
Abstract
The anatomical characteristics and density of wood were examined in 23-year-old Acacia mangium trees that had been planted in Yogyakarta, Indonesia. The seeds had been collected from trees of five different provenances. The distance from the pith of the boundary between juvenile and mature wood was also examined to clarify the maturity of the wood. Lengths of wood fibers near the pith and the distance from the pith of the boundary between juvenile and mature wood differed significantly among provenances. By contrast, other anatomical characteristics of the wood such as fiber wall area, fiber wall thickness, fiber diameter, vessel lumen area, vessel diameter, vessel frequency and wood density did not differ significantly among provenances. Wood density was strongly correlated with the area of fiber walls. Our observations suggest that Sidei and Daintree might be more appropriate provenances among those examined for the Acacia mangium tree-breeding programs in Indonesia that are aimed at improving wood quality, because these provenances are associated with longer initial wood fibers and narrower juvenile areas than the other provenances studied.
Similar content being viewed by others

Introduction
Acacia mangium is one of the most important woody species in tropical and sub-tropical countries. Acacia mangium trees have been planted extensively because of their rapid growth, ability to adapt to soil with low fertility and relatively high acidity, the production of good-quality pulp and paper, the relative ease of cultivation and the positive response to selection efforts [1]. The trees grow naturally from the Moluccas and Irian Jaya in eastern Indonesia to the River Oriomo in western Papua New Guinea and the northeastern part of Australia [2]. In Indonesia, plantations of Acacia mangium were originally established for the production of pulp and paper. Recently, they have provided timber for solid wood, furniture and light construction and have, thereby, replaced the supply of logs from natural forests.
Prior to December 2008, there were 227 units of industrial plantation forest (IPF) with 4.31 × 106 hectares planted in Indonesia [3]. Efforts to improve the productivity of IPF have exploited intensive silvicultural techniques. The silvicultural efforts were initiated with the selection of appropriate species for planting [4]. Success in the establishment and productivity of forest tree plantations is determined largely by the appropriate choice of seeds within species [5]. In Indonesia, breeding of Acacia mangium trees has focused mainly on improving phenotypic features, such as survival rate, growth rate, fewer branches and stem form. However, the trees have been planted not only as a source of raw materials for the pulp industry, but also as a source of saw logs and, therefore, wood quality has become an important focus of breeding programs. In particular, the volume of juvenile wood should be minimized because of the low suitability of such wood for end products.
Wood density is one of the most widely used parameters of wood quality, because it is closely correlated with many physical and technological properties [6]. Wood density is determined by aspects of wood structure, such as cell size and cell wall thickness. Therefore, variations in the structure of wood have a significant impact on the quality and yield of pulp and paper products, and on the strength and utility of solid wood products.
Significant differences among the properties and anatomical characteristics of wood have been recognized among provenances [6–10]. Therefore, differences in provenance might be expected to be closely related to the properties of wood. However, only limited information is available about differences in the properties of wood from Acacia mangium trees of different provenances. In this study, we examined radial variations in the anatomical characteristics and density of wood of Acacia mangium trees that had been grown from seeds of five different provenances, under identical conditions. The present results provide useful information for the selection of provenances for tree-breeding programs in Indonesia.
Materials and methods
Plant materials
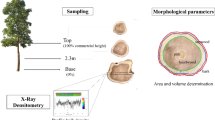
The materials were taken from the “Provenances Trial Site” in compartment no. 17 of Wanagama 1 Forest, an educational forest of the Faculty of Forestry, Gadjah Mada University, Yogyakarta, Indonesia. The Wanagama 1 Forest is located in Gunungkidul Regency, Yogyakarta Province, in the central part of Java (110° 30′ 38″–110° 33′ 03″ E and 7° 53′ 25″–7° 54′ 52″ S). Seeds were collected from several different provenances and seedlings were planted in 1984 under identical growth conditions with 4 × 4 m spacing in a 0.35-ha area. Five straight trees, grown from seeds from each of the five provenances of Acacia mangium trees, were examined. The provenances were Sidei provenance (Papua, Indonesia), West of Morehead provenance (Papua New Guinea), Daintree provenance (Queensland, Australia), Ellerbeck Rd. Cardwell provenance (Queensland, Australia) and El Arish provenance (Queensland, Australia). Twenty-five trees were felled in April 2007. The gross parameters of sample trees are shown in Table 1. Disks of wood were removed from main stems at the equivalent of 2 m above the ground and then a disk of wood from each tree was used for the measurements of the wood fiber length, characteristics of wood anatomy and wood density. Radial blocks were removed from each disk in both the north-facing and south-facing directions of original stems. Each radial block was divided into 2 parts, and then we used one block for the measurements of the wood fiber length and characteristics of wood anatomy and another one for the measurement of wood density.
Measurements of wood fiber length
Small pieces were removed continuously at 1-cm intervals, from pith to bark, from each radial block in both the north-facing and south-facing directions of original stems. The pieces of wood were macerated with Franklin solution, which is a mixture of equal volumes of glacial acetic acid and hydrogen peroxide, and heated at 60°C for 1 or 2 days [11]. Images of wood fibers were recorded under a light microscope (Axioscop; Carl Zeiss, Oberkochen, Germany) with a digital camera (Nikon Digital Sight DS-5M-L1; Nikon Corporation, Japan). Lengths of wood fibers were measured with image-analysis software (Image-J; National Institutes of Health, MD, USA). Fifty wood fibers in each sample were measured at random.
Determination of the boundary between juvenile and mature wood
The boundary between juvenile and mature wood is usually defined by the lengths of vessel elements, which are very similar to those of fusiform cambial cells [11, 12]. However, vessel elements in most trees of Acacia mangium were reported previously to be of constant lengths, 0.2 mm, across the stem [13]. Kim et al. [14] reported that lengths of vessel elements were also relatively constant in the radial direction in an Acacia hybrid planted in Vietnam. In the present study, we examined the radial variation in lengths of wood fibers to determine the boundary between juvenile and mature wood. Fujiwara and Yang [15] proposed that the juvenile area should be defined as the zone in which cell length undergoes a rapid increase and that mature wood area should be defined as the zone in which cell length eventually reaches a constant value toward the bark. Honjo et al. [13], in Acacia mangium, defined the boundary between juvenile and mature wood as the area in which the rate of increase in the lengths of wood fibers (∆D) reached 0.3%. Therefore, we defined the boundary as the point where the rate of increase reached 0.3%, as proposed by Honjo et al. [13]. Variations in lengths of wood fibers were approximated by a logarithmic relationship to the distance from the pith. We calculated ∆D from the logarithmic curve as described by Shiokura [16] and Honjo et al. [13]. This method is illustrated in Fig. 1. Furthermore, we measured proportion of mature wood area according to the data of boundary between juvenile and mature wood area. The mature wood proportion was calculated as the ratio between mature wood area and total area of disk for each individual tree and expressed as percentage.

Radial variation in length of wood fibers and its logarithmic curve for the Sidei provenance tree no. 1. The boundary between juvenile and mature wood was defined as the point where the rate of increase in lengths of wood fibers (∆D) reached 0.3%
Observation and measurement of characteristics of wood anatomy
Since it is difficult to section the wood of Acacia mangium on a microtome, we softened samples prior to preparation of transverse sections. We boiled radial blocks of wood in a solution of glycerin in water (20%) for approximately 10 h, and then we divided the blocks into 1 × 1 × 1 cm3 blocks at 1-cm intervals from the pith to the bark. Then transverse sections of 15- to 20-μm thickness were sectioned on a sliding microtome (Yamatokohki, Saitama, Japan). The transverse sections were stained with 1% safranin solution, mounted on glass slides, fixed with resin (Entellan New; Merck, Darmstadt, Germany) and covered with coverslips [17]. Images were recorded from each slide under the light microscope with the digital camera. Three digital images of transverse-sectional area of 35.3 × 103 μm2 were taken in each radial position to measure the fiber wall area (%), fiber wall thickness and fiber diameter. Fiber wall area (%) was defined as the ratio of total area of fiber wall per total area of measured image. Rays were excluded from total area of measured image. For the measurements of vessel lumen area (%), vessel diameter and vessel frequency (number of vessels/mm2), three digital images of transverse-sectional area of 8.1 × 106 μm2 from each radial position were observed. The percentage of vessel lumen area was determined by calculating the ratio of total area of vessel lumens per total area of observed image. Fiber wall area (%), fiber wall thickness, fiber diameter, vessel lumen area (%), vessel diameter and vessel frequency (number of vessels/mm2) were determined with Image-J software.
Observation and measurement of wood density
The radial blocks from pith to bark in the north-facing and south-facing directions were trimmed into strips that were approximately 1.57-mm thick (longitudinally) with a purpose-designed twin-blade saw [18]. Strips were air dried, under restraining conditions to prevent warping, in a thermostatically controlled chamber (air temperature, 20 ± 2°C; and relative humidity, 65%) for a week. Negative X-ray images of strips were recorded with a soft X-ray apparatus (Softex Co., Tokyo, Japan) on industrial X-ray films (Fuji Film Co., Ltd., Tokyo). The conditions for soft X-ray measurements were as follows: voltage 20 kVp; electric current 14 mA; lighting distance 2.2 m; and duration of irradiation for photography, 4 min. Films were washed several times and dried at room temperature in air. To measure wood density, we scanned the negative X-ray films in the radial direction using DENDRO2003 X-ray analysis instrumentation (Walesch Electronics, Effretikon, Switzerland) and collected density data at 10-μm intervals [19]. Wood densities near the pith, juvenile wood and mature wood were obtained from densitometric profiles.
Analysis of data
Analysis of variance was conducted to determine the significant differences of data on the gross parameter of sample trees, length of wood fibers, distance of boundary between juvenile and mature wood from pith, mature wood proportion, anatomical characteristics of wood and wood density among provenances. The length of wood fiber was analyzed at several positions on radial direction of disks, namely position near pith (1 cm from pith), boundary between juvenile and mature wood area, and near bark (1 cm from bark). For the anatomical characteristics of wood and wood density, data analysis was conducted at the position near the pith, juvenile wood area, and mature wood area.
Statistical analyses were performed using SPSS statistical software (SPSS base 11.5 for Windows; SPSS Inc., Chicago, IL, USA). The data of each measured parameter were analyzed using one-way analysis of variance (ANOVA) followed by Tukey’s post hoc test with the level of significant differences at P < 0.05. Linear regression analyses were carried out to analyze the relationship between the anatomical characteristics and the density of wood.
Results and discussion
Lengths of wood fibers
The lengths of wood fibers of all provenances exhibited very similar radial variations. The wood fibers were shortest near the pith. They increased in length rapidly from near the pith to approximately 5 cm from the pith and then remained constant toward the bark (Fig. 2). A similar radial pattern was reported in Acacia mangium grown in Malaysia and Indonesia by Honjo et al. [13]. Kim et al. [20] reported that in an Acacia hybrid grown in Vietnam, the lengths of wood fibers were shortest near the pith and increased toward the bark. The wood fibers of several hardwoods are shortest near the pith and then their lengths increase rapidly and nonlinearly, reflecting a prominent characteristic of juvenile wood [15, 21].

Radial variations in mean fiber length in Acacia mangium trees from five provenances (five trees for each provenance)
The lengths of wood fibers of all provenances examined ranged from 0.53 to 0.61 mm near the pith and from 1.04 to 1.08 mm near the bark (Table 2). Honjo et al. [13] reported that, in Acacia mangium from Malaysia and Indonesia, the wood fibers in the vicinity of the pith were 0.4- to 0.6-mm long and varied in length between 0.9 and 1.3 mm near the bark. In Acacia hybrid grown in Vietnam, the lengths of wood fibers ranged at around 0.5 mm near the pith and from 1.0 to 1.2 mm near the bark [20]. We found a significant difference (P = 0.001) in fiber lengths among provenances near the pith. Such a difference among lengths of wood fibers near the pith might be due to genetic factors. The Sidei and Daintree provenances had longer wood fibers near pith than the others. By contrast, lengths of wood fibers did not differ significantly among provenances in the boundary between juvenile and mature wood area (P = 0.584), near the bark (P = 0.494) and total average (P = 0.072). The pulp and paper industries require long wood fibers. Therefore, in an Acacia mangium breeding program, longer fiber length is one of the goals of improvements in wood quality. The Sidei and Daintree provenance might be more appropriate provenances for an Acacia mangium breeding program in Indonesia to increase the lengths of wood fibers in juvenile wood near the pith.
The boundary between juvenile and mature wood
We found that the lengths of wood fibers became stable at approximately 49 mm from the pith, where the increases in fiber length (∆D) were 0.3%. The distance of the boundary between juvenile and mature wood in all provenances examined ranged from 45.8 to 52.0 mm from the pith (Table 2).
The mechanical properties of juvenile wood are poorer than those of mature wood [22–25]. Therefore, the site of the border between juvenile and mature wood is an important determinant of wood quality. For tree-breeding programs focused on timber or saw logs from Acacia mangium, the selection of trees with minimized juvenile wood is essential. The distance from the pith of the boundary between juvenile and mature wood differed significantly among provenances (P = 0.016). The Daintree and Sidei provenances had narrower juvenile regions than the Ellerbeck Rd. Cardwell, West of Morehead and El Arish provenances. Therefore, these seed provenances might be good candidates for Acacia mangium tree-breeding programs. By contrast, there were no significant differences in terms of DBH (P = 0.733), tree height (P = 0.093), height of free bole (P = 0.932) and proportion of mature wood area (P = 0.985) among provenances (Tables 1, 2).
Figure 3 shows that the diameter of breast height (DBH) had no correlation with the distance of boundary between juvenile and mature wood from pith (P = 0.402). This result indicates that the distance of boundary between juvenile and mature wood from pith is not affected by the diameter of stem. Acceleration of radial growth might be useful for Acacia mangium tree-breeding programs to produce wider mature wood.

Relationship between diameter of breast height (DBH) and distance of boundary between juvenile and mature wood from pith. The DBH had no correlation with the distance of boundary between juvenile and mature wood from pith
Anatomical characteristics of wood
In general, wood density is a key indicator of wood quality. In hardwoods, wood density is determined by cell wall thickness, cell diameter and the frequency of vessel elements [26]. Thus, the anatomical characteristics of wood are closely related to the mechanical properties of wood. Parameters of wood fibers, such as fiber length, fiber wall thickness and fiber lumen diameter are important for pulp and paper production, because the physical properties of sheets made from hardwood pulp fibers are dependent on the characteristics of the fibers. The thickness of the fiber wall is an important indicator of most properties of paper, because thick-walled fibers are associated with high tearing strength.
In all studied provenances, the fiber wall area (%) and fiber wall thicknesses were lowest near the pith and increased from the pith to the bark side of the stem (Fig. 4a, b). There were no significant differences in terms of both fiber wall area (%) and fiber wall thickness among provenances in the region near the pith (P = 0.511 and P = 0.478), in the juvenile (P = 0.648 and P = 0.322), in mature wood (P = 0.867 and P = 0.711), and in total averages (P = 0.939 and P = 0.880). Fiber walls in the juvenile wood were thinner than in the mature wood. Fiber wall thicknesses in all studied provenances ranged from 1.1 to 1.3 μm near the pith, from 1.6 to 1.9 μm in the juvenile wood, and from 2.5 to 2.6 μm in the mature wood (Table 2). It was reported previously that the average thickness of fiber walls in Acacia mangium ranged from 1.5 to 2.5 μm [27], while the average of thickness of fiber walls of 8-year-old Acacia mangium grown in Malaysia was around 4.5 μm at breast height [28]. Differences among thicknesses of fiber walls might be due to genetic aspects of planted trees or to differences among site conditions.

Radial variations in mean fiber wall area percentage (a), fiber wall thickness (b) and fiber diameter (c) in Acacia mangium trees from five provenances (five trees for each provenance)
In contrast to fiber wall thickness, the diameters of wood fibers of all studied provenances decreased slightly from the pith to the bark side (Fig. 4c). The diameters of wood fibers of all observed provenances ranged from 26.5 to 28.7 μm near the pith, from 24.9 to 26.4 μm in the juvenile wood, and from 23.1 to 24.2 μm in the mature wood (Table 2). There were no significant differences among provenances in the region near the pith (P = 0.423), in the juvenile (P = 0.656), in mature wood (P = 0.568) and in the total average (P = 0.525) in terms of diameters of wood fibers.
Radial variations among vessel lumen area (%) and among diameters of vessels increased from the pith toward the bark side in all provenances (Fig. 5a, b). Images of transverse sections of Sidei provenance near the pith (Fig. 6a), in the intermediate region (Fig. 6b) and near the bark (Fig. 6c) showed that vessel size depended on the position within the stem. A similar pattern was observed in Eucalyptus camaldulensis, wherein vessel size increased from the pith to the bark side [29]. In all provenances that we examined, vessel lumen area (%) and the diameter of vessels ranged from 8.4 to 10.0% and 115 to 129 μm in the juvenile, and from 13.8 to 17.1% and 146 to 155 μm in the mature wood, respectively (Table 2). Similarly, in Acacia mangium grown in Malaysia, the average diameters of vessels at breast height ranged from 91 to 188 μm [28]. Ogata et al. [27] reported that the maximum tangential diameter of solitary vessels in Acacia mangium ranged from 150 to 280 μm. There were no significant differences among provenances in terms of vessel lumen area (%) and the diameter of vessels near the pith (P = 0.579 and P = 0.847), in the juvenile wood (P = 0.489 and P = 0.867), in mature wood (P = 0.570 and P = 0.978) and in the total averages (P = 0.411 and P = 0.982), respectively.

Radial variations in mean vessel lumen area percentage (a), vessel diameter (b), number of vessels/mm2 (c) and wood density (d) in Acacia mangium trees from five provenances (five trees for each provenance)

Light micrographs of transverse sections, stained with safranin, of Sidei provenance near the pith (a), the intermediate region of the disk (b) and near the bark (c) showing that vessel size depended on the position within the stem. Bars 500 μm
Unlike the radial variations in vessel lumen area (%) and the diameter of vessels, vessel frequency was almost constant from the pith toward the bark side in all provenances (Fig. 5c). The vessel number in all provenances ranged from 7.9 to 9.8/mm2 near the pith, from 7.1 to 8.5/mm2 in the juvenile wood, and from 8.0 to 10.0/mm2 in the mature wood (Table 2). There were no significant differences among provenances in the region near the pith (P = 0.774), in the juvenile (P = 0.424), in mature wood (P = 0.684) and in the total average (P = 0.318) in terms of respective vessel frequencies.
Wood density
As shown in Fig. 5d, wood density in all studied provenances increased from the pith toward the bark side. Kim et al. [14, 20] found, in Acacia hybrid trees in Vietnam, a similar radial variation in wood density from the pith to the bark side. The wood density in all our observed provenances ranged from 0.34 to 0.43 g/cm3 near the pith, from 0.48 to 0.53 g/cm3 in the juvenile wood and from 0.60 to 0.66 g/cm3 in the mature wood (Table 2). Chowdhury et al. [30] reported that the wood density of Acacia mangium grown in Bangladesh ranged from 0.50 to 0.62 g/cm3. In this study, wood densities did not differ significantly among provenances near the pith (P = 0.200), in the juvenile wood (P = 0.695), in the mature wood (P = 0.973) and in the total average (P = 0.327). Thus, on the basis of wood density, all of the observed provenances are acceptable for Acacia mangium tree-breeding programs in Indonesia.
Table 3 shows the correlation coefficients for the relationships between the anatomical characteristics and the density of wood. Fiber wall area (%) was more closely correlated with wood density than vessel lumen area (%). Thus, wood density is mainly influenced by fiber wall area (%) in Acacia mangium. In addition, fiber wall area (%) was more strongly correlated with fiber wall thickness than with fiber diameter. Therefore, trees with wood fibers with thicker cell walls are good targets for selection of higher-density wood. In terms of vessel lumen area (%), vessel diameter was more closely correlated than vessel frequency near the pith, but less closely correlated in the mature wood.
Conclusion
Many parameters of Acacia mangium trees grown in the same forest exhibited no significant differences among original provenances. However, there were significant differences in terms of the lengths of wood fibers near the pith and in the extent of the juvenile wood among the five original provenances. The Sidei and Daintree provenance had longer initial wood fibers and a narrower juvenile region than the other provenances examined. Therefore, Sidei and Daintree might be more appropriate provenances for breeding programs focused on improving wood quality of Acacia mangium in the central region of Java, Indonesia.
References
Hardiyanto EB (2004) Silviculture and breeding of Acacia mangium (in Indonesian). In: Hardiyanto EB, Arisman H (eds) Development of Acacia mangium plantation forest—experience in PT. Musi Hutan Persada, South Sumatera. PT. Musi Hutan Persada, pp 207–281
Eldoma A, Awang K (1999) Site adaptability of Acacia mangium, Acacia auriculiformis, Acacia crassicarpa and Acacia aulocarpa. APAFRI Publication Series no. 3
Indonesian Ministry of Forestry (2009) Forestry statistics of Indonesia 2008. Ministry of Forestry
Soekotjo (2004) Silviculture of plantation forest: basic principles (in Indonesian). In: Hardiyanto EB, Arisman H (eds) Development of Acacia mangium plantation forest—experience in PT. Musi Hutan Persada, South Sumatera. PT. Musi Hutan Persada, pp 143–206
Zobel B, Talbert J (1984) Applied forest tree improvement. Wiley, New York
Miranda I, Almeida MH, Pereira H (2001) Influence of provenance, subspecies and site on wood density in Eucalyptus globulus labill. Wood Fiber Sci 33:9–15
Bayramzadeh V, Funada R, Kubo T (2008) Relationships between vessel element anatomy and physiological as well as morphological traits of leaves in Fagus crenata seedlings originating from different provenances. Trees Struct Funct 22:217–224
Bhat KM, Priya PB (2004) Influence of provenance variation on wood properties of teak wood from the Western Ghat region in India. IAWA J 25:73–282
Liu J, Noshiro S (2003) Lack of latitudinal trends in wood anatomy of Dodonaea viscose (Sapindaceae), a species with a worldwide distribution. Am J Bot 90:532–539
Miranda I, Pereira H (2002) Variation of pulpwood quality with provenances and site in Eucalyptus globulus. Ann For Sci 59:283–291
Kitin P, Funada R, Sano Y, Beeckman H, Ohtani J (1999) Variation in the lengths of fusiform cambial cells and vessel elements in Kalopanax pictus. Ann Bot 84:621–632
Bailey IW (1920) The cambium and its derivative tissues. II. Size variations of cambial initials in gymnosperms and angiosperms. Am J Bot 7:355–367
Honjo K, Furukawa I, Sahri MH (2005) Radial variation of fiber length increment in Acacia mangium. IAWA J 26:339–352
Kim NT, Ochiishi M, Matsumura J, Oda K (2008) Variation in wood properties of six natural acacia hybrid clones in northern Vietnam. J Wood Sci 54:436–442
Fujiwara S, Yang KC (2000) The relationships between cell length and ring width and circumferential growth rate in five Canadian species. IAWA J 21:335–345
Shiokura T (1982) Extent and differentiation of the juvenile wood zone in coniferous tree trunks. Mokuzai Gakkaishi 28:85–90 (in Japanese)
Begum S, Nakaba S, Oribe Y, Kubo T, Funada R (2011) Changes in the localization and levels of starch and lipids in cambium and phloem during cambial reactivation by artificial heating of main stems of Cryptomeria japonica trees. Ann Bot 106:85–895
Yasue K, Funada R, Kobayashi O, Ohtani J (2000) The effects of tracheid dimensions on variations in maximum density of Picea glehnii and relationships to climatic factors. Trees Struct Funct 14:223–229
Kataoka Y, Kiguchi M, Fujiwara T, Evans PD (2005) The effects of within-species and between-species variation in wood density on the photodegradation depth profiles of sugi (Cryptomeria japonica) and hinoki (Chamaecyparis obtusa). J Wood Sci 51:531–536
Kim NT, Matsumura J, Oda K (2011) Effect of growing site on the fundamental wood properties of natural hybrid clones of Acacia in Vietnam. J Wood Sci 57:87–93
Bonham VA, Barnett JR (2001) Fibre length and microfibril angle in silver birch (Betula pendula Roth). Holzforschung 55:159–162
Bao FC, Jiang ZH, Jiang XM, Lu XX, Luo XQ, Zhang SY (2001) Differences in wood properties between juvenile wood and mature wood in 10 species grown in China. Wood Sci Tech 35:363–375
Bhat KM, Priya PB, Rugmini P (2001) Characterization of juvenile wood in teak. Wood Sci Tech 34:517–532
Evans JW, Senft JF, Green DW (2000) Juvenile wood effect in red alder: analysis of physical and mechanical data to delineate juvenile and mature wood zones. For Prod J 50(7/8):75–87
Haygreen JG, Boywer JL (1996) Forest products and wood sciences—an introduction, 3rd edn. IOWA State University Press/AMES, Oxford
Thomas DS, Montagu KD, Conroy JP (2004) Changes in wood density of Eucalyptus camaldulensis due to temperature—the physiological link between water viscosity and wood anatomy. For Ecol Manag 193:157–165
Ogata K, Fujii T, Abe H, Bass P (2008) Identification of the timbers of Southeast Asia and the Western Pacific. Forestry and Forest Product Research Institute, Kaisesha Press, Ohtsu, Japan
Sahri MH, Ibrahim FH, Shukor NAA (1993) Anatomy of Acacia mangium grown in Malaysia. IAWA J 14:245–251
Veenin T, Fujita M, Nobuchi T, Siripatandilok S (2005) Radial variation of anatomical characteristics and specific gravity in Eucayptus camaldulensis clones. IAWA J 26:353–361
Chowdhury MQ, Shams MI, Alam M (2005) Effect of age and height variation on physical properties of mangium (Acacia mangium Willd.) wood. Austr For 68:17–19
Acknowledgments
Widyanto Dwi Nugroho thanks the Hitachi Scholarship Foundation Japan for its support. This work was supported, in part, by Grants-in-Aid for Scientific Research from the Ministry of Education, Science, Sports and Culture of Japan (nos. 20120009, 21380107 and 2200104). This work was also supported by New Energy and Industrial Technology Development Organization (NEDO) in Japan through the Program of Development of Preparatory Basic Bioenergy Technologies.
Author information
Authors and Affiliations
Corresponding author
About this article
Cite this article
Nugroho, W.D., Marsoem, S.N., Yasue, K. et al. Radial variations in the anatomical characteristics and density of the wood of Acacia mangium of five different provenances in Indonesia. J Wood Sci 58, 185–194 (2012). https://doi.org/10.1007/s10086-011-1236-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10086-011-1236-4