
Abstract
Variation in communicative complexity has been conceptually and empirically attributed to social complexity, with animals living in more complex social environments exhibiting more signals and/or more complex signals than animals living in simpler social environments. As compelling as studies highlighting a link between social and communicative variables are, this hypothesis remains challenged by operational problems, contrasting results, and several weaknesses of the associated tests. Specifically, how to best operationalize social and communicative complexity remains debated; alternative hypotheses, such as the role of a species’ ecology, morphology, or phylogenetic history, have been neglected; and the actual ways in which variation in signaling is directly affected by social factors remain largely unexplored. In this review, we address these three issues and propose an extension of the “social complexity hypothesis for communicative complexity” that resolves and acknowledges the above factors. We specifically argue for integrating the inherently multimodal nature of communication into a more comprehensive framework and for acknowledging the social context of derived signals and the potential of audience effects. By doing so, we believe it will be possible to generate more accurate predictions about which specific social parameters may be responsible for selection on new or more complex signals, as well as to uncover potential adaptive functions that are not necessarily apparent from studying communication in only one modality.
Significance statement
Animals exhibit an astonishing diversity of communicative systems, with important variation in both the nature and the number of signals they produce. The roles of phylogenetic history, genetic drift, environmental factors, and sexual selection in shaping interspecific variation in communicative systems have long been acknowledged, whereas social complexity has only more recently emerged as a potential fundamental factor determining signal diversification. While a number of comparative studies support the key prediction of this hypothesis, i.e., that individuals living in more complex social environments exhibit more signals and/or more complex signals, we discuss several gaps in the current state of the art concerning this hypothesis and point out what we believe are neglected perspectives. By emphasizing the importance of the multimodal nature of communicative systems and the social context in which signals are exchanged, we hope to stimulate the development of new tests and specific questions based on this expanded framework.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Life in stable groups unfailingly entails repeated interactions between different individuals, creating particular costs and benefits for individual group members and the group as a whole. The “social intelligence hypothesis” suggests that increased socio-cognitive skills are necessary to effectively navigate this social network in order to reap the benefits of sociality (Jolly 1966; Humphrey 1976; Whiten and Byrne 1988; Dunbar 1993). Social competence, which describes the ability to reliably assess the behavior of others and to respond flexibly and adaptively to it, for example by remembering prior interactions, anticipating others’ behaviors, and by coordinating one’s own behavior with that of other group mates, summarizes the key cognitive abilities that are favored in this context (Cheney and Seyfarth 2005; Dunbar 2009; Taborsky and Oliveira 2012; Sewall 2015). Increased social competence has been suggested to trigger the need for more complex communicative systems, being the key behavioral mechanism mediating these interactions (McComb and Semple 2005; Freeberg 2006; Freeberg et al. 2012a, b; Sewall 2015).
The traditional approach to studying communication is based on the sender-receiver paradigm, i.e., a dyad connected by the transmission of a signal in a particular channel (Shannon and Weaver 1951). More recently, however, it has been recognized that signals may connect several senders and receivers within a group (McGregor and Peake 2000; Cheney and Seyfarth 2005; McComb and Reby 2005; Fichtel and Manser 2010; Pollard and Blumstein 2012). In particular, multiple individuals may communicate simultaneously, and conspicuous signals may draw the attention of unwanted receivers (McGregor and Peake 2000; Peake 2005; Fichtel and Manser 2010). Hence, specific mechanisms are required to direct a signal of a given sender towards a specific receiver (McGregor and Peake 2000; Freeberg et al. 2012b).
Moreover, group-living individuals are usually involved in a wider range of interactions with diverse social partners, ultimately triggering needs to transmit a broader diversity of information and to express a wider range of emotional and motivational states (Morton 1977; Freeberg et al. 2012b; Ord and Garcia-Porta 2012). Accordingly, the “social complexity hypothesis for communicative complexity” (SCHCC) posits that animals living in groups with comparatively greater social complexity will exhibit greater complexity in their signaling systems (Freeberg 2006; Freeberg et al. 2012a, b). This hypothesis formalizes ideas that have been acknowledged ever since Darwin (1872), but that had never been explicitly tested (see Freeberg et al. 2012b for a historical perspective). Accordingly, the complexity of social groups ought to drive communicative complexity in both a proximate (context for signal development) and ultimate (social selection pressure) sense towards increased signal diversity among species, populations, and groups (Freeberg et al. 2012a; Krams et al. 2012). Empirical evidence from diverse taxa and for three different communicative modalities (auditory, visual, and olfactory) has supported this hypothesis (Table S1). However, there are also several studies not reporting the predicted positive relationships (Table S2) and the work of Freeberg (2006) seems still to be unique in adopting an experimental approach to test predictions of this hypothesis. Building on influential previous work by Freeberg et al. (2012a, b), in the present paper, we point out new perspectives that significantly expand the existing framework.
Specifically, three key issues have emerged from the recent literature that may offer interesting new perspectives on the SCHCC (Fig. 1). The first issue concerns the operational definition of the main variables (1 and 2 in Fig. 1). Because the unspecified usage of the term complexity can lead to a lack of agreement on the relevant variables, there is a need for better quantitative tools to estimate social and communicative complexity more systematically across taxa (Freeberg et al. 2012b; Bergman and Beehner 2015; Fischer et al. 2016, 2017; Pika 2017; Kappeler 2019, topical collection on Social complexity). Secondly, while numerous studies have highlighted a link between social and communicative complexity, their correlative nature does not permit conclusions about the direction of causality (3 in Fig. 1). Hence, a systematic control for alternative hypotheses that invoke important effects of ecology, morphology, or phylogenetic history on communicative complexity appears essential when testing the link between social and communicative complexity. Finally, we note that researchers rarely address the actual ways in which variation in signaling is directly affected by social factors (4 in Fig. 1). Indeed, the underlying mechanisms of this link are usually not explored, failing to uncover the specific attribute of communication that would be co-evolving with specific aspects of sociality (Gustison et al. 2012, 2019, topical collection on Social complexity; Pollard and Blumstein 2012). In this review, we will address these three crucial issues, taking stock of the current state of the art and point out what we believe are the main associated perspectives. In doing so, we hope to provide guidelines and inspiration for future tests of the SCHCC.

Schematic representation of three key issues associated with tests of the “social complexity hypothesis for communicative complexity” (SCHCC)
What are we looking at? Need for good operational definitions
Whereas the term “complexity” is omnipresent in scientific publications, there is no general agreement on its definition, neither within nor among disciplines (Adami 2002; Bennett 2003). The two Latin roots com (together) and plectere (woven, plaited, twined, entwined) (Harper 2001) indicate that multiple parts are connected to each other in flexible ways. Pollard and Blumstein (2012) accordingly suggested that “a system is typically considered more complex if it contains more parts, more variability or types of parts, more connections or types of connections between parts or more layers of embedded meaning”. This verbal definition, though helpful, does not offer any operational criteria to evaluate social and communicative complexity, but highlights the importance of a holistic approach (Freeberg et al. 2012b). Hence, there is currently a need for better tools and concepts to estimate both social and communicative complexity more reliably across taxa (Freeberg et al. 2012b; Bergman and Beehner 2015; Fischer et al. 2016, 2017; Pika 2017; Kappeler 2019, topical collection on Social complexity).
Another issue when trying to define and quantify social and communicative complexity in the context of the SCHCC concerns the non-independence between communication and sociality that may even amount to circularity. If social complexity is measured through communicative interactions or communicative complexity through its social outcomes, then we should not be surprised to find correlations between these variables. An analogous problem exists between sociality and cognition, leading Bergman and Beehner (2015) to suggest to measure social cognition in a different context than the chosen measure of social complexity. In addition, communication, together with social interactions, constitutes the core of a species’ social structure (Kappeler 2019, topical collection on Social complexity). Hence, measures thereof that are correlated with measures of social organization, such as group size, may therefore reduce the risk of redundancy and non-independence, and this is what several studies effectively did (e.g., Blumstein and Armitage 1997; McComb and Semple 2005). However, this approach can and has been criticized on the grounds that measures of social organization (notably group size) alone do not fully capture all relevant dimensions of social complexity (Dunbar and Shultz 2010; Kappeler 2019, topical collection on Social complexity). Thus, comprehensive measures of social complexity, in particular, need to be developed (Kappeler et al. 2019, topical collection on Social complexity).
Social complexity: developing broadly applicable quantitative measures
The main variable used to assess social complexity in the context of the SCHCC has been group size (Table S1 and S2). Some studies have used various derived or more specific measures for sub-units (e.g., foraging group, grooming clique, or group density), but all of them are based on the notion that individuals in larger units are expected to interact with more partners, triggering a need to convey more diverse and flexible messages (Freeberg et al. 2012b). Group size is a convenient proxy of social complexity as it is relatively easy to assess for large numbers of species, but it is commonly recognized as only a crude measure of social complexity (McComb and Semple 2005; Shultz and Dunbar 2007; Freeberg et al. 2012b; Bergman and Beehner 2015). Indeed, even if group size is undeniably a key determinant of social complexity (Kappeler 2019, topical collection on Social complexity) and has been shown to be a good proxy for social complexity in some taxa (Bourke 1999; Jeanson et al. 2007), it may fail to capture differences in social complexity, especially across taxa. For example, a herd of ungulates, a troop of baboons and a colony of ants may be constituted of the same number of individuals, but no one in the field would consider them as equally socially complex.
Because the notion of complexity is not only based on the number of units but also on the connections between these parts (Freeberg et al. 2012b; Pollard and Blumstein 2012), some studies considered other social features, such as mating system, dominance style, or breeding strategies, as better or alternative proxies of social complexity. However, some of these alternative classifications have themselves been questioned, and their ranking can be controversial and largely dependent on the taxa considered (Kappeler 2019, topical collection on Social complexity). For example, pair-living species may be considered as the simplest form of social complexity because they have the smallest group size, but this type of social system typically also includes long-term bonds that require familiarity and a subtle coordination of each other’s behavior. In some taxa, they have therefore been considered as a very complex form of sociality (ungulates: Shultz and Dunbar 2006; bats: Pitnick et al. 2006), whereas in others certain pair-living species have been characterized as the baseline of social complexity (primates: Dröscher and Kappeler 2013).
Similarly, regarding the patterns of group-level dominance relationships, there is no agreement on which pattern is more socially complex. Some authors argued that linear dominance hierarchies are socially more complex than egalitarian ones because they require particular cognitive abilities, such as transitive inference (MacLean et al. 2008). However, non-linear hierarchies could equally be considered as more complex, especially from the animal’s perspective, as they represent more uncertainty and necessitate more social competence (Taborsky and Oliveira 2012). Hence, it is important to clarify the level of analysis; whereas some studies address complexity at the group level, others focus on the individual level, resulting in divergent interpretations (Aureli and Schino 2019, topical collection on Social complexity).
The ranking of social features in terms of social complexity may also be prone to circularity because social complexity is usually evaluated with regard to the number of cognitive skills or the type and frequency of communicative interactions. Hence, the usage of particular social features may not solve the definitional problem and may even enhance subjectivity. Against this background, the verbal definition by Freeberg et al. (2012b), who defined “complex social systems are those in which individuals frequently interact in many different contexts with many different individuals, and often repeatedly interact with many of the same individuals over time,” represents important progress. More recently, Bergman and Beehner (2015) suggested to measure social complexity as the “number of differentiated relationships that individuals have.” These verbal definitions, although non-operational, are essential pre-requisites for the development of meaningful quantitative measures, at least of variation in aspects of social structure, such as the one proposed by Fischer et al. (2017) based on several common social indices and cluster analysis, or the one offered by Weiss et al. (2019, topical collection on Social complexity) applying binomial mixture models to association indices.
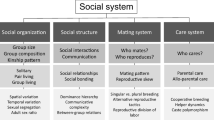
Facing this diversity of social complexity measures, it is evident that this concept encompasses different aspects and that a single measure may not be sufficient to assess it globally. Hence, a better appreciation of the complexity of a social system may rather be provided by a set of operational measures describing all of the four fundamental aspects proposed by Kappeler (2019, topical collection on Social complexity), i.e., social organization, social structure, mating system, and care system (Table 1). This systematic approach may facilitate progress with future comparative studies in this domain.
Hence, we believe that in the context of the SCHCC, more effort needs to be invested into the choice and definition of the variables used to assess social complexity. In particular, favoring broadly applicable quantitative measures rather than subjective categories should facilitate comparative work and clarify the level at which complexity is evaluated (Weiss et al. 2019, topical collection on Social complexity). Moreover, measures implicitly based on communicative variables should be avoided in order to stave off circularity. Rather than a single measure, we propose that a set of operational measures matching the criterion mentioned above and accounting for all four main components of social complexity (Kappeler 2019, topical collection on Social complexity) will help to move the field forward. Better identifying the social complexity aspects included in the test should also allow for better predictions and understanding of the mechanisms underlying the observed relationship (see below).
Communicative complexity: from partitioning signals towards a systematic multimodal approach
One of the main issues associated with communicative complexity results from the partitioning of the distinct modalities in which signals are produced. Each signaling modality, defined as the sensory system with which a signal is produced and perceived by the receiver (Smith and Evans 2013), has historically been highlighted in different taxa. This bias has led to the development of different research questions, approaches, and methodologies across these different modalities (Liebal et al. 2013). Our own sensory bias, together with differences in the ease of data collection and unequal development of technological tools, have resulted in large biases in the number of studies between the different modalities.
In the context of the SCHCC, but also more generally in communication research, mainly the auditory, olfactory, and visual modalities have been explored, with a strong bias towards acoustic communication (Baptista and Gaunt 1994; Slocombe et al. 2011). Its universality and the fact that most species produce acoustic signals easily perceptible by humans may explain this bias. Indeed, because many olfactory signals are not easily perceived by humans, more sophisticated analytical methods are required to study them in more detail. Only a few studies have addressed the complexity of these signals and even fewer from a comparative perspective (Kather and Martin 2015). Visual signals are highly diverse because they can be produced by completely different processes, such as movements of the whole body or body parts (e.g., foot-flagging of some frogs) as well as changes in coloration or shape (e.g., feather erection display of birds), making it difficult to define broadly applicable measures of visual signal complexity (Endler 1992; Cuthill et al. 2017). Moreover, visual signals vary widely in persistence, ranging from state signals, such as static features of coloration patterns, to dynamic signals with limited duration that require an action by the signaler to be initiated (Smith and Evans 2013).
However, despite this imbalance in previous studies of different modalities and the diversity of methods used to assess communicative complexity across these modalities, some commonalities exist (Table 1). According to the most common definitions, communicative complexity can be quantified via two main approaches: the number of distinct signals or signaling units or the number of bits of information included in signals (Griebel and Oller 2008; Freeberg et al. 2012b; Pollard and Blumstein 2012). In the following, we summarize how these two approaches have been applied across the three main modalities studied in the context of the SCHCC and identify their current limitations. At the end of this section, we argue in favor of enhanced acknowledgement of the multimodal aspect of communication within the framework of the SCHCC.
Number of distinct signals or signaling units
In all communicative modalities, the number of distinct signals or signaling units has been used as a proxy for communicative complexity. To assess vocal communicative complexity across species, vocal repertoire size, defined as the number of call type individuals of a population or species produce, has been mainly used. To this end, call types have been classified by visual inspections of spectrograms, a rather subjective approach. Although the more recently used quantitative approaches to estimate repertoire size are favored, subjectivity still persists in the way algorithms are implemented to classify call types (Fischer et al. 2016). This bias, together with the diversity of quantitative methods applied to identify call types, currently results in substantial inhomogeneity among studies. For example, the vocal repertoire of the Mongoose lemur (Eulemur mongoz) has been characterized by visual inspection as composed of either 8 (Petter and Charles-Dominique 1979), 10 (Curtis and Zaramody 1999), or 14 (Gamba et al. 2015) call types, whereas quantitative analyses suggested 9 (Gamba et al. 2015) or 15 (Nadhurou et al. 2015) different call types. Thus, there is currently no general agreement on how to identify different call types and, hence, how to assess vocal repertoire size objectively.
Similarly, the main variable used to access complexity in olfactory signaling is the number of distinct chemical compounds contained in the different excretions (guinea pigs: Beruter et al. 1973; primates: delBarco-Trillo et al. 2011, 2012; hymenoptera: Kather and Martin 2015). While some semiochemicals are composed of only one molecule (e.g., the female sex pheromone (Z)-7-dodecen-I-yl-acetate common to the Asian elephant (Elephas maximus) and several species of moths; Wyatt 2003), others are composed of a combination of several molecules. Moreover, the diversity of scent sources, as urine, feces, saliva, skin secretions, or specialized sent glands, has also been considered as a proxy to assess olfactory complexity (delBarco-Trillo et al. 2011; delBarco-Trillo and Drea 2014). In addition, olfactory communication is characterized by its temporal duality: while scent deposition is a one-off event, the scent itself usually persists for some time and can be perceived by receivers even after the departure of the sender. Because scent deposition is usually accompanied by a typical behavior, we propose that the number of scent-marking behaviors can also be used to characterize the complexity of an olfactory system, even though these behaviors often have a strong visual component. Although repertoires of scent-marking behaviors have been reported for some species, they have rarely been interpreted in the context of the SCHCC (Drea and Scordato 2008; Colquhoun 2011; delBarco-Trillo et al. 2011).
Regarding visual signals, the numbers of both static and visual signals have been used as a proxy for complexity, although only a few studies actually tested the SCHCC. For example, among 25 wasp species, flexibly nesting species (with either one or several foundresses) exhibited more intraspecific variation in body markings than species having a unique nesting strategy (Tibbetts 2004; Table S1). In addition, dynamic signals, such as the number of visually distinct facial movements correlate with group size in Old World primates (Dobson 2009; Table S1).
To summarize, the number of signals or signaling units is a widespread method of accounting for complexity across modalities (Table 1). However, this section also emphasizes the diversity of levels that may be evaluated through this approach. While some of these approaches may be applied at the system level across modalities (e.g., number of signaling behaviors), some others are more modality-specific and are usually applicable at the signal level (e.g., number of compounds, number of notes).
Number of bits of information or amount of uncertainty
This approach is based on information theory and represents a measure of the entropy of a system, which is an estimate of the amount of uncertainty (Shannon and Weaver 1951; Fischer et al. 2016). Thus, Shannon’s entropy is not a measure of what one communicates, but rather a measure of what one could communicate (Shannon and Weaver 1951; Robinson 2008). This is an essential conceptual difference that may help to avoid circularity. Indeed, considering the social information contained in signals, rather than the possibilities offered by the structure of these signals to convey flexible and diverse information as a measure of communicative complexity, may lead to non-informative tests of the SCHCC. From a fixed set of signals, more information may first emerge by combinations, either through assemblage (simultaneous expression of different signals) or sequence (succession of different signals). Hence, acoustic units, i.e., uninterrupted traces in a spectrographic representation, and discrete calls can be combined into sequences (Bouchet et al. 2013; Manser et al. 2014).
Vocal sequences have been an intense area of research in studies of birdsong (Kroodsma 2004; Catchpole and Slater 2008b), but the existence of non-random vocal structures has also been shown in several mammalian orders, including cetaceans (Riesch et al. 2008; Shapiro et al. 2011; Cholewiak et al. 2013; Zwamborn and Whitehead 2017), primates (Clarke et al. 2006; Arnold and Zuberbühler 2008; Zuberbühler and Lemasson 2014; Gustison et al. 2016), bats (Kanwal et al. 1994; Bohn et al. 2009), and hyraxes (Kershenbaum et al. 2012). The combination of signals is an alternative way to increase the amount of information transmitted, and several methods have been proposed to analyze the structure and rhythmicity of these sequences (reviewed in Kershenbaum et al. 2016; Ravignani and Norton 2017). Indeed, the number of call/note combinations correlates with group size in several species (Freeberg 2006; Bouchet et al. 2013; Manser et al. 2014), suggesting that this is a promising area for future research on communicative complexity.
Similarly to vocalizations, olfactory signals can also be produced in sequences and the combinational deposition of scents appears to be widespread in mammals (Mills et al. 1980; Clapham et al. 2014; Vogt et al. 2014). For instance, male ring-tailed lemurs (Lemur catta) flexibly use two specific scent glands, by either depositing only one type of scent or a mixture of the two secretions. The mixture elicited a higher responsiveness, suggesting that they contain a larger number of messages and persist for longer (Greene et al. 2016). The superposition or juxtaposition of different scent marks may also be studied following the same analytical methods as the ones mentioned above for acoustic sequences (Kappeler 1998). Similarly, numerous courtship displays have been described as assemblages or sequences of different visual signals (Lorenz 1941), but these combinations have rarely been related to sociality in a comparative perspective (Crook 1964).
Variability in the structure of signals presents another potential way to increase the amount of transmitted information. For example, vocal systems have been suggested to be either discrete, with acoustically distinct call types that are easily discriminated from each other, or graded, with the acoustic structure of the vocalizations forming a continuum without clear boundaries between different call types (Marler 1977; Keenan et al. 2013). The gradation level is a measure of the overlap in temporal and spectral features existing between two calls, and might be one of the main reasons for the inconsistency in measures of vocal repertoire sizes. The higher the level of gradation in a system, the more difficult it is to cluster its components, but the higher is the potential for information transmission. Gradation may therefore represent a measure of bits of information that may be contained in signals. Because vocal repertoires usually contain a combination of discrete and graded calls (Fichtel et al. 2001; Bouchet et al. 2013; Manser et al. 2014; Fischer et al. 2016), the level of gradation can also be used to operationalize the complexity of a vocal system (Fischer et al. 2016).
Different selective pressures seem to shape signals as a function of the quantity of information required from a signal. While stereotypical signals with high context specificity reduce ambiguity, more flexible signals can convey more subtle and complex information about the sender’s motivational state or a particular context (Fischer 1998; Fichtel et al. 2001; Griebel and Oller 2008; Manser et al. 2014). Accordingly, the “call social function hypothesis” proposes that the level of overlap in temporal and spectral features of a call type will be influenced by its social function (Snowdon et al. 1997; Griebel and Oller 2008; Lemasson and Hausberger 2011; Bouchet et al. 2013; Keenan et al. 2013). In female Campbell’s monkeys (Cercopithecus campbelli), calls associated with the highest affiliative social value exhibited the greatest gradation level, whereas calls associated with agonism were the most stereotyped, and less social alarm calls were of intermediate variability (Lemasson and Hausberger 2011; Table S1).
Furthermore, inter-individual variation in call structure is another source of gradation that might be impacted by group size. The more group members interact, the more they may benefit from accurately recognizing the sender of a call, but this recognition task becomes increasingly difficult with increasing group size (Freeberg 2006; Pollard and Blumstein 2011). Hence, if individual recognition is important, more individualistic signatures should evolve with increasing group size as shown, for example, in chick begging calls of swallows (cliff swallows Hirundo pyrrhonota and barn swallows Hirundo rustica; Medvin et al. 1993), alarm calls of ground-dwelling sciurids (Pollard and Blumstein 2011), and infant isolation calls of bats (Wilkinson 2003; Table S1).
Hence, the gradation level of the vocal system (1 in Fig. 2), the gradation level of different call categories (2 in Fig. 2), and the stereotypy of different call types (3 in Fig. 2), as well as the level of individuality in different call types (4 in Fig. 2), are all sources of gradation that may be impacted by social variables. However, a practical quantitative measure is still lacking to evaluate gradation at the system level. The distribution of call stereotypy coefficients has recently been suggested as a potential measure of gradation for comparative analyses, with right-skewed distributions (closer to 1) indicating higher differentiation and left-skewed distributions (closer to 0) indicating a higher degree of gradation (Wadewitz et al. 2015; Fischer et al. 2016). By applying a fuzzy-c-means cluster algorithm, which allows for imperfect membership, calls can be assigned to different clusters. For each call a membership value is attributed for each cluster, ranging from 1 (the call fully displays the properties of the cluster in question) to 0 (the call does not display any of the properties of the cluster). The call stereotypy coefficient (referred to as “typicality coefficient” in Wadewitz et al. 2015 and Fischer et al. 2016) can then be obtained by subtracting the two highest membership values associated with this call. Hence, this measure allows for quantifying the gradation level between two call types but does not provide a generic measure of the gradation level of the vocal repertoire. Therefore we suggest that the distribution of the standard deviations across membership values for each call might be a more appropriate measure of gradation at the system level.

Schematic representation of (i) the different levels of gradation: gradation level of the repertoire (for a given species), gradation level of the call category (for a given species and call category, e.g., affiliative calls), call stereotypy (for a given species and call type), and individuality (for a given call and a given individual) and (ii) the associated potential comparative levels (1 to 4). Each gray capital letter represents a different call type, e.g., A is a call type of category 1 produced by the individuals of species I
Although this approach to assess the gradation level between signals has almost exclusively been adopted in the acoustic modality, the level of overlap in signal features may theoretically also be applied to other modalities. Hence, the relative proportions of different chemical components may also be considered as a form of gradation, with possible variation among species, groups, and individuals (Symonds and Elgar 2008; delBarco-Trillo et al. 2011). This gradation approach has also been suggested to be applicable to dynamic visual signals, such as facial expressions and gestures (Fischer et al. 2016), but this remains to be explored in practice.
A final approach to assess communicative complexity is to evaluate the perceptive adaptations of a system. This approach is highly complementary with the precedent one as in all species, sensory capabilities co-evolved with the range of signals they produce. Moreover, it should provide information on the ability of the receiver to discriminate between different stimuli. Hence, from an evolutionary point of view, this approach is significant, but has only rarely been addressed (Ramsier et al. 2012; Elgar 2015). For example, foraging group size co-varied with the overall hearing sensitivity and high-frequency limits across 11 species of strepsirrhine primates, suggesting that social complexity favored enhanced hearing sensitivities, especially at higher frequencies (Ramsier et al. 2012; Table S1). Similarly, in the olfactory domain, social Hymenoptera species have higher densities of hair-like sensillae, with which they perceive olfactory signals, compared to species that became solitary (Wittwer et al. 2017; Table S1). Comparative studies on visual adaptations may also be informative (Endler et al. 2005), but studies of variation in visual communication have so far mostly considered effects of ecological factors, such as background vegetation (Fleishman 1992), habitat type (Malacarne et al. 1991), or diurnality and predatory habits (Veilleux and Kirk 2014).
To summarize, evaluating the amount of uncertainty in a signal or signaling system represents a common approach to evaluate communicative complexity across modalities (Table 1). Although the most common approach is to study of combinational use of signals, the concept of gradation, currently mainly used for the vocal modality, is a really promising one that may allow for a systematic evaluation of communicative of complexity at both the signal and system level. This approach may benefit by also considering the complexity of the associated perceptive adaptations.
Thus, across all three modalities discussed here, several complementary approaches of complexity appear particularly relevant. These approaches are based on the number of signals or signaling units and the uncertainty (combinations and gradation of signals). These approaches are applicable across modalities and address both signal and system levels and may therefore also be used for broad comparative studies. As for measures of social complexity, we encourage the development of complementary quantitative operational measures of communicative complexity that incorporate the insights addressed above.
Integrating the multimodal nature of communication
While the field of animal communication has moved in the early 2000s from a unimodal approach to a multimodal one, acknowledging the fact that most animals produce signals across different modalities (McGurk and Macdonald 1976; Partan and Marler 1999; Rowe 1999; Candolin 2003; Hebets and Papaj 2005), much research in the context of the SCHCC has remained focused on a single modality. However, there are several good reasons for acknowledging the multimodal nature of signals and communicative systems in this framework. Two fundamental aspects of communication effectively deserve some attention, first, at the signal level, the fact that numerous signals are by themselves multimodal and, second, at the system level, the fact that individuals, even when not using multimodal signals, usually communicate through different modalities (Liebal et al. 2013; Pika 2017).
Multimodal signals are by definition complex, because they involve more than one signaling and perceptive system (Rowe 1999; Smith and Evans 2013). Moreover, multimodal signals have been suggested to be widespread in group-living animals (Partan and Marler 1999), but this assumption has never been formally tested. Isolating one component of a multimodal signal may lead to false conclusions because some signals have a different function when they are expressed independently or together with another signal component (“multiple signals/messages hypothesis” and “disambiguation hypothesis”; Hebets and Papaj 2005; Liebal et al. 2013). For example, female red-winged blackbirds (Agelaius phoeniceus) use the same visual display for courtship and aggression, and only the addition of a vocalization allows discrimination between contexts (Beletsky 1983).
At the system level, focusing on one modality only may lead to over- or underestimation of the relationship between social and communicative complexity, respectively. For example, submission in primates can be expressed either by visual or acoustic signals (Jolly 1966; de Waal and Luttrell 1985), showing that the same social selective pressure may result in different signaling adaptations that are equally complex but expressed through different modalities. Second, the flexible use of different modalities in multimodal signals permits another level of complexity embedded in multimodal signaling (Liebal et al. 2013; Wilke et al. 2017). For example, captive chimpanzees (Pan troglodytes) used more tactile and vocal signals when the experimenter was facing away, but more visual-gestural signals when the experimenter was facing the chimpanzee (Leavens et al. 2010). Thus, depending on the species in question, it may also be highly relevant to include some modalities (e.g., vibratory, thermal, electrical) that are not addressed here because they have not yet been considered in the framework of the SCHCC.
Hence, we suggest that establishing a cross-modal signal repertoire would not only fill the gap in our current understanding of the multimodal nature of most signals (Partan and Marler 2005) and contribute to a more comprehensive assessment of communicative complexity, but would also allow more meaningful tests of the SCHCC (Liebal et al. 2013; Waller et al. 2013). Moreover, applying a multimodal approach may also help to uncover the different selective pressures acting on the communicative system and to better understand adaptive functions that might not be clear from the study of its components independently (Liebal et al. 2013).
Integration of expanded operational definitions
We argue that integrating appropriate quantitative measures of social complexity (as described above) together with cross-modal measures of communicative complexity (number of signaling units and uncertainty) in a holistic fashion should facilitate comparative research on the SCHCC and may reveal important avenues for future research. Although the development of appropriate analytical tools to study multimodal communication has been debated for a while (Partan 2013), a recent network approach has offered great promise. This method has been used to uncover the contributions of female mate choice and male-male competition in the North American barn swallows (Hirundo rusticaerythrogaster) via examining a comprehensive array of phenotypic variables by a correlation-based phenotype network (Wilkins et al. 2015; Hebets et al. 2016). A similar approach could be used for a systematic analysis of communication systems and their links with social parameters. For example, each node in a signal phenotype network may represent a different signal/signaling unit or a different communicative complexity measure, with different node shapes representing different modalities or types of signals. The edge between two signals may then represent the correlation between these two signals across individuals in terms of either frequency of occurrence (Example 1 in Fig. 3) or value (Example 2 in Fig. 3) or their temporal association (Example 3 in Fig. 3), depending on the chosen measures. The comparison of these communicative system representations across different social context (e.g., activities or audiences) may, for instance, reveal new interesting relationships between communicative signals and social parameters. Integrating relevant social parameters into these networks would also allow exploring the specific links between different social and communication variables at the system level (Wilkins et al. 2015). The application of such a system-inspired framework, for example, revealed dynamic changes in signal structure and function across environmental and social contexts in courtship displays of wolf spiders (Schizocosa floridana) (Rosenthal et al. 2018).

Three heuristic examples of possible network construction presented on a conceptual diagram showing different levels of system degeneracy and modularity. The meaning of the edge and nodes for each example are explained in the bottom-right quarter. This figure is adapted from Wilkins et al. 2015
Hence, we believe that this method would not only offer instructive visual representations of multimodal communicative systems, but also an opportunity to extract new quantitative and broadly applicable measures that could be interpreted in terms of communicative complexity. This method indeed offers the possibility to characterize a given system in terms of degeneracy, modularity and pluripotentiality (Wilkins et al. 2015; Hebets et al. 2016). Degeneracy is a measure of network density and represents the number of significant correlative relationships; a highly degenerated system will be composed of signals potentially carrying the same information or having the same function (see “redundancy” in Wilkins et al. 2015; Examples 1 and 2 in Fig. 3). Modularity represents the degree to which relationships exist across versus within clusters, here representing signal types or modalities; a highly modular system will be composed of more modality-specialized individuals (Wilkins et al. 2015; Example 2 in Fig. 3). Pluripotentiality is a measure of the degree to which identical display components function across contexts; a highly pluripotent system will be composed of more functionally flexible signals that are used across contexts (Hebets et al. 2016).
Hence, in the context of the SCHCC, we make a plea for more comprehensive study of communicative systems, integrating a set of quantitative and complementary measures of social and communicative complexity and accounting for the multimodal aspect of communication. We believe that his approach would bring considerable insights in the current debate, particularly allowing for the emergence of more specific predictions on the nature of the link between social and communicative factors.
The links between social and communicative complexity
Although several studies have supported the social complexity hypothesis by revealing correlative relationships between social and communicative variables, only a few studies have examined and discussed the nature of these links in detail. Hence, the direction of causality and the mechanisms underlying these links remain currently unclear (Freeberg et al. 2012b; Ord and Garcia-Porta 2012; Gustison et al. 2016, 2019, topical collection on Social complexity), and these questions would benefit from additional research.
Control for alternative hypotheses: directionality and strength of the links
When studying the SCHCC, it is essential to also consider potential alternative hypotheses driving the evolution of signals. This is fundamental in both situations when a correlational link is found or when no such link is found, even though a link would be predicted (Freeberg et al. 2012b). For both outcomes, several alternative explanations are possible (Fig. 4), revealing two fundamental problems. First, the relative role of social complexity in shaping communicative complexity needs to be assessed. Social complexity may shape the complexity of a particular set of signals synergistically with other selective pressures also selecting for more complexity (Fig. 4, scenarios A and B) or despite other additional selective pressures (selective forces act in opposite directions; Fig. 4, scenarios C and D), making it hard to extract the actual effect of social complexity itself (Fig. 4, scenarios B and D).

Schematic representation of different potential evolutionary scenarios depicting the link between social and communicative complexity. A: scenario in which a link is observed, social complexity is the main selective pressure driving communicative complexity, in parallel other factors also contribute selecting for communicative complexity; B: scenario in which no link is observed with social complexity because other selective pressures have a stronger effect on the evolution of communicative complexity; C: scenario in which a link is observed, social complexity is the main selective pressure driving communicative complexity even if other factors constrain the evolution of communicative complexity; D: scenario in which no link is observed with social complexity because the effect is counterbalanced by other selective pressures constraining the evolution of communicative complexity; E: scenario in which a link is observed but communicative complexity is under other selective pressures and drive itself the evolution of more complex social systems
Second, correlative studies do not permit conclusions about the direction of causality. Complex communicative systems may evolve in response to pressures related to social complexity, but complex communicative systems might also be driven by other selective pressures independent of changes in sociality that could in turn facilitate the evolution of greater social complexity (Fig. 4, scenario E; McComb and Reby 2005; Freeberg et al. 2012b; Ord and Garcia-Porta 2012). For example, a comparison across 22 species of lizards revealed that the number of head-bobbings was better explained by habitat use, i.e., the level of arboreality, than by the intensity of competition among males (Ord and Garcia-Porta 2012). Similarly, a comparison across 23 species of birds revealed that syllable repertoire size was better explained by body size than by the level of extra-pair paternity, and a comparison across 32 species of frogs revealed that the level of call amplitude modulation was better explained by phylogeny than by the intensity of competition among males (Ord and Garcia-Porta 2012).
Hence, non-social selective pressures such as ecology, anatomy or a phylogenetic null model explained changes in signaling complexity better than the specific aspects of social complexity considered in these analyses (Ord and Garcia-Porta 2012; Fig. 4, scenario B). The mechanisms underlying these selective pressures have been explicated by Freeberg et al. (2012b) and are summarized in Table 2. In general, the evolution of complex signaling systems is presumably not the result of a single specific selective pressure but rather the result of a combination of several ones (Freeberg et al. 2012b; Ord and Garcia-Porta 2012; Ramsier et al. 2012; Manser et al. 2014). Thus, if two species have similar social systems but only one experiences a specific ecological challenge, different types of signals may evolve, but these signals will not necessarily differ in complexity. And, as argued above, the same selective pressure may lead to different adaptations that may be equivalent in terms of complexity but involve different modalities.
To deal with these complications, appropriate statistical tools controlling for alternative hypotheses are required. For example, phylogenetic comparative studies can reveal the order in which different traits evolved in a given lineage (e.g., Podos 2001; Wittwer et al. 2017). Model fitting procedures may also allow for assessing the relative role of the different factors in selecting for communicative complexity and test the SCHCC against alternative hypotheses (Ord and Garcia-Porta 2012). Control of the alternative hypotheses may also be achieved by a quasi-experimental approach that includes the study of closely related species sharing similar habitats, climatic conditions, food resources, parasites, and predators, but exhibiting differences in their social systems (Krams et al. 2012; Ord and Garcia-Porta 2012).
Identification of the underlying mechanisms: exploring the relationship(s) between signal variation and social factors
How variation in signaling is directly affected by social factors is usually not explored, failing to uncover both the specific attributes of communication that would evolve in response to specific aspects of sociality and the actual way these social factors may influence signaling behavior (Pollard and Blumstein 2012; Gustison et al. 2012, 2019, topical collection on Social complexity). In the next sections, we offer suggestions on how to explore the nature of these relationship with both, ultimate (e.g., function of derived calls or complex signals) and proximate approaches (e.g., audience effect and social learning).
Studying the social function of derived signals
One approach to begin closing this gap is to study the social function of derived signals to obtain a better understanding of the functional meaning of the observed differences (e.g., why a larger vocal repertoire evolved in the more socially complex species). Identifying the relevant differences (e.g., which are the derived calls?), their social functions (e.g., what are the social functions of these derived calls?) and assessing potential fitness consequences of these derived signals in detail will help to identify the potential social selective factors driving the evolution of these signals (Gustison et al. 2012; Liebal et al. 2013).
Regarding the vocal modality, it has been shown that rather than an overall increase in repertoire size, an increased number of different calls are generally found only in one or two call categories, with species exhibiting greater social complexity having vocal repertoires including a greater proportion of vocalizations used in affiliative contexts (Le Roux et al. 2009; Taylor and Reby 2010; Fedurek and Slocombe 2011; Briefer 2012; Gustison et al. 2012). A descriptive identification of derived call categories has been conducted in a relatively small set of closely related species (Kroodsma 1977; Stirling and Thomas 2003; Manser et al. 2014), but recently also in a comparative study across 253 bird species (Leighton 2017; Table S1). Birds breeding cooperatively had a significantly larger vocal repertoire size than other species. This increase in repertoire size was due to specific increases in the number of alarm and contact calls but not aggression, flight, territorial, or mating calls. Although this contextual information is undeniably valuable for discussing the SCHCC, these broad categories nevertheless fail to uncover the specific social function at stake.
In contrast, a detailed comparison of the vocal repertoire of two closely related primate species (chacma baboons, Papio ursinus, and geladas, Theropithecus gelada) with different social systems, identified eight homologous calls, but also six derived calls present only in the geladas, suggesting that the socially more complex geladas have larger vocal repertoires (Gustison et al. 2012; Table S1). Gelada males use these calls to maintain social relationships with females in their units and specifically direct these calls towards females after conflicts. Interestingly, this difference parallels a social divergence between the two species because gelada males form long-term bonds with several females in a harem-like reproductive unit, whereas chacma males only form temporary associations with females. Hence, comparing the nature of the difference between signal repertoire sizes between species that differ in aspects of their sociality can help to uncover the role of the respective social factors in explaining the observed variation between the communicative systems.
Audience effects and signal complexity
The audience effect is defined as a change in signaling behavior caused by the presence of the audience; the audience being defined as any individual that is within the signal range (Marler et al. 1986; McGregor and Dabelsteen 1996). Therefore, signaling behaviors may be influenced not only by the characteristics of a targeted receiver (Fröhlich et al. 2016) or its attentional state (Leavens et al. 2010; Smith et al. 2011) but also by the nature of the audience (Slocombe and Zuberbühler 2007; Kalan and Boesch 2015; Crockford et al. 2017) including unwanted receivers (Matos and McGregor 2002; Smith et al. 2011). Hence, the SCHCC could provide a theoretical framework for testing audience effects by considering variation in the audience as an aspect of social complexity. Studying audience effects on complex signals may therefore help to uncover the particular social selective pressures that may have led to the evolution of flexible signals. Multimodal or multicomponent signals appear to be particularly good candidates to examine this audience effect.
First, signals including functionally redundant components may serve to increase signal detection and memorization (“backup hypothesis”: Johnstone 1997; “receiver psychology hypothesis”: Rowe 1999). This type of signal may optimize signal transmission but may also attract the attention of unwanted receivers. Hence, to balance benefits and costs, multicomponent signals should be associated with some behavioral flexibility, allowing variation in usage and signal structure (e.g., only one component is expressed) between contexts (e.g., more discrete signal when unwanted receivers are in proximity). Male jungle fowl (Gallus gallus), for example, exhibit variability in their tidbitting food-call recruitment behavior used not only to attract females but also to trigger aggression of male competitors (Smith et al. 2011). Tidbitting is a visual signal consisting of repetitive movements of the head and neck and can be combined with repetitive pulsatile sounds. Subordinate males adapt this display to the attentional state of the alpha male of the group by producing more frequently bimodal tidbitting when the alpha male is distracted (Smith et al. 2011).
Second, signals including non-functionally redundant components may carry additional information (“multiple messages hypothesis”: Møller and Pomiankowski 1993) that might be addressed at different audiences and may also be flexibly adapted between contexts. Even if it remains to be tested, this may be particularly true for mammal scent-marking behaviors (“demonstrative marking hypothesis”; Estes 1967; Johnston 2005) because of its temporal duality. Scent deposition behaviors often include obvious visual signals that may reach individuals present in the vicinity of the sender, while the scent itself may be directed towards both the present and future audiences.
Hence, studying the flexibility of complex signal usage (e.g., occurrence or structural modifications) across social contexts (audiences) will permit to identify the different individual social characteristics that may elicit or constrain complex signal expression. These social characteristics may later constitute social pressures acting for or against the evolution of these complex signaling behaviors.
Ontogeny of flexible signaling behavior: social contexts associated with vocal production learning flexibility in vertebrates
Despite fundamental similarities in the structure and mechanisms of vocal production systems across vertebrates, important differences exist between species at the level of flexibility in vocal production. Accordingly, a distinction is usually made between vocal learners and non-vocal learners. Non-vocal learner species produce adult-like vocalizations from birth onward and usually possess a relatively small vocal repertoire, which is genetically controlled and evolutionarily conserved. In contrast, vocal learner species learn their vocalizations from another conspecific (social tutor). Early vocalizations are usually structurally simple and highly variable, becoming more complex and stereotyped with age. In some species, vocal learning occurs only during a specific early sensitive period, whereas others exhibit open-ended vocal learning (Egnor and Hauser 2004; Catchpole and Slater 2008a).
In contrast to vocal contextual learning (in which “a pre-existing signal comes to be associated with a new context”, Janik and Slater 2000), vocal production learning refers more specifically to the modification in the structure of vocal signals through experience with another conspecific (Janik and Slater 2000; Ruch et al. 2018). These changes in vocalizations can be classified as either vocal accommodation, when existing vocalizations are modified, or lexical learning, when a new vocalization is acquired (Ruch et al. 2018). The latter form of vocal learning is particularly interesting in the context of the SCHCC as it may lead to an increasing number of signals used by an individual across its lifespan. So far, evidence for lexical learning was only found in three distantly related groups of birds (parrots, hummingbirds, and songbirds) and four distantly related groups of mammals, i.e., humans (but not in other primates), bats, cetaceans, and elephants (Nowicki and Searcy 2014; Ruch et al. 2018).
In most of these species, individuals exhibit babbling-like behavior in early life (bats: Knörnschild et al. 2006; cetaceans: Vergara and Barrett-Lennard 2008; parrots: Masin et al. 2004). Babbling not only is primarily considered as an essential practice for vocal learning but also seems to play a key role in social exchanges with caregivers (Vergara and Barrett-Lennard 2008; Tallerman and Gibson 2012). In humans, babbling triggers positive responses from kin as well as non-kin caretakers (Tallerman and Gibson 2012). Hence, babbling should be particularly adaptive in species displaying allomaternal care, as for example in cooperative breeders. In this social environment, young have to compete with each other for the attention of non-kin helpers. Interestingly, allomaternal care is present in all the families mentioned above exhibiting lexical learning (cetaceans: Hill and Campbell 2014; humans: Tallerman and Gibson 2012; elephants: Rapaport and Haight 1987; birds: Hatchwell 2009). For example, humans and callitrichids exhibit the greatest degree of allomaternal care among primates, and they are the only primate taxa with babbling infants (Tecot et al. 2012; Burkart et al. 2017). This potential link between the presence of allomaternal care and lexical learning is also consistent with the relatively recent idea of a possible co-evolution between vocal communication complexity and cooperation (Tallerman and Gibson 2012; Freeberg and Krams 2015). Hence, more elaborate vocal behavior and babbling-like behavior might be more common in species with allomaternal care. The examples illustrate the point that studying the social environments in which flexible vocal learning takes place may also illuminate the causal relationships between social variables and communicative complexity.
Conclusions
Many previous studies of communicative complexity have focused on specific links between single social and communicative variables, oversimplifying the complexity of these interrelations and ignoring the specific underlying mechanisms. We therefore make a plea for more specific predictions and a more comprehensive study of communicative systems. We particularly recommend:
- (1.)
To apply broadly applicable quantitative measures of social complexity in order to avoid the pitfalls of subjectivity and circularity and to clarify the level at which complexity is evaluated.
- (2.)
To assess communicative complexity at the system (repertoire) level, accounting for the multimodal nature of communication at both the signal and system levels.
- (3.)
To control for alternative hypotheses to the SCHCC through the application of appropriate statistical methods or careful selection of study species.
- (4.)
To develop more specific predictions about particular social factors that may impact a specific communication variable and the underlying mechanisms at play.
Moreover, we argue that integrating appropriate social and communicative complexity measures and studying the social contexts promoting complex signal expression in more detail may advance our current understanding of the links between social and communicative complexity. Integrating quantitative measures of social complexity with cross-modal measures of communicative complexity should help to uncover the selective pressures acting on the communication system and may additionally provide an opportunity to extract new quantitative and broadly applicable measures of communicative complexity at the system level. Studies of the social contexts associated with complex signal expression at both the ultimate and proximate level may identify the specific attributes of communication that evolve in response to specific aspects of sociality.
In summary, we propose that expanding tests of the SCHCC in scope (systematic approach across modalities) and depth (characterization of the observed relationships) will significantly advance our understanding of the intricate links between animal sociality and communication. Studying the SCHCC along these lines may also inform current debates in the study of social cognition, where communication is rarely discussed, as well as in studies of language origins and evolution.
References
Adami C (2002) What is complexity? BioEssays 24:1085–1094. https://doi.org/10.1002/bies.10192
Arnold K, Zuberbühler K (2008) Meaningful call combinations in a non-human primate. Curr Biol 18:R202–R203. https://doi.org/10.1016/j.cub.2008.01.040
Aureli F, Schino G (2019) Social complexity from within: how individuals experience the structure and organization of their groups. Behav Ecol Sociobiol. https://doi.org/10.1007/s00265-018-2604-5
Baptista LF, Gaunt SLL (1994) Advances in studies of avian sound communication. Condor 96:817–830. https://doi.org/10.2307/1369489
Beletsky LD (1983) Aggressive and pair-bond maintenance songs of female red-winged blackbirds (Agelaius pboeniceus). Z Tierpsychol 62:47–54. https://doi.org/10.1111/j.1439-0310.1983.tb02140.x
Bennett CH (2003) How to define complexity in physics, and why. In: Gregersen NH (ed) From complexity to life: on the emergence of life and meaning. Oxford University Press, UK, pp 34–43
Bergman TJ, Beehner JC (2015) Measuring social complexity. Anim Behav 103:203–209. https://doi.org/10.1016/j.anbehav.2015.02.018
Beruter J, Beauchamp GK, Muetterties EL (1973) Complexity of chemical communication in mammals: urinary components mediating sex discrimination by male guinea pigs. Biochem Biophys Res Commun 53:264–271. https://doi.org/10.1016/0006-291X(73)91429-0
Blumstein DT, Armitage KB (1997) Does sociality drive the evolution of communicative complexity? A comparative test with ground-dwelling sciurid alarm calls. Am Nat 150:179–200. https://doi.org/10.1086/286062
Bohn KM, Schmidt-French B, Schwartz C, Smotherman M, Pollak G (2009) Versatility and stereotypy of free-tailed bat songs. PLoS One 4:e6746. https://doi.org/10.1371/journal.pone.0006746
Bouchet H, Blois-Heulin C, Lemasson A (2013) Social complexity parallels vocal complexity: a comparison of three non-human primate species. Front Psychol 4:390. https://doi.org/10.3389/fpsyg.2013.00390
Bourke AFG (1999) Colony size, social complexity and reproductive conflict in social insects. J Evol Biol 12:245–257. https://doi.org/10.1046/j.1420-9101.1999.00028.x
Briefer EF (2012) Vocal expression of emotions in mammals: mechanisms of production and evidence. J Zool 288:1–20. https://doi.org/10.1111/j.1469-7998.2012.00920.x
Burkart JM, van Schaik C, Griesser M (2017) Looking for unity in diversity: human cooperative childcare in comparative perspective. Proc R Soc B 284:20171184. https://doi.org/10.1098/rspb.2017.1184
Candolin U (2003) The use of multiple cues in mate choice. Biol Rev 78:575–595. https://doi.org/10.1017/S1464793103006158
Cardoso GC, Hu Y (2011) Birdsong performance and the evolution of simple (rather than elaborate) sexual signals. Am Nat 178:679–686. https://doi.org/10.5061/dryad.1q424
Catchpole C, Slater PJB (2008a) How song develops. In: Catchpol CK, PJB S (eds) Bird song: biological themes and variations. Cambridge University Press, Cambridge, pp 49–81
Catchpole C, Slater PJB (2008b) Themes and variations. In: Catchpol CK, PJB S (eds) Bird song: biological themes and variations. Cambridge University Press, Cambridge, pp 203–234
Cheney DL, Seyfarth RM (2005) Social complexity and the information acquired during eavesdropping by primates and other animals. In: McGregor PK (ed) Animal communication networks. Cambridge University Press, Cambridge, pp 583–603
Cholewiak DM, Sousa-Lima RS, Cerchio S (2013) Humpback whale song hierarchical structure: historical context and discussion of current classification issues. Mar Mammal Sci 29:E312–E332. https://doi.org/10.1111/mms.12005
Clapham M, Nevin OT, Ramsey AD, Rosell F (2014) Scent-marking investment and motor patterns are affected by the age and sex of wild brown bears. Anim Behav 94:107–116. https://doi.org/10.1016/j.anbehav.2014.05.017
Clarke E, Reichard UH, Zuberbühler K (2006) The syntax and meaning of wild gibbon songs. PLoS One 1:e73. https://doi.org/10.1371/journal.pone.0000073
Colquhoun IC (2011) A review and interspecific comparison of nocturnal and cathemeral strepsirhine primate olfactory behavioural ecology. Int J Zool 2011:362976. https://doi.org/10.1155/2011/362976
Crockford C, Wittig RM, Zuberbühler K (2017) Vocalizing in chimpanzees is influenced by social-cognitive processes. Sci Adv 3:e1701742. https://doi.org/10.1126/sciadv.1701742
Crook JH (1964) The evolution of social organisation and visual communication in the weaver birds (Ploceinae). Behaviour 10:1–178
Curtis D, Zaramody A (1999) Social structure and seasonal variation in the behaviour of Eulemur mongoz. Folia Primatol 70:79–96. https://doi.org/10.1159/000021679
Cuthill IC, Allen WL, Arbuckle K, Caspers B, Chaplin G, Hauber ME, Hill GE, Jablonski NG, Jiggins CD, Kelber A, Mappes J, Marshall J, Merrill R, Osorio D, Prum R, Roberts NW, Roulin A, Rowland HM, Sherratt TN, Skelhorn J, Speed MP, Stevens M, Stoddard MC, Stuart-Fox D, Talas L, Tibbetts E, Caro T (2017) The biology of color. Science 357:eaan0221. https://doi.org/10.1126/science.aan0221
Darwin C (1872) The expression of the emotions in man and animals, vol 1998. Oxford University Press, New York, pp 55–115
de Waal FBM, Luttrell LM (1985) The formal hierarchy of rhesus macaques: an investigation of the bared-teeth display. Am J Primatol 9:73–85. https://doi.org/10.1002/ajp.1350090202
delBarco-Trillo J, Drea CM (2014) Socioecological and phylogenetic patterns in the chemical signals of strepsirrhine primates. Anim Behav 97:249–253. https://doi.org/10.1016/j.anbehav.2014.07.009
delBarco-Trillo J, Burkert BA, Goodwin TE, Drea CM (2011) Night and day: the comparative study of strepsirrhine primates reveals socioecological and phylogenetic patterns in olfactory signals: comparative study of primate urinary signals. J Evol Biol 24:82–98. https://doi.org/10.1111/j.1420-9101.2010.02145.x
delBarco-Trillo J, Sacha CR, Dubay GR, Drea CM (2012) Eulemur, me lemur: the evolution of scent-signal complexity in a primate clade. Philos Trans R Soc B 367:1909–1922. https://doi.org/10.1098/rstb.2011.0225
Dobson SD (2009) Socioecological correlates of facial mobility in nonhuman anthropoids. Am J Phys Anthropol 139:413–420. https://doi.org/10.1002/ajpa.21007
Drea CM, Scordato ES (2008) Olfactory communication in the ringtailed lemur (Lemur catta): form and function of multimodal signals. In: Hurst JL, Beynon RJ, Roberts SC, Wyatt TD (eds) Chemical signals in vertebrates. Springer, New York, pp 91–102
Dröscher I, Kappeler PM (2013) Defining the low end of primate social complexity: the social organization of the nocturnal white-footed sportive lemur (Lepilemur leucopus). Int J Primatol 34:1225–1243. https://doi.org/10.1007/s10764-013-9735-3
Dunbar RIM (1993) Co-evolution of neuro-cortex size, group size and language in humans. Behav Brain Sci 16:681–694
Dunbar RIM (2009) The social brain hypothesis and its implications for social evolution. Ann Hum Biol 36:562–572. https://doi.org/10.1080/03014460902960289
Dunbar RIM, Shultz S (2010) Bondedness and sociality. Behaviour 147:775–803. https://doi.org/10.1163/000579510X501151
Egnor SER, Hauser MD (2004) A paradox in the evolution of primate vocal learning. Trends Neurosci 27:649–654. https://doi.org/10.1016/j.tins.2004.08.009
Elgar MA (2015) Integrating insights across diverse taxa: challenges for understanding social evolution. Front Ecol Evol 3:124. https://doi.org/10.3389/fevo.2015.00124
Endler JA (1992) Signals, signal conditions, and the direction of evolution. Am Nat 139:S125–S153. https://doi.org/10.1086/285308
Endler JA, Westcott DA, Madden JR, Robson T, Phillips P (2005) Animal visual systems and the evolution of color patterns: sensory processing illuminates signal evolution. Evolution 59:1795–1818. https://doi.org/10.1554/04-669.1
Estes RD (1967) The comparative behavior of Grant’s and Thomson’s gazelles. J Mammal 48:189. https://doi.org/10.2307/1378022
Fedurek P, Slocombe KE (2011) Primate vocal communication: a useful tool for understanding human speech and language evolution? Hum Biol 83:153–173. https://doi.org/10.3378/027.083.0202
Fichtel C, Manser M (2010) Vocal communication in social groups. In: Kappeler P (ed) Animal behaviour: evolution and mechanisms. Springer Verlag, Heidelberg, pp 29–54
Fichtel C, Hammerschmidt K, Jürgens U (2001) On the vocal expression of emotion. A multi-parametric analysis of different states of aversion in the squirrel monkey. Behaviour 138:97–116. https://doi.org/10.1163/15685390151067094
Fischer J (1998) Barbary macaques categorize shrill barks into two call types. Anim Behav 55:799–807. https://doi.org/10.1006/anbe.1997.0663
Fischer J, Wadewitz P, Hammerschmidt K (2016) Structural variability and communicative complexity in acoustic communication. Anim Behav 134:229–237. https://doi.org/10.1016/j.anbehav.2016.06.012
Fischer J, Farnworth MS, Sennhenn-Reulen H, Hammerschmidt K (2017) Quantifying social complexity. Anim Behav 130:57–66. https://doi.org/10.1016/j.anbehav.2017.06.003
Fleishman LJ (1992) The influence of the sensory system and the environment on motion patterns in the visual displays of Anoline lizards and other vertebrates. Am Nat 139:S36–S61. https://doi.org/10.1086/285304
Freeberg TM (2006) Social complexity can drive vocal complexity. Psychol Sci 17:557–561. https://doi.org/10.1111/j.1467-9280.2006.01743.
Freeberg TM, Krams I (2015) Does social complexity link vocal complexity and cooperation? J Ornithol 156:125–132. https://doi.org/10.1007/s10336-015-1233-2
Freeberg TM, Ord TJ, Dunbar RIM (2012a) The social network and communicative complexity: preface to theme issue. Philos Trans R Soc B 367:1782–1784. https://doi.org/10.1098/rstb.2011.0294
Freeberg TM, Dunbar RIM, Ord TJ (2012b) Social complexity as a proximate and ultimate factor in communicative complexity. Philos Trans R Soc B 367:1785–1801. https://doi.org/10.1098/rstb.2011.0213
Fröhlich M, Wittig RM, Pika S (2016) Play-solicitation gestures in chimpanzees in the wild: flexible adjustment to social circumstances and individual matrices. Roy Soc open sci 3:160278. https://doi.org/10.1098/rsos.160278
Gamba M, Friard O, Riondato I, Righini R, Colombo C, Miaretsoa L, Torti V, Nadhurou B, Giacoma C (2015) Comparative analysis of the vocal repertoire of Eulemur: a dynamic time warping approach. Int J Primatol 36:894–910. https://doi.org/10.1007/s10764-015-9861-1
Greene LK, Grogan KE, Smyth KN, Adams CA, Klager SA, Drea CM (2016) Mix it and fix it: functions of composite olfactory signals in ring-tailed lemurs. R Soc Open Sci 3:160076. https://doi.org/10.1098/rsos.160076
Griebel U, Oller KD (2008) Evolutionary forces favoring communicative flexibility. In: Oller DK, Griebel U (eds) Evolution of communicative flexibility: complexity, creativity, and adaptability in human and animal communication. MIT Press, Cambridge, pp 9–41
Gustison ML, Le Roux A, Bergman TJ (2012) Derived vocalizations of geladas (Theropithecus gelada) and the evolution of vocal complexity in primates. Philos Trans R Soc B 367:1847–1859. https://doi.org/10.1098/rstb.2011.0218
Gustison ML, Semple S, Ferrer-i-Cancho R, Bergman TJ (2016) Gelada vocal sequences follow Menzerath’s linguistic law. Proc Natl Acad Sci U S A 113:E2750–E2758. https://doi.org/10.1073/pnas.1522072113
Gustison ML, Johnson ET, Beehner JC, Bergman TJ (2019) The social functions of complex vocal sequences in wild geladas. Behav Ecol Sociobiol. https://doi.org/10.1007/s00265-018-2612-5
Harper D (2001) Complex. In: Online etymology dictionary, https://www.etymonline.com/word/complex
Hasson O, Hibbard R, Ceballos G (1989) The pursuit deterrent function of tail-wagging in the zebra-tailed lizard (Callisaurus draconoides). Can J Zool 67:1203–1209. https://doi.org/10.1139/z89-174
Hatchwell BJ (2009) The evolution of cooperative breeding in birds: kinship, dispersal and life history. Proc R Soc Lond B 364:3217–3227. https://doi.org/10.1098/rstb.2009.0109
Hebets EA, Papaj DR (2005) Complex signal function: developing a framework of testable hypotheses. Behav Ecol Sociobiol 57:197–214. https://doi.org/10.1007/s00265-004-0865-7
Hebets EA, Barron AB, Balakrishnan CN, Hauber ME, Mason PH, Hoke KL (2016) A systems approach to animal communication. Proc R Soc B 283:20152889. https://doi.org/10.1098/rspb.2015.2889
Hill HM, Campbell C (2014) The frequency and nature of allocare by a group of belugas (Delphinapterus leucas) in human care. Int J Comp Psychol 27:501–514
Humphrey NK (1976) The social function of intellect. In: Humphrey NK, Bateson PPG, Hinde RA (eds) Growing points in ethology. Cambridge University Press, Cambridge, pp 303–317
Janik VM, Slater PJB (2000) The different roles of social learning in vocal communication. Anim Behav 60:1–11. https://doi.org/10.1006/anbe.2000.1410
Jeanson R, Fewell J, Gorelick R, Bertram S (2007) Emergence of increased division of labor as a function of group size. Behav Ecol Sociobiol 62:289–298. https://doi.org/10.1007/s00265-007-0464-5
Johnston RE (2005) Eavesdropping and scent over-marking. In: McGregor PK (ed) Animal communication networks. Cambridge University Press, Cambridge, pp 344–372
Johnstone RA (1997) The evolution of animal signals. In: Krebs JR, Davies NB (eds) Behavioural ecology: an evolutionary approach. Oxford University Press, Oxford, pp 155–178
Jolly A (1966) Lemur social behavior and primate intelligence. Science 153:501–506. https://doi.org/10.1126/science.153.3735.501
Kalan AK, Boesch C (2015) Audience effects in chimpanzee food calls and their potential for recruiting others. Behav Ecol Sociobiol 69:1701–1712. https://doi.org/10.1007/s00265-015-1982-1
Kanwal JS, Matsumura S, Ohlemiller K, Suga N (1994) Analysis of acoustic elements and syntax in communication sounds emitted by mustached bats. J Acoust Soc Am 96:1229–1254. https://doi.org/10.1121/1.410273
Kappeler PM (1998) To whom it may concern: the transmission and function of chemical signals in Lemur catta. Behav Ecol Sociobiol 42:411–421. https://doi.org/10.1007/s002650050455
Kappeler PM (2019) A framework for studying social complexity. Behav Ecol Sociobiol. https://doi.org/10.1007/s00265-018-2601-8
Kappeler PM, Clutton-Brock T, Shultz S, Lukas D (2019) Social complexity: patterns, processes, and evolution. Behav Ecol Sociobiol. https://doi.org/10.1007/s00265-018-2613-4
Kather R, Martin SJ (2015) Evolution of cuticular hydrocarbons in the Hymenoptera: a meta-analysis. J Chem Ecol 41:871–883. https://doi.org/10.1007/s10886-015-0631-5
Keenan S, Lemasson A, Zuberbühler K (2013) Graded or discrete? A quantitative analysis of Campbell’s monkey alarm calls. Anim Behav 85:109–118. https://doi.org/10.1016/j.anbehav.2012.10.014
Kershenbaum A, Ilany A, Blaustein L, Geffen E (2012) Syntactic structure and geographical dialects in the songs of male rock hyraxes. Proc R Soc Lond B 279:2974–2981. https://doi.org/10.1098/rspb.2012.0322
Kershenbaum A, Blumstein DT, Roch MA, Akçay Ç, Backus G, Bee MA, Bohn K, Cao Y, Carter G, Cäsar C, Coen M, DeRuiter SL, Doyle L, Edelman S, Ferrer-i-Cancho R, Freeberg TM, Garland EC, Gustison M, Harley HE, Huetz C, Hughes M, Hyland Bruno J, Ilany A, Jin DZ, Johnson M, Ju C, Karnowski J, Lohr B, Manser MB, McCowan B, Mercado E III, Narins PM, Piel A, Rice M, Salmi R, Sasahara K, Sayigh L, Shiu Y, Taylor C, Vallejo EE, Waller S, Zamora-Gutierrez V (2016) Acoustic sequences in non-human animals: a tutorial review and prospectus: acoustic sequences in animals. Biol Rev 91:13–52. https://doi.org/10.1111/brv.12160
Knörnschild M, Behr O, von Helversen O (2006) Babbling behavior in the sac-winged bat (Saccopteryx bilineata). Naturwissenschaften 93:451–454. https://doi.org/10.1007/s00114-006-0127-9
Krams I, Krama T, Freeberg TM, Kullberg C, Lucas JR (2012) Linking social complexity and vocal complexity: a parid perspective. Phil Trans R Soc B 367:1879–1891. https://doi.org/10.1098/rstb.2011.0222
Kroodsma DE (1977) Correlates of song organization among North American wrens. Am Nat 111:995–1008. https://doi.org/10.1086/283228
Kroodsma DE (2004) The diversity and plasticity of birdsong. In: Marler PR, Slabbekoorn H (eds) Nature’s music: the science of birdsong. Elsevier, New York, pp 108–131
Le Roux A, Cherry MI, Manser MB (2009) The vocal repertoire in a solitary foraging carnivore, Cynictis penicillata, may reflect facultative sociality. Naturwissenschaften 96:575–584. https://doi.org/10.1007/s00114-008-0506-5
Leavens DA, Russell JL, Hopkins WD (2010) Multimodal communication by captive chimpanzees (Pan troglodytes). Anim Cogn 13:33–40. https://doi.org/10.1007/s10071-009-0242-z
Leighton GM (2017) Cooperative breeding influences the number and type of vocalizations in avian lineages. Proc R Soc B 284:20171508. https://doi.org/10.1098/rspb.2017.1508
Lemasson A, Hausberger M (2011) Acoustic variability and social significance of calls in female Campbell’s monkeys (Cercopithecus campbelli campbelli). J Acoust Soc Am 129:3341–3352. https://doi.org/10.1121/1.3569704
Liebal K, Waller BM, Slocombe KE, Burrows AM (2013) A multimodal approach to primate communication. In: Liebal K, Waller BM, Burrows AM, Slocombe KE (eds) Primate communication: a multimodal approach. Cambridge University Press, Cambridge, pp 104–127
Lorenz K (1941) Vergleichende Bewegungsstudien an Anatiden. J Ornithol 89:194–293
MacLean EL, Merritt DJ, Brannon EM (2008) Social complexity predicts transitive reasoning in prosimian primates. Anim Behav 76:479–486. https://doi.org/10.1016/j.anbehav.2008.01.025
Malacarne G, Cucco M, Camanni S (1991) Coordinated visual displays and vocal duetting in different ecological situations among Western Palearctic non-passerine birds. Ethol Ecol Evol 3:207–219. https://doi.org/10.1080/08927014.1991.9525369
Manser MB, Jansen DAWAM, Graw B, Hollén LI, Bousquet CAH, Furrer RD, Le Roux A (2014) Vocal complexity in meerkats and other mongoose species. Adv Stud Behav 46:281–310. https://doi.org/10.1016/B978-0-12-800286-5.00006-7
Marler P (1977) The structure of animal communication sounds. In: Bullock TH, Evans EF (eds) Recognition of complex acoustic signals. Dahlem Konferenzen, Berlin, pp 17–35
Marler P, Dufty A, Pickert R (1986) Vocal communication in the domestic chicken: II. Is a sender sensitive to the presence and nature of a receiver? Anim Behav 34:194–198. https://doi.org/10.1016/0003-3472(86)90023-0
Masin S, Massa R, Bottoni L (2004) Evidence of tutoring in the development of subsong in newly-fledged Meyer’s parrots Poicephalus meyeri. An Acad Bras Cienc 76:231–236. https://doi.org/10.1590/S0001-37652004000200006
Mathevon N, Charrier I, Jouventin P (2003) Potential for individual recognition in acoustic signals: a comparative study of two gulls with different nesting patterns. C R Biol 326:329–337. https://doi.org/10.1016/S1631-0691(03)00072-6
Mathevon N, Aubin T, Vielliard J, da Silva ML, Sebe F, Boscolo D (2008) Singing in the rain forest: how a tropical bird song transfers information. PLoS One 3:e1580. https://doi.org/10.1371/journal.pone.0001580
Matos RJ, McGregor PK (2002) The effect of the sex of an audience on male-male displays of Siamese fighting fish (Betta splendens). Behaviour 139:1211–1221. https://doi.org/10.1163/15685390260437344
May-Collado LJ, Agnarsson I, Wartzok D (2007) Phylogenetic review of tonal sound production in whales in relation to sociality. BMC Evol Biol 7:136. https://doi.org/10.1186/1471-2148-7-136
McComb K, Reby D (2005) Vocal communication networks in large terrestrial mammals. In: McGregor PK (ed) Animal communication networks. Cambridge University Press, Cambridge, pp 372–389
McComb K, Semple S (2005) Coevolution of vocal communication and sociality in primates. Biol Lett 1:381–385. https://doi.org/10.1098/rsbl.2005.0366
McGregor PK, Dabelsteen T (1996) Communication networks. In: Kroodsma DE, Miller EH (eds) Ecology and evolution of acoustic communication in birds. Cornell University Press, Ithaca, pp 409–425
McGregor PK, Peake TM (2000) Communication networks: social environments for receiving and signaling behaviour. Acta Ethol 2:71–81. https://doi.org/10.1007/s102110000015
Mcgurk H, Macdonald J (1976) Hearing lips and seeing voices. Nature 264:746–748. https://doi.org/10.1038/264746a0
Medvin MB, Stoddard PK, Beecher MD (1993) Signals for parent-offspring recognition: a comparative analysis of the begging calls of cliff swallows and barn swallows. Anim Behav 45:841–850. https://doi.org/10.1006/anbe.1993.1105
Mills MGL, Gorman ML, Mills MEJ (1980) The scent marking behaviour of the brown hyaena Hyaena brunnea. Afr Zool 15:240–248
Møller AP, Pomiankowski A (1993) Why have birds got multiple sexual ornaments? Behav Ecol Sociobiol 32:167–176. https://doi.org/10.1007/BF00173774
Morton ES (1977) On the occurrence and significance of motivational-structural rules in some bird and mammal sounds. Am Nat 111:855–869
Nadhurou B, Gamba M, Andriaholinirina NV, Ouledi A, Giacoma C (2015) The vocal communication of the mongoose lemur (Eulemur mongoz): phonation mechanisms, acoustic features and quantitative analysis. Ethol Ecol Evol 28:241–260. https://doi.org/10.1080/03949370.2015.1039069
Nowicki S, Searcy WA (2014) The evolution of vocal learning. Curr Opin Neurobiol 28:48–53. https://doi.org/10.1016/j.conb.2014.06.007
Ord TJ, Garcia-Porta J (2012) Is sociality required for the evolution of communicative complexity? Evidence weighed against alternative hypotheses in diverse taxonomic groups. Phil Trans R Soc B 367:1811–1828. https://doi.org/10.1098/rstb.2011.0215
Ord TJ, Stamps JA (2008) Alert signals enhance animal communication in “noisy” environments. Proc Natl Acad Sci U S A 105:18830–18835. https://doi.org/10.1073/pnas.0807657105
Ouattara K, Lemasson A, Zuberbühler K (2009) Campbell’s monkeys concatenate vocalizations into context-specific call sequences. Proc Natl Acad Sci U S A 106:22026–22031. https://doi.org/10.1073/pnas.0908118106
Partan SR (2013) Ten unanswered questions in multimodal communication. Behav Ecol Sociobiol 67:1523–1539. https://doi.org/10.1007/s00265-013-1565-y
Partan SR, Marler P (1999) Communication goes multimodal. Science 283:1272–1273. https://doi.org/10.1126/science.283.5406.1272
Partan SR, Marler P (2005) Issues in the classification of multimodal communication signals. Am Nat 166:231–245. https://doi.org/10.1086/431246
Peake TM (2005) Eavesdropping in communication networks. In: McGregor PK (ed) Animal communication networks. Cambridge University Press, Cambridge, pp 13–37
Petter J, Charles-Dominique P (1979) Vocal communication in prosimians. In: Doyle GA, Martin RD (eds) The study of prosimian behavior. Academic Press, New York, pp 247–305
Pika S (2017) Unpeeling the layers of communicative complexity. Anim Behav 134:223–227. https://doi.org/10.1016/j.anbehav.2017.07.002
Pitnick S, Jones KE, Wilkinson GS (2006) Mating system and brain size in bats. Proc R Soc Lond B 273:719–724. https://doi.org/10.1098/rspb.2005.3367
Podos J (2001) Correlated evolution of morphology and vocal signal structure in Darwin’s finches. Nature 409:185–188. https://doi.org/10.1038/35051570
Pollard KA, Blumstein DT (2011) Social group size predicts the evolution of individuality. Curr Biol 21:413–417. https://doi.org/10.1016/j.cub.2011.01.051
Pollard KA, Blumstein DT (2012) Evolving communicative complexity: insights from rodents and beyond. Philos Trans R Soc B 367:1869–1878. https://doi.org/10.1098/rstb.2011.0221
Rakotonirina H, Kappeler PM, Fichtel C (2017) Evolution of facial color pattern complexity in lemurs. Sci Rep 7:15181. https://doi.org/10.1038/s41598-017-15393-7
Ramsier MA, Cunningham AJ, Finneran JJ, Dominy NJ (2012) Social drive and the evolution of primate hearing. Philos Trans R Soc B 367:1860–1868. https://doi.org/10.1098/rstb.2011.0219
Rapaport L, Haight J (1987) Some observations regarding allomaternal caretaking among captive Asian elephants (Elephas maximus). J Mammal 68:438–442. https://doi.org/10.2307/1381494
Ravignani A, Norton P (2017) Measuring rhythmic complexity: a primer to quantify and compare temporal structure in speech, movement, and animal vocalizations. J Lang Evol 2:4–19. https://doi.org/10.1093/jole/lzx002
Riesch R, Ford JKB, Thomsen F (2008) Whistle sequences in wild killer whales (Orcinus orca). J Acoust Soc Am 124:1822–1829. https://doi.org/10.1121/1.2956467
Robinson DW (2008) Entropy and uncertainty. Entropy 10:493–506. https://doi.org/10.3390/e10040493
Rosenthal MF, Wilkins MR, Shizuka D, Hebets EA (2018) Dynamic changes in display architecture and function across environments revealed by a systems approach to animal communication. Evolution 72:1134–1145. https://doi.org/10.1111/evo.13448
Rowe C (1999) Receiver psychology and the evolution of multicomponent signals. Anim Behav 58:921–931. https://doi.org/10.1006/anbe.1999.1242
Ruch H, Zürcher Y, Burkart JM (2018) The function and mechanism of vocal accommodation in humans and other primates. Biol Rev 93:996–1013. https://doi.org/10.1111/brv.12382
Ryan MJ, Tuttle MD, Rand AS (1982) Bat predation and sexual advertisement in a Neotropical anuran. Am Nat 119:136–139. https://doi.org/10.1086/283899
Santana SE, Dial TO, Eiting TP, Alfaro ME (2011) Roosting ecology and the evolution of pelage markings in bats. PLoS One 6:e25845. https://doi.org/10.1371/journal.pone.0025845
Santana SE, Alfaro JL, Alfaro ME (2012) Adaptive evolution of facial colour patterns in Neotropical primates. Proc R Soc Lond B 279:2204–2211. https://doi.org/10.1098/rspb.2011.2326
Santana SE, Alfaro JL, Noonan A, Alfaro ME (2013) Adaptive response to sociality and ecology drives the diversification of facial colour patterns in catarrhines. Nat Commun 4:2765. https://doi.org/10.1038/ncomms3765
Sewall KB (2015) Social complexity as a driver of communication and cognition. Integr Comp Biol 55:384–395. https://doi.org/10.1093/icb/icv064
Shannon CE, Weaver W (1951) The mathematical theory of communication. Philos Rev 60:398–400. https://doi.org/10.2307/2181879
Shapiro AD, Tyack PL, Seneff S (2011) Comparing call-based versus subunit-based methods for categorizing Norwegian killer whale, Orcinus orca, vocalizations. Anim Behav 81:377–386. https://doi.org/10.1016/j.anbehav.2010.09.020
Shultz S, Dunbar RIM (2006) Both social and ecological factors predict ungulate brain size. Proc R Soc Lond B 273:207–215. https://doi.org/10.1098/rspb.2005.3283
Shultz S, Dunbar RIM (2007) The evolution of the social brain: anthropoid primates contrast with other vertebrates. Proc R Soc Lond B 274:2429–2436. https://doi.org/10.1098/rspb.2007.0693
Slocombe KE, Zuberbühler K (2007) Chimpanzees modify recruitment screams as a function of audience composition. Proc Natl Acad Sci U S A 104:17228–17233. https://doi.org/10.1073/pnas.0706741104
Slocombe KE, Waller BM, Liebal K (2011) The language void: the need for multimodality in primate communication research. Anim Behav 81:919–924. https://doi.org/10.1016/j.anbehav.2011.02.002
Smith CL, Evans CS (2013) A new heuristic for capturing the complexity of multimodal signals. Behav Ecol Sociobiol 67:1389–1398. https://doi.org/10.1007/s00265-013-1490-0
Smith CL, Taylor A, Evans CS (2011) Tactical multimodal signaling in birds: facultative variation in signal modality reveals sensitivity to social costs. Anim Behav 82:521–527. https://doi.org/10.1016/j.anbehav.2011.06.002
Snowdon CT, Elowson AM, Roush RS (1997) Social influences on vocal development in new world primates. In: Snowdon CT, Elowson (eds) Social influences on vocal development. Cambridge University Press, Cambridge, pp 234–248
Stirling I, Thomas JA (2003) Relationships between underwater vocalizations and mating systems in phocid seals. Aquat Mamm 29:227–246
Symonds MRE, Elgar MA (2008) The evolution of pheromone diversity. Trends Ecol Evol 23:220–228. https://doi.org/10.1016/j.tree.2007.11.009
Taborsky B, Oliveira RF (2012) Social competence: an evolutionary approach. Trends Ecol Evol 27:679–688. https://doi.org/10.1016/j.tree.2012.09.003
Tallerman M, Gibson KR (2012) Introduction: the evolution of language. In: Tallerman M, Gibson KR (eds) The Oxford Handbook of Language Evolution. https://doi.org/10.1093/oxfordhb/9780199541119.013.0001
Taylor AM, Reby D (2010) The contribution of source-filter theory to mammal vocal communication research: advances in vocal communication research. J Zool 280:221–236. https://doi.org/10.1111/j.1469-7998.2009.00661.x
Tecot SR, Baden AL, Romine NK, Kamilar JM (2012) Infant parking and nesting, not allomaternal care, influence Malagasy primate life histories. Behav Ecol Sociobiol 66:1375–1386. https://doi.org/10.1007/s00265-012-1393-5
Tibbetts EA (2004) Complex social behaviour can select for variability in visual features: a case study in Polistes wasps. Proc R Soc Lond B 271:1955–1960. https://doi.org/10.1098/rspb.2004.2784
Uetz GW, Roberts JA, Clark DL, Gibson JS, Gordon SD (2013) Multimodal signals increase active space of communication by wolf spiders in a complex litter environment. Behav Ecol Sociobiol 67:1471–1482. https://doi.org/10.1007/s00265-013-1557-y
Vanden Hole C, Van Daele PAAG, Desmet N, Devos P, Adriaens D (2014) Does sociality imply a complex vocal communication system? A case study for Fukomys micklemi (Bathyergidae, Rodentia). Bioacoustics 23:143–160. https://doi.org/10.1080/09524622.2013.841085
Veilleux CC, Kirk EC (2014) Visual acuity in mammals: effects of eye size and ecology. Brain Behav Evol 83:43–53. https://doi.org/10.1159/000357830
Vergara V, Barrett-Lennard LG (2008) Vocal development in a Beluga calf (Delphinapterus leucas). Aquat Mamm 34:123–143. https://doi.org/10.1578/AM.34.1.2008.123
Vogt K, Zimmermann F, Kölliker M, Breitenmoser U (2014) Scent-marking behaviour and social dynamics in a wild population of Eurasian lynx Lynx lynx. Behav Process 106:98–106. https://doi.org/10.1016/j.beproc.2014.04.017
Wadewitz P, Hammerschmidt K, Battaglia D, Witt A, Wolf F, Fischer J (2015) Characterizing vocal repertoires—hard vs. soft classification approaches. PLoS One 10:e0125785. https://doi.org/10.1371/journal.pone.0125785
Waller BM, Liebal K, Burrows AM, Slocombe KE (2013) How can a multimodal approach to primate communication help us understand the evolution of communication? Evol Psychol 11:538–549. https://doi.org/10.1177/147470491301100305
Weiss MN, Franks DW, Croft DP, Whitehead H (2019) Measuring social complexity from association data using mixture models. Behav Ecol Sociobiol. https://doi.org/10.1007/s00265-018-2603-6
Whiten A, Byrne RW (1988) The Machiavellian intelligence hypotheses. In: Byrne RW, Whiten A (eds) Machiavellian intelligence: social expertise and the evolution of intellect in monkeys, apes, and humans. Clarendon Press/Oxford University Press, New York, pp 1–9
Wilke C, Kavanagh E, Donnellan E, Waller BM, Machanda ZP, Slocombe KE (2017) Production of and responses to unimodal and multimodal signals in wild chimpanzees, Pan troglodytes schweinfurthii. Anim Behav 123:305–316. https://doi.org/10.1016/j.anbehav.2016.10.024
Wilkins MR, Shizuka D, Joseph MB, Hubbard JK, Safran RJ (2015) Multimodal signaling in the North American barn swallow: a phenotype network approach. Proc R Soc B 282:20151574. https://doi.org/10.1098/rspb.2015.1574
Wilkinson G (2003) Social and vocal complexity in bats. In: de Waal FBM, Tyack P (eds) Animal social complexity: intelligence, culture, and individualized societies. Harvard University Press, Cambridge, pp 322–341
Wittwer B, Hefetz A, Simmon T, Murphy LEK, Elgar MA, Pierce NE, Kocher SD (2017) Solitary bees reduce investment in communication compared with their social relatives. Proc Natl Acad Sci U S A 114:6569–6574. https://doi.org/10.1073/pnas.1620780114
Wyatt TD (2003) Animals in a chemical world. In: Wyatt TD (ed) Pheromones and animal behaviour: communication by smell and taste. Cambridge University Press, Cambridge, pp 1–22
Zimmermann E (2017) Evolutionary origins of primate vocal communication: diversity, flexibility, and complexity of vocalizations in basal primates. In: Quam RM, Ramsier MA, Fay RR, Popper AN (eds) Primate hearing and communication. Springer, Cham, pp 109–140
Zuberbühler K, Lemasson A (2014) Primate communication: meaning from strings of calls. In: Lowenthal F, Lefebvre L (eds) Language and recursion. Springer, New York, pp 115–125
Zwamborn EMJ, Whitehead H (2017) Repeated call sequences and behavioural context in long-finned pilot whales off Cape Breton, Nova Scotia, Canada. Bioacoustics 26:169–183. https://doi.org/10.1080/09524622.2016.1233457
Acknowledgments
We are very grateful to Dieter Lukas and the three anonymous referees for numerous constructive comments on previous versions of this paper. Louise Peckre thanks Lluís Socías Martinez for precious discussions.
Funding
This research was funded by the Deutsche Forschungsgemeinschaft (DFG FI 929/13-1) and the German Primate Center.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by D. Lukas
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is a contribution to the Topical Collection Social complexity: patterns, processes, and evolution - Guest Editors: Peter Kappeler, Susanne Shultz, Tim Clutton-Brock, and Dieter Lukas
Electronic supplementary material
ESM 1
(DOCX 33 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Peckre, L., Kappeler, P.M. & Fichtel, C. Clarifying and expanding the social complexity hypothesis for communicative complexity. Behav Ecol Sociobiol 73, 11 (2019). https://doi.org/10.1007/s00265-018-2605-4
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00265-018-2605-4