikic I (2007) Role of ubiquitin- and Ubl-binding proteins in cell signaling. Curr Opin Cell Biol 19:199–205
Lan H, Tang Z, Jin H, Sun Y (2016) Neddylation inhibitor MLN4924 suppresses growth and migration of human gastric cancer cells. Sci Rep 6:24218
Leidecker O, Matic I, Mahata B, Pion E, Xirodimas DP (2012) The ubiquitin E1 enzyme Ube1 mediates NEDD8 activation under diverse stress conditions. Cell Cycle 11:1142–1150
Li T, Guan J, Huang Z, Hu X, Zheng X (2014) RNF168-mediated H2A neddylation antagonizes ubiquitylation of H2A and regulates DNA damage repair. J Cell Sci 127:2238–2248
Loftus SJ, Liu G, Carr SM, Munro S, La Thangue NB (2012) NEDDylation regulates E2F-1-dependent transcription. EMBO Rep 13:811–818
Ma T, Chen Y, Zhang F, Yang CY, Wang S, Yu X (2013) RNF111-dependent neddylation activates DNA damage-induced ubiquitination. Mol Cell 49:897–907
Mahata B, Sundqvist A, Xirodimas DP (2012) Recruitment of RPL11 at promoter sites of p53-regulated genes upon nucleolar stress through NEDD8 and in an Mdm2-dependent manner. Oncogene 31:3060–3071
Mergner J, Kuster B, Schwechheimer C (2017) DENEDDYLASE1 protein counters automodification of neddylating enzymes to maintain NEDD8 protein homeostasis in arabidopsis. J Biol Chem 292:3854–3865
Moldovan GL, Pfander B, Jentsch S (2007) PCNA, the maestro of the replication fork. Cell 129:665–679
Ohh M, Kim WY, Moslehi JJ, Chen Y, Chau V, Read MA, Kaelin WG Jr (2002) An intact NEDD8 pathway is required for Cullin-dependent ubiquitylation in mammalian cells. EMBO Rep 3:177–182
Papouli E, Chen S, Davies AA, Huttner D, Krejci L, Sung P, Ulrich HD (2005) Crosstalk between SUMO and ubiquitin on PCNA is mediated by recruitment of the helicase Srs2p. Mol Cell 19:123–133
Park JM, Yang SW, Yu KR, Ka SH, Lee SW, Seol JH, Jeon YJ, Chung CH (2014) Modification of PCNA by ISG15 plays a crucial role in termination of error-prone translesion DNA synthesis. Mol Cell 54:626–638
Pfander B, Moldovan GL, Sacher M, Hoege C, Jentsch S (2005) SUMO-modified PCNA recruits Srs2 to prevent recombination during S phase. Nature 436:428–433
Rabut G, Peter M (2008) Function and regulation of protein neddylation. ‘Protein modifications: beyond the usual suspects’ review series. EMBO Rep 9:969–976
Sale JE, Lehmann AR, Woodgate R (2012) Y-family DNA polymerases and their role in tolerance of cellular DNA damage. Nat Rev Mol Cell Biol 13:141–152
Watanabe K, Tateishi S, Kawasuji M, Tsurimoto T, Inoue H, Yamaizumi M (2004) Rad18 guides poleta to replication stalling sites through physical interaction and PCNA monoubiquitination. EMBO J 23:3886–3896
Watson IR, Blanch A, Lin DC, Ohh M, Irwin MS (2006) Mdm2-mediated NEDD8 modification of TAp73 regulates its transactivation function. J Biol Chem 281:34096–34103
Wei D, Li H, Yu J, Sebolt JT, Zhao L, Lawrence TS, Smith PG, Morgan MA, Sun Y (2012) Radiosensitization of human pancreatic cancer cells by MLN4924, an investigational NEDD8-activating enzyme inhibitor. Cancer Res 72:282–293
Xirodimas DP, Saville MK, Bourdon JC, Hay RT, Lane DP (2004) Mdm2-mediated NEDD8 conjugation of p53 inhibits its transcriptional activity. Cell 118:83–97
Xirodimas DP, Sundqvist A, Nakamura A, Shen L, Botting C, Hay RT (2008) Ribosomal proteins are targets for the NEDD8 pathway. EMBO Rep 9:280–286
Zhang J, Bai D, Ma X, Guan J, Zheng X (2014) hCINAP is a novel regulator of ribosomal protein-HDM2-p53 pathway by controlling NEDDylation of ribosomal protein S14. Oncogene 33:246–254
Zhou X, Tan M, Nyati MK, Zhao Y, Wang G, Sun Y (2016) Blockage of neddylation modification stimulates tumor sphere formation in vitro and stem cell differentiation and wound healing in vivo. Proc Natl Acad Sci USA 113:E2935–2944
Zlatanou A, Despras E, Braz-Petta T, Boubakour-Azzouz I, Pouvelle C, Stewart GS, Nakajima S, Yasui A, Ishchenko AA, Kannouche PL (2011) The hMsh2-hMsh6 complex acts in concert with monoubiquitinated PCNA and Pol eta in response to oxidative DNA damage in human cells. Mol Cell 43:649–662
Zuo W, Huang F, Chiang YJ, Li M, Du J, Ding Y, Zhang T, Lee HW, Jeong LS, Chen Y et al (2013) c-Cbl-mediated neddylation antagonizes ubiquitination and degradation of the TGF-beta type II receptor. Mol Cell 49:499–510
ACKNOWLEDGEMENTS
We sincerely thank Profs. Jun Huang, Wensheng Wei, and Caixia Guo for providing the plasmids used in this study. We thank Millennium Pharmaceuticals for providing the MLN4924 used in this study. This work was supported by the National Natural Science Foundation of China (Grant Nos. 31470754, 81730080 and 31670786), the National Key Research and Development Program of China (2016YFC1302401).
ABBREVIATIONS
CtIP, CtBP interacting protein; DDR, DNA damage response; H2O2, hydrogen peroxide; HR, homologous recombination; IP, immune precipitation; NEDD8, neural precursor cell expressed developmentally downregulated 8; NHEJ, Non-homologous end joining; Ni-PD, Ni2+ pull-down; PCNA, proliferating cell nuclear antigen; polη, polymerase η; PRR, post-replication repair; SKCM, skin cutaneous melanoma; SUMO, small ubiquitin-like modifier; TIF, Triton-X100 insoluble fraction; TLS, translesion synthesis; UV, ultraviolet; WT, wild type
COMPLIANCE WITH ETHICS GUIDELINES
Junhong Guan, Shuyu Yu, and Xiaofeng Zheng declare that they have no conflict of interest. This article does not contain any studies with human or animal subjects performed by the any of the authors.
AUTHOR CONTRIBUTIONS
J.H.G. and X.F.Z. designed the experiments, interpreted data and prepared the manuscript. J.H.G. performed all of the experiments. Y.S.Y. helped to screen Nedp1 −/− and Ubc12 −/− cell lines.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Guan, J., Yu, S. & Zheng, X. NEDDylation antagonizes ubiquitination of proliferating cell nuclear antigen and regulates the recruitment of polymerase η in response to oxidative DNA damage. Protein Cell 9, 365–379 (2018). https://doi.org/10.1007/s13238-017-0455-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13238-017-0455-x