
Abstract
Seeding of native grasses is widely used to restore plant communities and prevent establishment of introduced species following wildfire and prescribed burns. However, there is a lack of long-term data to evaluate the success of native grass seeding. Here, in the interior valley shrublands of southwestern Oregon, we resurveyed plots that had been masticated and burned, and then seeded with bunchgrasses seven years previously. The prescribed fires had resulted in bare ground that increased opportunities for bunchgrass germination as well as for invasion by introduced plants. After two years, native grass seeding was successful, with increased bunchgrass cover that correlated with decreased cover of introduced species. However, five years later, bunchgrass cover had declined by 80%, and the frequency of plots with bunchgrasses had declined by 60%. Cover of surviving bunchgrasses in year 7 correlated positively with bunch-grass cover in year 2 (R2 = 0.34; P = 0.003). Seven years after prescribed fire and seeding, native cover, introduced cover, and species richness were unchanged, and bunchgrass persistence was minimal. Basically, seeding following mastication and prescribed burning had a minimal effect. This study highlights the importance of longer-term monitoring to determine the efficacy of seeding treatments.
Resumen
La siembra de pastos nativos es ampliamente usada, luego de incendios naturales o de haber realizado quemas prescritas, para restaurar comunidades vegetales y prevenir el restablecimiento de especies introducidas. Sin embargo, la falta de datos a largo plazo impide poder evaluar el éxito de estas siembras. Nosotros re-evaluamos, en el valle de arbustales del interior en el sudoeste de Oregon, EEUU, parcelas que habían sido objeto, siete años antes, de tratamientos de triturado y quemado, y posteriormente sembradas con pastos. Las quemas prescritas habían dejado el suelo desnudo, lo que incrementó las oportunidades de germinación de pastos y también la invasión de especies introducidas. Después de dos años, la siembra de pastos fue exitosa, con un incremento en su cobertura que se cor relacionó con un decrecimiento en la cobertura de las especies introducidas. Sin embargo, cinco años después, la cobertura de pastos había declinado en un 80 %, y la frecuencia de parcelas con pastos lo habían hecho en un 60 %. La cobertura de los pastos sobrevivientes se correlacionó positivamente con la cobertura en el año 2 (R2 = 0.34; P = 0.003). Después de siete años de las quemas prescriptas y el sembrado, la cobertura de especies nativas, introducidas, y la riqueza de especies permaneció invariable, y la persistencia de pastos fue mínima. Básicamente, la siembra de pastos luego de los tratamientos de triturado y de quemas tuvo un efecto mínimo. Este estudio subraya la importancia del monitoreo a largo plazo para determinar la eficacia de los tratamientos de siembra.
Similar content being viewed by others

Introduction
Seeding of native grasses is widely used to restore particular species, plant communities, and ecosystem services following wildfire and prescribed burns (e.g., Hardegree et al. 2011). However, there is a lack of long-term data to evaluate the success of native grass establishment and its effect in preventing introduced species invasions (Robichaud et al. 2009, Peppin et al. 2010, Endress et al. 2012). Such studies are needed to evaluate the benefits of restoration projects and to improve restoration techniques (Duren and Muir 2010, Hardegree et al. 2011, Peppin et al. 2011).
Invasion by introduced species is a threat to the conservation of shrublands and creates a challenge to restoration (Hosten et al. 2006, Endress et al. 2012). Both mastication and fire lead to more introduced species in part because of increased bare ground (Beyers 2004, Potts and Stephens 2009). At a site with a Mediterranean climate (Hierro et al. 2006), prescribed fire created a favorable environment for invasion by yellow star-thistle (Centaurea solstitialis L.). However, in a ponderosa pine (Pinus ponderosa Lawson and C. Lawson) forest, the combination of mastication plus burning resulted in greater native plant diversity with only a slight increase in introduced species (Kane et al. 2010).
Seeding with native plants may be a way to prevent the spread of introduced species, but a review of post-fire seeding studies found equivalent numbers of effective and ineffective results on introduced species (Peppin et al. 2010). In a grassland-savannah ecosystem, post-fire seeding with native species reduced establishment of introduced species and increased native plant cover and species richness; however, the benefits of seeding occurred only at intermediate levels of introduced species richness (Suding and Gross 2006).
In dense, fire-suppressed shrublands that had been masticated, Coulter et al. (2010) seeded native bunchgrass species following prescribed burns. Post-fire seeding and subsequent increased bunchgrass cover correlated with decreased introduced species. Bunchgrasses were not present on unseeded plots. Initially, the native grass seeding appeared to be successful with 10 % cover of seeded species after two growing seasons. After five more years, we resurveyed their plots to determine the longer-term success of native grass seeding treatments.
Our objective was to assess the longer-term effectiveness of seeding with native bunchgrasses following brush mastication and prescribed fire in the shrublands of an interior valley in southern Oregon, USA. We compared the cover of bunchgrasses and introduced species on masticated plots resurveyed seven years after the prescribed burn with the cover two years after the prescribed burn. We hypothesized that (1) bunchgrasses that had survived two growing seasons would persist and expand; (2) greater bunchgrass cover in year 2 would predict cover in year 7; and (3) introduced species cover would be lower on plots with greater bunchgrass cover. In addition, we asked whether the treatments of prescribed fire and bunchgrass seeding would change the plant community.
Methods
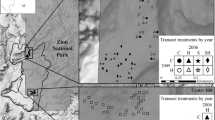
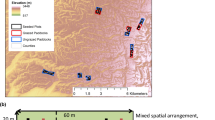
We resurveyed the sites used by Coulter et al. (2010). Sites were on south facing slopes in the Applegate Valley near Ruch, Oregon, USA: China Gulch (42° 15′ N, 122° 3′ W) and Hukill Hollow (42° 11′ N, 122° 59′ W). The climate is a Mediterranean type with cool, wet winters and hot, dry summers: mean temperature 4 °C in January and 21 °C in July. For the years 2005 to 2012, annual rainfall ranged from 343 mm to 734 mm, with only 10 % of total precipitation in the summer months, June through September (www.wrcc.dri.edu/cgi-bin/cli-MONtpre.pl?or7391, accessed 20 Oct 2014). Summer rainfall varied from 0.5 cm to 7 cm during the interval between surveys (Figure 1).

Summer rainfall (June through September) at the Ruch, Oregon, USA, gauging station. Between the second growing season (2007) and the time of resurvey (2012), there was an exceedingly dry summer (2008).
Prior to mastication, the sites were dominated by buckbrush (Ceanothus cuneatus [Hook.] Nutt.), sticky whiteleaf manzanita (Arctostaphylos viscida Parry), Pacific poison oak (Toxicodendron diversilobum [Torr. & A. Gray] Greene), and California yerba santa (Eriodictyon californicum [Hook. & Arn.] Torr.). Mastication was employed to reduce the threat of high-severity wildfire by lowering fuels from standing vegetation to ground level (Kane et al. 2009, Southworth et al. 2011, Duren et al. 2012). Five years after mastication, the sites were treated with prescribed fire to reduce woody fuels on the ground.
At each of two sites, China Gulch (CG) masticated in 2001 and Hukill Hollow (HH) masticated in 2002, 30 1 m2 plots were installed randomly and the vegetation surveyed (Coulter et al. 2010). All plots (60 total) were treated with a prescribed burn in October 2005, year 0. Two weeks after the burn, 15 plots at each site (30 total) were seeded with four native bunchgrasses: Lemmon’s needlegrass (Achnatherum lemmonii [Vasey] Barkworth, syn. Stipa lemmonii [Vasey] Scribn.), California brome (Bromus carinatus Hook. & Arn.), blue wildrye (Elymus glaucus Buckley), and Roemer’s fescue (Festuca roemeri [Pavlick] S. Aiken var. klamathensis B.L. Wilson, syn. F. idahoensis Elmer sp. roemeri [Pavlick] S. Aiken). An equal number of paired control plots (30 total) were unseeded. Cover of all species was assessed in the first growing season, year 1 (2006), and in year 2 (2007). In June 2012, year 7, we resurveyed the seeded (n = 30) and unseeded (n = 30) plots. We estimated percent cover of plant species using midpoint values of FIREMON classes (0 % to 1 %, >1 % to 5 %, >5 % to 15 %, >15 % to 25 %, >25 % to 35 %, >35 % to 45 %, >45 % to 55 %, >55 % to 65 %, >65 % to 7 %, >75 % to 85 %, >85 % to 95 %, and >95 % to 100 %) (http://frames.nbii.gov/projects/firemon/FIREMON_SamplingMethods.pdf).
We used a two sample t-test in MINITAB v. 15 (Minitab Inc., State College, Pennsylvania, USA) with a significance threshold of α = 0.05 to compare bunchgrass cover in seeded and unseeded plots at replicate sites and to evaluate the change in species richness and percent cover from year 2 to year 7. We analyzed changes in percent cover of species within the same plot using paired t-tests, and changes in the frequency of plots with bunchgrasses in different years using χ 2. We performed a linear regression to compare the relationship between bunchgrass and introduced species cover.
To determine differences between the plant communities of seeded and unseeded plots, we compared seeded and unseeded groups by multi-response permutation procedures (MRPP) in PC-ORD v. 6 using Sørensen (Bray-Curtis) distance measures (McCune and Grace 2002, Peck 2010). We calculated A, the chance-corrected within-group agreement, and the P-value. We analyzed the two sites separately because they differed in total number of species.
Results
Total plant alpha-diversity was 58 species at two sites (Appendix 1). Of these, 31 % were introduced species; 22 % were grasses. Beyond the seeded grasses, only one other grass species was native. Four shrub species—buckbrush, California yerba santa, pink honeysuckle (Lonicera hispidula [Lindl.] Douglas ex Torr. & A. Gray), and Pacific poison oak—were present in year 7.
On unseeded plots, bunchgrass cover was 0.4 %. On seeded plots, bunchgrass cover was 1.9 % in year 7, a decline in cover from 9.7 % in year 2 (t = 3.01; P = 0.005; Table 1). California brome and blue wildrye declined from year 2 to year 7 with cover losses of 91 % to 96 %, while cover of Lemmon’s needlegrass and Roemer’s fescue remained the same. All four seeded bunchgrass species remained at low cover levels (0.1 % to 0.9 %) and each was found on at least one plot.
Since bunchgrasses had not germinated in many plots, we looked closely at the set of 23 plots (21 seeded, 2 unseeded) in which bunchgrasses had been present by year 2 (Table 2). The total number of plots with bunchgrasses declined from 23 to 9 (7 seeded, 2 unseeded) in year 7 (χ 2 = 7.2, P = 0.007). Between year 2 and year 7, cover declined in California brome (t = 4.10, P = 0.001) and in blue wildrye (t = 3.08, P = 0.006) but did not change in Roemer’s fescue (t = −0.08, P = 0.94) or in Lemmon’s needlegrass (t = 3.08, P = 0.50) (Table 2). Bunchgrasses with higher cover by year 2 persisted into year 7 better than those with less cover (R2 = 0.34; P = 0.003; Figure 2).

Correlation between bunchgrass cover in year 7 with that in year 2 on plots with bunchgrass survival, at China Gulch and Hukill Hollow in southern Oregon, USA.
Introduced plant cover increased in both seeded and unseeded plots over the seven-year period (Table 3). However, changes in introduced cover between seeded and unseeded plots did not differ (P = 0.68). Introduced species richness increased in both seeded and unseeded plots. Species richness increased by an average of two species. There was no difference between seeded and unseeded plots in the percent increase of introduced species richness (P = 0.73). In plots with bunchgrasses present in year 7, introduced cover had not changed from pre-burn cover levels although there had been a substantial increase in year 2 (Figure 3; R2 = 0.05, P = 0.61).

Cover of introduced species in years 0, 1, 2, and 7 on all seeded (solid line) and unseeded (dashed line) plots at China Gulch and Hukill Hollow in southern Oregon, USA. Seeded and unseeded treatments did not differ.
Native plant cover decreased in both seeded and unseeded plots over the seven-year period (Table 3). Changes in native cover did not differ between seeded and unseeded plots (P = 0.46). Native species richness increased in both seeded and unseeded plots with species richness increasing by an average of one species. Seeded and unseeded plots did not differ in percent increase of native species richness (P = 0.81).
In year 7, plant communities on seeded and unseeded plots did not differ at either CG (A = −0.004, P = 0.62) or at HH (A = 0.0126, P = 0.08). This was consistent with the lack of differences in introduced or native species between seeded and unseeded plots.
Discussion
Hypothesis 1, that bunchgrasses that had survived two growing seasons would persist and expand, was not supported. Seeded bunchgrasses that were present two years following prescribed fire and seeding persisted poorly after seven years, with low bunchgrass cover occurring on only a few plots. Although bunchgrass cover in year 2 was nearly 10 % following fall burns of brush-masticated shrublands, bunchgrass cover declined to less than 2 % in year 7. While all four species of bunchgrasses remained on at least one plot, all cover estimates were low. Relative survival of the four seeded bunchgrass species differed. Lemmon’s needlegrass and Roemer’s fescue persisted with low cover while California brome and blue wildrye attained higher cover in year 2 and then declined by year 7.
Several factors may have prevented successful establishment of seeded bunchgrasses. One, our sites were on southerly aspects with a high concomitant heat load. In California, planted grasses established better on north facing than on south facing slopes (Lulow et al. 2007). Two, rainfall at our sites varied annually over the seven-year course of the study and included one exceedingly dry year (2008), which may have created conditions in which only well-established plants survived. Three, because California brome and blue wildrye are short-lived perennials in contrast to longer lived Roemer’s fescue and Lemmon’s needlegrass (http://plants.usda.gov), they may have reached the ends of their lifespans and failed to reseed due to site conditions or the drought year.
Supporting hypothesis 2, bunchgrass cover in year 7 correlated positively with cover in year 2. Greater cover may indicate better root development, better competitive ability, and consequently longer persistence. In other studies, larger bunchgrass transplants showed better survival and had greater final mass (Wilson 1994, Page and Bork 2005).
Hypothesis 3, that introduced species cover would be lower on plots with greater bunchgrass cover, was not supported. Seeding with bunchgrasses had no effect on the cover or species richness of introduced or native plant species. Although the cover of native and introduced species had decreased slightly and species richness had increased slightly after two years, after seven years neither cover nor species richness of native and introduced species differed between seeded and unseeded plots. Introduced species cover showed no correlation with bunchgrass cover. Similarly, neither richness nor cover of native species was affected by seeding with bunchgrasses. While native grass seeding limited the invasion of introduced grasses in a grassland-savanna ecosystem (Endress et al. 2012), a review of post-fire seeding studies found equivalent numbers of effective and ineffective results of seeding on introduced species (Peppin et al. 2010).
Treatments of prescribed fire and bunchgrass seeding did not change the plant community, due to the stability of both native and introduced components of vegetation. Apparently, species adapted to seasonally dry habitats through characteristics such as below-ground perennating structures and seed banks persist through prescribed fire.
These findings show the importance of longer-term monitoring of restoration treatments to determine their success (Robichaud et al. 2009, Peppin et al. 2010, Endress et al. 2012). Initial results at our study sites indicated a successful introduction of native bunchgrasses, but this introduction was considerably less successful after seven years. The outcome of seeding may depend on habitat conditions and on climatic variability. Although the trend has been to seed with native species on public lands to meet natural ecological system objectives, the success of such treatments remains uncertain (Peppin et al. 2011).
Climate change is likely to interfere with attempts at restoration of native plant communities, particularly at lower elevations and on south facing slopes with little summer rainfall (Harris et al. 2006, Bachelet et al. 2011). The shrubland communities of inland valleys may be adapted to Mediterranean-type climates, and thus will survive and expand. However, our ability to manage them or to promote native species restoration may be more limited. If reseeding is not effective, preservation of habitat becomes even more important.
Literature Cited
Bachelet, D., B.R. Johnson, S.D. Bridgham, P.V. Dunn, H.E. Anderson, and B.M. Rogers. 2011. Climate change impacts on western Pacific Northwest prairies and savannas. Northwest Science 85: 411–429. doi: 10.3955/046.085.0224
Beyers, J.L. 2004. Postfire seeding for erosion control: effectiveness and impacts on native plant communities. Conservation Biology 18: 947–956. doi: 10.1111/j.1523-1739.2004.00523.x
Coulter, C.T., P.E. Hosten, and D. Southworth. 2010. Prescribed fire and post-fire seeding in brush masticated oak-chaparral: consequences for native and non-native plants. Fire Ecology 6(2): 60–75. doi: 10.4996/fireecology.0602060
Duren, O.C., and P.S. Muir. 2010. Does fuels management accomplish restoration in southwest Oregon, USA, chaparral? Insights from age structure. Fire Ecology 6(2): 76–96. doi: 10.4996/fireecology.0602076
Duren, O.C., P.S. Muir, and P.E. Hosten. 2012. Vegetation change from the Euro-American settlement era to the present in relation to environment and disturbance in southwest Oregon. Northwest Science 86: 310–328. doi: 10.3955/046.086.0407
Endress, B.A., C.G. Parks, B.J. Naylor, S.R. Radosevich, and M. Porter. 2012. Grassland response to herbicides and seeding of native grasses 6 years posttreatment. Invasive Plant Science and Management 5: 311–316. doi: 10.1614/IPSM-D-11-00050.1
Hardegree, S.P., T.A. Jones, B.A. Roundy, M.L. Shaw, and T.A. Monaco. 2011. Assessment of range planting as a conservation practice. Pages 171–212 in: D.D. Briske, editor. Conservation benefits of rangeland practices: assessment, recommendations, and knowledge gaps. Allen Press, Lawrence, Kansas, USA.
Harris, J.A., R.J. Hobbs, E. Higgs, and J. Aronson. 2006. Ecological restoration and global climate change. Restoration Ecology 14: 170–176. doi: 10.1111/j.1526-100X.2006.00136.x
Hierro, J.L., D. Villarreal, O. Eren, J.M. Graham, and R.M. Callaway. 2006. Disturbance facilitates invasion: the effects are stronger abroad than at home. American Naturalist 168: 144–156. doi: 10.1086/505767
Hosten, P.E., G. Hickman, F. Lake, F. Lang, and D. Vesely. 2006. Oak woodland and savanna restoration. Pages 63–96 in: D. Apostol and M. Sinclair, editors. Restoring the Pacific Northwest: the art and science of ecological restoration in Cascadia. Island Press, Washington, D.C., USA.
Kane, J.M., J.M. Varner, and E.E. Knapp. 2009. Novel fuelbed characteristics associated with mechanical mastication treatments in northern California and southwestern Oregon. International Journal of Wildland Fire 18: 686–697. doi: 10.1071/WF08072
Kane, J.M., J.M. Varner, E.E. Knapp, and R.F. Powers. 2010. Understory vegetation response to mechanical mastication and other fuels treatments in a ponderosa pine forest. Applied Vegetation Science 13: 207–220. doi: 10.1111/j.1654-109X.2009.01062.x
Lulow, M.E., T.P. Young, J.L. Wirka, and J.H. Anderson. 2007. Variation in the initial success of seeded native bunchgrasses in the rangeland foothills of Yolo County, California. Ecological Restoration 25: 20–28. doi: 10.3368/er.25.1.20
McCune, B., and J.B. Grace. 2002. Analysis of ecological communities. MjM Software Design, Gleneden Beach, Oregon, USA.
Page, H.N., and E.W. Bork. 2005. Effect of planting season, bunchgrass species, and neighbor control on the success of transplants for grassland restoration. Restoration Ecology 13: 651–685. doi: 10.1111/j.1526-100X.2005.00083.x
Peck, J.E. 2010. Multivariate analysis for community ecologists: step-by-step using PC-ORD. MjM Software Design, Gleneden Beach, Oregon, USA.
Peppin, D.L., P.Z. Fulé, C.H. Sieg, J.L. Beyers, and M.E. Hunter. 2010. Post-wildfire seeding in forest of the western United States: an evidence-based review. Forest Ecology and Management 260: 573–586. doi: 10.1016/j.foreco.2010.06.004
Peppin, D.L., P.Z. Fulé, C.H. Sieg, J.L. Beyers, M.E. Hunter, and P. Robichaud. 2011. Recent trends in post-wildfire seeding in western US forests: costs and seed mixes. International Journal of Wildland Fire 20: 702–708. doi: 10.1071/WF10044
Potts, J.B., and S.L. Stephens. 2009. Invasive and native plant responses to shrubland fuel reduction: comparing prescribed fire, mastication, and treatment season. Biological Conservation 142: 1657–1664. doi: 10.1016/j.biocon.2009.03.001
Robichaud, P.R., S.A. Lewis, R.E. Brown, and L.E. Ashmun. 2009. Emergency post-fire rehabilitation treatment effects on burned area ecology and long-term restoration. Fire Ecology 5(1): 115–128. doi: 10.4996/fireecology.0501115
Southworth, D., J. Donohue, J.L. Frank, and J. Gibson. 2011. Mechanical mastication and prescribed fire in conifer-hardwood chaparral: differing responses of ectomycorrhizae and truffles. International Journal of Wildland Fire 20: 888–896. doi: 10.1071/WF10033
Suding, K.N., and K.L. Gross. 2006. Modifying native and non-native species richness correlations: the influence of fire and seed addition. Ecological Applications 16: 1319–1326. doi: 10.1890/1051-0761(2006)016[1319:MNAESR]2.0.CO;2
Wilson, S.D. 1994. Initial size and the competitive responses of two grasses at two levels of soil nitrogen: a field experiment. Canadian Journal of Botany 72: 1349–1354. doi: 10.1139/b94-165
Acknowledgments
We thank C.T. Coulter for help in relocating plots and in standardizing data collection protocols, D. Kendig for project support, and S. Joachims for field assistance. Research was supported in part by the US Department of Interior, Bureau of Land Management, Medford District. The views and conclusions contained in this document are those of the authors and should not be interpreted as representing the opinions or policies of the US government. Mention of trade names or commercial products does not constitute their endorsement by the US government.
Author information
Authors and Affiliations
Corresponding author
Appendix 1
Appendix 1
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made.
The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
To view a copy of this licence, visit https://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Busby, L.M., Southworth, D. Minimal Persistence of Native Bunchgrasses Seven Years after Seeding following Mastication and Prescribed Fire in Southwestern Oregon, USA. fire ecol 10, 63–71 (2014). https://doi.org/10.4996/fireecology.1003063
Published:
Issue Date:
DOI: https://doi.org/10.4996/fireecology.1003063