
Abstract
Forested areas provide important breeding habitats for the turtledove (Streptopelia turtur) in Dadia-Lefkimi-Soufli National Park, Northeastern Greece. We censused the birds in two forested habitat types using the point-count technique at 60 sites during the breeding season (from mid-April to mid-June) in 2001 and 2002. We sampled vegetation structure at the same sites by measuring horizontal (tree species and density in different size classes) and vertical (percentage canopy closure in dominant, intermediate, suppressed and shrub layer) characteristics within 0.04 ha circular plots centred on the established points. Univariate and multivariate statistical techniques were employed to examine the response of the turtledove′s presence to habitat differences between used and unused sites. The results of this study indicate that habitat structure influence the presence of the turtledove during the breeding season. Middle-aged forest stands particularly those dominated by pine trees with low percentage cover in understory are likely to be beneficial to breeding the turtledove population. A combination of multipurpose forestry operations allowing development of managed woodland in mosaics with other habitat types could provide high-quality habitats for a wide range of wildlife species including game and non-game species in the area. Silvicultural methods of maintaining appropriate breeding habitat for turtledove in Dadia-Lefkimi-Soufli National Park (DLS NP), which are in conflict with commercial forestry, are discussed.
Similar content being viewed by others


Introduction
The turtledove (Streptopelia turtur) is a summer visitor to Europe that has undergone a moderate decline between the 1970s and the 1990s both in range and abundance (Heath et al. 2000). The major threat to the turtledove is the nesting habitat degradation and removal (Browne et al. 2004) and its dietary changes, which are caused by the spatial and temporal availability of food during the breeding season (Browne and Aebischer 2003). In addition, other factors that contributed in the turtledove decline are associated with wintering grounds and during migration (Browne and Aebischer 2001). The turtledove numbers have been stabilised in most Central European countries in the last decades, but the species is continuing to decline in most of its breeding range and especially in countries containing sizeable populations, such as Russia, Spain, Turkey and Ukraine (Tucker and Heath 1994). Consequently, the species has an unfavourable conservation status (SPEC category 3, Species of European Conservation Concern), and it is estimated to be declining. However, the turtledove plays an important role in Greece due to its economic value. It is considered one of the main game bird species during the opening (late August) of the hunting season in Greece (Thomaides et al. 2000).
Turtledoves feed not only on weed seeds (Dias and Fontoura 1996) but also on cultivated seeds, particularly wheat and oilseed rapes (Browne and Aebischer 2001, 2003). The principal habitat requirement for nesting turtledoves is woodland areas close to open weedy patches for feeding (Gillings and Fuller 1998; Hinsley and Bellamy 2000; Browne and Aebischer 2003; Browne et al. 2004). Several studies have investigated the breeding ecology of the turtledove in Europe (Holzwarth 1971; Bijlsma 1985; Peiró 1990; Nankinov 1994; Dias and Fontoura 1996; Aubineau and Boutin 1998; Browne and Aebischer 2003; 2004), but no quantitative information is available on its breeding habitat preferences. Such information is needed for a more complete understanding of its breeding ecology to be able to help wildlife and forest managers decide how to manipulate habitat structure to achieve greater nesting sites availability. Understanding the species′ breeding habitat requirements are important implications for conservation. The aim of this study was to provide a tool for management by identifying the most suitable habitat management strategies to apply to the nesting habitats for the conservation and/or enhancement of the turtledove population, which is likely in conflict with other bird species in the area. Thus, the objectives of this study were: (a) to investigate the nesting habitat requirements of turtledoves in wooded areas and (b) to propose appropriate management practices in order to maintain suitable breeding habitats for the species.
Materials and methods
Study area
Dadia-Lefkimi-Soufli National Park (hereafter DLS NP) is well known for its valuable raptor and reptile populations (Bakaloudis et al. 1998a, b) as well as for its wildlife diversity (Grill and Cleary 2003; Kati and Sekercioglu 2006). It is located in the central part of Evros Province (40°59′–41°15′ N, 26°19′–26°36′ E) with elevations from 20 to 620 m above sea level (asl). The climate is sub-Mediterranean with a strong continental character: dry summers and cold winters. Mean monthly temperatures in the area range from 4°C in January to 25°C in July. Mean annual precipitation is 664 mm.
The area (43,326.01 ha) comprises different habitat types, but it is characterised by high landscape diversity and a patchy distribution of these habitat types. Forested habitats cover approximately 58.96% of the DLS NP (pine forest, oak forest and mixed pine–oak forest), followed by farmlands (intensively and non-intensively cultivated land, 18.84%), partially forested habitats (degraded oak and degraded pine forest 12.80%), barren (rocky areas and grasslands with sparse trees or shrubs, 7.13%) and scrublands (2.27%). The pine forest type dominated by pure Calabrian pine (Pinus brutia) or mixed Calabrian and black pine (Pinus nigra). Situated in the central part of the study area, approximately 80–500 m asl and characterised by a mixture of even- and uneven-aged stands. The mixed pine–oak forest type is principally found in the north-western part of the DLS NP, approximately 200–600 m asl and dominated by pine trees in the upper layer and oaks in the middle layer. Sessile oak (Quercus sessiliflora) and Hungarian oak (Quercus frainetto) are the main deciduous trees, while other shrub species (Phyllirea latifolia, Erica arborea, Juniperus oxycedrus, Arbutus andrachne, Carpinus orientalis, etc.) are found in small groups in the understory.
The DLS NP is particularly important for wildlife because it offers the only extensively wooded region along the Evros Valley. It had been a Wildlife Reserve from 1980, but recently (2006), it was declared a National Park. Two strictly protected cores were established for the conservation of birds of prey, covering approximately 16.8% of the DLS NP. There are basic differences concerning the forest management inside the cores and the buffer zone (forest surrounding the core areas). Human activities are prohibited in these cores and a Specific Management Plan should now take place in order to apply management measures. The forested and partially forested areas in the buffer zone are managed for a multiplicity of purposes (wood production, soil protection, water source management, livestock production, hunting, recreation development and aesthetics), but the major target is the wildlife diversity conservation. In order to meet the need for uneven-aged stands, which always contains some large trees across the compartments, a selection system (single-tree and group selective felling) with regeneration operations is planned. The DLS NP is controlled and managed by the local Forestry Service in Soufli, and recently, a Management Authority has taken over responsibilities for the region.
Methods
We censused turtledoves at 60 permanent survey plots (Morrison 1984) in two forested habitat types (pure pine forest and mixed pine–oak forest) using the point-count technique during the breeding season (from mid-April to mid-June) in 2001 and 2002 (Bibby et al. 2000). Survey plots were placed at least 200 m apart and 100 m away from habitat borders or roads to avoid edge effect. Doves′ abundance was recorded for 10 min at each of the 60 survey plots, six times in total. The turtledove coo vocalisations and sightings were noted within a radius of 30 m at each observation point (Bibby et al. 2000; Calladine et al. 1999; Slabbekoorn et al. 1999). Censuses started 1 h after sunrise and were performed within 4 h on days without rain and strong wind to reduce detectability problems. The order in which the survey plots were visited was randomised among the three observation days to avoid time of day effect.
We analysed vegetation structure at the same sites by measuring horizontal (tree species and density in different size classes) and vertical (percent canopy closure in dominant, intermediate, suppressed and shrub layer) characteristics within 0.04 ha circular plots centred on the established points using a modification of methods of James and Shugart (1970). Within each plot, all trees greater than 5 cm in diameter at breast height (DBH) were identified to species and classified according to DBH into five diameter classes (Table 1). Two randomly orientated cross lines-transects totalling 20 points at 2-m intervals were made within each plot, and the canopy closure (%) at different layers was estimated using a Geographic Resource Solutions vertical densitometer.
Statistical analysis
Variables expressed as percentage were arcsine transformed and those expressed as non-percentage were √x transformed prior to statistical analyses (Zar 1996). Univariate two-tailed tests were used to compare vegetation structure between used and unused sites. Normally distributed variables were analysed using t test for independent samples, but those not meeting normality assumptions were analysed using the non-parametric equivalent Mann–Whitney U test.
Multivariate statistical techniques were employed to examine the response of the turtledove′s presence to habitat differences between used and unused sites (Noon 1981; Morrison 1984; McGarigal et al. 2000). A two-group discriminant function analysis (DFA) was conducted in order to determine which vegetational variables were best discriminated between used and unused sites to assess the relative importance of each variable to this discrimination and to generate hypotheses explaining why turtledoves prefer to breed in some kind of habitats and not in others (Cavallaro et al. 1981).
All statistical analyses were performed using the statistical package SPSS (release 12.0 for windows), and statistical significance was set at P < 0.05.
Results
We recorded the presence of breeding turtledoves in 27 plots (used sites), while we did not observe birds in the remaining 33 plots (unused sites) during the six censuses of the study.
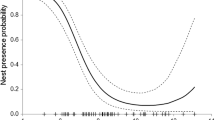
Univariate comparisons showed that six of 12 variables differed significantly (P < 0.05) between used and unused sites (Table 2). Used sites were in areas with a greater percentage of canopy cover of dominant and intermediate trees (Fig. 1) and a higher density of medium size class (pineDBH3 = 21–30 cm) pine trees (Fig. 2). Unused sites were more often in stands with a greater percentage canopy cover of suppressed trees and percentage of canopy cover of shrubs (Fig. 1) and a higher density of mature (pineDBH5 > 45 cm) pine trees (Fig. 2). Therefore, the results confirm that used sites were not randomly distributed within the forested land in the study area.

Mean percentage (%) of canopy closure at used and unused turtledove breeding sites, according to canopy layer. An asterisk indicates a statistically significant difference at 0.05 level of probability

Mean density of trees (no./0.04 ha) at used and unused turtledove breeding sites, according to diameter classes. An asterisk indicates a statistically significant difference at 0.05 level of probability
The stepwise discriminant analysis resulted in a significant discrimination (Wilk′s lambda = 0.417, F (2,57) = 39.827, P < 0.0001, eigenvalue = 1.397) between used and unused sites by turtledoves based on 12 habitat variables. The relative importance of the independent variables in predicting the dependent was determined from the standardised function coefficients (Table 3). The density of pine trees, which ranged from 21 to 30 cm and the percentage of canopy cover of intermediate tree layer were the most important contributors to the separation of the groups. These two variables were identified by the stepwise DFA as the subset of variables that achieve the maximum discrimination between used and unused sites. They were weighted positively (standardised function coefficients 0.895 and 0.653, for pineDBH3 and %intermediate, respectively) towards the used sites. Therefore, sites with high values for these variables were associated with potential breeding sites while sites with low values were less suitable for breeding.
Overall, the discriminant function correctly classified 85% of the sites indicating high success of classification for used and unused sites (Table 4). Of the 27 used sites, 23 (85.2%) were classified correctly, while five (15.2%) of the 33 unused sites were classified incorrectly as used sites. Thus, it could be used as a measure of differences between used and unused turtledove sites.
Discussion
Both univariate analysis and discriminant function analysis show that habitat structure is an important factor in determining its suitability for the turtledove during the breeding season in the DLS NP.
In the study area, turtledoves bred in forested stands with a low percentage of suppressed trees and shrubs layer, a smaller number of mature pine trees, a high density of medium-sized class pine trees and a higher percentage of canopy cover of intermediate layer than at unused sites. This result is similar to that reported in previous studies for woodland areas in the UK (Fuller and Moreton 1987; Hinsley et al. 1995; Dias and Fontoura 1996; Browne and Aebischer 2003; Browne et al. 2004). The habitat features highlighted by the present study not only occur in medium-aged pure pine stands but also in medium-aged mixed pine–oak stands where pines dominated oaks, and these stands were thinned to reduce the vegetation of understory (shrubs and young trees). Furthermore, these stands also experienced a mixture of single-tree and group selection silvicultural system (Smith et al. 1997). In the above silvicultural systems, the timber is removed either as single mature trees or in small groups of mature trees in a 10-year rotational period. This kind of tree removal creates small openings providing important sites for natural regeneration, while they can be used as feeding sites for many forest-dwelling bird species that depend on weed seeds from the forest floor (Browne and Aebischer 2003; Fuller et al. 2007). In addition, the forest structure with uneven-aged stands is important in wildlife management that requires horizontal and vertical diversity of habitats (Harris and Harris 1997).
However, it is known that the forested land in the study area supported high densities of birds of prey where most of them are nested in mature pine forest stands (Bakaloudis et al. 1998a, 2001). Generally, because bird species numbers varied considerably in their forest habitat requirements (Kati and Sekercioglu 2006), an integrated management plan to maintain the regional biodiversity should consider all stages of forest growth, from small forest openings to mature and old forest, in a broad area such as DLS NP.
Concluding, forest management recommendations in relation to the turtledove should be integrated with the results from any studies of other wildlife species with specific niche requirements (e.g. raptors, cavity nesting birds) to produce guidelines for sustainable forestry which would accommodate the needs of a wide range of wildlife species and not just one or two groups of bird species. For conservation purposes and to aid the biodiversity of the area, the maintenance of the proportion of the existing land uses (open land/forested land) should be followed to establish both feeding and nesting habitats for as many wildlife species as possible (Bakaloudis et al. 1998b). In particular, forest management practices that enhance the feature of suitable breeding habitats for the turtledove should be carefully applied in conjunction with those that benefit raptors′ breeding habitats (Smith et al. 1997; Petty 1998; Browne and Aebischer 2001). The turtledove is a game species providing an important economic activity in the region of the DLS NP (Thomaides et al. 2000), and so the forest management should be oriented towards the maintenance of a patchy distribution of uneven-aged stands and open habitats. This study demonstrates differences in habitat use by turtledoves as breeding sites among areas subjected to different forestry practices. This is done in order to predict the response of any wildlife species, including the turtledove. We, therefore, need to know more about the patterns of habitat use as breeding sites, which are associated with silvicultural manipulations when these alterations take place.
References
Aubineau J, Boutin J-M (1998) L’impact des modalités de gestion du maillage bocager sur les colombidés (Columbidae) nicheurs dans l’ouest de la France. Gib Faun Sauv. Game Wildl 15:55–63
Bakaloudis DE, Vlachos CG, Holloway G (1998a) Habitat use by Short-toed Eagle Circaetus gallicus and their reptilian prey during the breeding season in Dadia Forest (North-eastern Greece). J Appl Ecol 35:821–828
Bakaloudis DE, Vlachos CG, Nastis A, Holloway G (1998b) Distribution of raptors and reptiles in different habitat types in Dadia-Lefkimi-Soufli forest complex, north-eastern Greece. In: Waterhouse A, McEwan E (eds) Landscapes, livestock and livelihoods in European less favoured areas. SAC Auchincruive, Ayr, pp 63–67
Bakaloudis DE, Vlachos CG, Papageorgiou NK, Holloway G (2001) Nest site habitat selected by short-toed eagle (Circaetus gallicus) in Dadia-Lefkimi-Soufli forest complex, north-eastern Greece. Ibis 143:391–401. doi:10.1111/j.1474-919X.2001.tb04940.x
Bibby CJ, Burgess ND, Hill DA, Mustoe SH (2000) Bird Census Techniques. Academic, London
Bijlsma RG (1985) De broedbiologie van de Tortelduif Streptopelia turtur. Vogeljaar 33:225–232
Browne S, Aebischer N (2001) The role of agricultural intensification in the decline of the turtledove Streptopelia turtur. English Nature, Peterborough Research Project No. 421
Browne S, Aebischer N (2003) Habitat use, foraging ecology and diet of turtledoves Streptopelia turtur in Britain. Ibis 145:572–582. doi:10.1046/j.1474-919X.2003.00185.x
Browne S, Aebischer N (2004) Temporal changes in the breeding ecology of European Turtledoves Streptopelia turtur in Britain, and implications for conservation. Ibis 146:125–137. doi:10.1111/j.1474-919X.2004.00235.x
Browne S, Aebischer N, Yfantis G, Marchant J (2004) Habitat availability and use by turtledoves Streptopelia turtur between 1965 and 1995: an analysis of Common Bird Census data. Bird Study 51:1–11
Calladine J, Buner F, Aebischer NJ (1999) Temporal variations in the singing activity and the detection of turtledoves Streptopelia turtur: implications for surveys. Bird Study 46:74–80
Cavallaro JI, Menke JW, Williams WA (1981) Use of discriminant analysis and other statistical methods in analyzing microhabitat utilization of Dusky-footed woodrats. In: Capen DE (ed) The use of multivariate statistics in studies of wildlife habitats. USDA Forest Service, Vermont, pp 222–231 GTR RM-87
Dias S, Fontoura AP (1996) A dieta estival da rôla-brava (Streptopelia turtur) no sul de Portugal. Rev Flor 9:227–241
Fuller RJ, Moreton BD (1987) Breeding bird populations of Kentish sweet chestnut (Castanea sativa) coppice in relation to age and structure of the coppice. J Appl Ecol 24:13–27. doi:10.2307/2403784
Fuller RJ, Smith KW, Grice PV, Currie FA, Quine CP (2007) Habitat change and woodland birds in Britain: implications for management and future research. Ibis 149(Suppl. 2):261–268. doi:10.1111/j.1474-919X.2007.00775.x
Gillings S, Fuller RJ (1998) Changes in bird populations on sample lowland English farms in relation to loss of hedgerows and other non-crop habitats. Oecologia 116:120–127. doi:10.1007/s004420050570
Grill A, Cleary DFR (2003) Diversity patterns in butterfly communities of the Greek nature reserve Dadia. Biol Conserv 114:427–436. doi:10.1016/S0006-3207(03)00070-3
Harris E, Harris J (1997) Wildlife Conservation in Managed Woodlands and Forests, 2nd edn. Research Studies, Taunton
Heath M, Borggreve C, Peet C (2000) European Bird Populations: Estimates and Trends. BirdLife International (Birdlife Conservation Series No 10), Cambridge
Hinsley SA, Bellamy PE (2000) The influence of hedge structure, management and landscape context on the value of hedgerows to birds: A review. J Wildl Manage 60:33–49
Hinsley SA, Bellamy PE, Newton I, Sparks TH (1995) Habitat and landscape factors influencing the presence of individual breeding bird species in woodland fragments. J Avian Biol 26:94–104. doi:10.2307/3677057
Holzwarth G (1971) Ornithologischer Sammelbericht für Baden-Württemberg. Anz Ornithol Gesell Bayern 10:174–182
James FC, Shugart HH Jr (1970) A quantitative method of habitat description. Aubudon Field Notes 24:727–736
Kati V, Sekercioglu CH (2006) Diversity, ecological structure, and conservation of the landbird community of Dadia reserve, Greece. Divers Distrib 12:620–629. doi:10.1111/j.1366-9516.2006.00288.x
McGarigal K, Cushman S, Stafford S (2000) Multivariate Statistics for Wildlife and Ecology Research. Springer, Berlin
Morrison ML (1984) Influence of sample size on discriminant function analysis of habitat use by birds. J Field Ornithol 55:330–335
Nankinov D (1994) The breeding biology of the Turtledove (Streptopelia turtur) in Bulgaria. Gib Faun Sauv. Game Wildl 11:155–165
Noon BR (1981) Techniques for sampling avian habitats. In: Capen DE (ed) The use of multivariate statistics in studies of wildlife habitats. USDA Forest Service, Vermont, pp 42–52 GTR RM-87
Peiró V (1990) Aspectos de la reproducción de la tórtola común (Streptopelia turtur, L.) en Madrid. Mediterranea (Serie Biologia) 12:89–96
Petty SJ (1998) Ecology and Conservation of Raptors in Forests. Forestry Commission Bulletin 118, London
Slabbekoorn H, de Kort S, ten Cate C (1999) Comparative analysis of perch-coo vocalizations in Streptopelia doves. Auk 116:737–748
Smith DM, Larson BC, Kelty MJ, Ashton PMS (1997) The Practice of Silviculture: Applied Forest Ecology. Wiley, New York
Thomaides C, Logothetis G, Karabatzakis T, Christoforidou G (2000) Monitoring of game species population trends and harvest in Greece, 1994–1997. Hellenic Hunters Confederation, Athens
Tucker GM, Heath MF (1994) Birds in Europe. Their Conservation Status. BirdLife Conservation, Cambridge Series No. 3
Zar JH (1996) Biostatistical Analysis. Prentice-Hall, London
Acknowledgments
We wish to thank Mr A. Vasileiou who helped with map constructions and R. Taylor, S. Alatzias, K. Bakaloudis, P. Goudiakas, E. Vlachou, E. Dalmari, T. Papadopoulos and D. Tsalagas who provided assistance in the field. The authors declare that the experiments comply with the Greek and EU laws.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by E. Hadjisterkotis
Rights and permissions
About this article
Cite this article
Bakaloudis, D.E., Vlachos, C.G., Chatzinikos, E. et al. Breeding habitat preferences of the turtledove (Streptopelia turtur) in the Dadia-Soufli National Park and its implications for management. Eur J Wildl Res 55, 597–602 (2009). https://doi.org/10.1007/s10344-009-0287-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10344-009-0287-y