
Abstract
Wenxinia marina Ying et al. 2007 is the type species of the genus Wenxinia, a representative of the Roseobacter group within the alphaproteobacterial family Rhodobacteraceae, isolated from oilfield sediments of the South China Sea. This family was shown to harbor the most abundant bacteria especially from coastal and polar waters, but was also found in microbial mats, sediments and attached to different kind of surfaces.
Here we describe the features of W. marina strain HY34T together with the genome sequence and annotation of strain DSM 24838T and novel aspects of its phenotype. The 4,181,754 bp containing genome sequence encodes 4,047 protein-coding genes and 59 RNA genes. The genome of W. marina DSM 24838T was sequenced as part of the activities of the Genomic Encyclopedia of Type Strains, Phase I: the one thousand microbial genomes (KMG) project funded by the DoE and the Transregional Collaborative Research Centre 51 (TRR51) funded by the German Research Foundation (DFG).
Similar content being viewed by others

Introduction
Strain HY34T (= DSM 24838T = CGMCC 1.6105T = JCM 14017T) is the type strain of Wenxinia marina in the monospecific genus Wenxinia [1,2], which belongs to the widely distributed marine Roseobacter group [3]. The strain was isolated from sediments of the Xijiang oilfield located in the South China Sea (China) [1]. The genus Wenxinia was named after Professor Wen-Xin Chen, a Chinese pioneer in soil microbiology. The species epithet marina refers to the Latin adjective marina (‘of or belonging to the sea’) [1,2]. Current PubMed records do not indicate any follow-up research with strain HY34T after the initial description of W. marina [1].
In this study we analyzed the genome sequence of W. marina DSM 24838T. We present a description of the genome sequencing and annotation and present a summary classification together with a set of features for strain HY34T, including novel aspects of its phenotype.
Classifications and features
16S rRNA gene analysis
Figure 1 shows the phylogenetic neighborhood of W. marina in a 16S rRNA gene based tree. The sequences of the two identical 16S rRNA gene copies in the genome, differ by three nucleotides from the previously published 16S rRNA gene sequence (DQ640643).

Phylogenetic tree highlighting the position of W. marina relative to the type strains of the neighboring genera Citreicella and Rubellimicrobium. The tree was inferred from 1,381 aligned characters of the 16S rRNA gene sequence under the maximum likelihood (ML) criterion as previously described [4]. The branches are scaled in terms of the expected number of substitutions per site. Numbers adjacent to the branches are support values from 1,000 ML bootstrap replicates (left) and from 1,000 maximum-parsimony bootstrap replicates (right) if larger than 60% [4]. Lineages with type strain genome sequencing projects registered in GOLD [5] are labeled with one asterisk, those also listed as ‘Complete and Published’ with two asterisks [6].
A representative genomic 16S rRNA gene sequence of W. marina DSM 24838T was compared with the Greengenes database for determining the weighted relative frequencies of taxa and (truncated) keywords as previously described [4]. The most frequently occurring genera were Ruegeria (41.6%), Paracoccus (31.0%), Oceanicola (14.0%), Silicibacter (5.0%) and Loktanella (3.3%) (60 hits in total). Among all other species, the one yielding the highest score was Oceanicola granulosus (AAOT01000021), which corresponded to an identity of 94.7% and an HSP coverage of 99.6%. (Note that the Greengenes database uses the INSDC (= EMBL/NCBI/DDBJ) annotation, which is not an authoritative source for nomenclature or classification.) The highest-scoring environmental sequence was DQ640643 (Greengenes short name ‘Rhodobacteraceae South China Sea oil field sediment isolate HY34 Rhodobacteraceae str. HY34’), which showed an identity of 99.8% and an HSP coverage of 100.0%. The most frequently occurring keywords within the labels of all environmental samples that yielded hits were ‘microbi’ (4.3%), ‘coral’ (3.6%), ‘sea’ (2.6%), ‘diseas’ (2.5%) and ‘china’ (2.4%) (190 hits in total). The most frequently occurring keywords within the labels of those environmental samples which yielded hits of a higher score than the highest scoring species were ‘antecubit, fossa, skin’ (13.9%) and ‘china, field, oil, rhodobacteracea, sea, sediment, south’ (8.3%) (3 hits in total). Some of these keywords fit well to the isolation site of strain HY34T [1].
Morphology and physiology
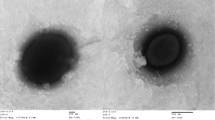
Cells of strain HY34T form Gram-negative, ovoid or short rods (0.7–0.8 µm in width and 1.3 µm in length) [Figure 2]. Motility and sporulation were not observed. Cells are strictly aerobic and display a heterotrophic lifestyle. When cultured on Marine Agar 2216 colonies with a weak pink color became visible, but bacteriochlorophyll a was not detected. The strain grows in a temperature range of 15–42°C with an optimum at 34–38°C. NaCl is required for growth (0.5–9%) with an optimum salt concentration at 1–4%. Further, the strain grows in a range of pH 6.5–8.5 with an optimum pH of 7.5–8.0. The strain is oxidase- and catalase-positive. Nitrate is reduced to nitrite. Indole and H2S are not produced. Cells hydrolyze urea and Tween 20, and a weak hydrolysis of Tween 40 and Tween 80 was also detected. The strain does not hydrolyze agar, casein, starch, DNA or CM-cellulose. Strain HY34T accumulates polyhydroxyalkanoates in its cells. Tests for arginine dehydrolase and lecithinase were negative. Further, cells utilize sucrose, lactose, galactose, maltose, melezitose, L-rhamnose, L-fucose, trehalose, cellobiose, gluconate, lactic acid, malate, L-glutamic acid. The strain utilizes D-melibiose, inulin, methyl α-D-glucoside, glycerol, sorbitol, butanol, pyruvate, formic acid, L-alanine and L-proline weakly. Utilization of D-raffinose, mannitol, L-sorbose, dulcitol, adonitol, myo-inositol, methanol, ethanol, citrate, malonate, butyric acid and caprate acid was not detected. Cells produce acid from D-xylose, cellobiose, lactose, L-rhamnose, L-arabinose, D-raffinose, and weakly from sucrose, maltose, mannose, trehalose, and ribose. Strong activities for esterase (C8) and α- and β-glucosidases were detected, as well as weak activities for alkaline phosphatase, leucine arylamidase, valine arylamidase and naphthol-AS-BI-phosphohydrolase. No activity was found for acid phosphatase, N-acetyl-β-cysteine arylamidase, glucosamidase, α- and β-galactosidase, α-mannosidase, α-chrymotrypsin, β-glucuronidase, α-fucosidase and lipase (C14). Cells of strain HY34T are found to be resistant to norfloxacin, tetracycline and gentamicin as well as sensitive to neomycin, polymyxin B, streptomycin, ampicillin, carbenicillin, vancomycin, ciprofloxacin, rifampicin, chloramphenicol, benzylpenicillin, kanamycin, and erythromycin (all data from [1] and presented in Table 1).

Phase contrast micrograph of W. marina DSM 24838T.
The utilization of carbon compounds by W. marina DSM 24838T grown at 28°C was also determined for this study using Generation-III microplates in an OmniLog phenotyping device (BIOLOG Inc., Hayward, CA, USA) [7]. The microplates were inoculated with a cell suspension at a cell density of 95–96% turbidity and dye IF-A. Further additives were vitamin, micronutrient and sea-salt solutions, which had to be added for dealing with such marine bacteria [8]. The plates were sealed with parafilm to avoid a loss of fluid.
The exported measurement data were further analyzed with the opm package for R [9,10], using its facilities for statistically estimating parameters from the respiration curves such as the maximum height, and automatically translating these values into negative, ambiguous, and positive reactions. The reactions were recorded in three biological replicates.
On the Generation-III plates, the strain was positive for pH 6, 1% NaCl, 4% NaCl, 8% NaCl, D-galactose, 3-O-methyl-D-glucose, D-fucose, L-fucose, L-rhamnose, 1% sodium lactate, myoinositol, rifamycin SV, L-aspartic acid, L-glutamic acid, L-histidine, L-serine, D-glucuronic acid, quinic acid, L-lactic acid, citric acid, α-keto-glutaric acid, D-malic acid, L-malic acid, nalidixic acid, and sodium formate.
W. marina HY34T was negative for the following tests: dextrin, D-maltose, D-trehalose, D-cellobiose, β-gentiobiose, sucrose, D-turanose, stachyose, pH 5, D-raffinose, α-D-lactose, D-melibiose, β-methyl-D-galactoside, D-salicin, N-acetyl-D-glucosamine, N-acetyl-β-D-mannosamine, N-acetyl-D-galactosamine, N-acetyl-neuraminic acid, D-glucose, D-mannose, D-fructose, inosine, fusidic acid, D-serine, D-sorbitol, D-mannitol, D-arabitol, glycerol, D-glucose-6-phosphate, D-fructose-6-phosphate, D-aspartic acid, D-serine, troleandomycin, minocycline, gelatin, glycyl-L-proline, L-alanine, L-arginine, L-pyroglutamic acid, lincomycin, guanidine hydrochloride, niaproof, pectin, D-galacturonic acid, L-galactonic acid-γ-lactone, D-gluconic acid, glucuronamide, mucic acid, D-saccharic acid, vancomycin, tetrazolium violet, tetrazolium blue, phydroxy-phenylacetic acid, methyl pyruvate, D-lactic acid methyl ester, bromo-succinic acid, lithium chloride, potassium tellurite, tween 40, γ-amino-n-butyric acid, α-hydroxy-butyric acid, β-hydroxy-butyric acid, α-keto-butyric acid, acetoacetic acid, propionic acid, acetic acid, aztreonam, butyric acid and sodium bromate.
The phenotype microarray results fit to those reported by Ying and colleagues [1] in large part. Only the utilization of lactose and D-trehalose could not be confirmed by respiration measurements under the given conditions. Interestingly, W. marina DSM 24838T showed a varying phenotype both in growth measurement [1] and in the respiration curves among replicates. Ying and colleagues reported eleven substrates yielding “weak” results, which complicates the exact comparison of substrate utilization [1]. In contrast to Ying and colleagues, the OmniLog measurements gave positive reactions for L-histidine and myoinositol. This may be due respiratory measurements being more sensitive than growth measurements [11].
Chemotaxonomy
The principal cellular fatty acids of strain HY34T are C18:1 ω7c (57.1%), C16:0 (16.5%), 11-methyl C18:1 ω7c (5.4%), C18:0 (3.9%), C14:0 (3.7%), C15:1 iso G and C15:1 iso I (3.4%), summed feature 3 C16:1 ω7c and/or C15:0 2-OH (1.9%), C12:0 (1.6%) and C13:0 2-OH (1.2%). The major respiratory lipoquinone was ubiquinone 10, which is a well-known characteristic of the Alphaproteobacteria. Phosphatidylglycerol and phosphatidylcholine were identified as the major polar lipids. In contrast to other representatives of the Roseobacter group such as Marinovum algicola FF3T (DSM 10251T) [12,13], strain HY34T also contains an unidentified glycolipid called L1, which shows similarities to an unidentified phospholipid of Ruegeria atlantica DSM 5828T (all data from [1]).
Genome sequencing and annotation
Genome project history
This strain was twice selected for genome sequencing on the basis of its phylogenetic position [14]. First as part of the DFG funded project “Ecology, Physiology and Molecular Biology of the Roseobacter clade: Towards a Systems Biology Understanding of a Globally Important Clade of Marine Bacteria” and later as part of the “Genomic Encyclopedia of Type Strains, Phase I: the one thousand microbial genomes (KMG) project” [15], a follow-up of the GEBA project [16], which aims in increasing the sequencing coverage of key reference microbial genomes. The strain was independently sequenced from the same source of DNA and produced draft sequences that were finally joined. The project information can found in the Genomes OnLine Database [5] and the Integrated Microbial Genomes database (IMG) [17]. A summary of the project information is shown in Table 2.
Growth conditions and DNA isolation
A culture of W. marina DSM 24838T was grown aerobically in DSMZ medium 514 [18] at 30°C. Genomic DNA was isolated using Jetflex Genomic DNA Purification Kit (GENOMED 600100) following the standard protocol provided by the manufacturer but modified by an incubation time of 60 min, incubation on ice overnight on a shaker, the use of an additional 50 µl proteinase K, and the addition of 100 µl protein precipitation buffer. The DNA is available from the Leibniz-Institute DSMZ through the DNA Bank Network [19].
Genome sequencing and assembly
The genome sequencing under the DFG funded part of the project was perform as previously described for Rubellimicrobium thermophilum [6], with 3.3 million reads delivered by the first run on an Illumina GAII platform. To increase the sequencing depth, a second Ilumina run was performed, providing another 8.1 million reads. The first draft assembly from 9,139,639 filtered reads (median read length 122 nt) resulted in more than 300 contigs. To gain information on the contig arrangement an additional 454 run was performed. The paired-end pyrosequencing jumping library resulted in 158,608 reads, with an average read length of 450 bp. Both draft assemblies (Illumina and 454 sequences) were fractionated into artificial Sanger reads of 1,000 nt in length plus 75 bp overlap on each site. These artificial reads served as an input for the phred/phrap/consed package [20]. In combination the assembly resulted in 265 contigs in 26 scaffolds.
The genome sequencing under the DoE funded part of the project was performed as previously described for Halomonas zhanjiangensis [21] also using the Illumina technology [22]. An Illumina Standard shotgun library was constructed and sequenced using the Illumina HiSeq 2000 platform. All general aspects of library construction and sequencing performed at the JGI can be found at [23]. The final assembly for this part of the project resulted in 41 scaffolds covering 4,175,892 bp (ARAY00000000).
The draft sequence from the first (DFG-funded) part was mapped to the permanent draft version ARAY00000000 using minimus2 [24]. By manual editing the number of contigs was reduced to 22 in 8 scaffolds (AONG00000000). The combined sequences provided a 356 × coverage of the genome.
Genome annotation
Genes were identified using Prodigal [25] as part of the JGI genome annotation pipeline. The predicted CDSs were translated and used to search the National Center for Biotechnology Information (NCBI) nonredundant database, UniProt, TIGR-Fam, Pfam, PRIAM, KEGG, COG, and InterPro databases. Identifications of RNA genes were carried out by using HMMER 3.0rc1 [26] (rRNAs) and tRNAscan-SE 1.23 (tRNAs) [27]. Other non-coding genes were predicted using INFERNAL 1.0.2 [28]. Additional gene prediction analysis and functional annotation was performed within the Integrated Microbial Genomes - Expert Review (IMG-ER) platform [29] CRISPR elements were detected using CRT [30] and PILER-CR [51].
Genome properties
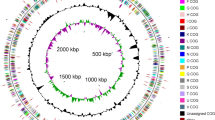
The genome statistics are provided in Table 3 and Figure 3. The genome of DSM 24838T has a total length of 4,181,754 bp and a G+C content of 70.5%. Of the 4,106 genes predicted, 4,047 were protein-coding genes, and 59 RNAs. The majority of the protein-coding genes (80.4%) were assigned a putative function while the remaining ones were annotated as hypothetical proteins. The distribution of genes into COGs functional categories is presented in Table 4.

Graphical map of the largest scaffold. From bottom to the top: Genes on forward strand (colored by COG categories), Genes on reverse strand (colored by COG categories), RNA genes (tRNAs green, rRNAs red, other RNAs black), GC content (black), GC skew (purple/olive).
Insights into the genome
Plasmids
Genome sequencing of W. marina DSM 24838T reveals the presence of one plasmid with a size of about 101 kb. The plasmid contains a characteristic replicase of the RepA-I type [31], but the typical module structure containing the replicase as well as a parAB partitioning operon was not found. A single parA gene (wenxma_04096) is located adjacent to the replicase and an additional parAB operon (wenxma_04090 to wenxma_04091) is located downstream of repA-I. The plasmid harbors neither a plasmid stability module nor a type-IV secretion system.
The plasmid contains a large RTX-toxin (wenxma_04058) and is dominated by genes that are required for polysaccharide biosynthesis. It includes all four genes of the rhamnose pathway [32], but the rmlA gene for the glucose-1-phosphate thymidylyltransferase (EC 2.7.7.24; wenxma_04097) is separated from the remaining clustered genes (rmlC, rmlB, rmlD; wenxma_04094 to wenxma_04092). The extrachromosomal replicon may be involved in surface attachment. Comparable RepA-I type plasmids with a similar genetic composition are also present in other Rhodobacterales including several Phaeobacter strains [33].
Phages
Many bacteria encode genome-inserted gene sequences, which are associated with prophages, one of the major reason for horizontal gene transfer and bacterial diversity [34,35]. The genome sequence of W. marina DSM 24838T was found to encode several prophage-associated gene sequences (e.g., wenxma_00641 to wenxma_00646, wenxma_00930 to wenxma_00936, wenxma_01496 to wenxma_01510).
Quorum Sensing
Analysis of the DSM 24838T genome sequence revealed the presence of gene sequences associated to quorum sensing (QS) [36–38]. QS is a bacterial communication system via chemical signal molecules called autoinducers, which are produced and released by QS bacteria to coordinate behaviors with respect to their population density [38]. Interestingly and surprisingly, QS induces also individual morphologies and cell division modes, which was recently shown for D. shibae DFL-12, another representative of the Roseobacter group [39,40]. Regarding to QS the genome of DSM 24838T codes for, e.g., two N-acyl-L-homoserine-lactone synthetases (LuxI homologues, wenxma_01086 and wenxma_03269) and two genes possibly encoding QS-involved response and transcriptional regulators (LuxR homologues, wenxma_01085 and wenxma_03267).
Morphological traits
With regard to morphological traits, several genes associated with the putative production, biosynthesis and export of exopopolysaccharides (wenxma_00281, wenxma_02363 and wenxma_02364, wenxma_03720 and wenxma_03721) and capsule polysaccharides (wenxma_00822, wenxma_02023 to wenxma_02025, wenxma_02704 and wenxma_02705, wenxma_04069) were detected.
Interestingly, the genome of DSM 24838T was found to encode several gene sequences putatively involved in pili formation (e.g., wenxma_01776 to wenxma_01787, wenxma_03426 to wenxma_03435) and chemotaxis (e.g., wenxma_3823 to wenxma_03830), although the strain was described as non-motile [1]. Hence, it could be that the formed pili play a role for adhesion or switching-type motility on solid surfaces.
Further, according to its genome strain DSM 24838T accumulates polyhydroxyalkanoates as storage compounds (wenxma_02601 to wenxma_02604), which is in accordance with the findings of Ying and colleagues for strain HY34T [1].
References
Ying JY, Wang BJ, Dai X, Yang SS, Liu SJ, Liu ZP. Wenxinia marina gen. nov., a novel member of the Roseobacter clade isolated from oilfield sediments of the South China Sea. Int J Syst Evol Microbiol 2007; 57:1711–1716. PubMed http://dx.doi.org/10.1099/ijs.0.64825-0
Euzéby JP. List of prokaryotic names with standing in nomenclature. http://www.bacterio.cict.fr/s/wenxinia.html.
Buchan A, González JM, Moran MA. Overview of the marine Roseobacter lineage. Appl Environ Microbiol 2005; 71:5665–5677. PubMed http://dx.doi.org/10.1128/AEM.71.10.5665-5677.2005
Göker M, Cleland D, Saunders E, Lapidus A, Nolan M, Lucas S, Hammon N, Deshpande S, Cheng JF, Tapia R, et al. Complete genome sequence of Isosphaera pallida type strain (IS1 BT). Stand Genomic Sci 2011; 4:63–71. PubMed http://dx.doi.org/10.4056/sigs.1533840
Pagani I, Liolios K, Jansson J, Chen IM, Smirnova T, Nosrat B, Markowitz VM, Kyrpides NC. The Genomes OnLine Database (GOLD) v.4: status of genomic and metagenomic projects and their associated metadata. Nucleic Acids Res 2012; 40:D571–D579. PubMed http://dx.doi.org/10.1093/nar/gkr1100
Fiebig A, Riedel T, Gronow S, Petersen J, Klenk HP, Göker M. Genome sequence of the reddish-pigmented Rubellimicrobium thermophilum type strain (DSM 16684T), a member of the Roseobacter clade. Stand Genomic Sci 2013; 8:480–490. PubMed http://dx.doi.org/10.4056/sigs.4247911
Bochner BR. Global phenotypic characterization of bacteria. FEMS Microbiol Rev 2009; 33:191–205. PubMed http://dx.doi.org/10.1111/j.1574-6976.2008.00149.x
Buddruhs N, Pradella S, Göker M, Päuker O, Michael V, Pukall R, Spröer C, Schumann P, Petersen J, Brinkhoff T. Molecular and phenotypic analyses reveal the non-identity of the Phaeobacter gallaeciensis type strain deposits CIP 105210T and DSM 17395. Int J Syst Evol Microbiol 2013; 63:4340–4349. PubMed http://dx.doi.org/10.1099/ijs.0.053900-0
Vaas LA, Sikorski J, Hofner B, Fiebig A, Buddruhs N, Klenk HP, Göker M. opm: an R package for analyzing Omnilog(R) phenotype microarray data. Bioinformatics 2013; 29:1823–1824. Med http://dx.doi.org/10.1093/bioinformatics/btt291
Vaas LA, Sikorski J, Michael V, Göker M, Klenk HP. Visualization and curve-parameter estimation strategies for efficient exploration of phenotype microarray kinetics. PLoS ONE 2012; 7:e34846. PubMed http://dx.doi.org/10.1371/journal.pone.0034846
Vaas LAI, Marheine M, Sikorski J, Göker M, Schumacher M. Impacts of pr-10a overexpression at the molecular and the phenotypic level. Int J Mol Sci 2013; 14:15141–15166; http://dx.doi.org/10.3390/ijms140715141.PubMed
Lafay B, Ruimy R, de Traubenberg CR, Breittmayer V, Gauthier MJ, Christen R. Roseobacter algicola sp. nov., a new marine bacterium isolated from the phycosphere of the toxin-producing dinoflagellate Prorocentrum lima. Int J Syst Bacteriol 1995; 45:290–296. PubMed http://dx.doi.org/10.1099/00207713-45-2-290
Martens T, Heidorn T, Pukall R, Simon M, Tindall BJ, Brinkhoff T. Reclassification of Roseobacter gallaeciensis Ruiz-Ponte et al. 1998 as Phaeobacter gallaeciensis gen. nov., comb. nov., description of Phaeobacter inhibens sp. nov., antibiotic-producing members of the Roseobacter clade, reclassification of Ruegeria algicola (Lafay et al. 1995) Uchino et al. 1998 as Marinovum algicola gen. nov., comb. nov., and emended descriptions of the genera Roseobacter, Ruegeria and Leisingera. Int J Syst Evol Microbiol 2006; 56:1293–1304. PubMed http://dx.doi.org/10.1099/ijs.0.63724-0
Göker M, Klenk HP. Phylogeny-driven target selection for large-scale genome sequencing (and other) projects. Stand Genomic Sci 2013; 8:360–374. PubMed http://dx.doi.org/10.4056/sigs.3446951
Kyrpides NC, Woyke T, Eisen JA, Garrity G, Lilburn TG, Beck BJ, Whitman WB, Hugenholz P, Klenk HP. Genomic Encyclopedia of Type Strains, Phase I: the one thousand microbial genomes (KMG-I) project. Stand Genomic Sci 2013; 9:628–634. http://dx.doi.org/10.4056/sigs.5068949
Wu D, Hugenholtz P, Mavromatis K, Pukall R, Dalin E, Ivanova NN, Kunin V, Goodwin L, Wu M, Tindall BJ, et al. A phylogeny-driven Genomic Encyclopaedia of Bacteria and Archaea. Nature 2009; 462:1056–1060. PubMed http://dx.doi.org/10.1038/nature08656
Markowitz VM, Chen IM, Palaniappan K, Chu K, Szeto E, Grechkin Y, Ratner A, Jacob B, Huang J, Williams P, et al. IMG: the integrated microbial genomes database and comparative analysis system. Nucleic Acids Res 2012; 40:D115–D122. PubMed http://dx.doi.org/10.1093/nar/gkr1044
List of growth media used at the DSMZ: http://www.dmsz.de/catalogues/cataloque-microorganisms/culture-technology/list-of-media-for-microorganisms.html.
Gemeinholzer B, Dröge G, Zetzsche H, Haszprunar G, Klenk HP, Güntsch AM, Berendsohn WG, Wägele JW. The DNA Bank Network: the start from a German initiative. Biopreserv Biobank 2011; 9:51–55. http://dx.doi.org/10.1089/bio.2010.0029
Gordon D, Abajian C, Green P. Consed: a graphical tool for sequence finishing. Genome Res 1998; 8:195–202. Med http://dx.doi.org/10.1101/gr.8.3.195
Zhou Y, Li R, Gao XY, Lapidus A, Han J, Haynes M, Lobos E, Huntemann M, Pati A, Ivanova NN, et al. High quality draft genome sequence of the slightly halophilic bacterium Halomonas zhanjiangensis type strain JSM 078169T (DSM 21076T) from a sea urchin in southern China. Stand Genomic Sci (under review).
Bennett S. Solexa Ltd. Pharmacogenomics 2004; 5:433–438. Med http://dx.doi.org/10.1517/14622416.5.4.433
The DOE Joint Genome Institute. http://www.jgi.doe.gov
Sommer DD, Delcher AL, Salzberg SL, Pop M. Minimus: a fast, lightweight genome assembler. BMC Bioinformatics 2007; 8:64. PubMed http://dx.doi.org/10.1186/1471-2105-8-64
Hyatt D, Chen GL, LoCascio PF, Land ML, Larimer FW, Hauser LJ. Prodigal: prokaryotic gene recognition and translation initiation site identification. BMC Bioinformatics 2010; 11:119. PubMed http://dx.doi.org/10.1186/1471-2105-11-119
Finn DR, Clements J, Eddy SR. HMMER web server: interactive sequence similarity searching. Nucleic Acids Res. Web Server Issue 2011; 39:W29–W37.
Lowe TM, Eddy SR. tRNAscan-SE: A Program for Improved Detection of Transfer RNA Genes in Genomic Sequence. Nucleic Acids Res 1997; 25:955–964. Med http://dx.doi.org/10.1093/nar/25.5.0955
Nawrocki EP, Kolbe DL, Eddy SR. Infernal 1.0: Inference of RNA alignments. Bioinformatics 2009; 25:1335–1337. PubMed http://dx.doi.org/10.1093/bioinformatics/btp_157
Markowitz VM, Ivanova NN, Chen IMA, Chu K, Kyrpides NC. IMG ER: a system for microbial genome annotation expert review and curation. Bioinformatics 2009; 25:2271–2278. PubMed http://dx.doi.org/10.1093/bioinformatics/btp_393
Bland C, Ramsey TL, Sabree F, Lowe M, Brown K, Kyrpides NC, Hugenholtz P. CRISPR recognition tool (CRT): a tool for automatic detection of clustered regularly interspaced palindromic repeats. BMC Bioinformatics 2007; 8:209. Med http://dx.doi.org/10.1186/1471-2105-8-209
Petersen J. Phylogeny and compatibility: plasmid classification in the genomics era. Arch Microbiol 2011; 193:313–321. PubMed
Giraud MF, Naismith JH. The rhamnose pathway. Curr Opin Struct Biol 2000; 10:687–696. PubMed http://dx.doi.org/10.1016/S0959-440X(00)00145-7
Thole S, Kalhoefer D, Voget S, Berger M, Engelhardt T, Liesegang H, Wollherr A, Kjelleberg S, Daniel R, Simon M, et al. Phaeobacter gallaeciensis genomes from globally opposite locations reveal high similarity of adaption to surface life. ISME J 2012; 6:2229–2244. PubMed http://dx.doi.org/10.1038/ismej.2012.62
Canchaya C, Proux C, Fournous G, Bruttin A. Prophage genomics. Microbiol Mol Biol Rev 2003; 67:238–276. PubMed http://dx.doi.org/10.1128/MMBR.67.2.238-276.2003
Paul JH. Prophages in marine bacteria: dangerous molecular bombs or the key to survival in the seas? ISME J 2008; 2:579–589. PubMed http://dx.doi.org/10.1038/ismej.2008.35
Bassler BL. How bacteria talk to each other: regulation of gene expression by quorum sensing. Curr Opin Microbiol 1999; 2:582–587. PubMed http://dx.doi.org/10.1016/S1369-5274(99)00025-9
Henke JM, Bassler BL. Bacterial social engagements. Trends Cell Biol 2004; 14:648–656. PubMed http://dx.doi.org/10.1016/j.tcb.2004.09.012
Waters CM, Bassler BL. Quorum Sensing: Cell-to-Cell Communication in Bacteria. Annu Rev Cell Dev Biol 2005; 21:319–246. PubMed http://dx.doi.org/10.1146/annurev.cellbio.21.012704.131001
Wagner-Döbler I, Ballhausen B, Berger M, Brinkhoff H, Buchholz I, Bunk B, Cypionka H, Daniel R, Gerdts G, Hahnke S, et al. The complete genome sequence of the algal symbiont Dinoroseobacter shibae: a hitchhiker’s guide to life in the sea. ISME J 2010; 4:61–77. PubMed http://dx.doi.org/10.1038/ismej.2009.94
Patzelt D, Wang H, Buchholz I, Rohde M, Gröbe L, Pradella S, Neumann A, Schulz S, Heyber S, Münch K, et al. You are what you talk: quorum sensing induces individual morphologies and cell division modes in Dinoroseobacter shibae. ISME J 2013; 7:2274–2286. PubMed http://dx.doi.org/10.1038/ismej.2013.107
Field D, Garrity G, Gray T, Morrison N, Selengut J, Sterk P, Tatusova T, Thomson N, Allen MJ, Angiuoli SV, et al. The minimum information about a genome sequence (MIGS) specification. Nat Biotechnol 2008; 26:541–547. PubMed http://dx.doi.org/10.1038/nbt1360
Field D, Amaral-Zettler L, Cochrane G, Cole JR, Dawyndt P, Garrity GM, Gilbert J, Glöckner FO, Hirschman L, Karsch-Mzrachi I, et al. The Genomic Standards Consortium. PLoS Biol 2011; 9:e1001088. PubMed http://dx.doi.org/10.1371/journal.pbio.1001088
Woese CR, Kandler O, Weelis ML. Towards a natural system of organisms. Proposal for the domains Archaea, Bacteria and Eucarya. Proc Natl Acad Sci USA 1990; 87:4576–4579. Med http://dx.doi.org/10.1073/pnas.87.12.4576
Garrity GM, Bell JA, Lilburn T. Phylum XIV. Proteobacteria phyl nov. In: Brenner DJ, Krieg NR, Stanley JT, Garrity GM (eds), Bergey’s Manual of Sytematic Bacteriology, second edition. Vol. 2 (The Proteobacteria), part B (The Gammaproteobacteria), Springer, New York, 2005, p. 1.
Garrity GM, Bell JA, Lilburn T. Class I. Alphapro teobacteria class. nov. In: Brenner DJ, Krieg NR, Stanley JT, Garrity GM (eds), Bergey’s Manual of Sytematic Bacteriology, second edition. Vol. 2 (The Proteobacteria), part C (The Alpha-, Beta-, Delta-, and Epsilonproteobacteria), Springer, New York, 2005, p. 1.
Validation List No. 107. List of new names and new combinations previously effectively, but not validly, published. Int J Syst Evol Microbiol 2006; 56:1–6. Med http://dx.doi.org/10.1099/ijs.0.64188-0
Garrity GM, Bell JA, Lilburn T. Order III. Rhodobacterales ord. nov. In: Brenner DJ, Krieg NR, Staley JT, Garrity GM (eds), Bergey’s Manual of Systematic Bacteriology, second edition. vol. 2 (The Proteobacteria), part C (The Alpha-, Beta-, Delta-, and Epsilonproteobacteria), Springer, New York, 2005, p. 161.
Garrity GM, Bell JA, Lilburn T. Family I. Rhodobacteraceae fam. nov. In: Brenner DJ, Krieg NR, Staley JT, Garrity GM (eds), Bergey’s Manual of Systematic Bacteriology, second edition. vol. 2 (The Proteobacteria), part C (The Alpha-, Beta-, Delta-, and Epsilonproteobacteria), Springer, New York, 2005, p. 161.
BAuA. Classification of Bacteria and Archaea in risk groups. TRBA 2010; 466:93.
Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT, et al. Gene ontology: tool for the unification of biology. The Gene Ontology Consortium. Nat Genet 2000; 25:25–29. PubMed http://dx.doi.org/10.1038/75556
PILER Genomic repeat analysis software. 2009.
Acknowledgements
The authors would like to gratefully acknowledge the assistance of Iljana Schröder for growing W. marina cultures and Meike Döppner for DNA extraction and quality control (both at the Leibniz-Institute DSMZ). This work was performed under the auspices of the German Research Foundation (DFG) Transregio-SFB51 Roseobacter grant and the US Department of Energy’s Office of Science, Biological and Environmental Research Program and by the University of California, Lawrence Berkeley National Laboratory under contract No. DE-AC02-05CH11231, Lawrence Livermore National Laboratory under Contract No. DE-AC52-07NA27344.
Author information
Authors and Affiliations
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Riedel, T., Fiebig, A., Han, J. et al. Genome sequence of the Wenxinia marina type strain (DSM 24838T), a representative of the Roseobacter group isolated from oilfield sediments. Stand in Genomic Sci 9, 855–865 (2014). https://doi.org/10.4056/sigs.5601028
Published:
Issue Date:
DOI: https://doi.org/10.4056/sigs.5601028