
Abstract
For structured populations with an annual breeding season, life-stage interactions and behavioral tactics may occur on a faster time scale than that of population dynamics. Motivated by recent field studies of the effect of rising sea surface temperature (SST) on within-breeding-season behaviors in colonial seabirds, we formulate and analyze a general class of discrete-time matrix models designed to account for changes in behavioral tactics within the breeding season and their dynamic consequences at the population level across breeding seasons. As a specific example, we focus on egg cannibalism and the daily reproductive synchrony observed in seabirds. Using the model, we investigate circumstances under which these life history tactics can be beneficial or non-beneficial at the population level in light of the expected continued rise in SST. Using bifurcation theoretic techniques, we study the nature of non-extinction, seasonal cycles as a function of environmental resource availability as they are created upon destabilization of the extinction state. Of particular interest are backward bifurcations in that they typically create strong Allee effects in population models which, in turn, lead to the benefit of possible (initial condition dependent) survival in adverse environments. We find that positive density effects (component Allee effects) due to increased adult survival from cannibalism and the propensity of females to synchronize daily egg laying can produce a strong Allee effect due to a backward bifurcation.






Similar content being viewed by others
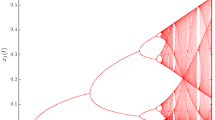
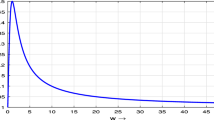
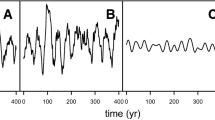
Notes
By \(a\lessapprox b\) is meant both \(a<b\) and \(\left| a-b\right| \) is small. Similarly, \(a\gtrapprox b\) means both \(a>b\) and \(\left| a-b\right| \) is small.
References
Alvarez-Fernandez S, Licandro P, van Damme CJG, Hufnagl M (2015) Effect of zooplankton on fish larval abundance and distribution: a long-term study on North Sea herring (Clupea harengus). ICES J Mar Sci 72(9):2569–2577
Barber RT, Chavez FP (1983) Biological consequences of El Niño. Science 222:1203–1210
Blight LK (2011) Egg production in a coastal seabird, the Glaucous-winged Gull (Larus glaucescens), declines during the last century. PLoS ONE 6(7):1–8
Burton D, Henson SM (2014) A note on the onset of synchrony in avian ovulation cycles. J Differ Equ Appl 20:664–668
Caswell H (2001) Matrix population models: construction, analysis, and interpretation, 2nd edn. Sinauer Associates, Sunderland
Caswell H, Trevisan MC (1994) Sensitivity analysis of periodic matrix models. Ecology 75:1299–303
Cushing JM (2014) Backward bifurcations and strong Allee effects in matrix models for the dynamics of structured populations. J Biol Dyn 8:57–73
Cushing JM (2016) One dimensional maps as population and evolutionary dynamic models. In: Cushing JM, Saleem M, Srivastava HM, Khan MA, Merajuddin M (eds) Applied analysis in biological and physical sciences. Springer proceedings in mathematics & statistics, vol 186. Springer, India, pp 41–62
Cushing JM, Henson SM, Hayward JL (2015) An evolutionary game theoretic model of cannibalism. Nat Resour Model 28(4):497–521
Galusha JG, Vorvick B, Opp M, Vorvick P (1987) Nesting season censuses of seabirds on Protection Island, Washington. Murrelet 68:103–107
Hayward JL, Verbeek NA (2005) Glaucous-winged gull (Larus glaucescens). In: Poole A (ed) The birds of North America. Cornell Laboratory of Ornithology, Ithaca
Hayward JL, Weldon LM, Henson SM, Megna LC, Payne BG, Moncrieff AE (2014) Egg cannibalism in a gull colony increases with sea surface temperature. Condor Ornithol Appl 116:62–73
Henson SM, Hayward JL, Cushing JM, Galusha JG (2010) Socially induced synchronization of every-other-day egg laying in a seabird colony. Auk 127:571–580
Henson SM, Cushing JM, Hayward JL (2011) Socially-induced ovulation synchrony and its effect on seabird population dynamics. J Biol Dyn 5:495–516
Irvine JR, Crawford WR (2011) State of the ocean report for the Pacific North Coast Integrated Management Area (PNCIMA). Fisheries and Oceans Canada, Science Branch, Pacific Region, Pacific Biological Stations, Nanaimo, BC
Irvine JR, Crawford WR (2013) State of physical, biological, and selected fishery resources of Pacific Canadian marine ecosystems in 2012. Research Document 2013/032. Canadian Science Advisory Secretariat, Fisheries and Oceans Canada, Pacific Region, Ottawa, ON
Kershner J, Samhouri JF, James CA, Levin PS (2011) Selecting indicator portfolios for marine species and food webs: a Puget Sound case study. PLoS ONE 6:e25248
Kielhöfer H (2004) Bifurcation theory: an introduction with applications to PDEs. Applied mathematical sciences, vol 156. Springer, New York
Lesnoff M (1999) Dynamics of a sheep population in a Sahelian area (Ndiagne district in Senegal): a periodic matrix model. Agric Syst 61:207–221
Lesnoff M, Lancelot R, Tillard E, Dohoo IR (2000) A steady-state approach of benefit-cost analysis with a periodic Leslie-matrix model: presentation and application to the evaluation of a sheep-diseases preventive scheme in Kolda, Senegal. Prev Vet Med 46:113–128
McGowan JA, Cayan DR, Dorman LM (1998) Climate ocean variability and ecosystem response in the northeast Pacific. Science 281:210–217
Mertens SK, Yearsley JM, van den Bosch F, Gilligan CA (2006) Transient population dynamics in periodic matrix models: methodology and effects of cyclic permutations. Ecology 87(9):2338–2348
Pearson SF, Hodum PJ, Good TP, Schrimpf M, Knapp SM (2013) A model approach for estimating colony size, trends, and habitat associations of burrow-nesting seabirds. Condor 115:356–365
Sandberg S, Awerbuch TE, Spielman A (1992) A comprehensive multiple matrix model representing the life cycle of the tick that transmits the agent of lyme disease. J Theor Biol 157:203–220
Sarukhan J, Gadgil M (1974) Studies on plant demography: Ranunculus repens L., R. bulbosus L. and R. acris L. Ill. A mathematical model incorporating multiple modes of reproduction. J Ecol 62:921–936
Schreiber S (2003) Allee effects, extinctions, and chaotic transients in simple population models. Theor Popul Biol 64:201–209
Schreiber RW, Schreiber EA (1984) Central Pacific seabirds and the El Niño Southern Oscillation: 1982 to 1983 perspectives. Science 225:713–716
Smith RS, Weldon LM, Hayward JL, Henson SM (2017) Time lags associated with effects of oceanic conditions on seabird breeding in the Salish Sea region of the northern California Current system. Mar Ornithol 45:39–42
Stenseth NC, Mysterud A, Ottersen G, Hurrell JW, Chan K-S, Lima M (2002) Ecological effects of climate fluctuations. Science 297:1292–1296
Strom A, Francis RC, Mantua NJ, Miles EL, Peterson DL (2004) North Pacific climate recorded in growth rings of geoduck clams: a new tool for paleoenvironmental reconstruction. Geophys Res Lett. https://doi.org/10.1029/2004GL019440
Veprauskas A, Cushing JM (2017) A juvenile-adult population model: climate change, cannibalism, reproductive synchrony, and strong Allee effects. J Biol Dyn 11:1–24
Vermeer K (1963) The breeding ecology of the glaucous-winged gull (Larus glaucescens). Occasional Papers of the British Columbia Provincial Museum No. 13, British Columbia Provincial Museum, Victoria, BC
Weir SK (2015) Ovulation synchrony as an adaptive response to egg cannibalism in a seabird colony. Honors thesis, Andrews University. http://digitalcommons.andrews.edu/honors/120/. Accessed 31 Jan 2018
Acknowledgements
We thank James L. Hayward for field work collaboration and discussions; Jennifer Brown-Scott, Lorenz Sollmann, and Sue Thomas, Washington Maritime National Wildlife Refuge Complex, for permission to work on Protection Island National Wildlife Refuge; and Rosario Beach Marine Laboratory for logistical support. We thank two anonymous reviewers for their careful reading of the manuscript and their suggestions for its revision. This research was supported by the U.S. National Science Foundation Grants DMS-1407564 (JMC) and DMS-1407040 (SMH).
Author information
Authors and Affiliations
Corresponding author
Appendix
Appendix
The following Lemma is an extension of results in Cushing (2016).
Lemma 1
Assume A1 and A2.
(a) Suppose \(\partial _{p}^{0}r>0\). Then the equilibrium \(\alpha =0\) of the difference equation (12) is locally asymptotically stable if \(p<p_{0}\) and is unstable if \(p>p_{0}\). Suppose, on the other hand, that \(\partial _{p}^{0}r<0\). Then \(\alpha =0\) is locally asymptotically stable for \(p>p_{0}\) and is unstable for \(p<p_{0}\).
(b) On an open interval I of \(p_{0}\) there exists a (twice continuously differentiable) continuum of equilibria \(\alpha =\alpha \left( p\right) \) of the difference equation (12) satisfying \(\alpha \left( p_{0}\right) =0\) and \(\alpha \left( p\right) \ne 0\) for \(p\ne p_{0}\).
(c) Suppose \(\partial _{p}^{0}r>0.\) If \(\partial _{\alpha }^{0}r<0\) then for \(p\in I\) the equilibrium \(\alpha \left( p\right) \) is positive and locally asymptotically stable if \(p>p_{0}\) (and are negative for \(p<p_{0}\)). On the other hand, if \(\partial _{\alpha }^{0}r>0\) then for \(p\in I\) the equilibrium \(\alpha \left( p\right) \) is positive and unstable for \(p<p_{0}\) (and are negative for \(p>p_{0}\)).
(d) Suppose \(\partial _{p}^{0}r<0.\) If \(\partial _{\alpha }^{0}r>0\) then for \(p\in I\) the equilibrium \(\alpha \left( p\right) \) is positive and locally asymptotically stable for \(p>p_{0}\) (and are negative for \(p<p_{0}\)). On the other hand, if \(\partial _{\alpha }^{0}r<0\) then for \(p\in I\) the equilibrium \(\alpha \left( p\right) \) is positive and unstable for \(p>p_{0}\) (and are negative for \(p>p_{0}\)).
Proof
(a) The linearization principle guarantees local asymptotic stability of \(\alpha =0\) if \(r\left( 0,p\right) <1\) and instability if \(r\left( 0,p\right) >1\).
(b) Under assumption A2 we can apply the implicit function theorem to the equation \(r\left( p,\alpha \right) =1\) for nontrivial equilibria of the difference equation (12) and obtain, on an open interval of \(p_{0}\), a twice continuously differentiable function \(\alpha =\alpha \left( 0\right) ,\) \(\alpha \left( p_{0}\right) =0\) satisfying \(r\left( p,\alpha \left( p\right) \right) =1.\) An implicit differentiation with respect to p yields \(\alpha ^{\prime }\left( 0\right) =-\partial _{p}^{0}r/\partial _{\alpha }^{0}r\ne 0\) from which it follows that \(\alpha \left( p\right) \ne 0\) for \(p\ne p_{0}\).
(c) If \(\alpha ^{\prime }\left( 0\right) >0\) then \(\alpha \left( p\right) >0\) (respectively \(\alpha \left( p\right) <0\)) for \(p\gtrapprox p_{0}\) (respectively \(p\lessapprox p_{0}\)). On the other hand, if \(\alpha ^{\prime }\left( 0\right) <0\) then \(\alpha \left( p\right) >0\) (respectively \(\alpha \left( p\right) <0\)) for \(p\lessapprox p_{0}\) (respectively \(p\gtrapprox p_{0}\)) as asserted. With regard to the stability of the equilibrium \(\alpha \left( p\right) \) for p near \(p_{0},\) we apply the linearization principle by calculating
Assume \(\partial _{p}^{0}r>0\). If \(\partial _{\alpha }^{0}r<0\) then \(\alpha ^{\prime }\left( 0\right) =-\partial _{p}^{0}r/\partial _{\alpha } ^{0}r>0\) and \(\alpha \left( p\right) \) is positive and \(\left| \lambda \right| <1\) for \(p\gtrapprox p_{0}\). If \(\partial _{\alpha }^{0}r>0\) then \(\alpha ^{\prime }\left( 0\right) =-\partial _{p}^{0}r/\partial _{\alpha }^{0}r<0\) and \(\alpha \left( p\right) \) is positive and \(\left| \lambda \right| <1\) for \(p\lessapprox p_{0}\).
(d) These conclusions are derived in closely similar manner to those in (c) by also making use of (24). \(\square \)
Proof of Corollary 1
It is obvious from the definition of \(\hat{x}\left( t,\alpha \right) \) that \(\hat{x}\left( t,0\right) =\hat{0}\) for all t. From (10) we have
where
By the chain rule
where
From
we obtain
for \(t=0,1,\ldots ,k-1.\) The matrix
leads to
The entries in the matrix \(D\left( t\right) \) are linear combinations of the derivatives \(\partial _{x_{l}}^{0}w_{ij}\). It follows that the same is true of the entries in the matrix \(\partial _{\alpha }^{0}Q\) and, as a result, the same is true of \(\partial _{\alpha }^{0}r\). \(\square \)
The quantity \(\partial _{\alpha }^{0}r\) for the gull model (17)–(19) with (18). We use the formula
where
with matrix W defined by (17) and entries (18) and with the vectors \(\hat{\nu }^{\tau }\) and \(\hat{a}\) given by (20). By the product rule
where
since
implies
From the gull model matrix entries (18) we have
Let \(M_{ij}\) be the \(4\times 4\) matrices with all zero entries with the exception of a 1 in the \(ij^{th}\) entry. Making use of these matrices, we write
Then
where we defined the nonnegative scalar coefficients
This leads to
Finally, making use of
we arrive at
The first two terms, deriving from the adult density dependent fertility and the effect of cannibalism on egg survival, are negative and therefore contribute to a forward bifurcation. The third and fourth terms are positive, deriving from the positive density effect of cannibalism on adult survival. Finally the sign of the last term, which involves the synchrony term \(g^{\prime }\left( 0\right) <0\), is ambiguous due to the difference \(c_{3}\left( t\right) -c_{4}\left( t\right) \) whose sign is dependent on model parameters.
Rights and permissions
About this article

Cite this article
Cushing, J.M., Henson, S.M. Periodic matrix models for seasonal dynamics of structured populations with application to a seabird population. J. Math. Biol. 77, 1689–1720 (2018). https://doi.org/10.1007/s00285-018-1211-4
Received:
Revised:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00285-018-1211-4
Keywords
- Structured population dynamics
- Discrete-time population dynamics
- Periodically-forced matrix equations
- Periodic orbits
- Stability
- Bifurcations
- Seabird population dynamics
- Cannibalism
- Reproductive synchrony
- Animal behavior
- Allee effect
- Tipping point