
Abstract
In this work we investigated the ability of four extremophilic bacteria from Archaea and Bacteria domains to resist to space environment by exposing them to extreme conditions of temperature, UV radiation, desiccation coupled to low pressure generated in a Mars’ conditions simulator. All the investigated extremophilic strains (namely Sulfolobus solfataricus, Haloterrigena hispanica, Thermotoga neapolitana and Geobacillus thermantarcticus) showed a good resistance to the simulation of the temperature variation in the space; on the other hand irradiation with UV at 254 nm affected only slightly the growth of H. hispanica, G. thermantarcticus and S. solfataricus; finally exposition to Mars simulated condition showed that H. hispanica and G. thermantarcticus were resistant to desiccation and low pressure.
Similar content being viewed by others

Introduction
Extremophiles are a group of microorganisms that belong to all three domains of life (Archaea, Bacteria and Eukarya) and that are found on the Earth in habitats considered extreme from an anthropocentric point of view. Such environments are characterized by harsh conditions of temperature, pH, salinity, pressure and radiation and they include hydrothermal vents and springs, the so called “black smokers”, nuclear reactors, acid mine drainages and rivers, permafrost sediments, sea ice and polar lakes, highly polluted soils, et cetera. The study of extremophiles microbiology is therefore relevant to the definition of the boundaries of life, one of main issues of astrobiology. Indeed astrobiology investigates the origin, evolution, distribution and future of life in the universe (Des Marais et al. 2008) that, according to the “panspermia” theory, was originated from microorganisms and simple cells transported from one planet to another and able to adapt to adverse conditions in space. In this frame studying microbial communities that are adapted to live on earth in very extreme conditions, and establishing the limits to life on our own planet, could help in getting insights into the potential of other worlds to support life, and thus determining whether life as we know it could exist elsewhere in the galaxy. (Saffary et al. 2002).
In this work we investigated the ability of some extremophilic species to survive and to resist to some stressing conditions simulating the space environment. For our study we chose four extremophilic species from different extreme environment and representing both Archaea and Bacteria domains: Sulfolobus solfataricus (Zilling et al. 1980) a thermo-acidophilic archeon isolated from Solfatara volcano, southern Italy; Haloterrigena hispanica (Romano et al. 2007) an extremely halophilic archaeon from Fuente de Piedra, southern Spain; Thermotoga neapolitana (Jannash et al. 1988) a thermo-anaerobic microorganism isolated from a black smoker in the bay of Naples (Italy) and finally Geobacillus thermantarcticus (Coorevits et al. 2012; Lama et al. 2004) a thermophilic microorganism isolated from geothermal soil from Mt. Melbourne, an active volcano in Antarctica. To assess their possible ability to survive in space environment, all the selected species were exposed to extreme conditions in terms of temperature, UV radiation, humidity and pressure resembling Mars conditions and that were assayed in separately or in combination (Onofri et al. 2008). Three different sets of conditions were assayed: storage at extreme temperatures, UV resistance (carried out separately and both in terrestrial atmosphere and pressure conditions), resistance to martian humidity and pressure (Nicholson et al. 2013) investigated at the same time in a Mars simulator set at terrestrial temperature and radiation conditions.
Experimental
Microorganisms culture conditions: Sulfolobus solfataricus (Zilling et al. 1980) (optimal T = 85 °C), Thermotoga neapolitana (Jannash et al. 1988) (optimal T = 80 °C), Haloterrigena hispanica type strain FP1T (Romano et al. 2007) (optimal T = 50 °C), Geobacillus thermantarcticus, initially named Bacillus thermantarcticus (Lama et al. 2004) (optimal T = 60 °C) were grown in the respective standard conditions as previously described.
Effect of Temperature Stress on Microbial Growth Kinetic
1.5 ml of each cell culture (with an O.D540 value of 0.600) were centrifuged at 10,000 rpm for 10 min and the resulting cell pellets were stored at −196 °C (by liquid N2), −80, −20, 0, 4, 10, 25, 50, 70, 85 °C for 1 week (in the case of S. solfataricus) and 3 months for all other species. After these treatments, cell pellets were recovered and then used as inoculum in the respective standard growth media to assess cell survival and starting O.D.540 was 0.010.
Effect of UV Radiation (λ = 254 nm) on Microbial Growth Kinetic
Experiments were carried out at terrestrial temperature (23 °C), pressure, and atmosphere conditions by using a bactericidal wavelength. In detail, 5 ml of each cell culture (with an O.D540 value of 0.600) were put in an open Petri dish that were exposed at room temperature to a UV-lamp set perpendicularly to the uncovered dishes at a distance of 7 cm. The lamp was set at λ = 254 nm with an irradiation power of about 6.00 W/m2 that was applied for 7.5, 15, 30 min and 1 h. Control experiments were carried out in the absence of UV radiation. After these treatments, cells were recovered after centrifugation of the liquid medium deposited in the Petri dish. The cells were then incubated in the respective standard growth conditions to assess cell resistance. Due to the aerobic conditions, this experiment could not be performed in the case of T. neapolitana, an anaerobic microorganism.
Mars Humidity and Pressure Conditions Simulation: Effect of Desiccation and low Pressure on Microbial Growth
100 μl of each cell sample (with an O.D540 value of 0.600) were desiccated by air drying to be subsequently exposed to the simulated low pressure conditions of Mars conditions. The desiccated cells were then set for 1 h, 1 and 8 days at low pressure (4 mbar) that was reproduced in a Martian environment Simulator built by Cosmic Physic Laboratory of the University “Parthenope”. After this treatment, cell pellets were recovered and then used as inoculum in the respective standard growth media to assess cellular resistance.
Results and Discussion
Effect of Temperature Stress on Microbial Growth
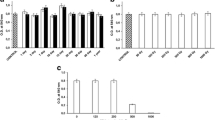
Cells of all extremophilic strains were stored at a wide range of temperatures (from −196 to 85 °C) to assess their resistance to the extreme variations of such parameter typical of space condition. Resistance was assessed by measuring the ability of treated colonies to give cell growth in comparison to that observable after storage in optimal standard conditions. In the case of S. solfataricus (Fig. 1) after one week storage at T from −196 to 25 °C cell growth registered after 1 day of incubation was comparable to that observed after storage in standard conditions; the same cell growth level (O.D540 value of 0.700) was gained after storage at 50, 70, 85 ° C but after a longer incubation time, i.e., 4, 6, and 7 days respectively.

S. solfataricus growth after 1 week storage at −196, −80, −20, 0, 4, 10, 25, 50, 70, 85 °C
After 3 months storage at T from −20 to 4 °C the cell growth of the halophilic species H. hispanica (Fig. 2, open bars) registered after 1 day of incubation, was comparable to that observed after storage in standard conditions; the same cell growth was observed after storage at 50, 70 and 85 °C but with a delay of 3, 4 and 5 days respectively. At −195 °C, −80 °C H. hispanica showed a lower resistance as showed by the low capability to grow again after the treatment.

H. hispanica (open bars), T. neapolitana (full bars), G. thermantarcticus (dashed bars) growth after 3 months storage at −195, −80, −20, 0, 4, 10, 25, 50, 70, 85 °C
The thermo-anaerobe T. neapolitana (Fig. 2, full bars) was able to survive to all the tested temperature stress although it showed an average resistance lower than the other species. Indeed for this strain, cell growth was always lower than that observed after storage in the standard conditions. The thermophilic species G. thermantarcticus (Fig. 2, dashed bars) showed to be highly resistant to 3 months storage at T values from 0 °C to 70 °C: in all these experiments indeed it reached an O.D540 value comparable to that observed after storage in the standard conditions; growth after storage at −196 °C and at 85 °C (the lowest and the highest T investigated) was less significant thus suggesting a minor resistance to these stressing conditions.
Effect of UV Radiation (λ = 254 nm) on Microbial Growth
The effect of ultraviolet radiations was investigated by monochromatic irradiation with a lamp whose power and wavelength allowed to simulate the UVC portion of solar radiation on Mars (Schuerger et al. 2003) that was applied in terrestrial conditions of temperature, pressure and atmosphere. This experiments could be carried out for all strains except the thermo-anaerobe T. neapolitana since UV irradiation was performed in terrestrial aerobic conditions that are lethal for this bacterium. In the case of H. hispanica (Fig. 3, open bars), exposition caused for almost all irradiation intervals only 20 % decrease of growth with respect to the control experiment. In the case of G. thermantarcticus (Fig. 3, full bars) UV irradiation resistance decreased only slightly with increasing time of exposition: indeed till after 30 min irradiation its growth level was 80 % with respect to the control experiment, and it was still about 40 % after 1 h irradiation at 254 nm.

H. hispanica (open bars), G. thermantarcticus (full bars), S. solfataricus (dashed bars) growth after UV radiation (λ = 254 nm) exposition
The highest resistance to UV irradiation was showed by the thermo-acidophilic archeon S. solfataricus (Fig. 3, dashed bars): after all irradiation treatment its growth was equal to that registered in standard conditions, only after 15 min irradiation a 40 % decrease was found, but further experiments are needed to confirm this trend. Nevertheless it is noteworthy to underline the surviving of such species that were treated in a vegetative state, while usually resistance to UVC radiations is displayed from the spore counterparts (Newcombe et al. 2005).
Mars Humidity and Pressure Conditions Simulation: Effect of Desiccation and low Pressure on Microbial Growth
The surface environment of Mars is now characterised by the scarcity of liquid water, extreme low temperatures and pressure, an anoxic atmosphere and exposition to solar and cosmic radiation. The experiments here described, allowed to assess at the same time the cell resistance to two of Mars environment parameters i.e., humidity and low pressure. All experiments were carried out on previously desiccated cells that were then exposed in the Mars simulator chamber at a 4 mbar pressure. Unfortunately, such testes could be carried out only with H. hispanica and G. thermantarcticus, since both the thermo-acidophilic archeon S. solfataricus and the thermo-anaerobe T. neapolitana did not survive to the initial step of desiccation; in this latter case it was probably lethal because it was performed in aerobic conditions.
Both H. hispanica and G. thermantarcticus species were able to survive to the initial step of desiccation, that was required to expose the cell pellets to the extremely low pressure conditions reached in the simulator. Exposition of desiccated G. thermantarcticus (Fig. 4, full bars) cells to Mars’s pressure for all the time intervals investigated didn’t affect significantly the growth of this bacterium: indeed after all the exposition time intervals investigated, a growth level comparable to the standard conditions was reached after only 1 day of incubation. Similar resistance was exhibited by H. hispanica (Fig. 4, open bars) desiccated cells but for this strain a delay in growth after the stressing treatment was observed: indeed the O.D. values comparable to the control experiment could be reached only by incubating for 2 days after 1 h exposition in the Mars simulator, or by incubating for 5 days after 24 h and 8 days of exposition in the simulator.

H. hispanica (open bars), G. thermantarcticus (full bars) growth after exposition to Mars simulated conditions
Conclusions
The described experiments were carried out in order to investigate extremophiles resistance to some stressing conditions typical of space environments i.e., wide variation of temperature, exposition to high energy UV radiations, desiccation and extremely low pressure values.
All the selected extremophilic strains showed a good resistance to the simulation of the temperature variation in the space: indeed all extremophiles were able to grow rapidly (from 1 to maximum 5 days of incubation) after prolonged storage also at temperature values very far from their respective optimal conditions.
The exposition to UV radiation affected only slightly the growth of H. hispanica, G. thermantarcticus and S. solfataricus: the latter showed to be the most resistant to this stress parameter whereas survival of H. hispanica and G. thermantarcticus decreased with increasing irradiation time. This result was interesting since the most significant resistance to UV radiation is displayed by some bacterial spores such as spores of different Bacillus species: the results here reported suggested that extremophilic vegetative cells were able to survive to Martian UVC irradiation at a wavelength usually employed as bactericidal radiation.
In the experiment of Mars humidity and pressure simulation the behaviour of S. solfataricus and T. neapolitana could not be investigated since the previous desiccation step was lethal for these two species. Nevertheless, it was possible to verify that both H. hispanica and G. thermantarcticus were resistant either to desiccation and Mars pressure, although H. hispanica growth after this simulation was strongly delayed in comparison to the standard conditions.
Overall these results suggested that all four extremophilic strains are good candidates for possible future experiments of microorganisms exposition in space missions.
References
Coorevits A, Dinsdale AE. Halket G, Lebbe L, De Vos P, Van Landschoot A, Logan NA (2012) Taxonomic revision of the genus Geobacillus: emendation of Geobacillus, G. stearothermophilus, G. jurassicus, G. toebii, G. thermodenitrificans and G. thermoglucosidans (nom. corrig., formerly ‘thermoglucosidasius’); transfer of Bacillus thermantarcticus to the genus as G. thermantarcticus; proposal of Caldibacillus debilis gen. nov., comb. nov.; transfer of G. tepidamans to Anoxybacillus as A. tepidamans and proposal of Anoxybacillus caldiproteolyticus sp. nov. Int J Syst Evol Microbiol 62:1470–1485
Des Marais DJ, Nuth JA, Allamandola LJ, Boss AP, Farmer JD, Hoehler TM, Jakosky BM, Meadows VM, Pohorille A, Runnegar B, Spormann AM (2008) The NASA astrobiology roadmap. Astrobiology 8:715–730
Jannash WJ, Huber R, Belkin S, Stetter O (1988) Thermotoga neapolitana sp. nov.of the extremely Thermophilic eubacterial genus Thermotoga. Arch Microbiol 150:103–104
Lama L, Calandrelli V, Gambacorta A, Nicolaus B (2004) Purification and characterization of thermostable xylanase and β-xylosidase by the thermophilic bacterium Bacillus thermantarcticus. Res Microbiol 155:283–289
Newcombe D, Schuerger A, Benardini J, Dickinson D, Tanner R, Venkateswaran K (2005) Survival of spacecraft-associated microorganisms under simulated Martian UV irradiation. App Environ Microbiol 71(12):8147–8156
Nicholson W, Krivushin K, Gilichinsky D, Schuerger A (2013) Growth of Carnobacterium spp. from permafrost under low pressure, temperature, and anoxic atmosphere has implications for Earth microbes on Mars. Proc Natl Acad Sci 110(2):666–671
Onofri S, Barreca D, Selbmann L, Isola D, Rabbow E, Horneck G, Zucconi L (2008) Resistance of Antarctic black fungi and cryptoendolithic communities to simulated space and Martian conditions. Stud Mycol 61:99–109
Romano I, Poli A, Finore I, Huertas FJ, Gambacorta A, Pelliccione S, Nicolaus G, Lama L, Nicolaus B (2007) Haloterrigena hispanica sp. nov., an extremely halophilic archaeon from Fuente de Piedra, southern Spain. IJSEM 57:1499–1503
Saffary R, Nandakumar R, Spencer D, Robb FT, Davila JM, Swartz M, Ofman L, Thomas RJ, DiRuggiero J (2002) Microbial survival of space vacuum and extreme ultraviolet irradiation: strain isolation and analysis during a rocket flight. FEMS Microbiol Lett 215:163–168
Schuerger A, Mancinelli R, Kern R, Rothschild L, McKay C (2003) Survival of endospores of Bacillus subtilis on spacecraft surfaces under simulated martian environments: implications for the forward contamination of Mars. Icarus 165(2):253–276
Zilling W, Stetter KO, Wunderl S, Schulz W, Priess H, Scholz I (1980) The Sulfolobus – “Caldariella” group: taxonomy on the basis of the structure of DNA-dependent RNA polymerases. Ach Microbiol 125:259–269
Acknowledgments
This work has been implemented in the frame of the project PON01_01966 “Integrated agro-industrial chains with high energy efficiency for the development of eco-compatible processes of energy and biochemicals production from renewable sources and for the land valorisation” funded by “MIUR- Ministero dell’Istruzione, dell’Università e della Ricerca”.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Mastascusa, V., Romano, I., Di Donato, P. et al. Extremophiles Survival to Simulated Space Conditions: An Astrobiology Model Study. Orig Life Evol Biosph 44, 231–237 (2014). https://doi.org/10.1007/s11084-014-9397-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11084-014-9397-y