
Abstract
Fuel hazard reduction treatments such as prescribed fire and mastication are widely used to reduce fuel hazard. These treatments help protect people from wildfire, yet may not be mutually beneficial for people and ecosystems in areas adapted to infrequent crown fire. Short-term studies indicate that some fuel hazard reduction treatments can be detrimental to biodiversity and ecosystem function, suggesting that land managers face an acute dilemma between protecting people or ecosystems. However, the long-term ecological trajectories and fuel hazard outcomes of fuel treatments are poorly understood. Using a 13-year replicated experimental study, we evaluated how shrub cover, non-native species abundance, native species diversity, and an obligate seeder responded to fuel treatments in California’s northern chaparral. The fuel hazard reduction treatments (fire and mastication) and their seasons of implementation (fall, winter, and spring) had unique influences on plant communities. Untreated controls had continuous shrub canopy with no understory throughout the study. Recovery of shrubs after mastication was slower than recovery after fire. Ten years after treatment, shrub cover in fire treatments and spring mastications produced 1 % to 2 % less cover than the control, whereas fall mastications produced 8 % less cover than the control. The number of non-native plants, including non-native annual grasses, was higher after mastication treatments compared to fire treatments after 10 years. Surprisingly, mastication treatments also increased cover of an uncommon native shrub that is an obligate seeder. The season of treatment also influenced these outcomes, but to a lesser extent than treatment type. Long-term shrub species composition did not follow the trends of short-term species composition of shrub recruitment. Based on these findings, we concluded that fuel hazard reduction treatments only reduce shrub cover for approximately 10 years, and can change plant community composition, suggesting that thorough consideration of the decision to use fuel hazard reduction treatments is warranted.
Resumen
Los tratamientos de reducción de combustibles como las quemas prescriptas y el triturado son ampliamente usados para reducir el peligro potencial de incendios. Estos tratamientos ayudan a proteger a la gente de los incendios, aunque pueden no ser mutuamente beneficiosos para la gente y los ecosistemas en áreas adaptadas a incendios de copa poco frecuentes. Estudios de corto plazo indican que los tratamientos de reducción de combustible pueden ser perjudiciales para la biodiversidad y el funcionamiento de los ecosistemas, sugiriendo que los gestores del territorio se enfrentan al dilema de tener que optar entre proteger a la gente o a los ecosistemas. Por otra parte, las trayectorias ecológicas y los resultados finales sobre el peligro de incendios por parte de los tratamientos de combustibles a largo plazo son aún poco entendidos. Usando un estudio experimental replicado durante 13 años, evaluamos como la cobertura de arbustos, la abundancia de especies no nativas, la diversidad de especies nativas y una especie que se reproduce obligadamente por semillas, responden a distintos tratamientos de reducción de combustible en el chaparral del norte de California. Los tratamientos de reducción de combustible (quemas prescriptas y triturado) y las estaciones en que fueron implementados (otoño, invierno, y primavera) tuvieron influencias únicas en las comunidades vegetales. Los tratamientos de control presentaban doseles arbustivos continuos sin sotobosque a través de todo el periodo de estudio. La recuperación de los arbustos después de aplicado el tratamiento de triturado fue más lento que en el caso de las quemas prescriptas. Diez años post tratamientos, la cobertura de arbustos en el tratamiento de quemas y en el de triturado en primavera produjeron de 1 % a 2 % menos de cobertura que el control, mientras que el triturado durante el otoño produjo 8 % menos cobertura que el control. Diez años después de haberse realizado los tratamientos, el número de especies no nativas incluyendo pastos anuales no nativos, fue mayor en el tratamiento de triturado que en el tratamiento de quemas prescriptas. Sorprendentemente, los tratamientos de triturado también incrementaron la cobertura de un arbusto nativo poco común que se reproduce estrictamente por semilla. La estación en que se llevaron a cabo los tratamientos también influenciaron estos resultados, aunque en menor medida que el tipo de tratamiento. La composición de arbustos en el largo plazo no siguió las tendencias del corto plazo en relación al reclutamiento de arbustos. Basados en estos resultados, concluimos que los tratamientos de reducción de combustible solo reducen la cobertura de arbustos por aproximadamente 10 años, y pueden cambiar la composición de la comunidad de plantas, sugiriendo que una profunda consideración en la decisión de usar estos tratamientos de reducción de combustible sea garantizada.
Similar content being viewed by others


Introduction
Wildfires near the wildland urban interface (WUI) can cause large losses of human life and structures (Stephens et al. 2009, Syphard et al. 2014). Wildfire losses and suppression costs are expected to increase with climate change in many areas in the US (Westerling and Bryant 2008). Wildland fire risk reduction through fuel reduction is therefore a high priority for WUI management (Dicus and Scott 2006, Stephens et al. 2009), yet can impact natural communities by, for example, shifting the composition of plant communities (Briese 1996, Merriam et al. 2006). In California’s chaparral, fuel hazard reduction treatments can facilitate the invasion of non-native species in the short term (Merriam et al. 2006, Potts and Stephens 2009), but the long-term influences of these treatments on plant communities and fire risk are poorly understood and have not been well studied.
Fuel reduction may be particularly harmful to California’s chaparral, which has unusually high biodiversity despite its small land area (Keeley 2002, Keeley and Davis 2007). Chaparral, like most mediterranean shrublands, is highly fire resilient and historically has experienced high-severity, stand-replacing fires every 30 to 100 years (Keeley and Davis 2007). Historically, Native Americans burned chaparral to promote grasslands for textiles and food (Vale 2002). Though adapted to infrequent fire, chaparral plant communities can be extirpated by more frequent disturbances (Syphard et al. 2007). Today, frequent accidental ignitions increasingly convert chaparral from native shrubland to non-native annual grassland and drastically reduce species diversity, especially in southern California (Haidinger and Keeley 1993). Chaparral conversion to grassland is especially likely under global-change-type drought (Syphard et al. 2007, Pratt et al. 2013). Non-native plant invasion can create a positive feedback cycle: once non-native plants establish, they can increase fire frequency, which favors more non-native plants (Brooks et al. 2004, Brennan and Keeley 2015). Changes in plant community structure can also negatively affect habitat for wildlife such as birds and small mammals (Lillywhite 1977, Longhurst 1978, Bleich and Holl 1982, Seavy et al. 2008). The effects of frequent disturbance on chaparral should not be confounded with disturbance effects on ecosystems adapted to frequent, low-severity fire, such as most California forests. In frequent-fire ecosystems, most fuel hazard reduction treatments both reduce fuel hazard and restore native plant communities (Stephens et al. 2012), but this is not necessarily the case for infrequent-fire ecosystems like chaparral (Schwilk et al. 2009).
The differences between forest fire regimes and those of chaparral are due to the unique characteristics of chaparral shrubs, which are the drivers of chaparral fire regimes. Chaparral shrubs form a continuous canopy layer, the foliage of which can be difficult to ignite but, once ignited, burns at 500 °C to 2000 °C due to volatile oils and dense twigs (Quinn and Keeley 2006). Continuous shrub canopy excludes understory plants, including highly flammable non-native annual grasses and other noxious weeds (D’Antonio and Vitousek 1992, Mack and D’Antonio 1998, Brooks et al. 2004, Keeley 2004). As such, in native chaparral, there is little to no surface fuel. Both prescribed fire and mastication in chaparral reduce fuel hazard in the short term, lowering potential flame length and rate of spread (Brennan and Keeley 2015). These two treatment types differ in that mastication reduces fuel hazard by converting upright shrub branches to surface fuels, whereas prescribed fire consumes fuels.
Land managers must consider operational, social, and ecological factors when choosing between mastication and prescribed fire treatments for fuel hazard reduction. Managers often prefer mastication, in which heavy machinery shreds vegetation, over prescribed fire because the latter involves more regulatory and social barriers (Gill and Stephens 2009, Moritz et al. 2014). Mastication treatments require less technical expertise than prescribed fire and their application is not reliant on an air quality permit. Nearby residents may also be more comfortable with mastication than high-intensity prescribed chaparral fire (Winter et al. 2002, Mayberry 2011).
In chaparral, the surface fuels that result from mastication are small pieces of dead wood 10 cm to 40 cm in length and 5 cm to 10 cm in width (Potts et al. 2010). This surface fuel layer would likely burn at lower intensity and for less time than intact shrubs because it is more compact and the fuel particles are smaller. In addition, the residence time of heat in this dead woody layer is likely short due to the small diameter and length of this debris. Brennan and Keeley (2015) found that, eight years after mastication, evidence suggested that downed woody debris is reduced, but that shrubs recovered and a novel herbaceous layer is formed. However, these findings may be difficult to extrapolate as the study was a chronosequence and does not have pretreatment measurements to account for the influence of previous plant communities.
There are limitations to mastication: it is infeasible on steep slopes, costs more per hectare than fire, and cannot occur in winter when rain softens the soil (Hartsough et al. 2008). Mastication also has conservation drawbacks relative to fire: it can promote non-native plant invasion because it kills more individual shrubs and delays canopy closure longer, allowing more time for non-natives to invade (Keeley et al. 2005). These herbaceous non-native plants, especially non-native annual grasses, are more flammable because they cure earlier than native plants, therefore lengthening the wildfire season (Brooks et al. 2004).
Despite the disadvantages of mastication, prescribed fire can also be controversial because it commonly occurs outside the historical fire season, (i.e., in winter or spring and not summer or fall; Parker 1987 b). Winter and spring burns are advantageous for managers because they have higher fuel moistures, leading to lower rates of fire spread and thus lower the risk of fire escaping. Burning outside the summer and fall wildfire season is also subject to fewer air quality constraints, and personnel and equipment are more readily available. However, winter and spring burns can be of ecological concern because they may lead to lower post-fire germination of obligate seeders like Ceanothus cuneatus (Hook.) Nutt. (buckbrush) and some rare herbs (Parker 1987 a, b; Keeley 2002). Research on fire season influence on chaparral shrub composition, however, is sparse and contradictory; one lab study suggests a seasonal influence on obligate seeders (Le Fer and Parker 2005) while field studies do not suggest an influence on shrub composition (Dunne et al. 1991, Beyers and Wakeman 2000). Season may also play an important role in competition between natives and non-natives; the timing of treatment may allow non-natives to establish before natives begin to germinate and grow.
Research on long-term treatment effects in chaparral is sparse. No studies have included pretreatment measurements to account for the influence of local site conditions and previous plant communities. To address this knowledge gap, we conducted a long-term experiment with pretreatment and post-treatment measurements to better understand how fuels and plant communities change after fuel hazard reduction treatments in chaparral. Broadly, we examined the long-term (10-year) ecological consequences of fuel hazard reduction in chaparral, a system adapted to infrequent high-severity fire. We focused on two main questions:
-
(1)
Which fuel hazard reduction treatment and season combination fosters native plant diversity and structure?
-
(2)
Which fuel hazard reduction treatment and season combination minimizes non-native species invasion and persistence?
By answering these questions, we will help managers decide if they should treat chaparral, what type of treatment they should use, and when treatments should be applied.
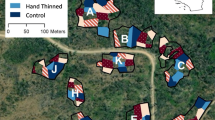
To address these questions, we conducted an experiment in chaparral and evaluated changes in plant communities over 13 years following fuel hazard reduction treatments (fire or mastication), with a seasonal component (fall, winter, and spring; Table 1; Figure 1). To our knowledge, this was one of only a few replicated and long-term studies in mediterranean shrublands with before-treatment measurements, controls, and two treatments applied across multiple seasons to address impacts on biodiversity.

Treatment type, season, and year were distributed across both northern and southern study regions in California, USA. The southern units were located at the University of California Hopland Research and Extension Center, and the northern units were mostly located at US Bureau of Land Management’s South Cow Mountain OHV (Off-Highway Vehicle) Recreation Area. Two other northernmost units were located on adjacent private lands.
Methods
Study Site
We conducted this study in northern California’s Interior Coast Range chaparral, approximately 50 km inland from the Pacific Ocean and 175 km north of San Francisco, near Ukiah, California, USA (39°N, 123°W). Vegetation is chaparral, dominated by Adenostoma fasciculatum Hook. & Arn. (chamise) and ecologically similar to chamise-dominated chaparral throughout California (Figure 2; Keeley and Davis 2007). In experimental units, chamise is approximately 75 % of the plant cover while other shrubs make up nearly 25 % of plant cover, including Ceanothus cuneatus (buckbrush), Baccharis pilularis DC (coyotebrush), Heteromeles arbutifolia (Lindley) Roemer (toyon), and assorted Arctostaphylos Adans. species. Soils are shallow, rocky, and moderately acidic, derived from weathered sandstone and shale. The experimental units are 214 m to 305 m above sea level on steep (25 % to 55 %), southern-facing and western-facing slopes. The region has a typical mediterranean climate with hot, dry summers and cool, wet winters. (Potts and Stephens 2009, Potts et al. 2010).

South-facing to west-facing chamise-dominated chaparral in the interior North Coast Range, California, USA. This area had six adjoining experimental units: fire only, mastication only, and fire and mastication burned in spring and winter. Photo credit: D. Fry.
Study Design
We chose late successional chaparral in areas where disturbances had been absent for at least 40 years. Experimental units were clustered in the University of California Hopland Research and Extension Center and at US Bureau of Land Management South Cow Mountain OHV Recreational Area and on adjacent private land. We divided the study area into 24 experimental units of nearly two hectares each, including four replicates of each treatment type and untreated control units (Table 1; Figures 1 and 2). There were two treatments (mastication and fire) across three seasons (fall, winter, and spring), which occurred between 2001 and 2003 (Table 1, Appendix A Figure 1). Fall treatments occurred in November, winter treatments occurred in January, and spring treatments occurred in April to early June (Table 1).
Operational limitations influenced study design in three ways. First, we limited mastication treatment to lower-grade slopes (3 % to 25 %) for equipment maneuverability and safety, whereas fire treatment and control units had slopes ranging from 4 % to 37 %. Second, there was no winter mastication treatment because masticators can damage seasonally wet roads and slopes. Third, prescribed fires require favorable weather conditions to safely burn, therefore not all fire treatments were completed in the same year. Beyond these limitations, we randomly assigned treatments and controls to experimental units and treatment year (Figure 1, Appendix A Figure 1).
The treatments were unique in application but had similar effects on vegetation cover, reducing it by 90 % to 100 %. We ignited prescribed fires with drip torches at the slopes’ bases, creating upslope head fires. Given the nature of fire, these treatments had some heterogeneity in their fuel reduction. Mastication by a track bulldozer with a front-mounted rotating toothed drum shredded aboveground biomass and left surface woody debris less than 5 cm deep and discontinuous (Figure 3). The debris ranged from 10 cm to 40 cm in length by 5 cm to 10 cm in width (Figure 4). The masticator did not disturb sub-surface soil or root systems, but there may have been some soil compaction and surface damage to the lignotubers of shrubs. Treatment heterogeneity was minimal in mastication treatments since the equipment operator performed systematic passes through vegetation.

Fuel hazard reduction treatments included (A) prescribed fire, (B) mastication, and (C) control. Photo credits: D. Fry, J. Potts, and K. Wilkin, respectively.

While the fuel hazard reduction treatments reduced vegetation, mastication redistributed all of the shrub biomass to the ground, creating a discontinuous layer of surface fuel (top left and bottom left) whereas fire consumed the fine materials and left larger diameter stems standing (top right and bottom right). Photos: J. Potts and D. Fry
Sample Design
Fifteen permanent line transects were randomly located and installed in each of the 24 experimental units prior to treatment (n = 360). Transects were 15 m in length. Shrub, vine, and tree (as defined by USDA NRCS 2016) species composition and cover were recorded continuously along each transect. Within each of the 24 experimental units, we established five permanent understory plots, 2.5 m in radius, in which we counted individuals of each species (n = 120). We measured burned and masticated treatment units in the summer prior to treatment; during the first, second, and third summers after treatment; and in 2012, which was nine, ten, or eleven summers after treatment. We measured untreated areas once between 2001 and 2004, and again in 2012.
We assessed non-native and native components of the plant community to investigate our questions.
Which fuel hazard reduction treatment and season combination fosters native plant communities? We assessed native plant community using shrub cover, shrub richness, and cover of the dominant obligate seeder, Ceanothus cuneatus (buckbrush), from transect measurements (Appendix B Table 1).
Which fuel hazard reduction treatment and season combination minimizes non-native plant invasion and persistence? We assessed non-native species invasion using non-native annual grass and all non-native plant densities from understory plots (Appendix B Table 1). The plants’ ecological harmfulness ratings were included as a covariate in the analysis. Harmfulness ratings are from the California Integrated Pest Management program based on a combination of ecological impact, invasive potential, and current distribution. (Bell et al. 2015).
Analysis
We constructed statistical models to test chaparral succession differences between treatment type and season of implementation. Particularly, we examined how the different treatment and season combinations affected the trajectory of the plant community and how they differed from untreated communities. We used linear and generalized linear mixed effects models to test the response variable significance between treatments types (prescribed fire, mastication, and the untreated control) and their season of application (spring, summer, and fall), and to account for the repeated measurements at each transect or quadrant through time (Bates et al. 2015, Kuznetsova 2016). In total, there were six treatment-season combinations including the control (Table 1). We examined the main effect of how each treatment-season combination changed through time and the interaction of treatment-season and years since treatment. We also included environmental variables that have been found to be significant in previous studies and may influence plants response to treatments (Appendix A Figures 1 and 2), including percent slope, solar radiation index (unitless; McCune and Keon 2002), and precipitation (cm). Yearly precipitation is often correlated with annual plant abundance and reduced precipitation is correlated with increased mortality in forests and shrublands (Pitt and Heady 1978, Pratt et al. 2013). Therefore, we used survey year annual precipitation for understory plants and precipitation one year post treatment for shrubs (August to July), as fixed effect explanatory variables in the models (data from University of California Hopland Research and Extension Center 2014; Appendix A Figure 1). In addition, we modeled the effect of random variability between and within sites (experimental unit and sample unit) and spatial blocking (University of California Hopland Research and Extension Center, US Bureau of Land Management’s South Cow Mountain OHV Recreational Area, or private ranch) to account for repeated measure.
The understory sample unit (five sample plots for each of the 24 experimental units, n = 120) was not included in the analysis because of the sparse and variable plant densities at that scale. Rather, we summarized understory plant density at the experimental unit scale (n = 24 for each year sampled).
We evaluated shrubs at the sample transect scale in which the response variables were shrub cover and species richness along a transect divided by the length of the transect. There were 15 transects for each experimental unit (n = 24) for each year sampled, resulting in 360 transects per year.
We selected models based on the type of data (including density and proportion), dispersion of the data, model residuals, and AIC. We tested model families and chose the model family with reasonable dispersion and residuals and the lowest AIC (Mazerolle 2017). For models with plant density as response variables, we tested Gaussian, square root transformation, and Poisson error (Appendix B Table 1). Density of non-native plants and annual grasses required transformed zero-inflated models (Fournier et al. 2012, Skaug et al. 2012). For shrub cover and its proportion data, we assumed a binomial error distribution and used the transect length as an offset (Appendix B Table 1). We conducted all analyses in R 3.1.2 (R Development Core Team 2008).
Results
Native Plant Diversity and Structure
Shrub cover. All shrubs that we encountered were native species. Untreated areas had nearly 100 % shrub cover throughout the study, which was significantly higher than all fire treatment units and mastication treatment units throughout the study (P < 0.001; Figure 5, Appendix C Table 1). All treatments reduced shrub cover in the near term (e.g., within the first three years of treatment). Differences between treatments were evident two growing seasons after treatments. Shrub cover in treated areas increased at greater rates than in untreated areas (P < 0.001). After about 10 years, shrub cover in areas treated by prescribed fire and mastication had rebounded to 98 % and 88 %, respectively, of the levels of shrub cover in untreated controls (P < 0.001).

Responses to treatments and the control throughout the study for shrub cover, shrub richness, and Ceanothus cuneatus (buckbrush) cover. Solid lines represent the regression coefficients, shaded areas are the 95 % confidence intervals, dotted lines are the boundaries of the 95% confidence interval, and points are experimental unit means for each survey year. The untreated controls were modeled as if they had been treated one year after their initial sampling period, and pretreatment measurements were modeled as controls.
Mastication treatments resulted in lower shrub cover than did fire (Figure 5, Appendix C Table 1). Ten years after treatments, areas with mastication applied in the fall had the lowest shrub cover compared to other treated areas: fall fire, spring fire, winter fire, and spring mastication had all produced 8 % to 10 % more shrub cover than did the fall mastication treatment (P < 0.001).
The season of application within each treatment type had significant effects on shrub cover (Figure 5, Appendix C Table 1). Within both fire and mastication treatments, fall treatments had lower shrub cover than spring treatments one year after treatment (by 18 % and 10 %, respectively), but the differences narrowed 10 years after treatment to 8 % and 2 %, respectively (P < 0.001 and P < 0.03, respectively). All treated areas experienced increases in shrub cover of between 3 % and 7 % per year throughout the study.
Slope and solar radiation did not influence shrub cover (P > 0.4 and P > 0.3, respectively).
Native shrub richness. Untreated areas had greater shrub richness than fall mastication areas and spring fire areas one year after treatment (P < 0.04; Figure 5, Appendix C Table 2, Appendix D Table 1). Untreated areas had a richness of 2.3 species per 15 m transect. One year after the fall mastication and spring fire treatments, each had 1.7 species per 15 m transect, and spring mastication had 1.9 species per 15 m transect (P < 0.03, P < 0.04, and P = 0.10, respectively). Through time, shrub species richness increased after mastication and, by 10 years after treatment, species richness in mastication treatment areas was similar to the untreated controls. In contrast, species richness after spring fire treatments remained lower than in the untreated areas throughout the study. Fall fire areas and winter fire areas had species richness similar to the untreated areas throughout the study (P > 0.3).
There were few significant differences in shrub species richness between fire and mastication treatment areas (Figure 5, Appendix C Table 2). Mastication applied in the fall produced slightly more species, about 0.3 species per 15 m transect, than winter fires one year after treatment (P < 0.03). Mastication applied in the spring produced a slightly lower rate of species accumulation through time than spring fires (only about 0.03 species per year per 15 m transect; P<0.01).
Solar radiation did not influence shrub richness (P > 0.34), and slope marginally influenced shrub richness (P > 0.07).
Dominant obligate seeder shrub cover. The untreated areas had significantly higher buckbrush cover than did fire and mastication treatment areas immediately after treatment (P < 0.001; Figure 5, Appendix C Table 3). Through time, the cover of buckbrush slightly decreased in the untreated areas from 0.2 % to 0.002 % cover (P < 0.001). After treatment, buckbrush cover increased in fall burned areas (P < 0.001), and it increased to a larger magnitude in mastication treatment areas (P < 0.001). A decade after treatment, buckbrush in masticated areas had between 0.44 % and 0.79 % greater cover than untreated areas (P < 0.001). In contrast, spring and winter burned areas still had ten times less buckbrush cover than the untreated areas. Fall fire treatment areas had 0.01 % buckbrush cover a decade after treatment, which is five times greater cover than the untreated areas 10 years after the study started (P < 0.001).
Season also influenced buckbrush cover within fire treatments: fall fire treatments resulted in greater buckbrush cover than did spring treatments (P < 0.001; Figure 5, Appendix C Table 3), and these differences marginally increase through time (P < 0.09). We could not compare buckbrush cover in winter fire treatment areas to other treatment areas because models describing winter fire treatments would not compute. Buckbrush was extremely uncommon in winter fire treatment areas and, if present, it had very low cover. In contrast, there was no seasonal differences in buckbrush cover within mastication treatment areas.
Slope and solar radiation did not influence buckbrush cover (P > 0.86 and P > 0.81, respectively).
Non-Native Plants
Non-native annual grass density. Throughout the study period, untreated areas had significantly lower non-native annual grass density than did the fall mastication areas (P < 0.001; Figure 6, Appendix C Table 4, Appendix E Table 1). No other treatments produced significantly different non-native grass density than the untreated areas. Mastication applied in the fall produced 4340 non-native annual grass plants m−2 one year after treatment (P < 0.001). In contrast, in the untreated areas, the number of non-native grasses was less than 10 plants per square meter per year throughout the study.

Responses to treatments and the control throughout the study for non-native plant densities, including annual grasses and all non-native plants by harmfulness rating from the California Integrated Pest Management (Bell et al. 2015). Estimates and standard error are based on the square root of the plant count per five meters squared. Solid lines represent the regression coefficients, shaded areas are the 95 % confidence intervals, dotted lines are the boundaries of the 95 % confidence interval, and points are the square root of the density within 5 m2 for each survey year. The untreated controls were modeled as if they had been treated one year after their initial sampling period, and pretreatment measurements were modeled as controls.
Mastication treatment areas generally had greater non-native annual grass density than did fire treatment areas (Figure 6, Appendix C Table 4). All fire treatment areas each had less than 10 non-native plants m−2 throughout the study.
Season of application only significantly influenced non-native annual grass density within the mastication treatment areas (Figure 6, Appendix C Table 4). Here, mastication applied in the fall produced 4200 more non-native annual grass plants per square meter than spring-applied mastication one year after treatment (P < 0.001). Through time, this difference persisted but declined in magnitude: ten years after treatment, fall mastication areas had 200 more non-native annual grass plants per square meter than spring mastication areas (P = 0.02).
Non-native plant density by harmfulness rating. All shrubs were native; therefore, we reported only understory non-native plants, including both graminoids and herbs (Appendix E Table 1). Untreated areas had fewer non-native plants than mastication treatment areas throughout the study (P < 0.04). Untreated areas had between 50 plants m−2 and 300 plants m−2 (Figure 6, Appendix C Table 5), and fire treatment areas had similar plant densities of between 0 plants m−2 and 500 plants m−2. Most of the non-native plants in untreated areas and fire treatment areas did not have significant ecological impact ratings (Bell et al. 2015). In contrast, non-native plants invaded and persisted after mastication treatments (P < 0.01). Mastication applied in the spring produced 550 plants m−2 10 years after treatment, and fall mastication produced 4000 plants m−2 10 years after treatment. Non-native plants in masticated treatment areas also were more likely to be harmful to the ecosystem than those found in both the untreated areas and fire treatment areas, although the most harmful plants did not persist, with density near zero, 10 years after treatment (P < 0.01).
Overall, mastication treatments produced greater non-native plant densities than fire treatments (P < 0.001; Figure 6, Appendix C Tables 5, Appendix E Table 1); only fall fire and spring mastication were marginally similar (P < 0.09). In mastication treatment areas, non-native plants invaded and persisted 10 years after treatment. Fall mastication treatment areas had fewer non-native plants as time since treatment increased, although they remained in higher density than in any other treatment area (P < 0.001). In contrast, non-native plants in spring mastication treatment areas continued to increase 10 years after treatment (P < 0.001).
There was a seasonal influence across treatments: both fall treatment areas had more non-native plants than spring treatment areas 10 years after treatment, with about four times the cover for fire and mastication (P < 0.01; Figure 6, Appendix C Table 5). Of the fire treatments, fall treatments produced the most non-native plants, but still 10 to 40 times fewer than mastication treatments 10 years after treatment (P < 0.01).
Increased annual precipitation also increased non-native plants in all experimental units (P < 0.01; Appendix F Figure 1).
Discussion
The results support our hypothesis that, in chaparral, a system adapted to infrequent high-severity fire, fuel hazard reduction treatments cause some long-term negative ecological consequences. Unfortunately, there is no “best” fuel hazard reduction treatment that promotes native plant communities. Rather, all treatments promote one or more aspects of native plant communities while degrading others.
Large differences between treatment types diminished through time, but lesser differences persisted. Ten years after treatment, most treatment areas had 96 % to 98 % shrub cover, but fall mastication areas only had 88 % shrub cover. All fire treatments and spring mastication may be preferred treatments for wildlife because continuous shrub cover provides perches for birds while also protecting birds and small mammals from birds of prey (Lillywhite 1977, Longhurst 1978, Bleich and Holl 1982, Seavy et al. 2008). In contrast, fall mastication may be a preferred fuel hazard reduction treatment because it reduces the shrub-based live fuel hazard for longer, although it also promotes non-native grasses.
The shrub recovery differences among treatments may relate to the disturbance responses of shrubs, their soil seed banks, and deer browsing. Shrubs resprouted aggressively after all treatments. The masticator may have killed some shrubs by damaging their burls, whereas fire top-killed shrubs, allowing them to resprout (Figure 4). The effects of deer browsed shrub resprouts after treatments were different during the first two years after treatment. Deer browse was most evident in mastication areas because there were no burnt shrub skeletons to protect the resprouting vegetation from deer (Appendix G Figure 1). Fire also stimulated more seedlings than mastication (Potts et al. 2010) and nutrients are often enhanced immediately post fire, which often increases growth (DeBano and Conrad 1978).
Burns outside of the natural fire season are safer and easier for managers to implement, but may have unintended consequences because of seasonal differences in plant physiology, plant competition, and treatment efficacy (Knapp et al. 2009). Our data support seasonal differences in shrub regrowth for both fire and mastication treatments. Fall treatment areas had the lowest shrub cover while winter and spring treatment areas had higher shrub cover. The seasonal differences in plant regrowth after fire are sometimes attributed to soil moisture, wherein imbibed seeds are more susceptible to heat (Le Fer and Parker 2005). However, soil moistures were statistically similar across seasons immediately before treatments (Potts and Stephens 2009), likely because fall and winter burns were only completed after a drying period following rainfall. Therefore, seasonal differences in surface soil moistures were minimized. Other physiological processes also correlate with seasonal drought, including reduced water potential in mature shrubs and reduced carbohydrate storage in lignotubers, which may leave plants more susceptible to disturbance by reducing resources needed for resprouting (Pratt et al. 2013).
Another possible driver of seasonal differences is fire intensity (Knapp et al. 2009). There is reason to believe that fire behavior could have been unique in each season (Stephens et al. 2008)—live fuel moistures were greater in the spring than in the fall or winter for fire treatments (Potts and Stephens 2009). However, seasonal differences in shrub regrowth existed for both fire and mastication treatments. Thus, fire intensity alone cannot explain the differences in shrub cover; plant physiology or competition may also play a role. Early trends in shrub seedling densities did not align with seasonal differences in shrub cover; at two and three years post treatment, both fall treatment areas had the greatest seedling densities (Potts et al. 2010). Therefore, early seasonal differences in shrub cover were likely based on pre-existing shrubs that resprouted. Shrubs may regrow more slowly or be less likely to sprout after fall treatments because of compounding stressors of a harsh environment and low storage and availability of resources such as carbohydrates or water (Pratt et al. 2013).
Shrub species richness had diverse responses to treatments. Richness was reduced during the entire study by spring fire treatments, reduced for a brief period by mastication, and was not altered by fall and winter fires. Most concerning, from a conservation perspective, were the spring fire treatments in which species richness was quasi-permanently reduced by the treatment. Further research is needed to understand how shrub species composition changes after fuel hazard reduction treatments.
Buckbrush cover changed throughout the study. Ten years after treatments, the control, spring fire areas, and winter fire areas had lower cover than before treatments; fall fire areas had slightly lower cover than before treatments; and fall and spring mastication areas had greater cover than before treatments. In control areas, buckbrush cover declined due to old-age mortality and minimal recruitment; whereas, in the other areas, treatments killed all buckbrush and all cover observed after treatments was from recruitment. Most buckbrush seeds are cued by heat shock (Keeley 1987), but the flush of post-fire buckbrush seedlings found early in this study did not correspond with high long-term cover (Potts et al. 2010). For example, three years post treatment, the fall fire treatment areas had the greatest buckbrush seedling densities, with about 2 seedlings m−2, which is more than twice as many as all other treatments and seasons (Potts et al. 2010). However, in mastication treatment areas, the soil seed bank may have continued to be stimulated by solar heat intensified by sparse shrub cover (Baskin and Baskin 1998). In addition, mastication likely promoted buckbrush because competitive resprouters like chamise grew slowly, allowing non-fire cued seedlings to prosper (Biswell 1961, Keeley 1987, Wilkin et al. 2013). While differences in buckbrush cover between fire and mastication treatment areas were less than 1 % total cover, these small differences may be biologically significant given that buckbrush is widespread and important as deer forage, despite being generally uncommon (Bleich and Holl 1982, Biswell 1989). Future research should seek to understand the mechanisms by which fire-cued species respond to mastication.
Non-native plants and annual grasses were complex to model because of their strong ecological reaction to the treatments. Non-native plants were rare in control units throughout the study, were rare in fire units ten years after treatment, and abundant and common throughout masticated units throughout the study. For example, non-natives were absent in half of the five square meter plots, and these plots were generally in control units. It is likely that established shrubs out-competed non-native plants for resources where shrub cover was high. Immediately after fuel hazard reduction treatments, non-natives had relatively high, homogenous cover within treatment type, as demonstrated by the narrow 95 % confidence intervals just after treatment (Figure 4). As shrubs grew, they created a heterogeneous habitat for non-native plants wherein non-native plants were either absent underneath shrubs or abundant in the open. Widening 95 % confidence intervals through time demonstrates how variation of non-native plant densities within treatments increased through time. At 10 years after treatment, there was great estimated variation within all treatments for non-native plant densities, and they had wide and overlapping confidence intervals. These large confidence intervals were in part due to the small sample size: six treatment units with four replicates each (n = 24). Therefore, these results should be applied cautiously and additional research is needed.
Masticated areas were more susceptible to invasion by non-native annual grasses compared to fire treated areas. In masticated areas, non-native plants, including non-native annual grasses, invaded and persisted for at least 10 years post treatment, creating a shrub-grass matrix. Fire treatment areas had low non-native annual grass presence, low non-native species richness, and no noxious weeds present despite their close proximity to maintained fuel breaks, which are often referred to as weed highways (Merriam et al. 2006). In contrast, mastication treatment areas generally were not placed next to fuel breaks and yet had high non-native grass presence and high non-native plant richness throughout the study.
In masticated areas, there are negative consequences of increasing non-native annual grasses and herbs, including increased flammability and even possible extirpation of native plants (Keeley 2000, Beyers 2004, Merriam et al. 2006, Syphard and Keeley 2015). Chaparral’s rich flora that emerges after fire and disturbance can be out-competed by non-native annual grasses (Keeley et al. 1981, Beyers 2004). The non-native grass and herb species that invade chaparral are much more likely to ignite than shrubs, and their incursion into chaparral can lead to increased fire frequency (Cione et al. 2002, Brooks et al. 2004). Non-native annual grasses increase fuel continuity between natural areas and homes, increase how quickly fire spreads, and lengthen the fire season by drying earlier than native species. Thus, non-native annual grass invasion and persistence in chaparral is detrimental to people and biodiversity in the WUI (Mack and D’Antonio 1998, Brooks et al. 2004, Dickens and Allen 2014).
To maintain fuel hazard reduction, it is likely that re-treatment will be needed every 10 to 20 years, especially in the fire and spring mastication treatment areas. Re-treatments applied too soon after the initial treatment could increase susceptibility to non-native plant invasion, and newly recruited shrubs may not have deposited a robust soil seed bank yet for their replacement (Zammit and Zedler 1988). Persistent non-native plants, including annual grasses, would likely increase quickly if another treatment occurred (Haidinger and Keeley 1993, Brennan and Keeley 2015). The results of this study suggest that reducing fuel hazard in chaparral can be detrimental to native plant biodiversity, especially when mastication is used. Therefore, fuels treatments should be applied carefully, non-native plant populations monitored carefully, and other wildfire risk reduction strategies should be considered.
These results should be applied cautiously throughout mediterranean-climate shrublands because of possible differences in fire behavior, climate, and plant communities between northern California’s chaparral and other mediterranean-climate regions. Fire behavior can have limited response to fuel hazard reduction treatments in areas like southern California where foehn winds drive many chaparral fires (Keeley 2002). Foehn winds foster fires that are nearly unstoppable, though fuel reductions may facilitate safer firefighting and evacuations. Fuel hazard reduction treatments are more successful in areas with limited foehn-wind-driven fires such as central and northern California (Moritz et al. 2010). Southern California also has more frequent and severe drought, which drives higher shrub mortality after fuel hazard reduction treatment (Pratt et al. 2013). Local climates and plant varieties may cause the seemingly similar dominant plants across California’s chaparral to respond to treatments differently. Despite regional differences, our findings expand upon results from other studies in California’s chaparral (Beyers and Wakeman 2000; Keeley 2000, 2002, 2004; Merriam et al. 2006; Perchemlides et al. 2008; Brennan and Keeley 2015) and can be used in combination with local studies to aid in fuel hazard reduction planning.
Summary
Implications drawn from this study can inform fuels management; however, as new information accrues, these implications could change.
Prescribed fire.
-
Generally fosters native plant richness and shrub cover, except for spring fires, which decrease shrub richness.
-
Reduces fuel hazard for a shorter time than mastication.
-
Decreases obligate seeder buckbrush, especially in winter and spring fires.
Mastication.
-
Fosters native shrub species, but non-native understory species invade and persist.
-
Reduces shrub fuel load more than fire, but may also increase fire frequency due to highly flammable and abundant annual grasses.
-
Increases certain native shrubs, such as the obligate seeder buckbrush.
Season of treatment.
-
Season influences shrub cover, buckbrush cover, and non-native annual grass presence to a lesser extent than treatment type.
-
Fall treatments have the slowest shrub recovery, followed by spring and then winter treatments.
-
Fall fire treatments increase buckbrush, whereas winter and spring fire treatments reduce it. Within mastication treatments, season does not influence buckbrush outcomes.
Conclusion
Land managers are challenged to simultaneously protect people from fire and protect ecosystems from harmful effects of fuel reductions. A decade after treatments, mastication and prescribed fire treatment areas had unique ecological responses. Mastication treatment areas had higher densities of non-native species, especially annual grasses, and lower shrub cover than burned areas. Fire treatment areas had little to no buckbrush, an obligate seeder and important deer browse. Lastly, the long-term plant community response did not follow the short-term response for shrub cover and seedling densities as suggested by Potts and Stephens (2009) and Potts et al. (2010).
In natural areas, we recommend fall prescribed fires because they reduce shrub cover while fostering native shrub communities and exclude non-natives ten years after treatment. When there are operational constraints to fall fires, winter or spring fires reduce shrub cover, but they also somewhat degrade the native shrub community. Increased non-native plants, especially grasses, can extend the season in which fire can ignite and spread. Despite these negative effects, mastication may still be a preferred management choice near structures because it reduces fuel load and the potential loss of people or structures during a fire. In this case, we recommend fall mastication because it reduces shrub cover and has the longest treatment efficacy. To counteract the flammable non-native plants promoted by mastication, we recommend annual mowing and grazing to reduce the flammability.
Questions remain about chaparral’s longer-term succession after fuel hazard reduction treatments, especially after repeated treatments. Shrubs regrow after treatments and treatments need to be repeated to remain effective at reducing fire risk. Treatment intervals may vary but they will inevitably need to be shorter now than the 30-year to 100-year historical fire intervals because shrubs regrow quickly and create fuel hazards. Altering fire regimes by increasing disturbance frequencies will leave these ecosystems vulnerable to possible adverse effects such as vegetation type conversion and species composition changes, especially during global-change-type drought (Keeley 2004, Pratt et al. 2013). Due to concerns for biodiversity, we recommend that land-use planners reduce the amount of WUI in chaparral to decrease the need for managers to make acute decisions between the needs of people and biodiversity (Moritz et al. 2014).
Literature Cited
Baskin, C.C., and J.M. Baskin. 1998. Seeds: ecology, biogeography, and evolution of dormancy and germination. Second edition. Academic Press, San Diego, California, USA.
Bates, D., M. Maechler, B. Bolker, and S. Walker. 2015. Fitting linear mixed-effects models using lme4. Journal of Statistical Software 67(1): 1–48. doi: 10.18637/jss.v067.i01
Bell, C.E., J.M. DiTomaso, and C.A. Wilen. 2015. Invasive plants. University of California Statewide Integrated Pest Management Program, Davis, California, USA.
Beyers, J.L. 2004. Postfire seeding for erosion control: effectiveness and impacts on native plant communities. Conservation Biology 18(4): 947–956. doi: 10.1111/j.1523-1739.2004.00523.x
Beyers, J.L., and C.D. Wakeman. 2000. Season of burn effects in southern California chaparral. Open-file report 00-62. Pages 45–55 in: J.E. Keeley, M. Baer-Keeley, and C.J. Fotheringham, editors. Second interface between ecology and land development in California. US Department of the Interior Geological Survey, Sacramento, California, USA.
Biswell, H.H. 1961. Manipulation of chamise brush for deer range improvement. California Fish and Game 47(2): 125–144.
Biswell, H.H. 1989. Prescribed burning in California wildlands vegetation management. University of California Press, Berkeley, USA.
Bleich, V.C., and S.A. Holl. 1982. Dynamics and management of mediterranean ecosystems: management of chaparral habitat for mule deer and mountain sheep in southern California. General Technical Report PSW-58, USDA Forest Service, Pacific Southwest Forest and Range Experiment Station, Berkeley, California, USA.
Brennan, T.J., and J.E. Keeley. 2015. Effect of mastication and other mechanical treatments on fuel structure in chaparral. International Journal of Wildland Fire 24(7): 949–963. doi: 10.1071/WF14140
Briese, D.T. 1996. Biological control of weeds and fire management in protected natural areas: are they compatible strategies? Biological Conservation 77(2): 135–141. doi: 10.1016/0006-3207(96)00007-9
Brooks, M.L., C.M. D’Antonio, D.M. Richardson, J.B. Grace, J.E. Keeley, J.M. DiTomaso, R.J. Hobbs, M. Pellant, and D. Pyke. 2004. Effects of invasive alien plants on fire regimes. Bioscience 54(7): 677–688. doi: 10.1641/0006-3568(2004)054[0677:EOIAPO]2.0.CO;2
Cione, N.K., P.E. Padgett, and E.B. Allen. 2002. Restoration of a native shrubland impacted by exotic grasses, frequent fire, and nitrogen deposition in southern California. Restoration Ecology 10(2): 376–384. doi: 10.1046/j.1526-100X.2002.02038.x
D’Antonio, C.M., and P.M. Vitousek. 1992. Biological invasions by exotic grasses, the grass/fire cycle, and global change. Annual Review of Ecology and Systematics 23: 63–87.
DeBano, L.F., and C.E. Conrad. 1978. The effect of fire on nutrients in a chaparral ecosystem. Ecology 59(3): 489–497. doi: 10.2307/1936579
Dickens, S.J.M., and E.B. Allen. 2014. Exotic plant invasion alters chaparral ecosystem resistance and resilience pre- and post-wildfire. Biological Invasions 16(5): 1119–1130. doi: 10.1007/s10530-013-0566-0
Dicus, C., and M.E. Scott. 2006. Reduction of potential fire behavior in wildland-urban interface communities in southern California: a collaborative approach. Pages 729–738 in: P.L. Andrews and B.W. Butler, editors. Proceedings of a conference: fuels management—how to measure success. RMRS-P-41, USDA Forest Service, Rocky Mountain Research Station, Portland, California, USA.
Dunne, J., A. Dennis, J.W. Bartolome, and R.H. Barrett. 1991. Chaparral response to a prescribed fire in the Mount Hamilton Range, Santa Clara County, California. Madroño 38(1): 21–29.
Fournier, D., H. Skaug, J. Ancheta, J. Ianelli, A. Magnusson, M. Maunder, A. Nielsen, and J. Sibert. 2012. AD Model Builder: using automatic differentiation for statistical inference of highly parameterized complex nonlinear models. Optimization Methods Software 27(2): 233–249. doi: 10.1080/10556788.2011.597854
Gill, A.M., and S.L. Stephens. 2009. Scientific and social challenges for the management of fire-prone wildland-urban interfaces. Environmental Research Letters 4(3). doi: 10.1088/1748-9326/4/3/034014
Haidinger, T.L., and J.E. Keeley. 1993. Role of high fire frequency in the destruction of mixed chaparral. Madroño 40(3): 141–147.
Hartsough, B.R., S. Abrams, R.J. Barbour, E.S. Drews, J.D. McIver, J.J. Moghaddas, D.W. Schwilk, and S.L. Stephens. 2008. The economics of alternative fuel reduction treatments in western United States dry forests: financial and policy implications from the National Fire and Fire Surrogate Study. Forest Policy and Economics 10(6): 344–354. doi: 10.1016/j.forpol.2008.02.001
Keeley, J.E. 2001. Fire and invasive species in mediterranean-climate ecosystems of California. Pages 81–94 in: K.E.M. Galley and T.P. Wilson, editors. Proceedings of the invasive species workshop: the role of fire in the control and spread of invasive species. Tall Timbers Research Station, Tallahassee, Florida, USA.
Keeley, J.E. 2002. Fire management of California shrubland landscapes. Environmental Management 29(3): 395–408. doi: 10.1007/s00267-001-0034-Y
Keeley, J.E. 2004. Invasive plants and fire management in California mediterranean-climate ecosystems. In: M. Arianoutsou and V.P. Papanastasis, editors. Proceedings of the 10th MEDECOS conference, Rhodes Island, Greece. Millpress, Rotterdam, The Netherlands.
Keeley, J.E. 1987. Role of fire in seed germination of woody taxa in California chaparral. Ecology 68(2): 434–443. doi: 10.2307/1939275
Keeley, J.E., M. Baer-Keeley, and C.J. Fotheringham. 2005. Alien plant dynamics following fire in mediterranean-climate California shrublands. Ecological Applications 15(6): 2109–2125. doi: 10.1890/04-1222
Keeley, J.E., and F.W. Davis. 2007. Terrestrial vegetation of California. Pages 339–366 in: M.G. Barbour, T. Keeler-Wolf, and A.A. Schoenherr, editors. University of California Press, Los Angeles, USA.
Keeley, S.C., J.E. Keeley, S.M. Hutchinson, and A.W. Johnson. 1981. Postfire succession of the herbaceous flora in southern California chaparral. Ecology 62(6): 1608–1621. doi: 10.2307/1941516
Knapp, E.E., B.L. Estes, and C.N. Skinner. 2009. Ecological effects of prescribed fire season: a literature review and synthesis for managers. General Technical Report PSW-GTR-224, USDA Forest Service, Pacific Southwest Research Station, Redding, California, USA.
Kuznetsova, A., P.B. Brockhoff, and R.H.B. Christensen. 2016. LmerTest: tests in linear mixed effects models. R package version 2.0-33. <https://CRAN.R-project.org/package=lmerTest>. Accessed 2 August 2017.
Le Fer, D., and V.T. Parker. 2005. The effect of seasonality of burn on seed germination in chaparral: the role of soil moisture. Madroño 52(3): 166–174. doi: 10.3120/0024-9637(2005)52[166:TEOSOB]2.0.CO;2
Lillywhite, H.B. 1977. Effects of chaparral conversion on small vertebrates in southern California. Biological Conservation 11(3): 171–184. doi: 10.1016/0006-3207(77)90002-7
Longhurst, W.M. 1978. Responses of bird and mammal populations to fire in chaparral. California Agriculture 32(10): 9–12.
Mack, M.C., and C.M. D’Antonio. 1998. Impacts of biological invasions on disturbance regimes. Trends in Ecology & Evolution 13(5): 195–198. doi: 10.1016/S0169-5347(97)01286-X
Mayberry, J.A. 2011. Community wildfire preparedness: balancing community safety and ecosystem sustainability in southern California chaparral. Thesis, Humboldt State University, Arcata, California, USA.
Mazerolle, M.J. 2017. AICcmodavg: model selection and multimodel inference based on (Q) AIC(c). R package version 2.1-1. <https://cran.r-project.org/package=AICcmodavg>. Accessed 2 August 2017.
McCune, B., and D. Keon. 2002. Equations for potential annual direct incident radiation and heat load. Journal of Vegetation Science 13(4): 603–606. doi: 10.1111/j.1654-1103.2002.tb02087.x
Merriam, K.E., J.E. Keeley, and J.L. Beyers. 2006. Fuel breaks affect nonnative species abundance in Californian plant communities. Ecological Applications 16(2): 515–527. doi: 10.1890/1051-0761(2006)016[0515:FBANSA]2.0.CO;2
Moritz, M.A., E. Batllori, R.A. Bradstock, A.M. Gill, J. Handmer, P.F. Hessburg, J. Leonard, S. McCaffrey, D.C. Odion, and T. Schoennagel. 2014. Learning to coexist with wildfire. Nature 515(7525): 58–66. doi: 10.1038/nature13946
Moritz, M.A., T.J. Moody, M.A. Krawchuk, M. Hughes, and A. Hall. 2010. Spatial variation in extreme winds predicts large wildfire locations in chaparral ecosystems. Geophysical Research Letters 37(4): L04801. doi: 10.1029/2009GL041735
Parker, V.T. 1987a. Can native flora survive prescribed burns? Fremontia 15(2): 3–6.
Parker, V.T. 1987b. Effects of wet-season management burns on chaparral vegetation: implications for rare species. Pages 231–237 in: T.E. Elias, editor. Conservation and management of rare and endangered plants. California Native Plant Society, Sacramento, California, USA.
Perchemlides, K.A., P.S. Muir, and P.E. Hosten. 2008. Responses of chaparral and oak woodland plant communities to fuel-reduction thinning in southwestern Oregon. Rangeland Ecology & Management 61(1): 98–109. doi: 10.2111/07-026R1.1
Pitt, M.D., and H.F. Heady. 1978. Responses of annual vegetation to temperature and rainfall patterns in northern California. Ecology 59(2): 336–350. doi: 10.2307/1936378
Potts, J.B., E. Marino, and S.L. Stephens. 2010. Chaparral shrub recovery after fuel reduction: a comparison of prescribed fire and mastication techniques. Plant Ecology 210(2): 303–315. doi: 10.1007/s11258-010-9758-1
Potts, J.B., and S.L. Stephens. 2009. Invasive and native plant responses to shrubland fuel reduction: comparing prescribed fire, mastication, and treatment season. Biological Conservation 142(8): 1657–1664. doi: 10.1016/j.biocon.2009.03.001
Pratt, R.B., A.L. Jacobsen, A.R. Ramirez, A.M. Helms, C.A. Traugh, M.F. Tobin, M.S. Heffner, and S.D. Davis. 2013. Mortality of resprouting chaparral shrubs after a fire and during a record drought: physiological mechanisms and demographic consequences. Global Change Biology 20(3): 893–907. doi: 10.1111/gcb.12477
Quinn, R.D., and S.C. Keeley. 2006. Introduction to California chaparral. University of California Press, Berkeley, California, USA.
R Development Core Team. 2008. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria.
Schwilk, D.W., J.E. Keeley, E.E. Knapp, J. McIver, J.D. Bailey, C.J. Fettig, C.E. Fiedler, R.J. Harrod, J.J. Moghaddas, K.W. Outcalt, C.N. Skinner, S.L. Stephens, T.A. Waldrop, D.A. Yaussy, and A. Youngblood. 2009. The National Fire and Fire Surrogate Study: effects of fuel reduction methods on forest vegetation structure and fuels. Ecological Applications 19(2): 285–304. doi: 10.1890/07-1747.1
Seavy, N.E., J.D. Alexander, and P.E. Hosten. 2008. Bird community composition after mechanical mastication fuel treatments in southwest Oregon oak woodland and chaparral. Forest Ecology and Management 256(4): 774–778. doi: 10.1016/j.foreco.2008.05.034
Skaug, H., D. Fournier, B. Bolker, A. Magnusson, and A. Nielsen. 2012. AD Model Builder: using automatic differentiation for statistical inference of highly parameterized complex nonlinear models. Optimization Methods and Software 27(2): 233–249. doi: 10.1080/10556788.2011.597854
Stephens, S.L., M.A. Adams, J. Handmer, F.R. Kearns, B. Leicester, J. Leonard, and M.A. Moritz. 2009. Urban-wildland fires: how California and other regions of the US can learn from Australia. Environmental Research Letters 4(1): 5. doi: 10.1088/1748-9326/4/1/014010
Stephens, S.L., J.D. McIver, R.E.J. Boerner, C.J. Fettig, J.B. Fontaine, B.R. Hartsough, P.L. Kennedy, and D.W. Schwilk. 2012. The effects of forest fuel-reduction treatments in the United States. Bioscience 62(6): 549–560. doi: 10.1525/bio.2012.62.6.6
Stephens, S.L., D.R. Weise, D.L. Fry, R.J. Keiffer, J. Dawson, E. Koo, J. Potts, and P.J. Pagni. 2008. Measuring the rate of spread of chaparral prescribed fires in northern California. Fire Ecology 4(1): 74–86. doi: 10.4996/fireecology.0401074
Syphard, A.D., T.J. Brennan, and J.E. Keeley. 2014. The role of defensible space for residential structure protection during wildfires. International Journal of Wildland Fire 23(8): 1165–1175. doi: 10.1071/WF13158
Syphard, A.D., and J.E. Keeley. 2015. Location, timing and extent of wildfire vary by cause of ignition. International Journal of Wildland Fire 24(1): 37–47. doi: 10.1071/WF14024
Syphard, A.D., V.C. Radeloff, J.E. Keeley, T.J. Hawbaker, M.K. Clayton, S.I. Stewart, and R.B. Hammer. 2007. Human influence on California fire regimes. Ecological Applications 17(5): 1388–1402. doi: 10.1890/06-1128.1
University of California Hopland Research & Cooperative Extension Center. 2014. Total precipitation. <http://hrec.ucanr.edu/Weather,_Physical,_and_Biological_Data/>. Accessed 4 March 2014.
USDA NRCS [US Department of Agriculture Natural Resources Conservation Service]. 2016. The PLANTS Database. <https://plants.usda.gov/java/>. Accessed 5 February 2016.
Vale, T.R. 2002. Fire, native peoples, and the natural landscape. Island Press, Washington, D.C., USA.
Westerling, A., and B. Bryant. 2008. Climate change and wildfire in California. Climatic Change 87(Suppl 1)): 231–249. doi: 10.1007/s10584-007-9363-z
Wilkin, K.M., V.L. Holland, D. Keil, and A. Schaffner. 2013. Mimicking fire for successful chaparral restoration. Madroño 60(3): 165–172. doi: 10.3120/0024-9637-60.3.165
Winter, G.J., C. Vogt, and J.S. Fried. 2002. Fuel treatments at the wildland-urban interface: common concerns in diverse regions. Journal of Forestry 100(1): 15–21.
Zammit, C.A., and P.H. Zedler. 1988. The influence of dominant shrubs, fire, and time since fire on soil seed banks in mixed chaparral. Vegetatio 75(3): 175–187.
Acknowledgements
Many people helped with this study including University of California Hopland Research and Extension Center, especially B. Keiffer, A. Shrum, and M. Lewman; US Bureau of Land Management staff, especially J. Dawson; CalFire in Mendocino and Lake counties, especially T. Derum, M. Tolbert, and B. Baxter with their support on the prescribed fires; Eaglepoint Ranch for hosting research sites; D. Ackerly of University of California Berkeley’s Integrative Biology Department with his thoughtful reviews; and funding from Joint Fire Science Program and California Cooperative Extension. Special thanks to J. Johnston for assistance in R.
Author information
Authors and Affiliations
Corresponding author
Appendices
Appendix A. Variability in Environment and Precipitation

The distribution of environmental variables (slope, solar radiation index [SRI], and precipitation [cm] one year after treatment) varied between treatments, and was incorporated into the models to determine their significance and effect size. The bold lines are the medians, the boxes represent 50 % of the data, and each whisker represents 25 % of the data. Dots are outliers and the extreme values represent maximum or minimum value if present. When there are no outliers, the whiskers’ ends depict minimum and maximum values. Slope and solar radiation did not significantly influences shrubs (shrub cover: P > 0.4 and P > 0.3, native shrub richness: P > 0.07 and P < 0.3, and buckbrush cover: P > 0.8 and P > 0.8, respectively).

Yearly precipitation varied between treatments and growing seasons, and was incorporated into the models to determine their significance and effect size. We graphed precipitation in relation to growing seasons because treatments occurred during two years, and precipitation after treatment may have influenced outcomes. Untreated controls were randomly assigned one of the two treatment years, and are included in the growing seasons since treatment groups. Gray boxes indicate whether measurements were taken pre-treatment (0) or a number of growing seasons after treatment (1 to 11). Bold lines are the medians, and the boxes represent 50 % of the data. Outliers are dots and are the maximum value if present. When there are no outliers, the whiskers’ ends depict minimum and maximum values.
Appendix B. Model Selection
Appendix C. P-Value And Coefficient Tables
Appendix D. Shrub Cover by Species
Appendix E. Non-Native Plants
Appendix F. Precipitation and Non-Native Plant Response

Non-native plant densities significantly increased with survey year precipitation: as precipitation increased from 20 cm to 45 cm, plant densities increased from 161 plants m−2 to 362 plants m−2 (P < 0.01). Non-native annual grasses were not influenced by precipitation (P > 0.19). Precipitation amounts beyond 38 cm were confounded by the number of years since treatment; these high precipitation years only occurred two to three years post treatment. Precipitation less than 38 cm occurred throughout the study period. Curves represent the fit regression coefficients, the shaded area represents 95 % confidence intervals, and points represent an experimental unit mean for each survey year.
Appendix G. Herbivory Effects on Shrubs

Exclosures had more chamise resprouts than the surrounding area nine months after mastication in plot 1 (top). Despite shrub height being significantly different in exclosures, exclosures had cover similar to their surrounding environment and browsing did not influence shrub cover (Potts et al. 2010). The differences in shrub cover and height diminished by two and a half years after treatment.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made.
The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
To view a copy of this licence, visit https://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wilkin, K.M., Ponisio, L.C., Fry, D.L. et al. Decade-Long Plant Community Responses to Shrubland Fuel Hazard Reduction. fire ecol 13, 105–136 (2017). https://doi.org/10.4996/fireecology.130210513
Published:
Issue Date:
DOI: https://doi.org/10.4996/fireecology.130210513