
Abstract
Exposure to smoke can influence the germination of seeds in many fire-prone ecosystems, but this effect is not well studied in grasslands. Smoke treatments such as smoke water could be useful as management and restoration tools if the response of target species in natural settings is well understood. We tested eight species native to the southern High Plains region in Texas, USA, that were already known to respond to smoke water in the laboratory, for their responses in a less controlled glasshouse environment. We exposed seeds to smoke water, heat, or a combination of the two, sowed them into greenhouse flats, and observed and recorded emergence. Emergence of nearly all species was influenced by smoke water, with most species experiencing either lower emergence or longer times for emergence when exposed to high-concentration smoke water. Smoke water exposure enhanced emergence of broom snakeweed (Gutierrezia sarothrae [DC.] A. Gray) seeds, with more than twice as many treated seeds emerging than untreated seeds (germination of control seeds = 26 % ± 4.39 % SE; germination of treated seeds = 69 % ± 4.62 % SE). Because many species displayed different results in the glasshouse as compared to the laboratory, smoke treatments should be tested in the field before being used on a larger scale. Doing so will allow a better understanding of how target species might respond to smoke treatments with more realistic soils, fluctuating temperatures, and other complications encountered in the field.
Resumen
La exposición al humo puede influenciar la germinación de semillas en muchos ecosistemas propensos al fuego, aunque este efecto no ha sido bien estudiado en pastizales. Los tratamientos con humo pueden ser exitosos como herramientas de manejo o de restauración, si la respuesta de las especies clave en ambientes naturales está bien entendida. Nosotros probamos ocho especies nativas de la región de las planicies altas de Texas, EEUU, que era conocido respondían al humo y posterior humidificación en laboratorio por su respuesta en ambientes poco controlados en invernaderos. Nosotros expusimos semillas a humo, calor, y a una combinación de ambos, y luego fueron sembradas en bandejas planas en invernadero, observando y registrando su emergencia. La emergencia de casi todas las especies fue influenciada por el humo, con la mayoría de las especies mostrando una baja emergencia o mucho tiempo para emerger cuando estuvieron expuestas a una alta concentración de humo. El humo aumentó la emergencia de semillas de hierba de la serpiente o hierba de San Nicolás (Gutierrezia sarothrae [DC.] A. Gray) a más del doble en semillas tratadas con humo que en aquellas sin tratar (germinación de semillas en el control = 26 % ± 4.39 % ES; germinación de semillas tratadas = 69 % ± 4.62 % ES). Dado que distintas especies pueden mostrar dispares resultados en el invernáculo cuando se las compara con el laboratorio, el tratamiento con humo debe probarse a campo antes de ser usado a gran escala. El hacer esto permitirá conocer mejor cómo las especies clave podrían responder en suelos reales, con temperaturas fluctuantes, y otras complicaciones que se encuentran en condiciones de campo.
Similar content being viewed by others


Introduction
The effect of smoke on seed germination has received attention in several ecosystems around the world (de Lange and Boucher 1990, Egerton-Warburton 1998, Keeley and Fotheringham 2000, Read et al. 2000, van Staden et al. 2004), and is especially well studied in fire-prone, mediterranean-climate ecosystems such as those in Australia, South Africa, and California (see Keeley and Fotheringham 2000, Jefferson et al. 2014 for reviews). This effect has received much less attention in other fire-prone ecosystems such as grasslands. Nevertheless, several species native to the southern High Plains in Texas, USA, recently have been shown to be responsive to smoke treatments, including smoke water and aerosol smoke (Jefferson et al. 2008, Chou et al. 2012, Schwilk and Zavala 2012). Because the response of these species was observed in the laboratory, their response in less controlled conditions, such as when sown in soil, is unknown. Fire has historically played an important role in the Great Plains grasslands, and is still used as a management treatment, so fire cues such as smoke and heat might be useful as on-the-ground restoration treatments. Therefore, understanding how seeds of grassland species respond to smoke in more natural settings is important.
In regions where rainfall is limited or is concentrated into short periods of time, such as occurs regularly in the southern High Plains, it is important that seeds germinate quickly when soil moisture is available so that they can establish successfully (Read et al. 2000). Smoke and heat treatments have the potential to increase germination of desirable species by reducing the time required for germination (Read and Bellairs 1999). In addition, smoke and heat can also increase both the density and richness of seedlings germinating from the seedbank (Read et al. 2000).
When applying smoke treatments, two methods are used most often: aerosol and liquid treatments. Aerosol smoke (also: dry smoke, plant-derived smoke, smoke fumigation) can be generated in laboratory settings by burning dry or fresh vegetation (Landis 2000). The active compounds that promote seed germination (karrikins; Chiwocha et al. 2009) exist in all plant species and are effective in responsive species, regardless of fuel origin (Jäger et al. 1996, Adkins and Peters 2001), although the effect might be slightly different depending on the type of plant fuel used to generate the smoke (Razanamandranto et al. 2005). Other active compounds also exist, and not all species are responsive to every promotive compound (Ghebrehiwot et al. 2013, Downes et al. 2014). The second method of smoke application is the use of smoke water (also: liquid smoke, aqueous smoke; Strydom et al. 1996, Read and Bellairs 1999, Clarke et al. 2000, Bhatia et al. 2005, Abella 2009). Because the active compound in aerosol smoke is water soluble (Flematti et al. 2004), liquid smoke can be created as needed by bubbling plant-derived smoke into distilled water (Landis 2000). However, commercial smoke water, advertised especially for germination enhancement, is also available for purchase and might be more effective than aerosol smoke (Dixon and Roche 1995). From the standpoint of evaluating the effect of smoke on seeds, using smoke water has the advantage of not requiring real-time combustion, and can potentially affect larger quantities of seeds at a time (Brown and van Staden 1997).
Heat is another important fire-related cue that can influence seed germination. Exposing seeds to temperatures of 50 °C to 150 °C for 1 min to 60 min has promoted germination of plant species from many families worldwide (Enright and Kintrup 2001, Buhk and Hensen 2006, Thomas et al. 2007, Bolin 2009, Tsuyuzaki and Miyoshi 2009). The temperature of the heat treatment does not necessarily correspond to the heat measured during actual wildfires; temperatures of 80 °C to 100 °C are effective for many species (Auld and Ooi 2009), yet typical wildfire temperatures commonly reach higher than 100 °C (Wright and Bailey 1982).
To date, few studies have been done to test the effect of smoke water and heat on species native to the southern High Plains, although fire is an important ecological influence in this ecosystem and such treatments could be used as management and restoration tools. Thus far, studies that have investigated this effect for the region have done so in the laboratory (Chou et al. 2012, Schwilk and Zavala 2012).
We selected several species of plants native to the southern High Plains region of Texas that were already known to respond to smoke water in the laboratory (Chou et al. 2012), and tested seeds of these species in the glasshouse to determine their germination response to both smoke water and heat, with the objective of understanding their potential field responses to smoke treatments at a broader scale.
Methods
In a previous laboratory study (Chou et al. 2012), we tested seeds of 10 species native to the southern High Plains region for germination response to smoke water, heat, and smoke water and heat in combination. To further characterize the ability of those species to respond to smoke water and heat in more natural conditions, we subsequently tested responsive species (the forbs Astragalus crassicarpus Nutt., Coreopsis tinctoria Nutt., Monarda citriodora Cerv. ex Lag., Salvia azurea Michx. ex Lam., Salvia reflexa Hornem., Solanum elaeagnifolium Cav.; the grass Digitaria ciliaris [Retz.] Koeler; and the shrub Gutierrezia sarothrae [DC.] A. Gray) in the glasshouse, and report results here.
For those species that responded to the interaction of smoke water and heat in the previous laboratory study, we applied both treatments and their combinations. For those that responded to only smoke water or heat, we applied all levels of the effective treatment only (Table 1). As described in Chou et al. (2012), we exposed seeds to four concentrations of commercial smoke water, diluted in distilled water (control of 0, 1:100, 1:10, and 1:5), and three levels of heat (no heat, 50 °C, and 80 °C). The treatments were organized as a completely randomized factorial experiment, replicated four times (in time), with 25 seeds for each replicate.
For smoke water treatment, we soaked seeds of each species in their respective smoke water-distilled water solution separately for 20 hr at room temperature, using the commercially available aqueous smoke solution Regen 2000® (Grayson Co., Bayswater, Australia), which recommends dilution at 1:10. To apply the heat treatment, seeds were placed in a preheated oven for 5 min at the prescribed temperature, and then cooled to room temperature. Seeds in the unheated control were not heated in the oven. When both smoke water and heat were applied, seeds were first treated with heat and then with smoke water after cooling.
After treatment, we sowed seeds of each species on top of sterile potting soil and covered them with a 1 mm layer of soil in a 12 cm × 12 cm paperboard glasshouse tray. Trays with seeds were randomly and evenly arranged on only one bench in the glasshouse. We applied distilled water to moisten the soil. We checked trays daily for emergence and once we observed a seedling emergence, we removed and recorded it. The experiment ran until emergence ceased (up to 45 days, depending on the species). During the experiment, glasshouse temperatures ranged from 24 °C to 30 °C.
We calculated emergence capacity (EC, also emergence, %) and mean emergence time (MET, days) for each species and treatment as:
where n. is the number of seeds germinating at each i day, N is total number of seeds sown, and t i. is the number of days from the date of sowing until all emergence ceased (Bewley and Black 1994).
We analyzed EC with Proc Glimmix in SAS (SAS Institute, Cary, North Carolina, USA) using a logit link function to model a binomial response variable. When we identified a significant difference between treatments, we conducted pair-wise comparisons with the t-test at the 0.05 level of significance. Significant differences in MET among smoke water, heat, and their interactions were tested with either a two-way ANOVA or one-way ANOVA, depending on the number of factors tested (smoke water or heat or both). We used Levene’s test (Levene 1960) and the Shapiro-Wilk test (Shapiro and Wilk 1965) to test homogeneity of variances across treatments and normality of experimental errors within each treatment, respectively, for MET. We also used the Brown and Forsythe (1974) test when variances were heterogeneous.
Results
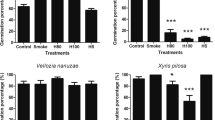
Of the eight species tested, only Monarda citriodora did not respond to smoke water (data not shown). Smoke water affected other species positively or negatively, depending on species, although germination of most species was inhibited. When treated with high-concentration (1:5) smoke water, Coreopsis tinctoria displayed approximately 14 % (± 3.47 % SE) emergence, compared to almost 80 % (± 4.14 % SE) emergence with no smoke water (Figure 1a). High-concentration (1:5) smoke water similarly suppressed the emergence capacity in Digitaria ciliaris (32 % ± 4.66 % SE emergence compared to 50 % ± 5.00 % SE emergence in untreated controls; (Figure 1b). Gutierrezia sarothrae, on the other hand, displayed a significant effect of promotion of EC as smoke water concentration increased, so that treatment by 1:100, 1:10, and 1:5 concentrations of smoke water increased the EC from the 26 % (± 4.39 % SE) of the control to 45 % (± 4.98 % SE), 59 % (± 4.92 % SE), and 69 % (± 4.62 % SE), respectively (Figure 1c). Emergence of Salvia azurea, however, was inhibited by higher smoke water concentrations, with the highest concentration treatment (1:5) essentially eliminating emergence of this species (Figure 1d).

Main effect of smoke water on the emergence capacity (%) of a) Digitaria ciliaris, b) Coreopsis tinctoria, c) Gutierrezia sarothrae, and d) Salvia azurea (mean ± SE) in the glasshouse. Means followed by the same letters are not significantly different (0.05 level).
Smoke water treatment also prolonged MET of most species. Moderate (1:10) concentration smoke water prolonged the MET of Astragalus crassicarpus (9 days ± 0.26 days SE, compared to 7 days ± 0.26 days SE for the control; Figure 2a). When treated with the 1:5 concentration of smoke water, Coreopsis tinctoria also emerged at a slower rate (11 days ± 0.94 days SE) than the control (7 days ± 0.21 days SE), but the 1:100 concentration of smoke water actually shortened the MET of this species to 5 days (± 0.24 days SE; Figure 2b). Effects on the MET of Digitaria ciliaris were only significantly different between the 1:5 and the 1:10 and 1:100 treatments; no treatment differed from the control (Figure 2c). Similar to the results for Coreopsis tinctoria, higher smoke water concentration prolonged MET of Salvia azurea by 9 days (± 2.40 days SE) compared to the control (5 days ± 0.28 days SE), but low-concentration (1:100) smoke water shortened emergence time to 4 days (± 0.21 days SE; Figure 2d). Finally, high-concentration smoke water also prolonged the MET of Salvia reflexa (21 days ± 2.91 days SE compared to approximately 9 days ± 0.65 days SE; Figure 2e).

Main effect of smoke water on the mean emergence time (days) of a) Astragalus crassicarpus, b) Coreopsis tinctoria, c) Digitaria ciliaris, d) Salvia azurea, and e) Salvia reflexa (Mean ± SE) in the glasshouse. Means followed by the same letters are not significantly different (0.05 level).
Heat was not generally effective as a single treatment in this study, although the MET of Salvia reflexa was prolonged by moderate (50 °C) and high (80 °C) heat from 6.8 days (± 1.10 days SE) to 10.6 days (± 1.49 days SE) and 10.7 days (± 1.89 days SE), respectively (data not shown). However, smoke water and heat did interact to influence the EC of three species. Medium (1:10) and high (1:5) smoke water concentrations inhibited emergence of Astragalus crassicarpus at all heat levels, although the magnitude of the effect varied by heat level (Figure 3a). High-concentration (1:5) smoke water suppressed the EC of Salvia reflexa across all levels of heat, while low (1:100) and moderate (1:10) concentrations of smoke water helped to compensate for the decrease of the EC due to the heat (50 °C or 80 °C) (Figure 3b). The interaction effects of smoke water and heat on Solanum eleagnifolum were complicated but were promotive in general. The 50 °C and 80 °C heat treatments promoted the EC of this species at 1:100 and 1:10 concentrations of smoke water. Moreover, at least one smoke water treatment enhanced the EC of this species at all levels of heat (Figure 3c).

Interaction effects of smoke water and heat on the emergence capacity (%) of a) Astragalus crassicarpus, b) Salvia reflexa, and c) Solanum eleagnifolum (mean ± SE) in the glasshouse. Means followed by the same letters are not significantly different (0.05 level) within groups.
Discussion
As in previous studies (Keeley and Fotheringham 1998, van Staden et al. 2000, Adkins and Peters 2001, Sparg et al. 2006), we found that smoke water inhibited the EC of some species (Coreopsis tinctoria and Digitaria ciliaris), especially at high (1:5) or medium (1:10) concentrations of smoke water. Similarly, the METs of Coreopsis tinctoria, Gutierrezia sarothrae, Astragalus crassicarpus, Salvia azurea, Salvia reflexa, and Digitaria ciliaris were prolonged by high- (1:5) or medium-(1:10) concentration smoke water, or both. This corresponds well to other examples in which smoke prolonged germination (Drewes et al. 1995, Daws et al. 2007). Interestingly, while most of these species experienced similar prolonging of germination in the previous laboratory study, Salvia azurea was inhibited only when also treated with heat in that same study (Chou et al. 2012). In this study, however, high-concentration smoke water inhibited emergence of Salvia azurea without heat. Other recent studies also used similar dilution levels as in this study (Norman et al. 2006, van Etten et al. 2014, Fowler et al. 2015).
Although smoke is known to inhibit some species, especially at high concentrations (Drewes et al. 1995, Daws et al. 2007), it is better known as a germination promoter. We found three species that were promoted by smoke water: Coreopsis tinctoria, Salvia azurea, and Gutierrezia sarothrae. In this study, these species experienced either increased emergence (Gutierrezia sarothrae) or decreased germination time (Coreopsis tinctoria, Salvia azurea) when exposed to some (but not all) concentrations of smoke water. This ability of smoke to promote germination in diverse species (Dixon and Roche 1995) has received increasing attention and could be the basis for using smoke or smoke water as a management or restoration tool. For example, smoke water can be sprayed over the soil surface (Abella 2009), thereby stimulating the germination of selected species. In the case of Gutierezia sarothrae, a species that is native but often considered undesirable over much of the western US (Ralphs and McDaniel 2011), smoke treatments could stimulate mass germination, thereby depleting the seedbank of seeds of this plant (Adkins and Peters 2001). Emergent plants could then be treated mechanically or chemically to remove them from the landscape before they produce further seeds. Alternatively, smoke treatment could also be used to treat seeds of desirable species before sowing, perhaps increasing their ability to quickly germinate when sowed under favorable conditions (Baxter and van Staden 1994).
However, before smoke treatments, including smoke water, are used in such a manner in the southern High Plains, their effect on seeds in the field must be further characterized. We observed striking differences between the earlier laboratory study, conducted under tightly controlled conditions, and this glasshouse study, in which conditions were less controlled (i.e., less control over temperatures, light exposure, and contact with soil rather than germination papers as in the laboratory). For example, the mean germination time (MGT) of Salvia reflexa did not respond to heat in the laboratory, but MET was retarded by heat (50 °C and 80 °C) in the glasshouse. The measures of MGT and MET are nearly similar measures of seedling development, although MET is used (as we did) when germination cannot be directly observed because seeds are covered with soil. Astragalus crassicarpus, Salvia azurea, and Solanum eleagnifolium all displayed nearly opposite results in the glasshouse as compared to the laboratory. In the glasshouse, smoke water or heat or both might interact with environmental factors such as temperature or soil (Roche et al. 1997 b), and measuring emergence, as we did in this study, rather than germination allows more time for such interactions to occur. In addition, because we used the same seedlots for both the laboratory study (Chou et al. 2012) and this study, seed afterripening may have altered some germination responses.
The inconsistent responses to smoke water or heat or both in the glasshouse compared to the laboratory suggest the necessity of testing these methods in the field to determine how factors such as soil (Read and Bellairs 1999), seed aging or ripening (Roche et al. 1997 a), darkness (Tsuyuzaki and Miyoshi 2009), and seasonality (Roche et al. 1998) might interact with smoke and heat treatments to affect the germination and produce different results. Likewise, it might be important to consider that different populations of the same species might respond to smoke treatments differently (Thomas et al. 2007). For management and conservation of shortgrass prairie, future work should focus on testing the effects of smoke water on soil-stored seedbank germination, seedling density, seedling vigor, and overall species diversity. In addition, understanding the dormancy status and germination process for each species to be tested with smoke water will greatly enhance our ability to predict the outcome of treatment. Finally, we further suggest testing with lower concentrations of Regen 2000® smoke water because, in general, 1:00, 1:10, and 1:5 concentration levels inhibited more than promoted shortgrass prairie species. Other brands of smoke water, or even direct use of the active compound (Flematti et al. 2004, Stevens et al. 2007) might work even more effectively.
Literature Cited
Abella, S.R. 2009. Smoke-cued emergence in plant species of ponderosa pine forests: contrasting greenhouse and field results. Fire Ecology 5(1): 22–37. doi: 10.4996/fireecology.0501022
Adkins, S.W., and N.C.B. Peters. 2001. Smoke derived from burnt vegetation stimulates germination of arable weeds. Seed Science Research 11: 213–222.
Auld, T.D. and M.K.J. Ooi. 2009. Heat increases germination of water-permeable seeds of obligate-seeding Darwinia species (Myrtaceae). Plant Ecology 200: 117–127. doi: 10.1007/s11258-008-9437-7
Bhatia, N.P., A.E. Nkang, K.B. Walsh, A.J. M. Baker, N. Ashwath, and D.J. Midmore. 2005. Successful seed germination of the nickel hyperaccumulator Stackhousia tryonii. Annals of Botany 96: 159. doi: 10.1093/aob/mci151
Baxter, B.J.M., and J. van Staden. 1994. Plant-derived smoke: an effective seed pre-treatment. Plant Growth Regulation 14: 279–282. doi: 10.1007/BF00024804
Bewley, J.D., and M. Black. 1994. Seeds: physiology of development and germination. Springer, New York, New York, USA. doi: 10.1007/978-1-4899-1002-8
Bolin, J.F. 2009. Heat shock germination responses of three eastern North American temperate species. Castanea 74: 160–167. doi: 10.2179/08-010.1
Brown, M.B., and A.B. Forsythe. 1974. The ANOVA and multiple comparisons for data with heterogeneous variances. Biometrics 30: 719–724. doi: 10.2307/2529238
Brown, N.A.C., and J. van Staden. 1997. Smoke as a germination cue: a review. Plant Growth Regulation 22: 115–124. doi: 10.1023/A:1005852018644
Buhk, C., and I. Hensen. 2006. “Fire seeders” during early post-fire succession and their quantitative importance in south-eastern Spain. Journal of Arid Environments 66: 193–209. doi: 10.1016/j.jaridenv.2005.11.007
Chiwocha, S.D.S., K.W. Dixon, G.R. Flmatti, E.L. Ghisalberti, D.J. Merrit, D.C. Nelson, J.M. Riseborough, S.M. Smith, and J.C. Stevens. 2009. Karrikins: a new family of plant growth regulators in smoke. Plant Science 177: 252–256. doi: 10.1016/j.plantsci.2009.06.007
Chou, Y.F., R.D. Cox, and D.B. Wester. 2012. Smoke water and heat shock influence germination of shortgrass prairie species. Rangeland Ecology and Management 65: 260–267. doi: 10.2111/REM-D-11-00093.1
Clarke, P.J., E.A. Davison, and L. Fulloon. 2000. Germination and dormancy of grassy woodland and forest species: effects of smoke, heat, darkness and cold. Australian Journal of Botany 48: 687–700. doi: 10.1071/BT99077
Daws, M.I., J. Davies, H.W. Pritchard, N.A.C. Brown, and J. van Staden. 2007. Butenolide from plant-derived smoke enhances germination and seedling growth of arable weed species. Plant Growth Regulation 51: 73–82. doi: 10.1007/s10725-006-9149-8
de Lange, J.H., and C. Boucher. 1990. Autecological studies on Audouinia capitata (Bruniaceae). I. Plant-derived smoke as a seed germination cue. South African Journal of Botany 56: 700–703. doi: 10.1016/S0254-6299(16)31009-2
Dixon, K.W., and S. Roche. 1995. The role of combustion products (smoke) in stimulating ex situ and in situ germination of Western Australian plants. Proceedings of the International Plant Propagators Society 45: 53–56.
Downes, K.S., M.E. Light, M. Posta, L. Kohout, and J. van Staden. 2014. Do fire-related cues, including smoke-water, karrikinolide, glyceronitrile and nitrate, stimulate the germination of 17 Anigozanthos taxa and Blancoa canescens (Haemodoraceae)? Australian Journal of Botany 62: 347–358.
Drewes, F.E., M.T. Smith, and J. Staden. 1995. The effect of a plant-derived smoke extract on the germination of light-sensitive lettuce seed. Plant Growth Regulation 16: 205–209. doi: 10.1007/BF00029542
Egerton-Warburton, L. 1998. A smoke-induced alteration of the sub-testa cuticle in seeds of the post-fire recruiter, Emmenanthe penduliflora Benth. (Hydrophyllaceae). Journal of Experimental Botany 49: 1317. doi: 10.1093/jxb/49.325.1317
Enright, N.J., and A. Kintrup. 2001. Effects of smoke, heat and charred wood on the germination of dormant soil-stored seeds from a Eucalyptus baxteri heathy-woodland in Victoria, SE Australia. Austral Ecology 26: 132–141. doi: 10.1046/j.1442-9993.2001.01096.x
Flematti, G.R., E.L. Ghisalberti, K.W. Dixon, and R.D. Trengove. 2004. A compound from smoke that promotes seed germination. Science 305: 977. doi: 10.1126/science.1099944
Fowler, W.M., J.B. Fontaine, N.J. Enright, and W.P. Veber. 2015. Evaluating restoration potential of transferred topsoil. Applied Vegetation Science 18: 379–390. doi: 10.1111/avsc.12162
Ghebrehiwot, H.M., M.G. Kulkarni, G. Szalai, V. Soós, E. Balázs, and J. van Staden. 2013. Karrikinolide residues in grassland soils following fire: implications on germination activity. South African Journal of Botany 88: 419–424. doi: 10.1016/j.sajb.2013.09.008
Jäger, A.K., M.E. Light, and J. van Staden. 1996. Effects of source of plant material and temperature on the production of smoke extracts that promote germination of light-sensitive lettuce seeds. Environmental and Experimental Botany 36: 421–429. doi: 10.1016/S00988472(96)01024-6
Jefferson, L.V., M. Pennacchio, and K. Havens. 2014. Ecology of plant-derived smoke: its use in seed germination. Oxford University Press, New York, New York, USA. doi: 10.1093/ac-prof:osobl/9780199755936.001.0001
Jefferson, L.V., M. Pennacchio, K. Havens, B. Forsberg, D. Sollenberger, and J. Ault. 2008. Ex situ germination responses of Midwestern USA prairie species to plant-derived smoke. The American Midland Naturalist 159: 251–256. doi: 10.1674/0003-0031(2008)159[251:ES-GROM]2.0.CO;2
Keeley, J.E., and C.J. Fotheringham. 1998. Smoke-induced seed germination in California chaparral. Ecology 79: 2320–2336. doi: 10.1890/0012-9658(1998)079[2320:SISGIC]2.0.CO;2
Keeley, J.E., and C.J. Fotheringham. 2000. Role of fire in regeneration from seed. Pages 311330 in: M. Fenner, editor. Seeds: the ecology of regeneration in plant communities. CAB International, Wallingford, England, United Kingdom.
Landis, T.D. 2000. Where there’s smoke… there’s germination? Native Plants Journal 1: 25–29. doi: 10.3368/npj.1.1.25
Levene, H. 1960. Robust tests for equality of variances. Pages 278–292 in: I. Olkin, S.G. Ghurye, W. Hoeffding, W.G. Madow, and H.B. Mann, editors. Contributions to probability and statistics: essays in honor of Harold Hotelling. Stanford University Press, Stanford, California, USA.
Norman, M.A., J.A. Plummer, J.M. Koch, and G.R. Mullins. 2006. Optimising smoke treatments for jarrah (Eucalyptus marginata) forest rehabilitation. Australian Journal of Botany 54: 571–581. doi: 10.1071/BT05140
Ralphs, M.H., and K.C. McDaniel. 2011. Broom snakeweed (Gutierrezia sarothrae): toxicology, ecology, control, and management. Invasive Plant Science and Management 4: 125–132. doi: 10.1614/IPSM-D-09-00038.1
Razanamandranto, S., M. Tigabu, L. Sawadogo, and P.C. Oden. 2005. Seed germination of eight savanna-woodland species from West Africa in response to different cold smoke treatments. Seed Science and Technology 33: 315–328. doi: 10.15258/sst.2005.33.2.05
Read, T.R., and S.M. Bellairs. 1999. Smoke affects the germination of native grasses of New South Wales. Australian Journal of Botany 47: 563–576. doi: 10.1071/BT97124
Read, T.R., S.M. Bellairs, D.R. Mulligan, and D. Lamb. 2000. Smoke and heat effects on soil seed bank germination for the re-establishment of a native forest community in New South Wales. Austral Ecology 25: 48–57. doi: 10.1046/j.1442-9993.2000.01031.x
Roche S., J.M. Koch, and K.W. Dixon. 1997a. Smoke enhanced seed germination for mine rehabilitation in the southwest of Western Australia. Restoration Ecology 5: 191–203. doi: 10.1046/j.1526-100X.1997.09724.x
Roche, S., K.W. Dixon, and J.S. Pate. 1997b. Seed aging and smoke: partner cues in the amelioration of seed dormancy in selected Australian native species. Australian Journal of Botany 45: 783–815. doi: 10.1071/BT96099
Roche, S., K.W. Dixon, and J.S. Pate. 1998. For everything a season: smoke-induced seed germination and seedling recruitment in a Western Australian Banksia woodland. Australian Journal of Ecology 23: 111–120. doi: 10.1111/j.1442-9993.1998.tb00709.x
Schwilk, D.W., and N. Zavala. 2012. Germination response of grassland species to plant-derived smoke. Journal of Arid Environments 79: 111–115. doi: 10.1016/j.jaridenv.2011.12.002
Shapiro, S.S., and M.B. Wilk. 1965. An analysis of variance test of normality (complete samples). Biometrika 52: 591–611. doi: 10.1093/biomet/52.3-4.591
Sparg, S.G., M.G. Kulkarni, and J. van Staden. 2006. Aerosol smoke and smoke-water stimulation of seedling vigor of a commercial maize cultivar. Crop Science 46: 1336–1340. doi: 10.2135/cropsci2005.07-0324
Stevens, J.C., D.J. Merritt, G.R. Flematti, E.L. Ghisalberti, and K.W. Dixon. 2007. Seed germination of agricultural weeds is promoted by the butenolide 3-methyl-2H-furo[2,3-c]pyran-2-one under laboratory and field conditions. Plant and Soil 298: 113–124. doi: 10.1007/s11104-007-9344-z
Strydom, A., A. Jäger, and J. Staden. 1996. Effect of a plant-derived smoke extract, N6-benzyladenine and gibberellic acid on the thermodormancy of lettuce seeds. Plant Growth Regulation 19: 97–100. doi: 10.1007/BF00024574
Thomas, P.B., E.C. Morris, and T.D. Auld. 2007. Response surfaces for the combined effects of heat shock and smoke on germination of 16 species forming soil seed banks in south-east Australia. Austral Ecology 32: 605–616. doi: 10.1111/j.1442-9993.2007.01730.x
Tsuyuzaki, S., and C. Miyoshi. 2009. Effects of smoke, heat, darkness and cold stratification on seed germination of 40 species in a cool temperate zone in northern Japan. Plant Biology 11: 369–378. doi: 10.1111/j.1438-8677.2008.00136.x
USDA NRCS [US Department of Agriculture Natural Resources Conservation Service]. 2012. The PLANTS Database. <http://plants.usda.gov>. Accessed 3 October 2012.
van Etten, E.J.B., B. Neasham, and S. Dalgleish. 2014. Soil seed banks of fringing salt lake vegetation in arid Western Australia—density, composition and implications for postmine restoration using topsoil. Ecological Management and Restoration 15: 239–242. doi: 10.1111/emr.12119
van Staden, J., N.A.C. Brown, A.K. Jäger, and T.A. Johnson. 2000. Smoke as a germination cue. Plant Species Biology 15: 167–178. doi: 10.1046/j.1442-1984.2000.00037.x
van Staden, J., A.K. Jäger, M.E. Light, and B.V. Burger. 2004. Isolation of the major germination cue from plant-derived smoke. South African Journal of Botany 70: 654–659. doi: 10.1016/S0254-6299(15)30206-4
Wright, H.A., and A.W. Bailey. 1982. Fire ecology, United States and southern Canada. John Wiley & Sons Inc., New York, New York, USA.
Acknowledgements
We gratefully acknowledge the editor and anonymous reviewers for their comments on the manuscript. This is manuscript T-91236 of the College of Agriculture and Natural Resources, Texas Tech University.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made.
The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
To view a copy of this licence, visit https://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Cox, R.D., Chou, YF. & Wester, D.B. Smoke Water and Heat Influence Emergence of Shortgrass Prairie Species. fire ecol 13, 138–148 (2017). https://doi.org/10.4996/fireecology.1301138
Published:
Issue Date:
DOI: https://doi.org/10.4996/fireecology.1301138