
Abstract
Lethal coalitionary aggression is of significant interest to primatologists and anthropologists given its pervasiveness in human, but not nonhuman, animal societies. Chimpanzees (Pan troglodytes) provide the largest sample of recorded lethal coalitionary aggression in nonhuman primates, and most long-term chimpanzee study sites have recorded coalitionary killing of conspecifics. We report an inferred lethal attack by resident males on a former alpha male chimpanzee (P. t. verus) at Fongoli in Senegal. We describe the male’s presence in the community, his overthrow, social peripheralization for >5 yr, and his attempt to rejoin the group as well as circumstances surrounding his death. We report attacks by multiple chimpanzees on his dead body, most frequently by a young adult male and an older female. The latter also cannibalized the body. Coalitionary killing is rare among West African chimpanzees compared to the East African chimpanzee (P. t. schweinfurthii). This pattern may relate to differences in population densities, research effort, and subspecies differences in biology and behavior.
Similar content being viewed by others


Introduction
Coalitionary killing among adults has been reported in several nonhuman primate species but is rare (Wilson et al. 2014; Wrangham and Peterson 1996). It is most common in chimpanzees (Pan troglodytes), one of the most intensively studied wild primates. Lethal aggression in and between chimpanzee communities has been reported at almost every long-term research site where these apes have been studied for more than a decade (Wilson et al. 2014). A recent summary of lethal events at 18 chimpanzee and 6 bonobo (Pan paniscus) study sites representing 518 observer years of records found that intracommunity killing of weaned individuals was rare, with only 9 observed and inferred cases reported for a total 426 observation years (Wilson et al. 2014). The analyses did not support the hypothesis that anthropogenic disturbance caused lethal aggression in chimpanzees (Wilson et al. 2014). Instead, the authors concluded that an evolutionary adaptive explanation better fit the pattern of lethal aggression recorded in chimpanzees because indicators of competition intensity, such as the number of adult males within a community and population density, were the best predictors of lethal events (Wilson et al. 2014). Further patterns in the data include the following: 1) Most lethal aggression of weaned individuals involves adult males as both the victims and attackers. 2) Most lethal events involve attacks on individuals from neighboring chimpanzee communities. 3) The behavior is less common in the West African chimpanzee subspecies (P. t. verus) and the bonobo (P. paniscus) compared to other Pan taxa (Wilson et al. 2014, Fig. 1).
Here, we add to the database on lethal coalitionary aggression in providing an account of what appeared to be a lethal attack on a peripheral, former alpha male at the Fongoli savanna chimpanzee study site in Senegal (Foudouko). We place our case in the context of his group. We describe 1) the victim’s social status, including his rise to and fall from alpha position, his peripheralization, and his attempts to reestablish himself in the male social group; 2) circumstances surrounding his death; 3) the treatment of his dead body by other community members; and 4) events that occurred in the Fongoli chimpanzee community immediately after his death. Finally, we 5) provide information on the different alpha males thus far recorded at Fongoli, in addition to brief descriptions of their fall from alpha status, for comparison with Foudouko.
Methods
Study Site
The Fongoli study site, under the auspices of the Fongoli Savanna Chimpanzee Project (FSCP), is located within the Kedougou Department in southeastern Senegal (12°40′N, 12°13′W). The Fongoli study site is the hottest, driest and most vegetatively open site where chimpanzees are habituated to the presence of observers. The extensive dry season lasts for >7 mo (November–May), with rainfall averaging <1000 mm annually. The site is a woodland–savanna, which includes open (tall and short) grassland, woodland, bamboo woodland, and small patches of gallery forest, in addition to ca. 5% anthropogenically disturbed areas such as fields, artisanal gold mines, and villages (Bogart and Pruetz 2011; Pruetz 2006). The majority of the chimpanzees’ home range is composed of woodland, while forest accounts for <3% of the area (Bogart and Pruetz 2011; Pruetz 2006). Temperatures exceed 40 °C at the end of the dry season, and all but a few water sources dry up by the late dry season.
Study Subjects
Observers began study of the Fongoli chimpanzee community in April 2001, and habituation took ca. 4 yr. We began systematic all-day follows of focal adult male subjects in early 2005 after habituation had proceeded to the point at which individual males tolerated the presence of observers and could be followed from night nest to night nest.
Community size between 2005 and 2013, when Foudouko was killed, ranged between 31 and 35 individuals. We defined age–sex classes as follows: infants, age ≤4 yr; juveniles, 4–7 yr; adolescents, >7 and <15 yr (males) or until a female gave birth; and adults as males >15 yr (Baldwin 1979). We refer to males known or estimated to be between 15 and 20 yr of age as young adult. During the study, the number of adult males ranged between 10 and 12. The number of adult females ranged between 7 and 8. The adult male/female sex ratio ranged between 1.4 and 1.7. Previous mitochondrial DNA analyses of individuals of juvenile age or older provide the matrilineal relatedness of all of the males and most of the females (excluding two females that had recently immigrated into the Fongoli community) of adult age at the time of Foudouko’s death (Stewart 2011).
We ranked adult males in the dominance hierarchy based on pant-grunt records (pant-grunts are made by subordinate individuals to dominant individuals) and noted when rank changes occurred. Where a pair of adult males never pant-grunted to one another, we scored them as tying for rank. During the early years of the project, observers did not collect all-occurrence data on focal subject social behavior, but we recorded any changes in the hierarchy as defined by observed pant-grunt records that did not coincide with the existing hierarchy.
Data Collection
Observers maintained a minimum distance of 10 m from most chimpanzees but, in the early years of the project, we followed mid-ranking adult male DF at a distance of 15–20 m when he was in small parties (<5 individuals) or when alone because of his relative timidity. Observers did not identify all adult females of the Fongoli community until January 2006.
Systematic behavioral data collection at Fongoli consists of all-day follows of focal adult males, with instantaneous sampling of general activity and location via a Global Positioning System (GPS) at 5-min intervals. We attempted to collect systematic behavioral data on adult male focal subjects ≥20 days per month, although seasonal factors such as river flooding result in some degree of monthly variation in data collection. We sampled rare but important behavioral events, such as contact aggression, copulations, hunts and meat sharing, and reactions to humans or predators opportunistically for all individuals. Operational definitions of chimpanzee behavior follow Nishida et al. (1999).
Data Collection and Analyses Specific to Foudouko
After March 2008, when Foudouko disappeared for almost 9 mo, we took detailed notes on his behavior whenever we encountered him. We recorded his reaction to observers and his social interactions and grouping with other individuals. Following the discovery of his body on June 15, 2013, three observers took detailed notes on how other community individuals reacted to his body over the course of 3.5 h, with two observers collecting data simultaneously (see Tables I, II and III). In addition, one observer took ca. 18 min of video as individuals interacted with Foudouko’s body (Electronic Supplementary Material [ESM] Tables SI–SX, corresponding to Video Clips 1–10). We transcribed records from video clips (ESM Video Clips 1–10) and combined these with detailed observations taken by three different researchers (ESM Tables SI–SX) to provide a chronological account of the Fongoli chimpanzees’ treatment of Foudouko’s body after his death (see Table II). To assess the frequency with which observers saw Foudouko before and after his disappearance in early 2008, we compared days of data collection where he was seen and not seen from January 2006 to December 2007 and January 2008 to June 2013 (N = 90 mo) (Fig. 1).

Data collection days on which Foudouko was seen or not seen in the Fongoli chimpanzee study 2006–2013: Foudouko was the alpha male until late 2007. Fongoli study site, Senegal.
Ethical Note
We conducted all research reported here with permission and in accordance with guidelines set forth by the Department of Forestry in the Republic of Senegal and the IACUC guidelines of Iowa State University. All research on chimpanzees was strictly observational.
Results
Foudouko’s Place in the Fongoli Community
We identified Foudouko as a sexually mature late adolescent or young adult male in 2003. We identified Foudouko as alpha male from early 2005, when adult males were habituated for nest-to-nest follows, until late 2007. He did not pant-grunt to any other adult males, and all other adult males pant-grunted to him. We estimated that Foudouko was likely in his late teens when he became an alpha male.
In 2006 and 2007, Foudouko was present in parties on 63.4% of observation days (N = 426 days) (Fig. 1). He lost alpha rank at the end of September 2007, at around the same time the beta male and Foudouko’s frequent coalitionary partner, MM, was severely injured (suffering a broken or dislocated hip or femur). We did not see MM with the other males in the community for ca. 6 weeks after this (October 19–December 3, 2007), and he was very frail when he returned, submitting to almost all adult males of the community, including a very aged male; females also ignored his displays.
The last day on which we collected focal observation data on Foudouko was March 27, 2008, and he disappeared at the end of March 2008. On one of the last days on which we saw Foudouko, observer D. Kante estimated that most or all of the community’s adult males chased him from one of the only permanent dry season water sources. We next saw Foudouko in December 2008, when K. Boyer Ontl and D. Kante found him with MM and two adult females (NI, NN) and their offspring (NE, TV), 263 days after he had disappeared. Foudouko was no longer tolerant of observers in that he repeatedly moved rapidly away from them and researchers abandoned their attempt to follow his party given the dry season heat stress.
Observers saw Foudouko once or twice per year with other chimpanzees in 2008, 2009, and 2010 but did not see him in 2011. In 2012 and 2013, Foudouko appeared to be slowly integrating back into the adult male social group. We saw him in small parties with several adult males and with the whole community on one occasion, but he was mainly associated with MM, who had regained beta male status (Table I). Although most males were aggressive toward Foudouko, alpha male DV and MM (maternal siblings, based on mtDNA analyses: Stewart 2011) affiliated with him and were not aggressive toward him as were other adult males; they did not participate when other males chased Foudouko. For example, on June 3, 2010, J. D. Pruetz witnessed Foudouko arriving at Sakoto ravine late in the day. Foudouko approached adult male DV and quietly pant-grunted to him. Afterwards, both males disappeared into the woodland. The next day, after the adult males had moved southeast, adolescent male LT alarm called, which resulted in the males returning rapidly and chasing something unseen >1.5 km south. We have not seen Fongoli chimpanzees chase predators (spotted hyenas [Crocuta crocuta], leopards [Panthera pardus]) or prey for such distances (Pruetz and Boyer-Ontl, in prep.), nor were the alarm calls of prey species, e.g., Papio papio, heard. We have never encountered extra-community males in the Fongoli home range. The alarm calls given by LT sounded different to observers from calls given in response to snakes and potential predators (Pruetz and Boyer-Ontl, in prep.). These factors led observers to speculate that the males were chasing Foudouko. In 2013, Foudouko quietly approached a party and directed his gaze at MM, who left to travel with Foudouko and disappeared for 1 week.
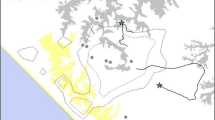
In late 2012 and in 2013, Foudouko spent more time in proximity to the social group of males, and continued to display nervousness around researchers, hiding or moving away. For instance, on February 26, 2013, K. Boyer Ontl followed a small party containing Foudouko, MM, NI, NN, TV (NI’s juvenile daughter), and VC (NI’s infant son) for most of the day (Table I). Foudouko stayed >75 m from the observer on an open grassland plateau when she first encountered the party at dawn, and although he tolerated her at closer distances, e.g., 20 m, later in the day, most of the time he positioned himself behind a tree or a tall tuft of grass. On several occasions, the behavior of Fongoli males suggested that Foudouko was in the vicinity, but observers did not see him. These encounters were almost always in the southwestern portion of the Fongoli chimpanzees’ range, near the core area used during the rainy season especially and in the vicinity of the Fongoli research camp. Once, J. D. Pruetz witnessed the adult males of the Fongoli group engage in what appeared to be an aggressive chase. Only alpha male DV and beta male MM refrained from taking chase. The observer then saw Foudouko as she was returning to camp at dusk, and she followed him for 15 min from a distance, undetected. He moved parallel to the other chimpanzees’ location at a nesting site at Maragoundi ravine (Fig. 2). He frequently climbed trees and gazed in the direction of the social group but disappeared on detecting the observer.

Map (a) of locations where Foudouko was seen during his peripheralization and attempted reentry into the adult male Fongoli chimpanzee social group, relative to the home range area (b) of the entire social group. Fongoli study site, Senegal.
Circumstances Surrounding Foudouko’s Death in 2013
We last saw Foudouko alive on May 5, 2013 when we observed him in the company of adult males MM and KL. During the days before his death, observers suspected that Foudouko was in the vicinity on several occasions. For example, on June 12, 2013, J. D. Pruetz followed the entire Fongoli community, excluding Foudouko, to Maragoundi ravine. At 19:30 h, after a chimpanzee screamed west of the nesting party, the males left their nests and ran in that direction. The observer followed adult male KL who stopped short of what sounded like an aggressive encounter, and they met adult male BI returning from the area. She then found adult males BN, MI, and LT returning from the area, followed by adult male SI returning from an area nearby moments later. Beta male MM was nearby, although he did not appear to have been involved in the confrontation. MM then disappeared until 18:00 h the following day. On June 14, 2013, two researchers (DK and LB) reported a similar incident in which all the adult males raced in a certain direction as if giving chase to another chimpanzee.
Foudouko likely approached the chimpanzee group during the night of the June 14/15 after they had nested at Sakoto ravine. Fongoli village is ca. 1 km from Sakoto and during the early morning of June 15 observers heard vocalizations (pant-hoots, screams) between 02:00 and 03:00 h. They were atypical compared to the pant-hoots often heard from camp when chimpanzees nested at Sakoto in that they were very loud, extensive, and their placement indicated that some individuals had left their nesting area and had moved south. Calls indicated that individuals were rapidly moving on the ground and involved in extensive agonism. Alpha male DV’s pant-hoot, for example, indicated he was somewhere between the nesting site and the location where Foudouko was discovered dead the next morning. Beginning with the rank of alpha male, the adult male dominance hierarchy at the time of Foudouko’s death was 1) DV, 2) MM, 3) BO, 4) KL, 5) JM, 6) LP, 7) MI, 8) LT, 9) DF, 10) BI, 11) BN, 12) SI.
Treatment of Foudouko’s Body After Death
On the morning of June 15, 2013, a research assistant (M. Sadiakho) arrived before dawn at the chimpanzees’ nesting site. He returned to camp at ca. 08:00 h and reported that he had seen a dead chimpanzee, which he presumed to be Foudouko because all 12 other community males were present. He reported that most of the males had attacked Foudouko’s body (Table II). At this time, two additional researchers (J. D. Pruetz, E. G. Wessling) left to observe the body (Table II). On arrival at the scene, one observer identified the dead chimpanzee as Foudouko. When J. D. Pruetz and E. G. Wessling arrived in the vicinity of the body, they saw adult female FA (alpha male DV’s mother and presumably beta male MM’s mother) termite fishing <40 m from Foudouko and other chimpanzees in the area. For the next 2 h, observers recorded the chimpanzees’ reactions to Foudouko’s body, until the party departed (Table II). We observed fresh wounds on Foudouko’s hands, feet, back, and anus, with most of the wounds on the neck, torso, and appendages (Figs. 3 and 4) being inflicted postmortem (Tables II and III). Given their locations and severity, none of the bite wounds would have been lethal, save for possibly a severe bite to the right foot (Fig. 5) that could have considerably hindered his movement and escape and that could have resulted in significant blood loss. This foot wound was the most severe wound seen on Foudouko’s body when observers arrived at the scene. Chimpanzees may have inflicted this wound at or around the time of Foudouko’s death. We observed few wounds on any other individuals recorded in the vicinity of Foudouko’s body, i.e., potential attackers. Only older adult male BN exhibited fresh wounds, on his arm, and these appeared to be superficial. We approached Foudouko’s body after the party left the area. We found little sign of rigor mortis when we left his body at 11:20 h, suggesting he died within the previous 24 h. We judged livor mortis as minimal given the extent to which his neck wound still bled on rupture during the postmortem attacks. However, because multiple chimpanzees turned over and dragged Foudouko’s body, pooling patterns could have been interrupted. We found no extensive evidence of bleeding on the ground in the vicinity of Foudouko’s body.

Body of male chimpanzee Foudouko, 11:00 h, June 15, 2013. Fongoli study site, Senegal.

Close-up of male chimpanzee Foudouko’s genital area with wounds inflicted after death. Fongoli study site, Senegal.

Male chimpanzee Foudouko’s fingers and toes with signs of bites. Fongoli study site, Senegal.
During observations of Foudouko’s body, several young adult males beat his body and bit it repeatedly (Table III; Fig. 6), with young adult male BO most often attacking the body. Most group members, including several adult females, pulled on the body and smelled it intensely. Following BO, older adult female FA, mother of current alpha male DV and of adult male MM, attacked the body most frequently, and consumed more of it than any other chimpanzee. Neither of her sons attacked the body in similar fashion. These males had not behaved aggressively toward Foudouko earlier, and had engaged in affiliative behavior with him. MM displayed around the body several times, pulling Foudouko once and later stamping on him once. MM seemed to be attempting to rouse Foudouko, rather than trying to inflict injury, as his behavior was less aggressive than the other males’ displays, e.g., adult males BN, KL, BI, MI, BO, LT, and JM and adolescent males LX and DW. Alpha male DV approached and smelled the body several times but did not display toward Foudouko or attempt to move him. Numerous females and their offspring approached and smelled the body. FA and a young adult female (TM) fed on different parts of the body but ingested only small pieces of flesh.

Adult male chimpanzee Mike (MI) (left) and adolescent male LX (right) examine Foudouko’s body. Fongoli study site, Senegal.
No attackers were matrilineally related to Foudouko (Stewart 2011). Two of the frequent attackers, young adult males BO and LT, were matrilineally related to one another (Stewart 2011). LT is the son of adult female LU, brother to adolescent male LX, and likely nephew of BO, as BO’s mother WI (now deceased) was apparently LU’s mother. BO, LT, LU, and LX attacked Foudouko’s body. No other attackers were matrilineally related.
We buried Foudouko’s body near where we discovered his corpse, under the supervision of the local authorities. His bones will be exhumed for examination in the future.
Events in the Community Immediately after Foudouko’s Death
On June 16, 2013, an observer contacted the same chimpanzee party (N = 29 individuals) at dawn at their nesting site. At 06:05 h, almost all of the males and several females (TM, EV) traveled silently and rapidly toward the site where they had left Foudouko’s body. They began a pant-hoot chorus a few minutes after beginning travel, fell silent again, then reassured one another physically, e.g., touch fingers to mouth, hold genitals, embrace, as they traveled together, listening and looking toward the location where they left Foudouko’s body. All of the males were present except BI, BN, and alpha male DV. Alpha male DV then arrived in a silent, charging display. The party continued traveling to Foudouko’s burial site. Young adult male LT led the party, along with adolescent male DW and young adult males BO, MI, and JM. Young adult males JM and BO reassured one another by touching and embracing. DV and MM were further behind the party. All traveled silently and vigilantly. BN arrived at the rear of the party. At 06:35 h, the party neared the site where Foudouko’s body was last seen, and multiple males smelled the ground.
The party traveled quickly to the exact site where they last left Foudouko’s body and smelled the ground intensely. They then moved ca. 15 m to where we buried Foudouko. They smelled the rocks around the burial site, including where we laid the body to prepare to lower it into the grave. After smelling other areas where the body had been dragged, the party left the site at 06:50 h and traveled back to Sakoto ravine, backtracking along the same route taken from the night nests to the burial site. The group gave alarm calls nervously and frequently in the direction of Foudouko’s site, e.g., at 07:30, 08:41, 09:00, 09:20, and 12:40 h, despite having moved several hundred meters east. Third-ranked and young adult male BO hit beta male MM during a fight among the males at 09:25 h. MM and BI left the area, screaming. MM recruited his mother (based on behavioral indications and mtDNA analysis), adult female FA, who then helped recruit older adult males DF, LP, and BN to retaliate against BO. BO also attacked (hit) fourth ranked male KL at 13:35 h after the party returned to Sakoto. Such an attack by BO, on MM especially, was unusual. The party, including all adult males, nested at Sakoto on June 16, 2013. On June 17, the party left Sakoto ravine in the morning and traveled north ca. 2 km to Petit Oubadji ravine, in the direction opposite of Foudouko’s death site.
Alpha Males at Fongoli
Thus far at Fongoli, we have recorded four different alpha males in 11 yr of study following habituation for systematic data collection. The alpha male (YO) that rose from the third position in the hierarchy to take Foudouko’s place disappeared in 2010, <1 yr after LP usurped him from the dominant position. YO held the alpha position for ca. 15 mo, while LP was alpha for just over 3 yr. Following LP’s fall from alpha position in 2012, he was absent for a several weeks but then returned to range with community. LP dropped to the middle-lower end of the hierarchy, but he has since risen to middle ranking male. The ascendancy of young males into the dominance hierarchy (N = 12 adult males) created an unstable hierarchy for several years. Only the highest and lowest ranks remained stable over a period of >2 yr at Fongoli (2013–2014), with the hierarchy stabilizing in early 2015. The current alpha male DV is the longest reigning alpha thus far, having usurped LP in March 2012.
Discussion
Our report of lethal coalitionary aggression in a community of wild, West African chimpanzees fits the general pattern among chimpanzees in which adult males are usually both attackers and victims (Wilson et al. 2014). However, most lethal coalitionary aggression in chimpanzees is between communities, rather than within communities as was the case here (Wilson et al. 2014). Foudouko’s case is also only one of a few recorded for West African chimpanzees, a subspecies characterized, along with bonobos, as having relatively little lethal coalitionary aggression compared to the East African chimpanzee subspecies (Wilson et al. 2014).
The Fongoli chimpanzee community is more cohesive than other study groups, and the entire community often ranges together, especially during the early rainy season (Pruetz and Bertolani 2009). However, Foudouko was estranged from most members of his natal group for >5 yr, was rarely seen, and then only in the company of specific individuals until he began to attempt to rejoin the adult male social milieu. Chimpanzees are highly social, especially males, and living even a semisolitary life would have been exceptionally stressful, as has been shown in chimpanzees experiencing social isolation (Reimers et al. 2007). The peripheralization of an adult male is rarely reported in the literature (Nishida 2012). However, at Mahale, the former alpha male Ntologi was absent from M group for ca. 9 mo, ranging alone and some distance from the rest of the group save for occasional socializing with a male age-mate (Nishida 2012), similar to our observations of Foudouko and MM. Ntologi rejoined M group, along with another adult male, Jilba, that had been similarly ostracized by the other males (Nishida 2012). Unlike Foudouko, Ntologi rose back up the ranks to alpha male in several months and retained the position for several years (Nishida 2012). However, like Foudouko, Ntologi was ultimately killed by community males. The M group chimpanzee community at Mahale decreased in number from 76 to 44 individuals over the course of the 5 yr that included Ntologi’s return and rise to alpha, his fall from alpha status, his death, and the following year (Nishida 2012). Although only three corpses were recovered, Nishida (2012) surmised that an epidemic had likely caused such high mortality, as they never discovered a splinter group. Although researchers did not witness the attack on Ntologi that left him in a coma and ultimately killed him, Nishida (2012) speculated that a gang attack by the alpha male and other males in M group resulted in Ntologi’s condition and was likely retaliation for the former alpha’s 15 yr of domination. The same may be true for Foudouko’s case at Fongoli, although our data on details of Foudouko’s tenure as alpha is incomplete compared to that of later alphas.
The unstable male social hierarchy at Fongoli appears to be related to the relatively low number of adult females in this community. Of the 18 chimpanzee communities included in Wilson et al. (2014), Fongoli had the lowest chimpanzee density at 0.4 individuals per km2 compared to the overall average of 3.6 individuals per km2 (N = 18 communities at 10 sites: Wilson et al. 2014) but the highest male to female sex ratio at 1.7 (Wilson et al. 2014, Extended Data Fig. 1a). The sex ratio of the communities for which we have evidence of intragroup lethal aggression of weaned individuals averaged 0.51 (N = 4). With Fongoli included, the average was 0.75 (N = 5). A sex ratio of 1.0 in Ateles has also been suggested as a contributing factor in lethal aggression among males (Campbell 2006; Valero et al. 2006).
The high level of male reproductive competition at Fongoli, along with a low population density and extremely low rate of intercommunity interactions (FSCP, unpubl. data) might explain the attack on Foudouko. This pattern supports the hypothesis that intracommunity aggression occurs when male–male social bonds relax as a consequence of low rates of intercommunity aggression and high rates of intracommunity competition (black-handed spider monkeys [Ateles geoffroyi], Barro Colorado Island, Panama: Campbell 2006; Yucatan, Mexico: Valero et al. 2006; East African chimpanzees [Pan troglodytes schweinfurthii], Ngogo, Uganda: Watts 2004) (see Table IV). Foudouko copulated with at least one receptive female and associated with another on two of the occasions he was with a mixed-sex party in 2013, several months before his death, indicating that he presented some degree of competition for other adult males in the community.
The skewed sex ratio at Fongoli may be due to human pressure. Local residents in the Kedougou region have noted that rare but opportunistic hunting of female chimpanzees has occurred in the area to obtain infants for the pet trade (P. Stirling, pers. comm.). One case of infant poaching has occurred at Fongoli since the project began in 2001 (Pruetz and Kante 2010), but given the slow life history of chimpanzees, even such a low rate could eventually reduce the number of reproductive females in a social group significantly. Moreover, the Fongoli group’s home range is bordered on three sides by anthropogenic (a highway and the town of Kedougou) and natural (Gambia River) landscape features that could create movement challenges for potential female immigrants. Therefore, we cannot reject the hypothesis that anthropogenic disturbance is a contributing factor of lethal aggression at Fongoli. However, this does not preclude the possibility that lethal intragroup aggression is also adaptive under such circumstances.
Treatment of Foudouko’s Body After Death
Observers saw multiple adult males hit or bite Foudouko’s body, including repeated and intense biting attacks by particular males, such as BO and BI. Several individuals consumed small pieces of flesh taken from his body. Adult females in particular cannibalized Foudouko’s body, similar to reports in West African chimpanzees at Taí Forest, Ivory Coast (Boesch 2009). However, at Fongoli, females ate only small pieces of flesh, unlike a Taí female that ate the entire testes (Boesch 2009).
The wounding patterns on Foudouko’s body were similar to those described in cases of lethal intergroup aggression (Muller 2002; Wrangham 1999). However, most of these wounds were made postmortem, including the trauma to the throat and most trauma to the genitals. Given the nature of these wounds, the video footage and descriptions of other lethal attacks by multiple males on lone males at other study sites (Watts 2004), and the patterns seen at Fongoli where multiple males have attacked a single individual (FSCP, unpubl. data; Marshack 2016), we suggest multiple individuals likely attacked Foudouko, which ultimately ended in his death during the night or early morning hours of June 15, 2013. On first encountering Foudouko’s body, we recorded a wounding pattern similar to those reported in the case of wounds inflicted during lethal aggression directed at an incumbent alpha male at Mahale, Tanzania (Kaburu et al. 2013). The wounds we observed on Foudouko’s hands, feet, and appendages suggest he was being held by some individuals, and it is likely that others were beating and stamping on him, as has been observed in East African chimpanzees (Watts 2004). He likely died of internal injuries. We also observed individuals using branches and rocks to attack Foudouko’s dead body, although it was unclear whether these caused wounds. Kaburu et al. (2013) similarly reported that an attacking male used a branch to hold the victim’s body down during a lethal coalitionary attack on the alpha. This is of interest given the focus on the use of weapons in reconstructing a hominin–human niche (Gintis et al. 2015; Phillips et al. 2014). Even though the weapons in the described cases could not be described as lethal weapons, they illustrate the use of tools as weapons during conspecific attacks.
Several studies have reported behaviors akin to caregiving or even mourning after the death of a conspecific social group member, especially kin (Boesch 2012; Buhl et al. 2012; Fiore 2013; King 2013). However, most of the reports in the literature on thanatology describe compassionate reactions to group members’ deaths or dying (Fashing and Nguyen 2011). In our case, save for alpha male DV and a few individuals that did not approach Foudouko’s body, almost all Fongoli chimpanzees behaved aggressively toward him. Similar behavior has been seen in rhesus monkeys (Macaca mulatta), in which adult males inferred to have killed another adult male also directed aggression at his dead body >20 times the baseline rate of aggression (Buhl et al. 2012). Adult female mandrills (Mandrillus sphinx) also directed coalitionary aggression at an incapacitated male (Setchell et al. 2006). A report of chimpanzees’ reactions to a dead adolescent male at Chimfunshi Sanctuary in the Zambia notes that individual relationships affected the manner with which individuals reacted to their dead group member (van Leeuwen et al. 2016). This seemed to be the case at Fongoli with MM in particular. Even though MM pulled and stamped on Foudouko’s body at the end of his final display, all of his earlier displays involved vocalizations directed toward the body and, specifically, into Foudouko’s face. The observer interpreted MM’s behavior toward Foudouko as protective when he first approached the corpse early on the morning of June 15. The interpretation that MM’s behavior was an attempt to rouse Foudouko coincides with that described for chimpanzees at Taí, Ivory Coast (Boesch 2012).
The chimpanzees’ travel to the site of Foudouko’s death the morning after he was killed is reminiscent of a report for bonobos at Lui Kotale, Democratic Republic of the Congo (Tokuyama et al. 2012). In that case, a large group traveled 1.8 km without feeding or foraging to a site where they had left a group member with a snare on his hand the previous night. The Fongoli chimpanzees exhibited nervous behavior for much of that morning, until the party had moved some distance from the site. Their direct and oriented travel in a nonfeeding context is notable given this chimpanzee community has not been observed to exhibit boundary patrolling behavior that is typical of chimpanzees elsewhere (Mitani and Watts 2005) despite the fact they have at least one other chimpanzee community bordering their home range.
Variation in Lethal Aggression Between Chimpanzee Subspecies
Chimpanzees contribute significantly to our understanding of lethal aggression in nonhuman mammals because this behavior is well documented, perhaps because this primate species is one of the best-studied wild animals in terms of observer-year effort (Wilson et al. 2014). Lethal aggression involving adults has been recorded in few other primates. It occurs in some Atelines (black-handed spider monkeys: Campbell 2006; muriquis [Brachyteles arachnoides]: Talebi et al. 2009), a taxon that is also characterized by high fission–fusion dynamics and female emigration from the natal group, as well as in Cebus that, unlike Ateles and Pan, exhibits male dispersal. At Lomas Barbudal in Costa Rica, the rate of observed and inferred lethal aggression among white-faced capuchins (Cebus imitator) over the course of 12 yr has been likened to that of the East African chimpanzee (Gros-Louis et al. 2003). Intergroup killing (N = 2) is more common than intragroup (N = 1) killing (Gros-Louis et al. 2003), but sample sizes are small.
Species and subspecies differences in lethal aggression appear to exist within the genus Pan. Records of observed, inferred or suspected lethal aggression are uncommon in West African chimpanzees compared to such data on the East African subspecies (Wilson et al. 2014), although the latter have been studied more intensively in terms of observation years and number of study sites. On average, lethal aggression in western chimpanzees occurs once every 5 yr when observed or inferred cases are considered, and if only observations are considered, once every 9 yr. Of the 58 total observed killings (infanticidal, intergroup, intragroup) reported for chimpanzees, 15 were of adults between groups, and only one involved the West African subspecies (Wilson et al. 2014). Statistically, there was no difference between lethal aggression in general in West African chimpanzees and bonobos, but East African chimpanzees had significantly higher rates of lethal aggression than bonobos (Wilson et al. 2014). Lethal aggression within a social group or community is rarer than intercommunity killing, however, and small sample sizes preclude statistical testing of differences between species and subspecies. Nevertheless, of the nine recorded killings of weaned victims within a community (four observed, five inferred), only two are from Fongoli (Foudouko and adolescent male FR). Explanations for the differences in the recorded frequency of lethal coalitionary aggression between chimpanzee subspecies include possible variation in population densities, as well as the relative intensity of study of East African chimpanzees compared to other subspecies (Wilson et al. 2014).
The variable rates of lethal aggression in Pan, parsed along subspecies and species lines, suggest that genetic, cultural, or environmental mechanisms, or some combination of these factors, may explain the rarity of killing in western chimpanzees and bonobos. Although we cannot explicitly test these causal factors, it is intriguing that some studies recognize the western chimpanzee as the most genetically differentiated subspecies (Gonder et al. 2011), while others link Pan troglodytes verus more closely with the Nigerian subspecies (P. t. ellioti) than with East African (P. t. schweinfurthii) and Central African (P. t. troglodytes) chimpanzee subspecies (Prado-Martinez et al. 2013). Traits related to social cohesion and behavior also differ between the subspecies, such as larger party size and more frequent male–female cohesion during ranging (Boesch et al. 2008; Lehmann and Boesch 2005; Pruetz and Bertolani 2009; Yamakoshi 2004), as well as biological differences such as the quicker resumption of swelling after parturition and extended female genital swelling (FSCP, unpubl. data; Stumpf 2007, Table 19.6) in West African chimpanzees. These similarities prevail across habitat types, supporting the hypothesis that variation in lethal aggression between chimpanzee subspecies may not be explained completely by environmental factors. Compared to East African chimpanzees, over evolutionary time, bonobos and western chimpanzees appear to have faced selective pressures under which lethal coalitionary aggression was less likely to provide fitness benefits for attackers, resulting in a decreased propensity toward such behavior.
Our contextualization of the ostracization, peripheralization, and subsequent death of a former alpha male chimpanzee at Fongoli adds to the database of lethal coalitionary aggression in chimpanzees. This case illustrates the potential complications of dichotomizing “natural” vs. “anthropogenic” explanations for the root causes of lethal coalitionary aggression and highlights the variation in lethal aggression between taxa of the genus Pan.
References
Baldwin, P. (1979). The natural history of the chimpanzee (Pan troglodytes verus) at Mt. Assirik, Senegal. Ph.D. dissertation, University of Stirling, UK.
Boesch, C. (2009). The real chimpanzee: Sex strategies in the forest. Cambridge: Cambridge University Press.
Boesch, C. (2012). Wild cultures: A comparison between chimpanzee and human cultures. Cambridge: Cambridge University Press.
Boesch, C., Crockford, C., Herbinger, I., Wittig, R., Moebius, Y., & Normand, E. (2008). Intergroup conflicts among chimpanzees in Tai National Park: lethal violence and the female perspective. American Journal of Primatology, 70, 519–532.
Bogart, S. B., & Pruetz, J. D. (2011). Insectivory of savanna chimpanzees at Fongoli, Senegal. American Journal of Physical Anthropology, 145, 11–20.
Buhl, J. S., Bonn, A., Ruiz-Lambides, A., Gonzalez-Martinez, J., Platt, M. L., & Brent, L. J. N. (2012). Response of rhesus macaques (Macaca mulatta) to the body of a group member that died from a fatal attack. International Journal of Primatology, 33, 860–871.
Campbell, C. J. (2006). Lethal intragroup aggression by adult male spider monkeys (Ateles geoffroyi). American Journal of Primatology, 68, 1197–1201.
Fashing, P., & Nguyen, N. (2011). Behavior toward the dying, diseased, or disabled among animals and its relevance to paleopathology. International Journal of Paleopathology, 1, 128–129.
Fawcett, K., & Muhumuza, G. (2000). Death of a wild chimpanzee community member: possible outcome of intense sexual competition. American Journal of Primatology, 51, 243–247.
Fedigan, L. M., Rose, L. M., & Morera Avila, R. (1997). See how they grow: Tracking capuchin monkey (Cebus capucinus) populations in a regenerating Costa Rican dry forest. In M. A. Norconk, A. L. Rosenberger, & P. A. Garber (Eds.), Adaptive radiations of Neotropical primates (pp. 289–308). New York: Plenum.
Fiore, R. R. (2013). What defines us: An analysis of grieving behavior in non-human primates as a potential evolutionary adaptation. Ph.D. dissertation, University of Colorado at Boulder.
Gintis, H., van Schaik, C., & Boehm, C. (2015). Zoon Politikon. Current Anthropology, 56, 327–353.
Gonder, M. K., Locatelli, S., Ghobrial, L., Mitchell, M. W., Kujawski, J. T., et al. (2011). Evidence from Cameroon reveals differences in the genetic structure and histories of chimpanzee populations. PNAS, 108, 4766–4771.
Gros-Louis, J., Perry, S., & Manson, J. (2003). Violent coalitionary attacks and intraspecific killing in wild white-faced capuchin monkeys (Cebus capucinus). Primates, 44, 341–346.
Kaburu, S. S., Inoue, S., & Newton-Fisher, N. (2013). Death of the alpha: Within-community lethal violence among chimpanzees of the Mahale Mountains National Park. American Journal of Primatology, 75, 789–797.
King, B. J. (2013). How animals grieve. Chicago: University of Chicago Press.
Lindburg, D. G. (1971). The rhesus monkeys in north India: An ecological and behavioural study. In L. A. Rosenblum (Ed.), Primate behaviour: Developments in the field and laboratory research (pp. 83–104). New York: Academic.
Mitani, J., & Watts, D. (2005). Correlates of territorial boundary patrol behaviour in wild chimpanzees. Animal Behaviour, 70, 1079–1086.
Muller, M. N. (2002). Agonistic relations among Kanyawara chimpanzees. In C. Boesch, G. Hohmann, & L. F. Marchant (Eds.), Behavioural diversity in chimpanzees and bonobos (pp. 112–124). Cambridge: Cambridge University Press.
Nishida, T. (1996). Reports from the field: Mahale, Tanzania: the death of Ntologi, the unparalleled leader of M group [4]. Pan Africa News, 3(1), 3–4.
Nishida, T. (2012). Chimpanzees of the Lakeshore: Natural history and culture at Mahale. Cambridge: Cambridge University Press.
Nishida, T., Kano, T., Goodall, J., McGrew, W. C., Nakamura, M. (1999). Ethogram and ethnography of Mahale chimpanzees. Anthropological Science 107, 141–188.
Perry, S. (1998). A case report of a male rank reversal in a group of wild white-faced capuchins (Cebus capucinus). Primates, 39, 51–70.
Perry, S. (2008). Manipulative monkeys: The capuchins of Lomas Barbudal. Cambridge: Harvard University Press.
Phillips, T., Li, J., & Kendall, G. (2014). The effects of extra-somatic weapons on the evolution of human cooperation towards non-kin. PLoS ONE, 9, e95742.
Prado-Martinez, J., Sudmant, P. H., Kidd, J. M., Li, H., Kelly, J. L., et al. (2013). Great ape genetic diversity and population history. Nature, 499, 471–475.
Pruetz, J. D. (2006). Feeding ecology of savanna chimpanzees at Fongoli, Senegal. In C. Boesch, G. Hohmann, & M. Robbins (Eds.), The feeding ecology of great apes and other primates (pp. 161–182). Cambridge: Cambridge University Press.
Pruetz, J. D., & Kante, D. (2010). Successful return of a wild infant chimpanzee (Pan troglodytes verus) to its natal group after capture by poachers. African Primates, 7, 35–41.
Reimers, M., Schwarzenberger, F., & Preuschoft, S. (2007). Rehabilitation of research chimpanzees: stress and coping after long-term isolation. Hormones and Behavior, 51, 428–435.
Rose, L. (1998). Behavioral ecology of white-faced capuchins (Cebus capucinus) in Costa Rica. Ph.D. dissertation, Washington University in St. Louis.
Scarry, C. J., & Tujague, M. P. (2012). Consequences of lethal intragroup aggression and alpha male replacement on intergroup relations and home range use in tufted capuchin monkeys (Cebus paella nigritus). American Journal of Primatology, 74, 804–810.
Setchell, J. M., Knapp, L. A., & Wickings, E. J. (2006). Violent coalitionary attack by female mandrills against an injured alpha male. American Journal of Primatology, 68, 411–418.
Stewart, F. (2011). The evolution of shelter: Ecology and ethology of chimpanzee nest building. Ph.D. dissertation, University of Cambridge.
Stumpf, R. (2007). Chimpanzees and bonobos: Diversity within and between species. In C. J. Campbell, A. Fuentes, K. C. Mackinnon, M. Panger, & S. K. Bearder (Eds.), Primates in perspective. Oxford: Oxford University Press.
Talebi, M. G., Beltrao-Mendes, R., & Lee, P. C. (2009). Intra-community coalitionary lethal attack of an adult male southern muriqui (Brachyteles arachnoides). American Journal of Primatology, 7, 860–867.
Tokuyama, N., Emikey, B., Bafike, B., Isolumbo, B., Iyokango, B., et al. (2012). Bonobos apparently search for a lost member injured by a snare. Primates, 53, 215–219.
Valero, A., Schaffner, C. M., Vick, L. G., Aureli, F., & Ramos-Fernandez, G. (2006). Intragroup lethal aggression in wild spider monkeys. American Journal of Primatology, 68, 732–737.
van Leeuwen, E. J. C., Mulenga, I. C., Bodamer, M. D., & Cronin, K. A. (2016). Chimpanzees’ responses to the dead body of a 9-year-old group member. American Journal of Primatology. doi:10.1002/ajp.22560.
Watts, D. P. (2004). Intracommunity coalitionary killing of an adult male chimpanzee at Ngogo, Kibale National Park, Uganda. International Journal of Primatology, 25, 507–521.
Wilson, M. L., Boesch, C., Gilby, I. C., Hohmann, G., Itoh, N., et al. (2014). Rates of lethal aggression in chimpanzees depend on the number of adult males rather than measures of human disturbance. Nature, 513, 414–417.
Wrangham, R. W. (1999). Evolution of coalitionary killing. American Journal of Physical Anthropology, 110, 1–30.
Wrangham, R. W., & Peterson, D. (1996). Demonic males: Apes and the evolution of human aggression. Boston: Houghton Mifflin Harcourt.
Yamakoshi, G. (2004). Food seasonality and socioecology in Pan: are West African chimpanzees another bonobo? African Study Monographs, 25, 45–60.
Acknowledgements
Research was conducted with the permission of the Republic of Senegal and the Eaux et Forets of that country. Inspecteur Guisse provided assistance following the death of Foudouko per Fongoli Savanna Chimpanzee Project and Eaux et Foret protocol. Dondo Kante, Michel Sadiakho, Mboule Camara, and numerous students provided assistance in the field. Suggestions from three anonymous reviewers as well as the editor significantly improved this manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Joanna M. Setchell
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(DOCX 38 kb)
(MOV 41133 kb)
(MOV 34485 kb)
(MOV 18147 kb)
(MOV 41434 kb)
(MOV 40792 kb)
(MOV 40513 kb)
(MOV 9840 kb)
(MOV 22269 kb)
(MOV 4255 kb)
(MOV 3257 kb)
Rights and permissions
About this article

Cite this article
Pruetz, J.D., Ontl, K.B., Cleaveland, E. et al. Intragroup Lethal Aggression in West African Chimpanzees (Pan troglodytes verus): Inferred Killing of a Former Alpha Male at Fongoli, Senegal. Int J Primatol 38, 31–57 (2017). https://doi.org/10.1007/s10764-016-9942-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10764-016-9942-9