
Summary
Mesanthemum is a genus comprising 16 species in the family Eriocaulaceae and is native to Africa and Madagascar. Eriocaulaceae are characterised by a basal tuft or rosette of narrow leaves and small flowers in heads. Mesanthemum can be recognised by diplostemonous flowers and fused glandular pistillate petals. While most Mesanthemum species are large perennial herbs, two small ephemeral species from West Africa, M. albidum and M. auratum differ from the rest of the genus by their shorter life cycle, smaller size, simpler floral structures and different seed surface patterning. A molecular phylogenetic study, morphological comparisons and scanning electron microscope (SEM) examination of seed coat sculpture were carried out to determine whether they should be separated as a new genus. The molecular results indicate that the two ephemeral species are nested in the Mesanthemum clade. However, they are not closely related to each other. All species of Mesanthemum are here revised, including the description of a new species M. alenicola from Equatorial Guinea. An identification key is provided, together with taxonomic descriptions, synonymy and notes. Images of the seeds as seen under SEM are provided where available. Lectotypifications are provided for Mesanthemum albidum, M. bennae, M. pilosum, M. prescottianum, M. pubescens and M. variabile. A neotype is selected for M. rutenbergianum, which is synonymised with M. pubescens.
Similar content being viewed by others


Introduction
Mesanthemum Körn. belongs to the pantropical and warm temperate monocot family Eriocaulaceae, comprising ten genera and c. 1200 species (WCSP 2018). Most of the generic diversity occurs in South America, especially Brazil (Giulietti et al. 1995; Dokkedal et al. 2008), and the type genus Eriocaulon L. is pantropical, extending into warm temperate parts of Asia, Australia, North America and western Europe. Mesanthemum is unusual in being confined to tropical Africa and Madagascar. It has been little studied from a molecular phylogenetic point of view (Andrade et al. 2010), most previous work being associated with African Flora accounts (e.g. Phillips 1997, 2010, 2011, 2016). The family is characteristically found in permanently or seasonally wet habitats on a variety of substrates (Stützel 1998: 203), often favouring sandy soils. A few species, including M. reductum H.E.Hess, are true aquatics, occurring in rivers and lakes.
Mesanthemum was last revised by Jacques-Félix (1947), who recognised 10 species. Since then, new material has been collected, and several new species have been described (Hess 1955; Kimpouni 1994; Moldenke 1949) bringing the total to 16 accepted species (WCSP 2018). The genus is morphologically well defined, most species being robust perennials with scapes up to c. 60 cm high arising from a basal rosette of linear or linear-lanceolate leaves sometimes up to 50 cm long. These species are easily distinguished from most Eriocaulon in Africa by their more robust habit and often hairy leaves and scapes (see e.g. Mesanthemum africanum Moldenke in Fig. 1). The tiny unisexual flowers are gathered into dense heads (capitula) which are solitary on the scapes, the white-hairy flowers being supported below by an involucre of several series of usually tough, involucral bracts (Fig. 2). Sometimes, the innermost involucral bracts greatly exceed the floral disc. The flowers have six stamens and glandular petals as in Eriocaulon, but the petals of the female flowers are connate into a tube above the ovary, not free as in Eriocaulon. The seed surface structure is uniform between the perennial species, as discussed below.

Mesanthemum africanum: A in habitat (Wursten 3699-2A); B capitulum from above (Wursten 8528A); C capitulum from below (Wursten 8530A). photos: bart wursten.

Floral heads (capitula) of some Mesanthemum species. A M. angustitepalum (Desenfans 4121); B M. cupricola (Duvigneaud 4500E); C M. glabrum (Hess 52/746); D M. pilosum (Codd 7297); E M. reductum (Hess 52/2106); F M. variabile (Richards 12427).
Two slender ephemeral species from West Africa, Mesanthemum albidum Lecomte (Fig. 3A, B) and M. auratum Lecomte (Fig. 3C, D), do not conform to the general pattern set out above. Apart from their smaller short-lived habit, they have completely different seed surface structures compared to the perennial species. Therefore, the question arose as to whether these two species should be removed from Mesanthemum and placed in a separate genus. A molecular phylogenetic study was carried out to explore relationships within the genus, and especially to determine whether the two ephemerals are congeneric with the perennial Mesanthemum species. This paper revises all known Mesanthemum species with updated descriptions, nomenclature, typifications, distribution and habitat notes. A new species to science is described from Equatorial Guinea.

Mesanthemum albidum: A whole plant; B capitulum. M. auratum: C whole plant; D capitulum. photos: martin cheek.
Materials & Methods
Molecular study
For Mesanthemum albidum, M. auratum, M. prescottianum (Bong.) Körn. and M. tuberosum Lecomte, total genomic DNA was extracted from silica-gel-dried leaf materials. For M. bennae Jacq.-Fél., leaves for DNA extraction were taken from two recently-collected herbarium specimens, van der Burgt 2185 and Konomou 311. Additional species of Mesanthemum, and of two other Eriocaulaceae genera Eriocaulon and Syngonanthus were included using existing sequence data (Y. Ito et al. unpublished data). All sampling information is provided in Table 1. CTAB procedure following Doyle & Doyle (1987) was applied for DNA extraction. Four plastid regions matK, rbcL, rpoB and rpoC1, and the nuclear region PHYC were amplified by polymerase chain reactions (PCR) with PCR Premix ‘DreamTaq’ Mix in a total volume of 25 μL. Sequences of primer pairs used and the PCR profiles are presented in Table 2 and Table 3, respectively.
Amplified fragments were examined with 1% agarose gel electrophoresis and purified with Macherey-Nagel Gel and PCR Clean-up Columns. To separate the forward and reverse chains, cycle-sequencing using BigDye™ Terminator v.3.1 Cycle Sequencing Kit was performed, and the products were loaded on sequencing plates. The fragments were cleaned using sodium acetate (NaAc) precipitation and sequenced on an Applied Biosystems 3730 DNA Analyzer following the manufacturer’s protocol.
The raw sequences of Mesanthemum were preliminarily viewed and checked in Geneious R8 (http://www.geneious.com) (Kearse et al. 2012), as well as the DNA sequences shared by Y. Ito et al. (unpublished data). Forward and reverse sequences with clear pherograms were selected and combined to produce consensus sequences. MAFFT v.7 (Katoh et al. 2009; Katoh & Standley 2013) was used to align the sequences. The alignments produced by MAFFT were then manually checked and improved in PhyDE v.0.9971 (Müller et al. 2010) by examining the gaps and excluding the dubious regions.
Trees were produced with CIPRES (http://www.phylo.org, Miller et al. 2010) for the five regions separately and for a concatenated (plastid + nuclear) alignment. Syngonanthus species were used as outgroup. The concatenated alignment is available as a supplementary data file (ESM 1).
Before running the analyses, PartitionFinder v.2.1.1 (Lanfear et al. 2012) was used to determine the best fitting partition scheme and the best fitting model for each partition in the concatenated dataset. In this study, the best partition scheme is matK, PHYC, and rbcL+rpoB+rpoC1. For maximum likelihood (ML), GTR+G is the best fitting model for each partition. For Bayesian inference (BI), GTR+G is the best fitting model for matK and rbcL+rpoB+rpoC1, while HKY+G was determined as the best fitting model for PHYC.
Maximum likelihood analyses of the partitioned data were performed using RAxML v.8. 2.1 (Stamatakis 2014). Bayesian inference analyses were performed in MrBayes v.3.2.6 (Ronquist et al. 2012), with the same parameters of partitions as in the ML analyses. MrBayes was run for 100,000,000 generations for 4 runs with 4 chains, keeping a sampling frequency of every 10,000 generations. The first 25% of trees of all runs were discarded as burn-in before generating a majority-rule consensus tree. The final ML and BI trees were viewed and annotated in FigTree v.1.4.3 (http://tree.bio.ed.ac.uk/software/figtree/).
Morphological study
The morphological work was based on specimens in the Kew herbarium (K), with additional material on loan from Brussels (BR, BRLU), Paris (P) and Zurich (ZT) (Thiers continuously updated). The seeds of seven species available at K, including the two small ephemerals, were examined under a scanning electron microscope (SEM). Three seeds were taken from a representative specimen of each species and mounted dry on stubs with carbon tabs. The stubs were coated for 3 minutes and investigated with a SEM (HITACHI S-4700) with a camera system connected to a computer. The contrast, brightness and background of the images were optimised in Adobe Photoshop CC 2018.
Conservation status
For the conservation assessments, the IUCN (2012) and IUCN Standards and Petitions Subcommittee (2017) were followed. Area of Occupancy (AOO) and Extent of Occurrence (EOO) were calculated with GeoCAT (Bachman et al. 2011).
Results
Molecular study
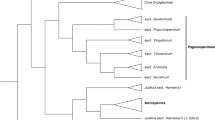
Here, trees resulting from the concatenated analyses are presented. Maximum likelihood analysis (Fig. 4), with bootstrap (BS) values) and Bayesian inference analysis (Fig. 5), with posterior probability (PP) values) show similar topologies with some small differences. Ericaulon and Mesanthemum represent monophyletic groups in both the ML and BI trees, and Syngonanthus appears as a sister to the clade comprising these two genera (Figs 4, 5).

Maximum likelihood analysis based on concatenated alignment for four plastid regions (matK, rbcL, rpoB and rpoC1) and one nuclear region (PHYC). Bootstrap (BS) values are shown.

Bayesian inference analysis based on concatenated alignment for four plastid regions (matK, rbcL, rpoB and rpoC1) and one nuclear region (PHYC). Posterior probabilities (PP) values are shown.
The monophyly of Mesanthemum is well supported (BS = 100, PP = 1) and both M. albidum and M. auratum form part of the Mesanthemum clade (Figs 4, 5), although M. auratum is sister to the rest of Mesanthemum in the BI tree (Fig. 5). In the results of the ML analysis (Fig. 4), M. auratum is sister to M. bennae (BS = 73), and the M. auratum + M. bennae clade is sister to the other species of the genus (BS = 100). A sister relationship between M. albidum and M. radicans Körn. is strongly supported in both analyses (BS = 100, PP =1). Mesanthemum prescottianum and M. tuberosum also appear as sister to each other in both trees (BS = 100, PP = 1).
Morphological study
Detailed characters for species are presented in the descriptions (see Taxonomic Treatment). The main morphological differences that distinguish large perennial Mesanthemum species from the two ephemeral Mesanthemum species, M. albidum and M. auratum, are their size, reduced floral structures and different seed coat patterning. The height of the ephemeral Mesanthemum species is generally <25 cm, while the perennial species usually grow far beyond this size, even up to 1 m (e.g. in M. radicans). Floral bracts and sepals are absent from the female flowers in M. albidum. Mesanthemum auratum has floral bracts but lacks sepals in both male and female flowers. The perennial aquatic species M. reductum with submerged heads also lacks floral bracts and sepals in both male and female flowers.
In the seed of perennial Mesanthemum species, the cells are irregularly shaped, usually 5 – 6-sided, with straight, rather prominent anticlinal cell walls. The angles bear long, thickened, pillar-like projections with expanded truncate tips, as seen in M. radicans (Figs 6A, B). The projections appear as a fringe of colourless hairs under a light microscope, when soaking the seeds in water will usually release the projections. They are held flat in the young seed by the outer cell layer of the outer integument. This disintegrates as the seed matures, as shown in the young and mature seeds of M. alenicola which is formally described below (Fig. 7). Remnants of the disintegrating outer cell layer are also visible in the images of M. bennae (Fig. 6C, D), M. prescottianum (Fig. 6E, F) and M. tuberosum (Fig. 6G, H). The background seed surface is smooth or almost so.

Seeds and details of testa morphology (SEM). A, B Mesanthemum radicans (Kent 6); C, D M. bennae (Konomou 311); E, F M. prescottianum (Baldwin 9516); G, H M. tuberosum (Porembski 97).

Seeds and details of testa morphology (SEM). Mesanthemum alenicola (Lejoly 99/25): A, B young seed with intact upper cell layer; C, D mature seed with disintegrated upper cell layer.
In Mesanthemum albidum the cells are irregularly shaped, 5 – 6-walled, and often ± isodiametric as in the perennial species (Fig. 8A, C). However, they lack pillar-like projections from the angles. Instead the anticlinal walls are strongly thickened and prominent, appearing as a white network under a light microscope. These walls in the seed from Adames 363 are heavily coated in a white deposit of crumbly appearance (Fig. 8D). This is a variable feature, as seed from Cheek 18615 has almost smooth walls, with only traces of deposit visible on some walls at high magnification (Fig. 8B). The background relief of the seed surface is verrucose.

Seeds and details of testa morphology (SEM). Mesanthemum albidum: A, B from Cheek 18615; C, D from Adames 363.
The seeds of Mesanthemum auratum differ from those in the rest of the genus, including M. albidum, as the cells are not isodiametric but are elongated in rows along the long axis of the seed (Fig. 9). There is no difference between seeds from plants with white capitula (Fig. 9A, B) or yellow capitula (Fig. 9C, D) from Guinea. The anticlinal walls are prominent but there are no pillar-like projections. Instead, each cell has a linear, slightly sinuous, raised ridge running down the centre, parallel to the long axis but separate from the cell walls. This pattern has not been observed elsewhere in African Eriocaulaceae. Occasionally, these ridges may fork or branch, or are discontinuous (Fig. 9F). The background relief of the seed surface is ruminate, including over the ridges. Seeds from a Cameroon collection (Fig. 9E, F) are larger than seeds from Guinea (Fig. 9A – D), but show no other significant differences.

Seeds and details of testa morphology (SEM). Mesanthemum auratum: A, B Mesterházy 224 (white capitula, Guinea); C, D Chillou 676 (yellow capitula, Guinea); E, F Cheek 14121 (Cameroon).
Discussion
Molecular study
Taxonomy of Eriocaulaceae has long been determined by morphological characters, especially by floral structures. In his taxonomic revision, Ruhland (1903) accepted two subfamilies, i.e. Eriocauloideae, comprised of Eriocaulon and Mesanthemum, and Paepalanthoideae, which included the rest of the family. Stützel (1985) suggested that Mesanthemum should be placed with Philodice Mart. (now accepted within Syngonanthus), Syngonanthus Ruhland and Wurdackia Moldenke (= Rondonanthus Herzog) to form Syngonanthoideae, a third subfamily, based on the fusion of the syngonanthoid corolla in the pistillate flowers. Stützel (1985) also proposed that Eriocauloideae is only comprised of Eriocaulon, and the other two subfamilies derived from it independently. However, this hypothesis was doubted by later researchers.
Unwin (2004) conducted a phylogenetic study using molecular data and proposed some evolutionary trends in the Eriocaulaceae. However, Mesanthemum was not included in that study. The phylogenetic position of Mesanthemum in the family was first confirmed by Andrade et al. (2010). Using plastid data of M. radicans, their results placed Mesanthemum as sister to Eriocaulon, and the two genera form a clade that appears as sister to the rest of the family. Their results rejected the third subfamily and strongly supported the monophyly of Eriocauloideae sensu Ruhland (Andrade et al. 2010), providing molecular support for the classification proposed by Ruhland (1903) based on floral morphology.
In the present study, besides Mesanthemum species, molecular data from six Eriocaulon species were included and Syngonanthus angolensis H.E.Hess was selected as outgroup. Our results show that the clade including both Mesanthemum and Eriocaulon is strongly supported (Figs 4, 5), which is consistent with Andrade et al. (2010).
As only one Mesanthemum species (M. radicans) was included in the previous molecular study (Andrade et al. 2010), the intrageneric relationships in Mesanthemum are revealed here for the first time. Despite the different position of M. auratum in the results of the ML and BI analyses (Figs 4, 5), both M. albidum and M. auratum are part of the Mesanthemum clade. Differing from our starting hypothesis based on vegetative and floral characters, these two ephemeral species do not appear to be closely related to each other, which accords with the significant differences in their seed coat patterns. On present evidence the two ephemeral species do not represent a different taxon but are congeneric with the perennial Mesanthemum species.
Mesanthemum albidum appears to be sister to M. radicans (Figs 4, 5), despite the considerable differences between them in morphology and life cycle. The phylogenetic position of M. auratum is less clear, appearing either as sister to M. bennae (Fig. 4) or sister to the rest of the genus (Fig. 5). This uncertainty in placement is likely the consequence of a lack of data. While sequence data for all the five regions was available for M. albidum, only two regions (matK and rpoC1) could be sequenced for M. auratum.
The similarity in size, life cycle and reduced floral parts in the two ephemeral Mesanthemum species is probably the result of convergent evolution. The very distinct seed coat sculpture of M. auratum (Fig. 9) and its potential early branching position in the tree (Fig. 5) leaves a degree of uncertainty about its phylogenetic position. Better plant material of this seldom collected species is needed for further molecular work.
Two accessions of Mesanthemum prescottianum and one of M. tuberosum are strongly supported as forming a clade (Figs 4, 5). The two species share many morphological characters, especially the daisy-like capitula, so M. tuberosum was reduced to a synonym of M. prescottianum by Jacques-Félix (1947: 144). In a recent paper (Phillips et al. 2018), M. tuberosum has been resurrected from synonymy, based primarily on field observations including different habitat preferences. Our results show M. prescottianum to be paraphyletic, but based on the limited sampling of this study we do not suggest taxonomic changes at this time.
This study provides a first view of the phylogenetic relationships in Mesanthemum, although further study is needed since some of the perennial Mesanthemum species could not be included at this time, either because the herbarium material available was insufficient or the DNA was too fragmented to enable Sanger sequencing. Further study is also needed to place M. auratum with more certainty.
Morphological study
Six reduced stamens, differentiated into filament and anther, were found around the base of the ovary in dissected female flowers of Mesanthemum pubescens (Lam.) Körn. (Fig. 10) and may be a regular feature in this species. Sterile anthers were also found in some abnormal female flowers of M. cupricola Kimp., but have not been seen elsewhere in Mesanthemum, nor in any of the many species of Eriocaulon in Africa. According to Stützel (pers. comm.) ontogenetical studies in Eriocaulaceae have shown that anthers are always initiated in the female flowers, but do not develop beyond small tissue bumps visible only in extremely young flowers under SEM.

Mesanthemum pubescens. Light microscope photos of reduced stamens around ovary base in dissected female flower (de la Bathie 16938): A ovary present; B ovary removed.
Eriocaulaceae are known to be entomophilous (Stützel 1998: 202). Insects (Coleoptera and Thysanoptera) were noted visiting the flowers of Mesanthemum radicans in Gabon (Wieringa 1139), presumably attracted by nectar from the epipetalous glands. In Mesanthemum, these glands are linear and attached only at the base. They are dark brown, often with a yellow band at the summit, especially well marked in M. bennae and M. pubescens (Fig. 11). The function of the yellow portion and details of the secretions are unknown.

Light microscope photos of glands on the petals. A, B glands topped with cap-like yellow structure in female flower of Mesanthemum bennae (Konomou 311); C tube-like gland with swollen base in female flower of M. pubescens (de la Bathie 16938).
Seeds of Eriocaulaceae species bear various characteristic ornamentations (sculpture) on the seed coat surface, and they have furnished information for the taxonomy of several genera (e.g. Giulietti et al. 1988; Ansari & Balakrishnan 1994; Phillips 1994; Zhang 1999; Barreto et al. 2013). Giulietti et al. (1988) were the first to use SEM to observe the seed coat surface in Eriocaulaceae, and they described different patterns for 10 genera in the family (Blastocaulon Ruhland is now considered as a synonym of Paepalanthus Mart., and Philodice as a synonym of Syngonanthus). Barreto et al. (2013) studied the seed coat sculptures in Comanthera L.B.Sm. and discovered that the seeds for its two subgenera have very distinct surface ornamentation. The results of these studies suggest that the seed coat sculpture in Mesanthemum may provide information that supports taxonomic hypotheses at a generic or infrageneric level.
Seed morphology in Mesanthemum has been little studied to date. Only the commonest species, M. radicans, was included in previous publications on seed coat sculpture (Giulietti et al. 1988; Phillips 1994). In this study, the seeds of several perennial species and the two ephemeral species were examined. Our results show that there are big differences in seed morphology between the ephemeral species and the rest of the genus. Interestingly, the two ephemeral species are also very distinct from each other.
In Eriocaulaceae, the testa consists of two integuments, an inner and an outer, each integument being itself composed of two cell layers (Zhang 1999). Frequently any thickened ornamentation arises in the inner layer of the outer integument, and this appears to be the case in typical perennial Mesanthemum species (Figs 6, 7). Seed morphology conforms to the same basic plan in all the robust perennial species of Mesanthemum examined: larger seeds with regular-shaped cells and characteristic projections at the corners of the anticlinal walls (Figs 6, 7). The reticulate, uniform cell arrangement formed by 5 – 6-sided cells can also be seen in other Eriocaulaceae genera. Similar patterns with long projections at the cell corners were reported in a few Eriocaulon species from Sri Lanka (Phillips 1994) and Paepalanthus gardnerianus Walp. (Giulietti et al. 1988).
The seeds of the two West African ephemeral species, Mesanthemum albidum and M. auratum, are completely different from seeds of the perennial species. No evidence was found in the SEM images of an evanescent outer cell layer (Figs 8, 9), but it may disappear very early, as is the case in some ephemeral Eriocaulon species. Furthermore, the differences in seed coat sculpture in M. albidum compared with M. auratum suggest that the two ephemeral species are not closely related to each other, which is consistent with our molecular results.
Conclusion
This study provides the first insight into the relationships between Mesanthemum species based on molecular phylogenetic data. The molecular results and the SEM study of seed coat patterns indicate that the two ephemeral species from West Africa, M. albidum and M. auratum, are not closely related to each other, and their similar habit is likely the result of convergent evolution in similar habitats. Mesanthemum albidum is nested within the generally perennial genus Mesanthemum. The case for retaining M. auratum within Mesanthemum is weaker, but we maintain it as part of Mesanthemum until further study. All Mesanthemum species are here revised. Mesanthemum alenicola is described as a new species from Equatorial Guinea, and only one species is recognised in Madagascar giving a total of 16 species. An identification key is provided, together with taxonomic notes. Images of the seeds as seen under the SEM are provided where available.
Taxonomic Treatment
Mesanthemum Körn., Linnaea 27: 572 (Körnicke 1856). Type species: Eriocaulon radicans Benth. (= Mesanthemum radicans (Benth.) Körn.).
Perennial herbs from a stout tough rhizome or rootstock, rarely aquatic with a slender rhizome; rarely slender ephemerals. Leaves in a basal tuft, linear to linear-lanceolate or ensiform, usually thick, spongy, frequently sparsely to densely hairy, old leaves glabrescent. Scapes many-ribbed, arising from the leaf axils; sheaths cylindrical, obliquely slit with lanceolate limb, scapes and sheaths often sparsely to densely pilose. Capitulum globose or flattened, 4 – 35 mm wide. Involucral bracts coriaceous or crustaceous with scarious margins (scarious in M. albidum), in several imbricate series, innermost sometimes radiating beyond the periphery of the floral disc. Floral bracts filiform with expanded, usually hairy tips, rarely absent. Flowers unisexual, trimerous, pedicellate, embedded in a woolly cushion of long receptacular hairs. Sepals free or sometimes basally connate in the male flower, usually membranous, oblanceolate-oblong, concave, often caducous, rarely absent; male flower with a short fleshy stipe between sepals and petals. Petals connate into a fleshy infundibular or oblong tube with shallowly 3-lobed upper margin, petal bases free in female flower resulting in slits in petal-tube around ovary, small slits sometimes also present in male flower, 3 linear brown glands inserted about \( \raisebox{1ex}{$2$}\!\left/ \!\raisebox{-1ex}{$3$}\right. \) up inside, petal-tube glabrous or female petal-tube often villous outside and/or within. Stamens 6 arising from base of male petal-tube; anthers usually pale creamy-yellow, rarely dark brown; vestigial gynoecium present. Ovary protruding between petal-tube slits of female flower at maturity; stigmas 3 without appendages. Seeds subglobose, brown, densely covered in white hair-like projections in perennial species.
16 species in tropical Africa, 1 species endemic in Madagascar.
Key to the species of Mesanthemum
-
1. Slender ephemerals; scapes < 25 cm high 2
-
Robust perennials from a rootstock or rhizome; scapes 25 – 75 cm high 3
-
2. Involucral bracts creamy-buff, soft, scarious; floral bracts absent; sepals present in male flower; seed white-reticulate M. albidum
-
Involucral bracts dark brown, glossy, crustaceous; floral bracts filiform, upper part conspicuously pilose; sepals absent; seed with indistinct longitudinal ridges M. auratum
-
3. Innermost involucral bracts much longer than head of flowers, widely spreading and resembling ray-florets of a daisy 4
-
Innermost involucral bracts shorter than or only slightly longer (<3 mm) than head of flowers, not widely spreading and ray-like 6
-
4. Leaves 15 – 65 mm wide; longest involucral bracts with indistinct veins, velvety on both surfaces; floral bracts bearded M. bennae
-
Leaves 2.6 – 6.0 mm wide; longest involucral bracts with obvious veins, glabrous on upper surface, glabrous or pubescent below; floral bracts glabrous 5
-
5. Longest involucral bracts shiny white, lanceolate-oblong, acute; plant from slightly thickened base M. prescottianum
-
Longest involucral bracts dull white, broadly linear, obtuse; plant from well developed fibrous corm M. tuberosum
-
6. Leaves thin, flaccid, translucent, cellular lattice structure visible; floral bracts and sepals absent; aquatic in fast flowing rivers M. reductum
-
Leaves spongy or herbaceous without lattice structure; floral bracts and sepals present; plants of marshes or seasonally wet places 7
-
7. Capitula 5 – 8 mm wide, viviparous; female flowers with petal-tube glabrous on both faces (free tips villous), sepals ˂ ½ length of petal-tube M. alenicola
-
Capitula 9 – 17 mm wide, not viviparous; female flowers with petal-tube hairy on one or both faces (M. cupricola glabrous), sepals ½ as long to equalling petal-tube 8
-
8. Innermost involucral bracts extending 0.5 – 3.0 mm beyond floral disc 9
-
Innermost involucral bracts ± equalling floral disc 11
-
9. Leaves 3 – 7 mm wide, bicoloured red-brown/green, hirsute; involucral bracts acute to cuspidate M. cupricola
-
Leaves 7 – 20 mm wide, uniformly green, usually subglabrous or with sparse hairs; involucral bracts obtuse to subacute10
-
10. Innermost involucral bracts extending 2 – 3 mm beyond floral disc; sepals colourless; female petals connate above ovary, villous within with straight grey hairs M. radicans
-
Innermost involucral bracts extending up to 1 mm beyond floral disc; sepals blackish; female petals connate near tips, lanate within with tangled blackish hairs M. pubescens
-
11. Plant with a swollen often bulbous base clothed in whitish papery leaf bases; involucral bracts soft, membranous, apiculate or acute M. jaegeri
-
Plant from a tough rootstock or rhizome, base not swollen; involucral bracts firm, cartilaginous or coriaceous (margins thinner), acute to obtuse 12
-
12. Female flowers surrounded by dense circle of long grey silky hairs (easily seen on disarticulated flowers); burned leaf remains often present at tough plant base M. pilosum
-
Female flowers without circle of surrounding long hairs outside sepals 13
-
13. Sepals colourless 14
-
Sepals black or brownish-black 16
-
14. Involucral bracts ovate-oblong with conspicuous broad pale margins, back glabrous, tip obtuse; receptacle hairs pale to dark grey-brown M. glabrum
-
Involucral bracts broadly ovate with membranous margins, back patchily hairy, tip subacute to cuspidate; receptacle hairs black 15
-
15. Plant rhizomatous; leaves 4 – 8 mm wide; capitulum flat-topped; involucral bracts sericeous; female petal-tube glabrous inside M. variabile
-
Plant tufted; leaves 7 – 10 mm wide; capitulum hemispherical; involucral bracts pilose; female petal-tube densely villous inside M. angustitepalum
-
16. Mature capitulum subglobose; female petals connate from middle, villous with dark straight hairs outside and within M. africanum
-
Mature capitulum shallowly hemispherical; female petals connate towards tips, glabrous outside, lanate inside with blackish woolly tangled hairs M. pubescens
Mesanthemum africanum Moldenke (1949: 113); Phillips (2010: 74). Type: Mozambique, Chimanimani Mts, 9 June 1948, Munch 72 (holotype NY [NY00102772]; isotypes K!, SRGH).
Rosulate perennial from a woody rootstock. Leaves yellowish-green, linear to narrowly subulate, 10 – 35 cm long, 5 – 12 mm wide, softly pilose, sometimes thinly, older leaves glabrescent, margins firm, tapering to a subacute tip. Scapes 1 – 5, 30 – 60 cm high, 1.2 – 1.5 mm thick, glabrous. Sheaths pilose, limb straight, glabrous or a few hairs within. Capitulum 10 – 14 mm wide, densely white-hairy becoming black speckled, hemispherical at first with a shallow involucre slightly smaller than capitulum width, later depressed-globose, involucre reflexing and obscured at maturity. Involucral bracts in 3 – 4 series, light brown, all similar, cartilaginous with scarious margins, ovate, 3.0 – 4.5 mm long, hirsute, veins visible, glossy brown on inner face, acute or obtuse, inner bracts with an apical patch of white hairs inside. Floral bracts filiform, tip clavate, white-pilose. Receptacle hairs black, curling and tangled. Flowers 2 – 3 mm long; sepals black. Male flowers: sepals free, oblanceolate, tips white-pilose, truncate; petals densely white-pilose at tips. Female flowers: sepals as long as petals, oblanceolate-oblong to obovate, tips erose-truncate and white-pilose, otherwise glabrous; petals free below middle, villous with long black hairs on both faces, hairs arising near petal bases, tips densely white-pilose. Seed ellipsoid, c. 1 mm long, dark brown. Fig. 1.
distribution. Zimbabwe, Mozambique; endemic to the Chimanimani mountains.
specimens examined. zimbabwe. Martin Forest Reserve, 14 Nov. 1976, Mavi 632 (K!, SRGH); Chimanimani Mts, Nov. 1947, McCosh 2 in G.H.17692 (K!; SRGH). mozambique. Manica, Madzirene Forest, Murere R. camp, 15 Nov. 2015, Darbyshire 942 (K!, LMA) & Madzireni area, W of Chiira R., 17 Nov. 2015, Darbyshire 966 (K!, LMA); Mombane, 2 March 1907, Johnson 243 (K!); Eastern slopes of Chimanimani Mts, 27 Oct. 2014, Mapaura 701 (K!); Chimanimani Mts, 8 June 1949, Munch 213 (K!, SRGH); Sussundenga Distr., slopes of Mt Binga, 20 April 2014, Timberlake 5969 (BR, K!, LMA, SRGH); E of Makurupini R., 8 Sept. 1969, Wild A71 (K!, SRGH).
habitat. Wet grassy areas around watercourses; 350 – 2400 m.
conservation status. Although this species has a restricted distribution range, it is not under any specific threat. It is locally common and reported to be spreading, especially on disturbed ground such as footpaths (J. Timberlake pers. comm.). As a consequence, it has been assessed at Least Concern (LC) by Darbyshire et al. (2017).
notes. Mesanthemum africanum can be recognised by its hirsute involucre, which becomes hidden as the floral disc matures and expands, together with the copious black receptaclar hairs, black sepals and dense black hairs on both sides of the petal-tube in the female flowers.
The petal-tube hairs arise on the basal claws and lower tube of the petals, not spread over the central portion, as in the similar but more widespread Mesanthemum pilosum. Mesanthemum rutenbergianum from Madagascar also has black receptacular hairs, black sepals and densely black-hairy female petals, but this has a different obconical, non-hirsute involucre still obvious at maturity, and only slightly connate female petals which lack hairs on the outer face.
Mesanthemum albidum Lecomte (1908, publ. 1909: 601); Jacques-Félix (1947: 144); Meikle (1968: 65). Type: Guinea (Conakry), Kindia, Pobéguin 1359 (lectotype P [P00443232], designated here).
Eriocaulon guineënse Moldenke (1962: 386). Type: Guinea (Conakry), Friguiagbé, Chillou 3904 (holotype IFAN [IFAN31807]).
Eriocaulon hirsutulum Moldenke (1962: 387). Type: Senegal, Asamame, Adam 18370 (holotype LL [LL00374547]).
Delicate ephemeral. Leaves in a basal rosette, broadly subulate, 1 – 3.5 cm long, 2 – 4 mm wide, pilose with spreading hairs to subglabrous, attenuate. Scapes erect, slender, up to about 20 but usually fewer, to c. 15 cm high, 0.2 – 0.4 mm thick, pilose with spreading hairs. Sheaths slightly longer than leaves, pilose, cylindrical in lower half, inflated above to a subacute limb. Capitulum bright white and fluffy-villous above the crateriform involucre at anthesis, 5 – 6 (– 10 mm) wide, at maturity white hairs lost and capitulum appearing pale buff. Involucral bracts in 2 – 3 series, creamy-buff, scarious, margins membranous, very broadly oblong, 2.5 – 3.0 mm long, several-veined, villous with tubercle-based hyaline hairs on exposed parts, inner bracts narrower, tips deeply lacerate. Floral bracts absent. Receptacle densely villous, hairs pallid. Flowers 1 – 2 mm long, pedicellate, a single circle of barren pedicels c. 2 mm long within involucral bracts, rarely these bearing fully developed male flowers which extend ray-like beyond the involucre, other flowers with shorter pedicels. Male flowers: sepals connate at base, oblong, villous on upper margins and back with white hairs; petals broadly obovate, connate below middle into infundibular cup, free tips very broad, slightly emarginate, villous with white hairs on upper margin and back, penicillate on inner blade from fleshy cushion above filament insertion; anthers 6, cream-coloured, vestigial gynoecium present in penicillate hair-tuft. Female flowers: sepals absent; petals spathulate, free in lower third, lightly connate above but sometimes separating, petal-tube villous outside with hyaline hairs, white-penicillate on inner face of each petal from fleshy cushion below tip, tips broadly rounded, slightly emarginate, densely villous with white hairs. Seed subspherical, 0.4 mm long, brown, conspicuously white-reticulate. Figs 3A & B, 8, 12.

Mesanthemum albidum. A habit; B capitulum; C outer involucral bract; D innermost involucral bract; E male flower; F inside view of male petal tube; G sepal of male flower; H female flower; J inside view of female petal tube; K ovary and stigmata. From Cheek 18615 and van der Burgt 2018. drawn by yuxi liang.
distribution. Guinea (Conakry), Senegal, Sierra Leone.
specimens examined. guinea (conakry). Fouta Djalon: Pita, 16 Sept. 1962, Adames 363 (K!). Forécariah: Benna Plateau, 5 km E of Gbereboun village, 25 Sept. 2016, van der Burgt 2018 (HNG, K!, P, WAG); Sengulen, Moofanyi inselberg, 28 Sept. 2012, Cheek 16749 (HNG, K!) & 23 Sept. 2015, Molmou 919 (HNG, K); Tonkoyah, inselberg near to village, 7 Oct. 2012, Molmou 560 (K!). Kindia: Kindia-Mamou rd., outskirts of Kindia, 3 Oct. 2015, Cheek 18241 (HNG, K!) & Cheek 18256 (HNG, K!); Mayon Khoure village, 20 June 2016, Cheek 18552 (HNG, K!, WAG) & Cheek 18553 (HNG, K!); N of Kindia town, towards Mt Gangan, 28 Sept. 2016, Cheek 18563 (HNG, K!) & 29 Sept. 2016, Cheek 18573 (HNG, K!); from Kindia town to Kiriya village, 1 Oct. 2016, Cheek 18615 (K!); c. 9 km N of Kindia on the Telimele road, 3 Oct. 2015, Molmou 957 (HNG, K!); near Mt Gangan, along road from Kindia to Samaya, 2 Aug. 1992, Porembski 120 (K!); 10 km S of Friguiagbé, 3 Aug. 1992, Porembski 142(K!). Kouroussa: Kouroussa, Aug. 1902, Pobéguin 1153 (K!). sierra leone. Gbap, Nonguba, Bullom, 10 Oct. 1951, Jordan 588 (K!).
habitat. Shallow wet soil in seepages on sandstone flats and on granite inselbergs, plants scattered in open populations; below 400 m.
conservation status. Mesanthemum albidum is fairly widespread but not common in West Africa. Although the extent of occurrence (EOO = 161,444 km2) is large, the area of occupancy (AOO) is only 60 km2, locations are 10, and there are major threats at several of the locations. It is assessed as Vulnerable VU B2ab(i,ii,iii,iv).
typification. There are three specimens of Pobéguin 1359 in the Paris herbarium. The specimen designated here as lectotype bears an original handwritten label and also includes a floral dissection.
Two specimens exist of the type of Eriocaulon hirsutulum Moldenke (Adam 18370), both from Senegal and collected on 8 October 1961, but with different localities: Asamame (LL) and Kabrousee (IFAN). The specimen in LL is from Moldenke’s personal herbarium and the locality matches the protologue, so this specimen is the holotype. In the protologues of Eriocaulon guineense and E. hirsutulum Moldenke describes the female flowers as having 3 oblong sepals, but no sepals were seen in any flowers of M. albidum examined. There are also further differences in the protologue of E. hirsutulum, which has only been seen as an electronic image, but the specimen Adam 18370 in IFAN was seen by Meikle and cited by him as Mesanthemum albidum Meikle (1968).
notes. This species has showy, bright white, conspicuously fluffy hairy capitula, and can be further recognised by its conspicuously white-reticulate seeds. Van der Burgt 2018 has a peripheral circle of fully developed, long pedicellate, exserted male flowers in the capitula. In all other specimens seen there are only slender rudiments resembling the pedicels of male flowers.
Mesanthemum alenicola S.M.Phillips sp. nov. Type: Equatorial Guinea, Monte Alen National Park, Lejoly 99/25 (holotype BRLU [BRLU0018216]!; isotype K!].
http://www.ipni.org/urn:lsid:ipni.org:names:77202344-1
Tufted perennial from a rootstock or short rhizome. Leaves in a loose rosette, ascending or arching outwards when short, linear or linear-subulate, flat or folded, 8 – 30 cm long, 5 – 12 mm wide, papery, pilose to hispid on both surfaces, tapering to a hard acuminate tip. Scapes up to 10, relatively slender, 30 – 56 cm high, 0.5 – 1.1 mm thick, erect at first, later drooping, hirsute with spreading colourless hairs 1 mm or longer. Sheaths shorter or as long as the leaves, narrowly cylindrical, hirsute, limb 2 – 3 cm long, acuminate. Capitulum viviparous, tuft of small leaves arising from centre of cup-shaped involucre, 5 – 8 mm wide, a circlet of white-villous flowers visible around periphery. Involucral bracts in 3 – 4 series, pale straw tinged green with paler thinner margin, crustaceous, indistinctly veined, silky-pilose on back with appressed hairs, upper margins ciliate, outermost bracts ovate, 2.3 – 2.5 mm long, inner progressively longer and usually more oblong, innermost 3.5 – 3.6 mm long, subacute or obtuse. Floral bracts few, filamentous, tip not expanded, thinly penicillate with short white hairs above middle or towards tip. Receptacle hairs fuscous or dark grey. Flowers 3 – 4 mm, female subsessile, male pedicellate; sepals colourless. Male flowers: sepals free, oblong-spathulate narrowed to a slender base, laterals keeled, median flat, membranous, back densely white-villous above middle, inner face white-villous towards tip; petal-tube fleshy with 3 small slits near base, upper margin 3-lobed, lobes rounded, densely white-villous on margin and back; anthers 6, creamy-white. Female flowers: sepals free, \( \raisebox{1ex}{$1$}\!\left/ \!\raisebox{-1ex}{$3$}\right. \) – ½ length of petal-tube, 1.0 – 1.3 mm, ovate, concave, membranous, acute or denticulate; petals linear-oblong, lightly adherent and easily splitting in central area, connate portion glabrous outside and within, free lobes densely white-villous on outer face and tips, lateral petals with dark gland, median petal eglandular. Seed almost spherical, c. 0.8 mm, dark reddish-brown, covered in clinging white prickle-hairs. Figs 7, 13.

Mesanthemum alenicola. A habit; B viviparous capitulum; C outer involucral bract; D inner involucral bract; E male flower; F female flower; G female flower opened, one sepal removed; H seed. From Lejoly 99/25. drawn by margaret tebbs.
recognition. Mesanthemum alenicola differs from the widespread species M. radicans by its more slender drooping scapes, smaller viviparous capitula with a cup-shaped (not turbinate) involucre of more obviously hairy involucral bracts, and by the female flowers with very short sepals and a glabrous (not villous) petal-tube.
distribution. Equatorial Guinea.
specimens examined. equatorial guinea. Centro Sur, Monte Alén N. P.: 3 km E of Ecofac station, 2 Jan. 1999, Lejoly 99/25 (holotype BRLU!; isotype K!) & 1°39'N 10°17'E, 8 Aug. 2001, Cabezas 134 (MA) & 1h30 walk from Ecofac site, 15 May 2002, Parmentier & Esono 3070 (BRLU).
habitat. Rock pavement in grassland and low bushland, on black soil rich in humus; 1110 – 1130 m.
conservation status. Mesanthemum alenicola is only known from three collections at one location in Monte Alén National Park in Equatorial Guinea. More information is needed to confirm that it is endemic to this location only, to assess the population size and to assess threats to this species, currently unknown. It is here assessed as Near Threatened (NT) in the absence of any credible threat being known. Should a threat be discovered the species should be reassessed as Critically Endangered (CR) under Criterion B2ab.
etymology. The specific epithet refers to the mountain range where this species occurs.
notes. This species exhibits the only known case of vivipary in Mesanthemum, although vivipary occurs in several species of Eriocaulon. It may be facultative, as a single very mature capitulum without a central viviparous leaf tuft contained a number of ripe seeds. The capitula are smaller than is usual in the genus, although the habit is otherwise typical. The sepals of the female flowers are very short, barely extending above the level of the ovary. Mesanthemum radicans is similar, but besides the differences given above usually has slightly wider leaves, and grows in swampy grassland and bog generally at lower elevations, not on rock pavement.
Mesanthemum angustitepalum Kimp. (Kimpouni 1994: 149). Type: D.R. Congo, Katanga, Upemba National Park, Mitwaba, Mumbale, April 1948, de Witte 3701 (holotype BR [BR855434]!).
Robust rosulate perennial from a tough rootstock. Leaves linear or narrowly linear-lanceolate, up to 55 cm long, 7 – 10 mm wide, herbaceous, hirsute on both surfaces, tip, subacute, hard. Scapes 1 -- 3, 28 – 55 cm high, stout, 1.6 – 1.8 mm thick, hirsute with patent hairs. Sheaths hirsute, limb acute. Capitulum 13 – 16 mm wide, hemispherical, white-hairy from an involucre with flat base as wide as floral disc. Involucral bracts in 3 – 4 series, pale greenish-brown, crustaceous, outer bracts broadly ovate, 3.0 – 4.5 mm long, pilose on exposed part, hairs dense towards tip, veins and margins inconspicuous, tip acute, innermost bracts narrowly oblong, white-pilose above middle and on exposed inner face. Floral bracts filiform, subulate and densely white-pilose above middle. Receptacle hairs black, some hairs also on pedicels of female flowers. Flowers 3.0 – 3.5 mm long; sepals colourless. Male flowers: sepals free, narrowly oblong, scattered hairs on back, tips densely white-pilose, obtuse; petal-tube with slits at base, upper margin shallowly 3-lobed, lobes broadly rounded, white-ciliate; glands at midpoint of petals inside. Female flowers: sepals c. \( \raisebox{1ex}{$1$}\!\left/ \!\raisebox{-1ex}{$3$}\right. \) petals, membranous, narrowly elliptic-oblong, tips densely white-pilose, acute; petals free around ovary, connate above, villous outside and within with dense black hairs extending to middle or above from base and margins of free lower portion, hairs outside grading into white hairs along upper seams of petal-tube, hairs inside extending to level of glands c. \( \raisebox{1ex}{$1$}\!\left/ \!\raisebox{-1ex}{$3$}\right. \) up tube, tips shallowly rounded, densely white-ciliate. Seed subglobose, 0.6 – 0.8 mm long. Fig. 2A.
distribution. D.R. Congo (Katanga).
specimens examined. d. r. congo. Upemba National Park: Mitwaba, Mumbale, 20 Aug. 1953, Desenfans 4117 (BRLU [BRLU0020235]!) & Desenfans 4121 (BRLU [BRLU0020237]!); near source of the R. Sweba, 24 Aug. 1949, de Witte 7539 (BR!, WAG).
habitat. Ponds on iron pan pavement in savanna grassland; 1600 – 1800 m.
conservation status. Mesanthemum angustitepalum is a perennial herb restricted to pools on iron pan, and is known only from two locations in the Upemba National Park in Katanga, southeastern Democratic Republic of Congo. Upemba National Park is confronted with many threats ranging from poaching, illegal settlements and the presence of illegal permits to mining and hydroelectric interests (https://saveupemba.com/). However, the effects of these threats on M. angustitepalum, and the occurrence, number of locations and distribution of this species, remain unknown — with the last known collection of this species made in 1949 despite numerous surveys for rare plant species in Katanga connected with the mining industry in recent years. It is here assessed as Endangered (EN) B1ab(iii)+2ab(iii).
notes. Young specimens of Mesanthemum pilosum Kimp. are similar to M. angustitepalum, but the leaves and involucral bracts are usually less hairy, and the receptacle hairs are grey rather than black. At maturity the capitula of M. pilosum become globose with inturned involucral bracts, whilst those of M. angustitepalum remain hemispherical. Both species have villous female flowers, but the hairs arise differently. The sepals of M. angustitepalum are glabrous except for white hairs at the acute tips, whereas the sepals of M. pilosum are long-villous from the base and margins as well as having white hairs at the denticulate tips. The female petal-tube is villous inside in M. angustitepalum, but glabrous inside in M. pilosum.
Mesanthemum angustitepalum is contrasted in the protologue with M. jaegeri Jacq.-Fél., a West African species found among rocks and on inselbergs. It has large rosettes from a whitish, swollen base, with larger capitula (15 – 20 mm wide), female flowers with the sepals as long as the flower (not shorter), and the petal-tube is glabrous inside (not villous).
Mesanthemum auratum Lecomte (1908, publ. 1909: 599). Type: Guinea-Conakry, Friguiagbé à Bambaya, Dec. 1899, Pobéguin 30 (holotype P [P00443224]).
M. rubrum Moldenke, Phytologia 8: 390 (1962). Type: Guinea-Conakry, Mayon-Couré plantation, Friguiagbé, valley of Quatamba, 10 Feb. 1940, Chillou 1835 (holotype LL [LL00374648]).
Slender ephemeral. Leaves in a basal tuft, ensiform or narrowly lanceolate, 1 – 4.5 cm long, 1.2 – 4.0 mm wide, pilose, narrowed to a hard, subulate tip. Scapes erect, slender, 1 – c. 10, to 25 cm high, 0.2 – 0.3 mm thick, pilose with spreading, tubercle-based, colourless hairs. Sheaths much exceeding leaves, pilose, cylindrical in lower half, inflated above, deeply slit with acuminate limb. Capitulum 4 – 8 mm wide, bright golden-yellow, orange or white from copious fluffy hairs arising from an infundibular dark brown involucre. Involucral bracts in 3 – 4 series, lanceolate-oblong, crustaceous, dark brown with broad, pale, scarious margins, 2.5 – 3 mm long, villous with appressed scurfy white hairs on exposed surfaces, 5-veined, tip scarious, obtuse or inner bracts acute. Floral bracts filiform, plumose in upper half with golden-yellow, orange or white hairs 1.0 – 1.5 mm long, arranged in a single spreading circle within involucral bracts, plumose tips exserted from capitulum, floral disc exposed, inner flowers lacking plumose bracts. Receptacle villous, flowers embedded in colourless hairs. Flowers c. 1.5 mm, long raised on filiform pedicels of varying length; shorter filaments scattered among them. Male flowers: sepals absent; petals connate into an infundibular tube with free lobes above middle, lobes 0.3 – 1.0 mm long, oblong, yellow- or white-pilose on outer face, obtuse, a spongy linear penicillate gland on inner face; anthers 6, creamy-yellow or white; vestigial gynoecium present. Female flowers: sepals absent; petals free at base around ovary, connate above into a slender, cylindrical, glabrous tube expanded at apex into 3-lobed cup, lobes glabrous or yellow- or white-pilose on outer face, obtuse, a pale spongy penicillate gland on inner face. Seed elliptic-oblong in outline, c. 0.5 mm long, brown, indistinctly striate. Figs 3C & D, 9, 14.

Mesanthemum auratum. A habit; B capitulum; C outer involucral bract; D innermost involucral bract; E floral bract; F male flower; G inside view of male petal tube; H female flower; J inside view of female petal tube. From Chillou 675 & Cheek 18599. drawn by yuxi liang.
distribution. Guinea-Bissau, Guinea (Conakry), Sierra Leone.
specimens examined. guinea-bissau. Bijagós, Orango Grande, Raimundo & Guerra 931, 24 Jan. 1961 (K). guinea (conakry). Fouta Djalon, Dalaba, 18 Oct. 2016, Couch 830 (HNG, K). Kindia Prov.: before Mambia on Coyah to Kindia road, 2 Oct. 2015, Cheek 18231 (HNG, K); Kindia – Mamou road, not far beyond outskirts of Kindia, 3 Oct. 2015, Cheek 18256 (HNG, K); W of Mambia on road from Conakry, 2 Feb. 2016, Cheek 18307 (HNG, K); Mayon Khoure village, 30 Sept. 2016, Cheek 18599 (HNG, K); Kindia to Kiriya village, 1 Oct. 2016, Cheek 18636 (HNG, K); Friguiagbé, vallée du Samou, 25 Jan. 1938, Chillou 675 (K, WAG) & plantation du Mayon-Couré, 25 Jan. 1938, Chillou 676 (K); Kindia, Dec. 1956, Jacques-Felix 7451 (K, P); Mt Gangan, 28 Oct. 2011, Mesterházy MAGUI 224 (K) & 13 Oct. 1954, Schnell 7479 (K, P, WAG). sierra leone. Mt Loma, Nov. 1944, Jaeger 576 (K); Mt Bintumani, 26 Oct. 2012, Mesterházy MASIE 302 (K).
habitat. Sandstone outcrops with flowing water during the rainy season, flat sandstone bowal and on thin sandy soil over almost bare rock, with surface water 1 cm deep or less; up to 1000 (– 1900) m.
conservation status. Mesanthemum auratum as currently circumscribed is fairly widespread in West Africa. However, its estimated AOO is smaller than 2000 km2 which meets the threshold for Vulnerable; its number of locations nearly meets the threshold for Vulnerable (13 > 10); and continuing decline of area, extent and quality of habitat has been observed and/or inferred at various locations. Therefore, Mesanthemum auratum is here assessed as Near Threatened NT B2ab(iii). However, the taxonomic status of this species is not entirely resolved (see notes below).
notes. There appear to be two colour variants of this species. The extraordinary yellow-orange colour of the hairs on the floral bracts and flowers of some collections (including the type) is unique in Eriocaulaceae. Yellow-flowered specimens have only been collected from a restricted area around the town of Kindia in Guinea. Despite the striking colour of the flowering plants, this variant has not been collected since 1956 and recent searches of the area have failed to find it. There appear to be no significant morphological differences between this and the more widespread white-flowered form, which also grows in the same habitat around Kindia. Both colour forms have the same seed ornamentation (Fig. 9A – F).
According to the protologue (Lecomte 1909) Mesanthemum auratum flowers in profusion towards the end of the wet season, forming carpets, then completely disappears in the dry season. Material collected in early October is rather young, the male flowers unfurling their stamens, but the female flowers still immature. A plant with ripe seed was collected in late October. By early February all seed has been shed and no male flowers were present.
Mesanthemum albidum, several Utricularia species and Genlisea species occur in similar habitat. Collections of M. albidum are often mixtures with M. auratum. When white, the fluffy heads and delicate habit are very similar to those of M. albidum, but M. auratum can be distinguished by its obvious brown-black involucre of much tougher bracts at the base of the capitulum. Mesanthemum auratum lacks sepals in both the male and female flowers, whereas M. albidum lacks them only in the female flowers.
The conspicuous plumose floral bracts of Mesanthemum auratum are lacking in M. albidum, where the fluffy white hairs are borne on the flowers. However, the plumose bract-tips are fragile, easily breaking off and are absent in seeding capitula. Sometimes even most plumose floral bracts are absent, or only one or two are present. The fluffy appearance of well-developed capitula is then lacking, the dark brown involucre being the most obvious feature of the capitulum. This is the case with the specimens from Guinea-Bissau and Sierra Leone cited here and by Meikle (1968: 65), which have very sparse white-plumose floral bracts.
On well-developed specimens (e.g. Cheek 18636) the petals, although tiny in both flowers, are villous and provided with a penicillate gland on the inner face. However, frequently the petal lobes are scarcely developed and then are glabrous with the gland on the tip. The filaments found within the cushion of receptacular hairs were tentatively suggested to be sepals by Lecomte (caption to Fig. 1, p. 600, 1909). However, they are inserted on the receptacle separately from the flowers, and as they sometimes have a minute yellow spot at the tip, are more likely to be reduced floral bracts (although not obviously subtending flowers) or perhaps sterile pedicels. Insects (Coleoptera and Thysanoptera) were noted visiting the flowers of Wieringa 1139 (Gabon).
This species remains enigmatic, and possibly more than one species is included here. The variably reduced flowers, colour differences and often over-mature collections make comparisons difficult. The herbarium material available was not ideal for molecular study. The two specimens cited above from Sierra Leone were collected between 1600 – 1900 m, considerably higher than collections from Guinea. However, no substantive differences were found in the specimens examined, and the dark brown involucral bracts are distinctive.
Two collections from the Mone Forest Reserve in Cameroon may represent a new species close to Mesanthemum auratum (Cheek 13938 & 14121). They were collected from a granite inselberg in late November and early December, already dried up and dead. The plants have the typical dark brown bracts of M. auratum and remaining seed found in a capitulum has the same remarkable surface patterning (Fig. 9E, F).
Mesanthemum bennae Jacq.-Fél. (Jacques-Félix 1947: 145). Type: Guinea (Conakry), environs de Kindia, falaise du Benna, Oct. 1937, Jacques-Félix 2091 (lectotype P [P00443218, designated here]; isolectotypes K!, NY, WAG).
Robust tussocky perennial from a stout rootstock. Leaves ascending, ensiform or linear-elliptic and tapering to base, 20 – 48 cm long, 15 – 65 mm wide, fleshy becoming tough when dry, hirsute on both surfaces when young, old leaves glabrescent, margins firm, tapering to a hard acuminate-cuspidate tip. Scapes 2 – 6, erect, stout, 30 – 74 cm high, 1 – 2 mm thick, densely pilose with patent hairs, many-ribbed. Sheaths shorter than leaves, hirsute with patent hairs, limb hirsute within, tip hard, acuminate-cuspidate. Capitulum 20 – 45 mm wide, daisy-like, white becoming buff when old, involucre much exceedng floral disc. Involucral bracts in 5 or more irregular series of increasing length, indistinctly many-veined, outer bracts forming a shallow cup at capitulum base, 4.5 – 7 mm long, ovate, hirsute, cream-coloured or pale green, acuminate, inner series radiating ray-like much beyond floral disc, 9 – 20 mm long, lanceolate-oblong, papery, white-velutinous on both surfaces, acute. Floral bracts filiform, tips expanded, bearded with white hairs. Receptacle hairs grey or colourless. Flowers 3.5 – 3.8 mm long, pedicellate; sepals colourless. Male flowers: sepals free except at extreme base, narrowly oblong or oblanceolate, membranous, concave, tips pilose, obtuse; petal tube glabrous within, upper margin shallowly 3-lobed, white-pilose, glands dark brown with yellow tips; anthers cream-coloured. Female flowers: sepals free, narrowly oblong, membranous, concave, laterals slightly keeled, tips white-pilose; petals free in lower third, connate above ovary, petal-tube densely white-pilose outside towards tip, glabrous below or with a few longer colourless hairs or sometimes these denser, glabrous within. Figs 6C & D, 11A & B.
distribution. Known only from the Kounounkan area of SW Guinea.
specimens examined. guinea (conakry). Coyah Préfecture, above Fossikouré village, 8 Dec. 2017, van der Burgt 2185 (HNG, K!); Forécariah Préfecture, above Foret Classée de Kounounkan, southern part of Benna Plateau, 28 Nov. 2017, Konomou 311 (HNG, K!).
habitat. Cracks in vertical sandstone cliffs with seasonal water seepage; 360 – 1300 m.
conservation status. Mesanthemum bennae is the most spectacular species of the genus in flower with several brilliant white heads up to 4.5 cm in diam. It is restricted to sandstone cliffs on the Benna-Kounounkan Plateaus near Kindia in Guinea, and has been observed to be threatened by fires set artificially (X. van der Burgt pers. comm.). It is likely that the entire global population amounts to some thousands of plants, and is assessed as Endangered EN B1ab(iii)+2ab(iii).
typification. There are three specimens of Jacques-Félix 2091 in the Paris herbarium. The specimen with a handwritten label is designated here as lectotype.
notes. Mesanthemum bennae is a showy species, with its large white daisy-like heads from a broad-leaved basal rosette. It appears to have a very restricted distribution in Guinea, on vertical sandstone cliffs that are difficult to access. The only other species with similar showy daisy-like flowers is M. prescottianum, which also occurs in Guinea. This can be distinguished immediately by its much narrower, grassy leaves and almost glabrous long ray-like involucral bracts. Additionally, M. prescottianum is found on granite outcrops, not sandstone cliffs. Van der Burgt notes that M. bennae is not at all resistant to fire, never occurring in grassland that is burned annually.
Mesanthemum cupricola Kimp. (Kimpouni 1994: 151). Type: D.R. Congo, Katanga, Duvigneaud 4500E (holotype BRLU [BRLU000090021503]!).
Perennial from a thick tough rhizome. Leaves loosely tufted at rhizome tip, narrowly linear, up to 35 cm long, 3 – 7 mm wide, herbaceous, hirsute on both surfaces, bicoloured, adaxial surface reddish-brown and abaxial surface green, tip subacute, often with terminal hair tuft. Scapes 1 – 3, 30 – 50 cm high, 1.0 – 1.3 mm thick, pilose with scattered hairs. Sheaths hirsute when young, limb acuminate, tomentellous within. Capitulum 10 – 15 mm wide, dull white, flat-topped from a crateriform involucre, floral disc with blackish involucral hairs visible among white-hairy floral bracts, innermost involucral bracts extending 0.7 – 0.8 mm beyond floral disc. Involucral bracts in 4 – 5 series, brownish, cartilaginous, outer bracts broadly ovate with thinner ciliolate margins, 4 – 6 mm long, back patchily appressed silky-pilose, veins visible, tip acute to cuspidate, inner bracts narrower and longer, innermost narrowly oblanceolate, 5.2 – 6.5 mm long, acuminate-cuspidate. Floral bracts filiform, tip clavate, densely white-pilose. Receptacle hairs coarse, blackish, wiry. Flowers 2.5 – 3.4 mm long; sepals colourless. Male flowers: sepals connate at extreme base, oblanceolate-oblong, as long as flower, concave, back appressed-pilose towards tip, tip obtuse to truncate; petal-tube shallowly 3-lobed, white-pilose on upper back near tip. Female flowers: sepals \( \raisebox{1ex}{$1$}\!\left/ \!\raisebox{-1ex}{$3$}\right. \) length of petals, oblanceolate-oblong, tip rounded, erose, ciliate; petals variably connate into a fleshy cylindrical tube, free claws around ovary short or extending to middle, often hairy on upper claw margins, connate portion usually glabrous within, rarely villous with grey hairs from near base, free tips densely white-pilose on back, obtuse. Fig. 2B.
recognition. Mesanthemum cupricola is close to M. variabile, both being relatively slender species with narrowly linear leaves; flat-topped capitula; coarse blackish receptacle hairs; short broad sepals in the female flowers; and hairy margins to the claws of the petal-tube. However, they can be easily distinguished by the involucral bracts which are sharply pointed, extending shortly beyond the floral disc, and with longer shaggier hairs in M. cupricola. Mesanthemum variabile has more rounded involucral bracts with an obvious margin and a fine silky appressed pubescence.
Mesathemum cupricola is compared in the protologue to the common species M. radicans (Benth.) Körn. Mesanthemum radicans is a more robust, usually less hairy species with longer (up to 50 cm), wider (8 – 15 mm), scattered-pilose or glabrescent leaves. The involucral bracts are rounded to obtuse (not sharply pointed) with conspicuous veins towards the tips, and a shorter silky pubescence. The innermost bracts extend clearly 2 – 3 mm beyond the floral disc. The receptacle hairs in M. radicans are a paler grey colour, not blackish and wiry. M. radicans is a species of lowland marshes, not known above 1200 m.
distribution. D.R. Congo (Katanga).
specimens examined. d.r. congo. Katanga, Kolwezi, colline cuprifère de Dikuluwe, 10 Dec. 1959, Duvigneaud 4500E (holotype BRLU!) & Tilwizembe, 15 Dec. 1959, Duvigneaud 4580E (BRLU!).
habitat. Ponds on ironpan pavement and copper rich soils; c. 1400 – 1500 m.
conservation status. It is endemic to Katanga in southeastern Democratic Republic of Congo. It is only known from two collections made on heavy-metal polluted sites. The copperbelt area in the Democratic Republic of Congo is heavily impacted by open cast mining. This species has not been observed since 1959 at its only known location. Field surveys are needed to confirm whether the species is extant. Therefore, it is assessed as Critically Endangered CR B2ab(ii,iv) and possibly Extinct.
notes. Mesanthemum cupricola appears to be endemic to the copper rich soils of the copperbelt in southern D.R. Congo.
Some flowers from the type specimen show abnormalities. Apparently female flowers with a narrow cylindrical petal-tube and free basal claws were found to contain stamens when opened as well as an ovary, although the anthers may not be well developed. The number of anthers is variable: 3, 4 and 6 anthers were observed in three separate flowers. The sepals are longer, resembling those of the male flowers, but the ovary is larger than the vestigial ovary of typical male flowers. Typical male flowers always have an infundibular petal-tube containing six anthers, a vestigial ovary at the base, and sepals as long as the flower, as is usual in the genus. Additionally, the female petal-tube is normally glabrous within, but one flower was found which was densely villous inside. This character is usually stable within species and useful for identification. One flower was found with branching petal glands. These floral abnormalities and perhaps the bicoloured leaves may be due to the polluted habitat.
Mesanthemum glabrum Kimp. (Kimpouni 1994: 153); Phillips (2010: 73). Type: D.R. Congo, Kasai, by Kashidishi river, Kapanga, Oct. 1933, Overlaet 807 (holotype BR [BR855401]!).
Tufted perennial, occasionally developing a short rhizome. Leaves in a loose rosette, linear or subulate, up to 20 (– 40) cm long, 3 – 6 (– 8) mm wide, spongy, glabrous on lower surface, hirsute above, sometimes sparsely or glabrescent, margins incurling (at least when dry), tip thickened, obtuse. Scapes 1 – 3, slender, 40 – 75 cm high, 0.7 – 1.1 mm thick, glabrous to hirsute. Sheaths hirsute, often longer than the leaves, limb narrowed to a shortly rostrate obtuse tip. Capitulum 9 – 15 mm wide, bright white, flat topped with a turbinate involucre, floral disc densely white-pilose, involucral bracts not exceeding floral disc. Involucral bracts in 4 – 5 series, brownish-green with a broad pale scarious margin, strongly ribbed in upper half, smooth below, essentially glabrous (some short marginal cilia), outermost 3.7 – 5.0 mm long, ovate-oblong, inner progressively longer and more oblong, innermost 4.5 – 5.5 mm long, tips obtuse to broadly rounded. Floral bracts filiform, a dense brush of white hairs on expanded rounded tip. Receptacle hairs pale to dark grey-brown. Flowers 3 – 4 mm long; sepals colourless. Male flowers: sepals connate at extreme base, narrowly oblong, tips white-pilose, hairs eventually caducous. Female flowers: sepals resembling male sepals, as long as petals, glabrous on back, white-pilose at the truncate, denticulate tips; petals narrowly oblong, variably free up to about halfway, connate above, villous with long grey-brown hairs both outside and within up to level of glands (but margins glabrous), free tips densely white-pilose. Seed broadly ellipsoid, c. 0.9 mm long, dark brown. Fig. 2C.
distribution. Angola, Democratic Republic of Congo (Kasai), Zambia.
specimens examined. angola. Lazingua (Longa), 7 Jan. 1900, Baum 645 (K!); Benguella, received 1910, Gossweiler 2615, 2719, 3469 (all K!). Cuando Cubango Province: 4 km N Cuchi Mission, 120 km E Vila da Ponte, 12 Feb. 1952, H. Hess 52/669, 52/670, 52/671 (all K!, ZT); R. Senge, 120 km E Vila da Ponte, 24 June 1952, H. Hess 52/2059 (K!, ZT); Baixo Cubango, R. Cuevi, Caiundo area, 6 Feb. 1952, H. Hess 52/638 (K!, ZT); Baixo Cubango, 176 km von Cuangar in Richtung Caiundo am Rio cubango, 1070 m, 29 Jan. 1952, H. Hess 52/514 (K!); Baixo Cubango, 61 km von Caiundo in Richtung Cuangar an Rio cubango, 1100 m, 4 Feb. 1952, H. Hess 52/614 (K!); Longa R. (Cuito drainage), just north of rice project, 85 km E of Menongue, 14°33'22"S 18°24'25"E, 1320 m, 8 Feb. 2016, D. J. Goyder 8201 (K001333118!); Longa R. (Cuito drainage), 27 km NW of Longa village and 75 km ENE of Menongue, 14°24′09"S 18°20'11"E, 1334 m, 6 Nov. 2016, A. Frisby 3065 (K001333885!); NE Huila Province: 1 km E of Galange Mission, H. Hess 52/746 (K!, ZT); Moxico Province: Near head of lake at source of Cuito R., 62 km SSW of Munhango, 12°41'24"S 18°21'32"E, 1419 m, 22 May 2015, D. J. Goyder 8004 (K001333003!); R. Cuito source lake, c. 62 km SSW of Munhango, 12°41'22"S 18°21'34"E, 1353 m, 15 Feb. 2016, D. J. Goyder 8238 (K001333151!); Cuanavale R. source lake, c. 110 km SSE of Munhango, 13°05'22"S 18°53'37"E, 1353 m, 28 Feb. 2016, D. J. Goyder 8358 (K001333263!). d.r. congo. Kasai, by Kashidishi R., Kapanga, Oct. 1933, Overlaet 807 (BR!). zambia. Chama Distr., dambo of Manshya R., 28 Dec. 1963, Symoens 10781 (BR, BRLU, K!). Chinsali Distr., Ishiba Ngandu (Shiwa Ngandu), Lake Young, 15 Jan. 1959, Richards 10656 (K!). Mwinilunga Distr., Dobeka dambo, 54 km W of Mwinilunga, 22 Jan. 1975, Brummitt, Chisumpa & Polhill 13998 (K!).
habitat. Wet peaty soil of dambos and pools or marshy ground near rivers; 1100 – 1500 m.
conservation status. Mesanthemum glabrum occurs in at least 13 locations and has a very broad distribution with an EOO of 496,512 km2. Its known area of occupancy is small (60 km2) although it may have been undercollected. There are a number of continuing threats to its habitat associated with agriculture and housing development. However, recent collections (2015 – 2016) have been made in four locations where it was found to be frequent. One of these locations had recently undergone extensive agricultural development, meaning that this species is still locally common despite the change of land use. Since it is close to the threshold of ten locations for Vulnerable under Criterion B2, it has been assessed as Near Threatened (NT). There are planned hydroelectric developments (2015 – 2025) in Angola within this species’ distribution range, which might have future impacts on habitat and increase the threat level to this species. It is therefore suggested that seeds should be banked for ex situ conservation and the assessment is repeated in 5 years.
notes. This relatively slender species occurs on wet peaty soil at middle altitudes in central Africa. The heads have a top-shaped greenish involucre of neat, rather narrow, conspicuously pale-margined and prominently veined glabrous bracts, which support the bright white floral disc. The petal-tube of the female flowers is conspicuously villous with grey-brown hairs both outside and within.
Drummond & Cookson 6444 (W Zambia: Kalabo, 13 Nov. 1959) is a vigorous specimen matching Mesanthemum glabrum, but with dark brown coloration in the involucral bracts and a brush of long hairs from the pedicel-tip of the male flowers. Further collections are needed to determine its status.
Mesanthemum jaegeri Jacq.-Fél. (Jacques-Félix 1947: 146); Meikle (1968: 64); Phillips (1997: 34; 2011: 32; 2016: 10). Type: Sierra Leone, Jaeger 1625 (holotype P [P00135473]; isotype K!).
Robust perennial from a short, very stout rhizome clothed in decaying leaf remnants, base often bulbously swollen. Leaves linear to broadly lanceolate, 15 – 30 cm long, 7 – 25 mm wide, thick, hirsute at first, glabrescent, expanded bases papery and whitish, margins firm, tip acute. Scapes up to about 10, stout, 25 – 45 cm high, 1.6 – 2.2 mm thick, hirsute with spreading hairs. Sheaths about half as long as leaves, hirsute with spreading hairs, limb with hard acuminate tip. Capitulum 15 – 20 mm wide, flat-topped with a broad, shallow, cup-like involucre slightly shorter than capitulum width, white-hairy flowers and bracts protruding from a conspicuous cushion of black hairs. Involucral bracts in 3 – 4 series, pale brown, firmly membranous or finally coriaceous with broad hyaline margins, c. 5 mm long, ovate-oblong to broadly ovate, 5-veined, softly pilose on exposed portion, margins ciliate above middle, outer bracts apiculate, inner bracts acute, innermost not extending beyond floral disc. Floral bracts filiform, tip clavate, white-villous. Receptacle hairs wiry, black. Flowers c. 5 mm long; sepals pallid. Male flowers: sepals connate at extreme base, narrowly oblanceolate-oblong, back thinly pilose with scattered long hyaline hairs, tip obtusely triangular and densely pilose with stout white hairs; free petal-lobes densely white-pilose around margins. Female flowers: sepals longer than petals, slightly narrower than male sepals but otherwise similar; petals free near base, central connate portion glabrous outside, villous with long grey hairs within, free tips truncate, densely pilose with short stout white hairs. Seed plumply ellipsoid, c. 0.8 mm long, dark brown.
distribution. SW Cameroon, SE Nigeria, Ivory Coast, Sierra Leone.
specimens examined. cameroon. Kupe-Muanenguba, Muadelengoh rock, 23 Oct. 2001, Etuge 4434r (K!, MO, P, SCA, WAG, YA). ivory coast. Momi Mt, 29 Oct. 1966, Aké Assi 9145 (K!). nigeria. Ogoja Prov., Sonkwala area of Obudu Division, N side of Koloishe Mt, 21 Dec. 1948, Savory & Keay FHI25079 (K!). sierra leone. Loma Mts, Sérélen Konko, 20 Dec. 1965, Adam 22666 (G, K!, SL, WAG) & 10 Sept. 1945, Jaeger 1576 (K!); Sérélen Konko summit, 27 Sept. 1964, Jaeger 7655 (K!) & 20 Dec. 1965, Jaeger 8636 (K!); Sankan Biriwa summit, 5 Jan. 1960, Cole 155 (K!); Mt Loma, 2 Oct. 1964, Jaeger 7798 (K!); Tingi Mts, N Kono, 13 March 1966, Morton & Gledhill SL3148 (K!, SL, WAG).
habitat. Among rocks on mountains and damp rockfaces of inselbergs; (600 –) 1300 – 1500 m.
conservation status. Mesanthemum jaegeri is fairly widely distributed. However, based on herbarium label data of collections made in the past 80 years, the habitat is limited to mountainous regions (AOO = 44 km2), which form a small fraction of the EOO (957,614 km2). Applying Criterion B, the AOO is smaller than the thresholds for Vulnerable and Endangered, and the number of locations (8) falls within the threshold for Vulnerable. Furthermore, loss or degradation of habitat due to human activities is inferred. Therefore, the species is assessed as Vulnerable VU B2ab(iii).
notes. Mesanthemum jaegeri is a West African species found in wet rocky places in uplands. It usually has hairier leaves than M. radicans, which grows in marshy places at lower elevations. The bulbous plant base and conspicuous cushion of black wiry hairs in the capitulum are distinctive.
Mesanthemum pilosum Kimp. (Kimpouni 1994: 155); Phillips (2010: 74). Type: D.R. Congo, Kundelungu Plateau, 27 Oct. 1969, Lisowski, Malaisse & Symoens 7438 (lectotype BR [BR855368]!, designated here).
Robust tussocky perennial from a woody rootstock or shortly elongating rhizome, old burned leaf remains often present at base. Leaves clustered at rhizome branch tips, linear, grass-like, pale green, 15 – 30 cm long, 8 – 12 mm wide, usually pilose on both surfaces but varying from glabrescent or pubescent to velutinous, tip hard. Scapes solitary, 25 – 75 cm high, stout, 1.3 – 1.8 mm thick. Sheaths pilose, limb glabrous within. Capitulum 12 – 16 mm wide, densely white-hairy, hemispherical becoming subglobose at maturity, involucre flat across capitulum base at first, outer bracts extending almost to capitulum periphery, inner equalling floral disc, reflexing and inconspicuous at maturity. Involucral bracts in 4 – 5 series, brownish-green, coriaceous with narrow hyaline margins not extending round tip, outer bracts 4.5 – 6.0 mm long, ovate, glabrous or puberulous, veins visible, margins ciliate, tip subacute, inner more oblong, white-tomentose on inner face towards tip. Floral bracts filiform, tip subulate, white-villous, extending above flowers. Receptacle hairs pale to dark grey. Flowers 2.5 – 4.0 mm long; sepals brownish-grey. Male flowers: sepals free, thinly cartilaginous, narrowly oblong, tips white-pilose. Female flowers: surrounded by a dense circlet of long, soft, grey hairs from base of flower and sepal-bases; sepals half as long as petals, oblong or obovate-oblong, long-villous with straight colourless or pale grey hairs at base, some similar hairs also along margins, tips irregularly dentate, glabrous or few to many long marginal hairs; petals unequally free to middle or above, silky-villous near and along free margins outside, glabrous within, glands at midpoint inside, tips membranous, c. 0.5 mm, white-pilose or subglabrous. Seed elliptic-oblong in outline, c. 0.9 mm long, brown. Fig. 2D.
distribution. Angola, D.R. Congo (Katanga Prov.), Malawi, Zambia.
specimens examined. angola. Benguella, received 1910, Gossweiler 2205, 3037 (both K!); Central Mbaluku, near Quingenge, 28 Sept. 1959, Damann 58/717 (K!, ZT). d.r. congo. Kundelungu Plateau, 27 Oct. 1969, Lisowski, Malaisse & Symoens 7438 (BR!). malawi. Mzimba-Nkhata Bay Distr., Vipya Plateau, 51 km [32 ml] SW of Mzuzu, 11 Nov. 1973, Pawek 7488 (K!, MO, SRGH, UC); Nkhata Bay Distr., [10 ml] NE Chikangawa, 29 Oct. 1978, E. Phillips 4155 (K!, MO). zambia. Broken Hill [Kabwe] Distr., just beyond Mulunguishi R., 23 Sept. 1947, Brenan & Greenway 7938 (K!); Barotseland, 16 km [10 ml] N of Senanga, 31 July 1952, Codd 7297 (K!, PRE); Samfya, 2 Oct. 1953, Fanshawe 335 (K!); Kalabo, 15 Oct. 1963, Fanshawe 8088 (K!); Muchinga Prov., Mutinondo Wilderness Area, 30 Nov. 2013, Merrett 1696 (K!); Mwinilunga Distr., between Boma and Matonchi Farm, 28 Aug. 1930, Milne-Redhead 983 (K!) & SW of Dobeka Bridge, 13 Oct. 1937, Milne-Redhead 2748 (K!); Ntenke, Kawambwa, 10 Sept. 1963, Mutimushi 454 (K!); Kitwe, 24 Sept. 1968, Mutimushi 2716 (K!)
habitat. Grassland often subjected to burning, in wet boggy areas (dambos); 1000 – 1700 m.
conservation status. Mesanthemum pilosum is fairly widely distributed in Angola, the Democratic Republic of Congo (Katanga Province), Malawi, and Zambia (EOO = 887,045 km2 yet AOO is only 56 km2), where it is found in grassland often subjected to burning, and in wet boggy areas (dambos). Although the area, extent and/or quality of its habitat may be impacted by human activities and/or drought in some locations, its presence has been confirmed in at least two protected areas, and other known occurrences are near protected areas (particularly in Zambia). Targeted fieldwork will likely confirm its presence in the some of the latter. It is here assessed as Near Threatened (NT) since 13 locations are known, close to the threshold of 10 for Vulnerable B2a.
typification. The holotype was destroyed during transport from Belgium to Poland in the 1990s. An isotype is selected above as lectotype.
notes. Mesanthemum pilosum occurs across south-central Africa in wet patches in open grassland. The tussocky rosettes bear globose, white-hairy heads at maturity. The long circlet of silky hairs surrounding the disarticulated female flower is a good spot character, but female flowers are sometimes few. Damann (Angola s.n., 7 Nov. 1959 (ZT)) notes “first leaves appear on burned swampland in early September; plants in full flower by early October; most flowers finished by early November”.
Mesanthemum prescottianum (Bong.) Körn. (Körnicke 1863: 472, t.60 f.1); Durand & Schinz (1895: 504); N. E. Brown (1901: 261); Ruhland (1903: 118); Jacques-Félix (1947: 144); Meikle (1968: 327).
Eriocaulon prescottianum Bong. (Bongard 1831: 635). Type: Sierra Leone (“Brasilia”), com. Lindley s.n. (sphalm. Lindly), 1828 (LE lectotype [LE 00001102], selected by Phillips et al. (2018); K, isolectotype (K000346197)!).
Tufted perennial from slightly swollen base or with a small corm, sometimes producing small tubers on horizontal stolons; roots dense, dry, fibrous. Leaves up to 5, linear or rarely ensiform, up to 35 cm long, 2.6 – 6 mm wide, flat or lightly folded, outwardly curving when short, pilose especially on upper surface and margins towards base, tip acute. Scapes solitary or up to 4, slender to moderately stout, 25 – 50 (– 67) cm high, 0.5 – 1.0 mm thick, pilose. Sheaths shorter than leaves, pilose, limb acuminate, glabrous or sparsely pilose within. Capitulum daisy-like, shiny white, (17 –) 20 – 35 mm wide, with a shallowly cup-shaped involucre at base. Involucral bracts in several series of increasing length, outer bracts greenish straw-coloured, 2.5 – 7.0 mm long, lanceolate-oblong, coriaceous with thinner margins, pilose with soft dense hairs, subacute, innermost bracts shiny-white, radiating ray-like and much exceeding the floral disc, 9 – 18 mm long, narrowly lanceolate-oblong, thickly papery, outer (lower) surface glabrous or pubescent, strongly veined, veins appearing as furrows on upper surface, tips acute. Floral bracts filiform below middle, upper part fusiform, fleshy, glabrous, acute. Receptacle hairs usually colourless, occasionally pale brownish-grey, rarely blackish. Flowers 2.0 – 2.5 mm long; sepals usually colourless. Male flowers: sepals free, narrowly oblong, concave, glabrous, shiny, broadly obtuse or minutely denticulate; petal-tube densely villous along shallowly 3-lobed upper margin with colourless or infrequently grey hairs, also pilose within around the 3 linear brown glands; anthers dark brown or black. Female flowers: sepals resembling male sepals; petals free below middle, connate above, connate portion with a band of hairs outside from upper margins of free petal-bases, some hairs within around glands, upper margin shallowly 3-lobed, densely villous. Seed plumply ellipsoid, 0.55 mm long. Fig. 6E, F.
distribution. Guinea (Conakry), Ivory Coast, Liberia, Sierra Leone.
specimens examined. (Three selected from each country where more are available). guinea (conakry). Beyla Distr., Simandou Ridge, 8 Nov. 2005, Cheek 12665 (HNG, K!); Nzérékoré, Nimba Mts, near pumping station Zougué R., 24 July 2008, Jongkind 8265 (K!, WAG); Macenta Prefecture, Ziama Mts, Mt Wokou just S of Macenta, 24 Oct. 2008, Pearce 22 (HNG, K!, WAG). ivory coast. Mt Tonkoui, Aug. 1954, Schnell 6363 (K!, P); Nimba Mts, 10 Aug. 1954, Boughey G.C.18053 (K!). liberia. Central Province, Sanokwele Distr., near Sanokwele, 26 Sept. 1947, Baldwin 9516 (K!, M, NY, S). sierra leone. Makeni Hill, 26 Aug. 1951, Jordan 499 (K!); Tonkolili Distr., S Sula Mts, NE of Farangbaaya, 12 Sept. 2009, Kanu 6 (FBC, K!, SL); N of Makeni on Kalaba road, 21 Aug. 1967, Morton 4534 (K!, SL).
habitat. Thin peaty soil over exposed rock with seasonal seepages, especially in crevices on granite outcrops, often growing with grasses and species of Cyperaceae; 630 – 1600 m.
conservation status. Mesanthemum prescottianum is widespread and frequent over its area of occurrence and can therefore be considered as Least Concern (LC), although there is some threat from exploration for iron ore open-cast mining (Phillips et al. 2018).
notes. Mesanthemum prescottianum has a restricted distribution in the far west of Africa and is known only from thin soil over rock, to which it is very tightly affixed. It varies considerably in vigour, capitulum size, hairiness of the involucral bracts, and the colour of the hairs in the capitulum. However, it is always immediately recognisable by its shiny white daisy-like capitula with long, ray-like, usually pointed involucral bracts. The dark anthers of this species and M. tuberosum appear to be unique in the genus, other species having creamy-white or yellowish anthers.
Mesanthemum prescottianum flowers late in the wet season, reaching peak flowering in August and September and seeding in late October and November, the dried capitula persisting on the plant. It typically grows directly on rock in seasonally wet places, never in permanent standing water or marsh.
Mesanthemum pubescens (Lam.) Körn. (Körnicke 1856: 575); Jacques-Félix (1947: 148).
Eriocaulon pubescens Lam. (Lamarck 1789: 276). Type: Madagascar, Fort-Dauphin, Commerson s.n. (lectotype P-LAM [[P00563414], designated here).
M. rutenbergianum Körn. (Körnicke 1880: 34; Jacques-Félix (1947: 148)). Type: Madagascar, Ost-Amerina, Andrangoloaka, Nov. 1880, Hildebrandt 3714 (neotype K!, designated here; isoneotypes M, P), synon. nov.
Mesanthemum platyphyllum Baker (Baker 1883: 278). Type: Central Madagascar, Oct. 1882, Baron 1863 (K lectotype [K000346208]!, designated here; isolectotypes K [K000346207]!, P [P00102953]).
M. pubescens Körn. f. alba Jacq.-Fél. (Jacques-Félix 1947: 144, 149). Based on: Madagascar, without locality, Commerson s.n. (P-LAM) & Baron 458 (K).
M. pubescens Körn. f. nigra Jacq.-Fel. (Jacques-Félix 1947: 144, 149). Based on: Madagascar, Manankozo, Perrier de la Bathie 2245 [P01712831] & Decary 17200 [P01712832] (both P).
Robust tussocky perennial from a short stout rhizome. Leaves clustered at rhizome tip, linear to linear-lanceolate, up to c. 40 cm long, (4 –) 7 – 20 mm wide, thick, smooth, glossy, usually glabrous or with a few stiff scattered hairs, occasionally hispid, margins firm, tip hard, obtuse, sometimes shortly bearded in youngest leaves, hairs soon lost. Scapes up to c. 4, 50 – 65 cm high, 1.0 – 1.7 mm thick, wiry, glabrous or with scattered hairs. Sheaths glabrous to hispid, limb acuminate or attenuate with a few marginal cilia, sometimes shortly ciliate in the bottom angle. Capitulum white, 10 – 16 mm wide, shallowly hemispherical from a crateriform involucre. Involucral bracts in 3 – 5 series, yellowish-green, broadly ovate, 3.5 – 5.0 mm long, coriaceous with scarious margins, veins obvious, back minutely papillose, essentially glabrous or with scattered hairs, subacute to obtuse, inner bracts patchily appressed silky-pilose on thinner marginal tissue, margins ciliate, innermost bracts oblong, equalling or exceeding floral disc by up to 1 mm, densely appressed white-tomentose on exposed inner face. Floral bracts filiform, tip oblanceolate, white-tomentose. Receptacle hairs brownish-black. Flowers: 3.0 – 4.0 mm long, female flowers peripheral, subsessile; male flowers central, pedicellate; sepals brownish-black. Male flowers: sepals almost free, oblanceolate-angulate, densely white-pilose towards broadly obtuse tip, hairs spreading slightly inside. Female flowers: sepals \( \raisebox{1ex}{$3$}\!\left/ \!\raisebox{-1ex}{$4$}\right. \) as long to subequalling petals, narrowly oblong, broadly obtuse, glabrous to densely white-pilose around tip; petals oblanceolate, lightly connate from upper \( \raisebox{1ex}{$1$}\!\left/ \!\raisebox{-1ex}{$3$}\right. \) to subapically, easily separating, glabrous on outer face, lanate from lower \( \raisebox{1ex}{$1$}\!\left/ \!\raisebox{-1ex}{$3$}\right. \) inside, the woolly blackish hairs extending almost whole length of petals and protruding out of sides, upper margin membranous, truncate-crenate, white-pilose; 6 vestigial yellow stamens c. 0.5 mm long at base of ovary. Seed not seen. Figs 10, 11C.
distribution. Madagascar.
specimens examined. madagascar. Central Madagascar, Oct. 1882, Baron 1863 (K!, P!); Ost-Amerina, Andrangoloaka, Nov. 1880, Hildebrandt 3714 (K!, M); Ambatolampy, Feb. 1925, Perrier de la Bathie 16948 (K!).
habitat. Boggy stream margins and other marshy places on peaty soil over sand and quartz; 1100 – 1600 m.
conservation status. Mesanthemum pubescens and M. rutenbergianum (which we here place in synonymy of the first) have been assessed separately for the IUCN Red List (IUCN 2018). Both species have been assessed as near threatened (Rakotonirina & Faranirina 2017a, 2017b). Although the species experiences continuing decline due to threats including agriculture, mining & quarrying, nomadic grazing, wild fires, soil erosion and sedimentation, the combined distribution range and number of locations will unlikely qualify for any of the IUCN threat categories (IUCN 2012; IUCN Standards and Petitions Subcommittee 2017).
notes. The differences given by Moldenke (1955: 30) between Mesanthemum pubescens and M. rutenbergianum were not apparent in the specimens studied here. Jacques-Félix (1947: 144) distinguished these two mainly by leaf shape and pubescence, and whilst plants with narrower linear leaves do tend to be hairier, there are also narrow-leaved plants with glabrous leaves. Leaf width and pubescence appears to be variable, as it is in some other species of the genus. Therefore, they are combined here as the single species M. pubescens, which is endemic to Madagascar.
The drawing of Mesanthemum rutenbergianum in Ruhland (1903: 119) shows the female flowers with petals connate above the ovary in the upper two-thirds. This is an error, as the petals are loosely connate only towards the tip as described in his description, often being almost free with the dense curly hairs inside protruding through the slits. This is a very characteristic feature of M. pubescens. Vestigial stamens surrounding the base of the ovary in the female flowers have not been reported previously in Mesanthemum (Fig. 10). Apparently female flowers with a narrow petal-tube were also found to contain quite well-developed anthers in M. cupricola Kimp.
typification. The specimen in the Lamarck herbarium (P-LAM; microfiche LM-667/4) must be the basis of Lamarck’s protologue and is designated here as lectotype. Another specimen in Paris collected by Commerson (labelled “isotype” by Moldenke) consists of a single scape and sheath without any leaves [P00102948]. A further Commerson specimen from Herb. Jussieu [P00675322], comprises a whole plant.
The holotype of Mesanthemum rutenbergianum, collected in Madagascar by Rutenberg at Mahamba, near the Alaotra, on 18 Dec. 1877, appears to be lost. According to Dorr (1997: 418), Rutenberg was murdered in Madagascar, but some of his specimens were later returned to Bremen. Evidently his specimen forming the basis of this species was among them, as it was described by Körnicke who worked in Bonn. Soon after WWII many types in German herbaria were taken by Soviet troops and deposited in LE. These were later returned to Germany, but it has not proved possible to locate this particular type specimen. The specimen Hildebrandt 3714 (K) is a good match with the protologue, so is designated here as neotype.
The two forms of Mesanthemum pubescens, f. alba and f. nigra, described by Jacques-Félix are of no significance. He himself proposed them with some doubt, dependent on further collections. The specimen of Baron 458 (Kew), cited under f. alba, in fact has black (not whitish) sepals, and no specimens of this species have been seen with pale sepals. The names are included in the synonymy above for completeness only.
Mesanthemum radicans (Benth.) Körn. (Körnicke 1856: 573); N. E. Brown (1901: 260); Ruhland (1903: 119); Jacques-Felix (1947: 147); H. E. Hess (1955: 180); Meikle (1968: 64); Kimpouni et al. (1992: 134); Philllips (2010: 71; 2011: 31).
Eriocaulon radicans Benth. (Bentham 1849: 547). Type: Angola, without locality, Curror s.n. (lectotype K [K000346201]!, designated by Phillips in Fl. de Cameroun (2011)).
E. giganteum Afzel. ex Körn. (Körnicke 1856: 573), in synon.
E. guineense Steud. (Steudel 1855: 273). Type: Gabon, 1847, Jardin s.n.. (holotype P [P00443133]).
Mesanthemum erici-rosenii T.C.E.Fr. (in Fries 1916: 218); Jacques-Félix (1947: 148); Kimpouni et al. (1992: 134). Type: Zambia, L. Bangweulu (Bangweolo), Mbawala Is., Eric von Rosen 806 (holotype UPS!; isotype B [B100160928]).
Robust tussocky perennial from a stout, short or elongating rhizome, rhizomes horizontal below ground with branches turning vertically to surface. Leaves clustered at rhizome tip, light green, narrowly to broadly linear, 12 – 50 cm long, (8 –) 10 – 15 mm wide, slightly glossy, scattered-pilose to glabrescent, rarely hirsute with patent hairs, margins firm, tapering to an obtuse tip. Scapes 1 or 2, 30 – 60 cm high, 1.0 – 1.7 mm thick, tough. Sheaths usually pilose, limb elongate, tomentellous within. Capitulum 10 – 15 mm wide, creamy-white, flat-topped from a turbinate involucre, innermost involucral bracts extending 1 – 3 mm beyond floral disc. Involucral bracts in 5 – 6 series, straw-coloured with greenish conspicuously-veined tips, coriaceous with broad, thinner, pale margins, patchily appressed-sericeous, outer bracts 3.0 – 3.6 mm long, broadly ovate, rounded, obtuse or subacute, innermost oblong, appressed white-pilose on inner face, obtuse or subacute. Floral bracts capillary, tip subulate, white-pilose. Receptacle hairs pale to dark grey. Flowers 2.2 – 2.8 mm long; sepals colourless. Male flowers: sepals free or connate at the base, broadly oblong, back glabrous, tips truncate, white-ciliate; petal-tube with rounded, white-villous apical lobes. Female flowers: sepals half as long to subequalling petals, ovate-oblong, truncate-denticulate, glabrous or tips white-ciliate in a central tuft; petals free around ovary, connate in upper \( \raisebox{1ex}{$2$}\!\left/ \!\raisebox{-1ex}{$3$}\right. \), tube usually villous with long grey hairs outside and more densely within, occasionally only very sparsely hairy outside, free tips densely pilose with short stout white hairs. Figs 6A & B, 15.

Mesanthemum radicans. A in habitat; B capitulum from above; C capitulum from side. All from van der Burgt 1732. photos: xander van der burgt.
recognition. Mesanthemum radicans has flat-topped heads with a short fringe of protruding narrow involucral bracts around the periphery (Fig. 15), the heads appearing rather small in proportion to the size of the leaf rosette. Mesanthemum cupricola Kimp. from Zaire (Shaba Prov.) also has inner involucral bracts which exceed the floral disc, but differs from M. radicans by its more slender habit with narrower hairy leaves, acute outer bracts, and by the petals of the female flowers which are glabrous (not villous) on the inner face. Mesanthemum variabile also has ovate, appressed-sericeous involucral bracts, but differs by the broad pale margins and the conspicuously nerved tips of these bracts. On dissection, M. radicans can also be distinguished from M. variabile by the dense, long grey hairs inside the petal-tube of the female flowers.
distribution. Throughout West Africa, southwards to Angola, N Zambia and Mozambique, eastwards to W Uganda and W Tanzania.
specimens examined. (Two selected from each country where more are available). angola. Congo Prov., area of Cuango R. 5 km N of Posten Macocola, 12 April 1952, Hess 52/1289 (K!, ZT!); Lunda Norte, Lagoa Carumbo region, between the Luxico and Luele Rs N of Lagoa Carumbo, 7°45'18"S 19°57'19"E, 3 May 2011, Darbyshire 737 (K!). benin. Ouémé, Sakété, 6°43'N 2°39'E, 14 Aug. 1999, Adjakidjè 3111 (K!, WAG); Sémé, 20 km E Cotonou, Station des Eaux & Forêts, 27 Feb. 1965, Raynal 13543 (K!, P). cameroon. Hill Nkol Tsia, 18 km NW of Bipindi near Gouap, 12 Jan. 1974, Letouzey 12734 (K!, P); 10 km E of Kinsassa, 4 March 1971, Letouzey & Villiers 10495 (K!, P). central african republic. Manovo-Gounda-St Floris N.P., 300 m N of Camp Koumbela, meadow of Kpania creek, 8°29'N 21°12'E, 9 June 1982, Fay 3404 (K!); Ubangi (Haut Oubangui), R. Mbali, 45 km SW Wadda, 1 Dec. 1921, Le Testu 3513 (K!, P). congo (brazzaville). Kouilou, Pointe Noire-Pointe-Indienne, 4°41'36.9"S 11°48'55.7"E, 13 Sept. 2012, Kami 1404 (IEC, K!); ogooue-Leketi proposed N.P. near Simonbondo, 11 Oct. 2009, Moutsambote 6611 (IEC, K!, P, WAG). d.r. congo. Orientale Prov., Isangi Territory, Yambao and Mongandjo, R. Limbete, 15 Jan. 1957, Evrard 2106 (BR, K!); Kivu Prov., Kindu Territ., Kindu à Katako-Kombe, 12 Jan. 1960, Bamps 763 (BR, K!). equatorial guinea. A few km to the S of Bata, 22 March 1998, Porembski et al. 3628 (K!). gabon. Ogooué-Ivindo, Lopé Reserve, 0°07'S 11°37'E, 23 Aug. 1986, Blom LOPE 200 (K!, LBV); Ogooue-Maritime, Loango N.P., 1°55'23"S 9°18'41"E, 24 April 2005, Walters et al. 1544 (K!, MO). ghana. Near N Kroful, 24 Jan. 1970, Easterly 1216 (BGSU, CCG, K!); just beyond Essiana, 2 March 1953, Morton GC8498 (GC, K!). guinea (conakry). Boké, Boffa Prefecture, Bel-Air at Balandougou, 10°20′8"N 14°22'19"W, 22 Oct. 2016, Bidault et al. 2506 (K!, MO); 7 km from Kindia, 6 Sept. 1968, Kent 6 (K!). ivory coast. Savane d’ Abou-abou, between Abidjan and Gd Bassam, 7 Dec. 1963, Oldeman 731 (K!, WAG); Grand Bassam, 5°14'N 3°44'W, 29 Jan. 1968, Geerling & Bokdam 1962 (K!, WAG). liberia. New Univ. Site near Senya, Careysburgh, 35 km from Monrovia, 13 Sept. 1963, van Harten 85 (K!, WAG); Buchanan, 19 Nov. 1958, Adam 16006 (K!, P). malawi. N Prov., Chishimga Raneh near Luwingu, 27 April 1961, Astle 560 (K!, SRGH). mozambique. Beira Distr, Cheringoma Coast, Nyamaruza Dambo, between camp and road junction to Chiniziun Lighthouse, May 1973, Tinley 2915 (K!, SRGH). nigeria. Calabar Prov., Eket Distr., Ibeno, compound of Qua Ibo Mission School, 12 May 1953, Onochie FHI32099 (FHI, K!); Uyo Prov., Eket Distr., along footpath from Unyene to the Reserve, 15 Oct. 1964, Daramola FHI55317 (FHI, K!). senegal. Niokolo, Koba, June 1958, Adam 14198 (K!, P); Ziguinchor (Casamance), 4 Sept. 1962, Broadbent 92 (K!). sierra leone. Northern Prov., Port Loko Distr., Nyama, 8°42'9.2"N 13°7'2.3"W, 8 Nov. 2012, Lopez Poveda 185 (FBC, K!, SL); Tonkolli Distr., S Sula Mts, E of Bumbuna, 8°58'48"N 11°38'59"W, 27 Feb. 2010, Kanu 65 (FBC, K!, SL). tanzania. Bukoba Distr., Kagera, 1°19'S 31°48'E, 1 Nov. 1992, Breteler 11676 (K!, WAG) & Bukoba, June 1931, Haarer 2038 (K!); Mafia Is., near Kiombeni, 4 Oct. 1937, Greenway 5393 (K!). uganda. Masaka Distr., S of Luunga (Jubiya), 0°18'S 31°58'E, 31 May 1971, Katende 945 (K!, MHU) & Bukakata, 3 km [2 ml] from Bukakata old Port, 8 Dec. 1951, Norman 54 (K!). zambia. Northern Prov., Fort Rosebery [Mansa] Distr., Swamp mushitu near Samfya, Lake Bangweulu, 26 Aug. 1952, Angus 317 (K!, FHO).
habitat. Swampy grassland, lake and river margins and acid sand near the coast, sometimes growing in shallow water or periodically inundated places; sea level – 1500 m.
conservation status. Mesanthemum radicans is the most widely distributed species of Mesanthemum. Although the area, extent and/or quality of its habitat may be impacted by human activities and/or drought in some locations, due to its very extensive EOO (8,128,165 km2), it is here assessed as Least Concern (LC).
notes. Mesanthemum radicans is the most widespread species of Mesanthemum in Africa, but is apparently common only in West Africa and south to Angola. It is known in East Africa from the vicinity of Lake Victoria, Lake Bangweulu, and from coastal Tanzania and Mozambique. It appears to be common in the coastal wet savannas of Gabon. Very few female flowers were found in these populations. Insects (Coleoptera and Thysanoptera) were noted visiting the flowers of Wieringa 1139 (Gabon).
Mesanthemum reductum H.E.Hess (1955: 183, 179 Figs. 1 – 3). Type: Angola, H. Hess 52/2106 (holotype ZT!; isotypes BR [BR860335], K [K001089549]!, P [P00443054]).
Aquatic perennial from a slender elongating rhizome clothed in roots and fibrous leaf remnants; rhizome up to 45 cm long. Leaves clustered at rhizome tip, pale green, linear, 20 – 24 cm long, 5 – 7 mm wide, flaccid, membranous, translucent, glabrous, finely lattice-veined, tapering to an obtuse tip. Scapes 1 – 8, 40 – 55 cm high, 1.3 – 2.0 mm thick, glabrous. Sheaths glabrous, limb slightly inflated and flattened, acute or bifid. Capitulum 6.0 – 9.5 mm wide, involucre brown, cup-shaped, surrounding but not exceeding the paler floral disc. Involucral bracts in 3 – 5 series, hard, dark brown in upper half, often scurfy, with scarious, white, ciliolate margins, cream-coloured below, outermost bracts broadly ovate, 3 – 4 mm long, inner progressively narrower and longer, innermost oblong, c. 4.5 mm long, tip obtuse to rounded. Floral bracts absent. Receptacle hairs colourless. Flowers 1.5 – 2.0 mm long, embedded in the dense receptacular hairs on pedicels 2 – 3 mm long. Male flowers: (from protologue, none seen): sepals absent; petals connate into a campanulate hyaline tube, laciniate apex usually sparsely pilose, epipetalous glands absent; anthers 6, white, 0.3 – 0.4 mm long. Female flowers: sepals absent; petals free below middle around ovary, connate into a delicate narrow tube above, connate portion villous in lower part, hairs forming a penicillate tuft as long as and enclosing petal tube, this with 3 fine filamentous petal-tips at upper margin, epipetalous glands within absent. Seed not seen. Fig. 2E.
distribution. Angola.
specimens examined. angola. Cuando Cubango Province: 20 km E of Longa, Quiriri R., 29 June 1952, H. Hess 52/2106 (ZT type!, K!) & Longa R., 55 km SW of Cuito Cuanavale [Cuito drainage], 7 June 2015, Barker et al. 115 (K!).
habitat. Submerged in fast flowing rivers, forming patches and rooting in the sandy substrate; 1230 – 1290 m.
conservation status. Mesanthemum reductum is restricted in distribution to two locations on rivers on Kalahari sand in SE Angola. Further study is needed to better assess its distribution range and assess the threats faced by this species. Based on its very restricted distribution, it is here assessed as Near Threatened (NT) since no threats are known. Should threats be discovered the species should be re-assessed as Endangered EN B2ab.
notes. According to Hess (1955) the plants remain completely submerged, even the ripe capitula not appearing above the surface. The leaves are very thin and flaccid, with a fine cellular lattice structure visible when held against the light. No male flowers were found in the two collections examined.
Mesanthemum tuberosum Lecomte (1908 publ. 1909: 598). Type: Guinea (Conakry), Kouroussa to Timbo, June 1907, Pobéguin 734 (lectotype P [P00443181], selected by Phillips et al. 2018).
Perennial from a broad-based corm buried 2 cm or deeper, corm well developed, covered in brown fibres. Leaves few, 5 – 26 cm long, 2.4 – 10.0 mm wide, flat or lightly folded, longer leaves linear, shorter leaves narrowly lanceolate and outwardly curving from sheathing base, pilose with patent hairs especially towards base and on margins, tip hard, subacute. Scapes 1 – 3, up to 33 cm high, 0.9 – 1.2 mm thick, densely pilose with patent hairs. Sheaths pilose, opening abruptly into a flattened acuminate limb. Capitulum 12 – 25 (– 30) mm wide, dull white, flat topped from a crateriform involucre, inner bracts much longer than floral disc. Involucral bracts in several series of increasing length, outer bracts broadly lanceolate, green or brown, coriaceous in lower \( \raisebox{1ex}{$1$}\!\left/ \!\raisebox{-1ex}{$3$}\right. \), upper part and margins herbaceous, 5 – 7 mm long, softly pilose, abruptly narrowed above, inner bracts progressively more caudate, becoming broadly linear, fleshy, dull white, 3 – 5-veined, obtuse or subacute, extending ray-like beyond floral disc, innermost bracts shorter and grading into floral bracts, all except outermost subtending flowers. Floral bracts broadly linear to clavate from a slender base, fleshy, white, glabrous, obtuse or subacute. Receptacle with flowers and slender bases of floral bracts embedded in dense cushion of grey hairs, fleshy white tips of floral bracts protruding above. Flowers 2.5 – 3.5 mm; sepals colourless. Male flowers: sepals unequally connate at base, elliptic-oblong to obovate, concave, glabrous, obtuse; petal-tube shallowly 3-lobed, upper margin densely villous with grey-brown hairs; anthers dark brown. Female flowers: sepals resembling male sepals; petals free in lower \( \raisebox{1ex}{$1$}\!\left/ \!\raisebox{-1ex}{$3$}\right. \), connate above, villous in 3 longitudinal bands along seams of upper connate portion, glabrous inside, upper margin shallowly 3-lobed and densely villous with grey-brown hairs. Seed ellipsoid, 0.6 mm long. Fig. 6G, H.
distribution. Guinea (Conakry) in the Fouta Djallon highlands.
specimens examined. guinea (conakry). Fouta Djallon: Dalaba, 10°37'41.9"N 12°16'46.2"W, 10 June 2016, Cheek 18489 (K!) & near Forestry House, 21 June 1962, Adames 273 (K!); Labe, between Labe and Pita on the road c. 15 km from Pita, 11°10'47.9"N 12°22'51.3"W, 12 June 2016, Konig 30 (K!, HNG); 5 km S of Sériba, 11°4'N 3°1'W, 28 July 1992, Porembski 44 (K!); 40 km W of Labé, 11°2'N 2°4'W, 29 July 1992, Porembski 64 (K!); 20 km N of Dalaba, 10°4'N 12°2'W, 30 July 1992, Porembski 84 (K!); 20 km S of Mamou, 10°1'N 12°3'W, 31 July 1992, Porembski 97 (K!); Mamou, Sept. 1954, Schnell 6789 (K!); Mali, Sept. 1954, Schnell 7063 (K!).
habitat. Sandstone bowal with stepped horizontal flats covered in sandy soil; 140 – 1500 m.
conservation status. Mesanthemum tuberosum is a localised species endemic to the high altitude seasonally wet sandstone bowal grassland of the Fouta Djallon highlands of Guinea. It bears flowers in June in the wet season, and is unusual in having a fleshy basal corm buried in its sandy habitat. It is known from 9 locations, although some of these have not been verified for a number of years. Its habitat is impacted at several locations by cattle grazing and trampling and by modification of habitat for cultivation (Cheek pers. obs. June 2016; Phillips et al. 2018). There is also the threat of mining for bauxite. Based on current evidence and the lack of any protection for this species, it is assessed as Vulnerable VU B1ab(iii)+2ab(iii) .
notes. Mesanthemum tuberosum is closely related to M. prescottianum, confined to the Fouta Djallon area of Guinea. It occupies a different habitat from M. prescottianum, on sandstone flats rather than on rocky outcrops, and also flowers earlier at the start of the wet season (June – July). In dissected specimens the female flowers were few in the axils of the involucral bracts, flowers of the central floral disc being apparently all male.
Mesanthemum variabile Kimp. (Kimpouni 1994: 157); Phillips (2010: 73). Type: D.R. Congo, Manika Plateau, 3 km W of Katema, 19 Jan. 1969, Lisowski, Malaisse & Symoens 423 (lectotype BR [BR8603326], designated here).
Perennial from a thick tough rhizome. Leaves in erect tuft at rhizome tip, narrowly linear, 13 – 30 cm long, 4 – 8 mm wide, tapering to base and apex, glabrous with marginal setae or pubescent to hirsute especially on upper surface, tip subacute. Scapes 1 – 3, 40 – 70 cm high, c. 1 mm thick, glabrous or pubescent. Sheaths glabrous or pilose, limb long acuminate. Capitulum 13 – 15 mm wide, white, flat-topped from a crateriform involucre, black involucral hairs visible in floral disc, innermost involucral bracts very slightly longer (< 0.8 mm) than floral disc. Involucral bracts in 5 series, buff-coloured, cartilaginous, outer bracts broadly ovate, 2.5 – 4.0 mm long, veins obscure, margins membranous, ciliolate, tip subacute, inner bracts up to 5.5 mm long, sometimes obtuse, all patchily appressed-sericeous on back, innermost sometimes also on inner face. Floral bracts filiform, tip clavate, white-pilose. Receptacle hairs black, coarse, straight, wiry. Flowers 2.7 – 3.5 mm long; sepals colourless. Male flowers: sepals ± free, oblanceolate-oblong, white-pilose on upper back and rounded tips; petal-tube shallowly 3-lobed, free lobes white-pilose on back at tip. Female flowers: sepals ½ – ¾ length of petals, obovate-oblong to narrowly oblong, tips truncate-dentate with a few white hairs on teeth; petals free in lower \( \raisebox{1ex}{$1$}\!\left/ \!\raisebox{-1ex}{$4$}\right. \) – \( \raisebox{1ex}{$1$}\!\left/ \!\raisebox{-1ex}{$3$}\right. \), connate into a spongy tube above, white-pilose towards subtruncate tip or from middle upwards, villous with long, grey hairs arising near margins at top of free claws outside, a few marginal hairs sometimes also projecting inside petal-tube, otherwise glabrous within. Fig. 2F.
recognition. Mesanthemum variabile has sericeous involucral bracts like M. radicans, but is a more slender species with narrower leaves and an involucre scarcely extending beyond the floral disc. The capitula are similar to those of M. glabrum, but are slightly flatter, with a cup-shaped rather than conical involucre when young, and coarse black receptacular hairs. A glabrous interior to the female petal-tube will confirm the identification.
distribution. D.R. Congo (Kasai, Katanga), NE & NW Zambia.
specimens examined. d.r. congo. Haut-Katanga, towards the Poto Poto, 19 Dec. 1974, Malaisse 8752 (BR!). zambia. Abercorn [Mbala] Distr., Mpukutu Forest, Chinakila, 14 Jan. 1965, Richards 19522 (K!); Kawambwa Distr., Masenga R., 18 April 1957, Richards 9251 (K!) & M’tunatusha R., 28 Nov. 1961, Richards 15412 (BR; K!); Mporokoso Distr., Mporokoso – Sanga Hill road, 19 Jan. 1960, Richards 12427 (K!); Mwinilunga Distr., Zambesi rapids, 6 km N of Kalene Hill, 21 Feb. 1975, Hooper & Townsend 296 (K!) & 18 km E of Kalene Hill, 16 Dec. 1963, Robinson 6116 (K!); Northern Prov., Chishinga Ranch near Luwingu, 27 April 1961, Astle 560 (K!).
habitat. Wet hollows and marshy grassland near rivers on sandy alluvium; 1000 – 1600 m.
conservation status. Mesanthemum variabile is found in areas of D.R. Congo and Zambia that are heavily impacted by mining activities. The AOO has been estimated as 52 km2 which meets the threshold for Vulnerable and Endangered under criterion B, and the number of locations (8) meets the threshold for Vulnerable. Due to open cast mining, decline of area, extent and/or quality of habitat has been observed, and decline of AOO, EOO and number of locations are inferred. Therefore, Mesanthemum variabile is assessed as Vulnerable VU B2ab(i,ii,iii,iv).
typification. The holotype was destroyed during transport from Belgium to Poland in the 1990s. An isotype is selected above as lectotype.
References
Andrade, M. J. G. de, Giulietti, A. M., Rapini, A., Queiroz, L. P. de, Conceição, A. D. S., Almeida, P. R. M. de & van den Berg, C. (2010). A comprehensive phylogenetic analysis of Eriocaulaceae: Evidence from nuclear (ITS) and plastid (psbA-trnH and trnL-F) DNA sequences. Taxon 59: 379 – 388.
Ansari, R. & Balakrishnan, N. P. (1994). The Family Eriocaulaceae in India. Dehra Dun.
Bachman, S., Moat, J., Hill, A., de la Torre, J. & Scott, B. (2011). Supporting Red List threat assessments with GeoCAT: Geospatial Conservation Assessment Tool. ZooKeys 150: 117 – 126.
Baker, J. G. (1883). Contributions to the Flora of Madagascar III. J. Linn. Soc., Bot. 20: 237 – 358.
Barreto, L. C., Echternacht, L. & Garcia, Q. S. (2013). Seed coat sculpture in Comanthera (Eriocaulaceae) and its implications on taxonomy and phylogenetics. Pl. Syst. Evol. 299: 1461 – 1469.
Bentham, G. (1849). Flora Nigritiana. In: W. J. Hooker (ed.), Niger Flora, pp. 203 – 577. J. Cramer, Lehre.
Bongard, M. (1831). Essai monographique sur les espèces d'Ériocaulon du Brésil. Mém. Acad. Imp. Sci. St. Pétersbourg, Sér. 6. Sci. Math. 1: 601 – 655.
Brown, N. E. (1901). Eriocaulaceae. In: W. T. Thiselton-Dyer (ed.), Flora of Tropical Africa Vol. 8: 230 – 264. Lovell Reeve & Co, London.
Darbyshire, I., Timberlake, J., Mapaura, A., Chelene, I. & Hadj-Hammou, J. (2017). Mesanthemum africanum. The IUCN Red List of Threatened Species 2017: e.T66106115A66106124. doi: https://doi.org/10.2305/IUCN.UK.2017-1.RLTS.T66106115A66106124.en. Downloaded on 16 November 2018.
Dokkedal, A. L., Campaner dos Santos, L., Takeo Sano, P. & Vilegas, W. (2008). Chemistry in Eriocaulaceae. Z. Naturf. C 63: 169 – 175.
Dorr, L. (1997). Plant Collectors in Madagascar and the Comoro Islands. Royal Botanic Gardens, Kew.
Doyle, J. J. & Doyle, J. L. (1987). A Rapid DNA Isolation Procedure for Small Quantities of Fresh Leaf Tissue. Phytochem. Bull. 19: 11 – 15.
Durand, T. & Schinz, H. (1895). Conspectus Florae Africae 5. Jardin Botanique de l'Etat, Bruxelles.
Fries, R. E. (1916). Wissenschaftliche Ergebnisse der Schwedischen Rhodesia-Kongo-Expedition, 1911 – 1912. Bd. 1, Botanische Untersuchungen. Stockholm.
Giulietti, A. M., Monteiro, W. R., Mayo, S. J. & Stephens, J. (1988). A Preliminary survey of testa sculpture in Eriocaulaceae. Beitr. Biol. Pflanzen 62: 189 – 209.
Giulietti, A. M., Amaral, M. C. E. & Bittrich, V. (1995). Phylogenetic analysis of inter-and infrageneric relationships of Leiothrix Ruhland (Eriocaulaceae). Kew Bull. 50: 55 – 71.
Hess, H. E. (1955). Zur Kenntnis der Eriocaulaceae von Angola und dem unteren Belgischen Kongo. Ber. Schweiz. Bot. Ges. 65: 115 – 204.
IUCN (2012). IUCN red list categories and criteria: version 3.1, Vol 4, 2nd ed. IUCN, Gland.
IUCN (2018). The IUCN Red List of Threatened Species. Version 2018-2. http://www.iucnredlist.org/ Downloaded on 14 November 2018.
IUCN Standards and Petitions Subcommittee. (2017). Guidelines for Using the IUCN Red List Categories and Criteria. Version 13. Prepared by the Standards and Petitions Subcommittee. Downloadable from http://www.iucnredlist.org/documents/RedListGuidelines.pdf. Dowloaded on 23 May 2018.
Jacques-Félix, H. (1947). Le genre Mesanthemum (Eriocaulaceae). Bull. Soc. Bot. France 94: 143 – 151.
Katoh, K., Asimenos, G. & Toh, H. (2009). Multiple alignment of DNA sequences with MAFFT. Methods Molec. Biol. 537: 39 – 64.
Katoh, K. & Standley, D. M. (2013). MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Molec. Biol. Evol. 30: 772 – 780.
Kearse, M., Moir, R., Wilson, A., Stones-Havas, S., Cheung, M., Sturrock, S., Buxton, S., Cooper, A., Markowitz, S., Duran, C., Thierer, T., Ashton, B., Meintjes, P. & Drummond, A. (2012). Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28: 1647 – 1649.
Kimpouni, V. (1994). Nouvelles espèces africaines du genre Mesanthemum (Eriocaulaceae). Fragm. Florist. Geobot. 39: 147 – 160.
Kimpouni, V., Lejoly, J. & Lisowski, S. (1992). Les Eriocaulaceae du Congo. Fragm. Florist. Geobot. 37: 127 – 145.
Körnicke, F. (1856). Eriocaulacearum monographie supplementum. Linnaea 27: 561 – 692.
Körnicke, F. (1863). Eriocaulaceae. In: C. F. P. H. von Martius & A. W. Eichler (eds), Flora Brasiliensis, Vol. 3 (1): 274 – 507. Typographia regia, Monachii.
Körnicke, F. (1880). Reliquiae Rutenbergianae. Eriocaulaceen. Abh. Naturwiss. Vereine Bremen 7: 34 – 36.
Lamarck, J. B. A. P. (1789). Encyclopedie Méthodique: botanique 3. Paris.
Lanfear, R., Calcott, B., Ho, S. Y. & Guindon, S. (2012). PartitionFinder: combined selection of partitioning schemes and substitution models for phylogenetic analyses. Molec. Biol. Evol. 29: 1695 – 1701.
Larridon, I., Tanaka, N., Liang, Y., Phillips, S. M., Barfod, A., Cho, S.-H., Gale, S., Jobson, R. W., Kim, Y.-D., Li, J., Muasya, A. M., Parnell, J. A. N., Prajaksood, A., Shuto, K., Souladeth, P., Tagane, S., Tanaka, N., Yano, O., Mesterhazy, A., Newman, M. & Ito, Y. (2019). First molecular phylogenetic insights in the evolution of Eriocaulon (Eriocaulaceae, Poales). J. Pl. Res. 132: 589 – 600.
Lecomte, H. (1908, publ. 1909). Ériocaulacées d’Afrique. Bull. Soc. Bot. France 55: 594 – 602.
Meikle, R. D. (1968). Eriocaulaceae. In: F. N. Hepper (ed.), Flora of West Tropical Africa, Ed. 2, 3: 57 – 67. Crown Agents, London.
Miller, M. A., Pfeiffer, W. & Schwartz, T. (2010). Creating the CIPRES Science Gateway for inference of large phylogenetic trees. Gateway Computing Environments Workshop (GCE), pp. 1 – 8.
Moldenke, H. (1949). Notes on new and noteworthy plants VIII. Phytologia 3: 106 – 121.
Moldenke, H. (1955). Flore de Madagascar et des Comores, 36 Famille, Eriocaulacées. Typ. Firmin-Didot, Paris.
Moldenke, H. (1962). Notes on new and noteworthy plants XXXII. Phytologia 8: 385 – 395.
Müller, J., Müller, K., Neinhuis, C. & Quandt, D. (2010). PhyDE–Phylogenetic Data Editor, version 0.9971. http://www.phyde.de/.
Phillips, S. M. (1994). Notes on some Eriocaulon species from Ceylon. Kew Bull. 49: 287 – 303.
Phillips, S. M. (1997). Eriocaulaceae. In: R. M. Polholl (ed.), Flora of Tropical East Africa. Royal Botanic Gardens, Kew.
Phillips, S. M. (2010). Mesanthemum. In: J. R. Timberlake & E. S. Martins (eds), Flora Zambesiaca 13 (4): 70 – 75. Royal Botanic Gardens, Kew.
Phillips, S. M. (2011). Eriocaulaceae. Flore du Cameroun 38. Ministry of Scientific Research and Innovation, Yaoundé.
Phillips, S. M. (2016). Eriocaulaceae. In: Flora du Gabon Vol. 48: 1 – 19. Margraf, Weikersheim.
Phillips, S. M., Fofana, F. & Cheek, M. (2018). Mesanthemum tuberosum Lecomte resurrected from M. prescottianum (Bong.) Körn. (Eriocaulaceae), variation and lectotypification. Kew Bull. 73: 13 (8 pp.). doi: https://doi.org/10.1007/S12225-018-9744-7.
Rakotonirina, N. & Faranirina, L. (2017a). Mesanthemum pubescens. The IUCN Red List of Threatened Species 2017: e.T88514892A88514899. doi: https://doi.org/10.2305/IUCN.UK.2017-2.RLTS.T88514892A88514899.en. Downloaded on 16 November 2018.
Rakotonirina, N. & Faranirina, L. (2017b). Mesanthemum rutenbergianum. The IUCN Red List of Threatened Species 2017: e.T88514905A88514908. doi: https://doi.org/10.2305/IUCN.UK.2017-2.RLTS.T88514905A88514908.en. Downloaded on 16 November 2018.
Ronquist, F., Teslenko, M., Van Der Mark, P., Ayres, D. L., Darling, A., Höhna, S., Larget, B., Liu, L., Suchard, M. A. & Huelsenbeck, J. P. (2012). MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 61: 539 – 542.
Ruhland, W. (1903). Eriocaulaceae. In: A. Engler (ed.), Das Pflanzenreich Vol. 13: 1 – 294 (IV – 30). Wilhelm Engelmann, Leipzig.
Stamatakis, A. (2014). RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30: 1312 – 1313.
Steudel, E. G. (1855). Synopsis Plantarum Glumacearum. II. J. B. Metzler, Stuttgart.
Stützel, T. (1985). Die Bedeutung monothecat-bisporangiater Antheren als systematisches Merkmal zur Gliederung der Eriocaulaceen. Bot. Jahrb. Syst. 105: 433 – 438.
Stützel, T. (1998). Eriocaulaceae. In: K. Kubitzki (ed.), The Families and Genera of Vascular Plants, Vol. IV: 197 – 211. Springer, Berlin, Heidelberg.
Thiers, B. (continuously updated). Index Herbariorum: A global directory of public herbaria and associated staff. New York Botanical Garden's Virtual Herbarium. http://sweetgum.nybg.org/science/ih/. Accessed 22 sept 2018.
WCSP (2018). World Checklist of Selected Plant Families. Facilitated by the Royal Botanic Gardens, Kew. Published on the Internet. http://wcsp.science.kew.org/. Accessed 19 Oct 2018.
Unwin, M. M. (2004). Molecular systematics of the Eriocaulaceae Martinov. Doctoral dissertation, Miami University.
Zhang, Z. (1999). Monographie der Gattung Eriocaulon in Ostasien. Dissertationes Botanicae Band 313. Cramer, Stuttgart.
Acknowledgements
We thank Laszlo Csiba for laboratory assistance and Chrissie Prychid for help with the electron microscope at Kew. We also thank the curators of the herbaria in Paris (P), Brussels (BR & BRLU) and Zurich (ZT) for the loan of type material and other specimens. Véronique Durand-Andro is thanked for supplying a high-resolution image of a type from the Lamarck herbarium (P-LAM). Bart Wursten and Xander van der Burgt gave permission for use of their images. The research of IL is supported by the B. A. Krukoff Fund for the Study of African Botany. YL completed her part in this manuscript as part of the requirements for the MSc in Plant and Fungal Taxonomy, Diversity and Conservation at Royal Botanic Gardens, Kew and Queen Mary University of London.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
ESM 1.
Supplementary data file of the concatenated alignment (matK, PHYC, rbcL, rpoB, rpoC) (FAS 82 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Liang, Y., Phillips, S.M., Cheek, M. et al. A revision of the African genus Mesanthemum (Eriocaulaceae). Kew Bull 74, 71 (2019). https://doi.org/10.1007/s12225-019-9853-y
Accepted:
Published:
DOI: https://doi.org/10.1007/s12225-019-9853-y