
Abstract
Three experiments with rats investigated whether adding or removing elements of a context affects generalization of instrumental behavior. Each of the experiments used a free operant procedure. In Experiments 1 and 2, rats were trained to press a lever for food in a distinctive context. Then, transfer of lever pressing was tested in a context created either by adding an element to the context of initial acquisition or by removing one of the acquisition context’s elements. In Experiment 3, a similar generalization test was conducted after rats received acquisition and extinction within the same context. For Experiments 1 and 2, we observed that removing elements from the acquisition context disrupted acquisition performance, whereas the addition of elements to the context did not. Experiment 3 revealed that removing elements from but not adding elements to the original context improved extinction performance. Our results are consistent with an elemental view of context representation.
Similar content being viewed by others
Learning and behavior occur within a particular set of stimuli referred to as background or context (Smith, 2007). Contextual cues have been found to strongly influence information retrieval and performance (Bouton, 2010; Smith & Vela, 2001). For example, Godden and Baddeley (1975) showed that participants remember more words of a list under water when they learned it under water than on land, and that recall performance on land is better when acquisition took place on land rather than under water. Contexts also play a fundamental role in the modulation of voluntary behavior as indicated by research on instrumental (operant) conditioning (for a review, see Bouton & Todd, 2014; Todd, Vurbic, & Bouton, 2014).
Instrumental learning refers to the ability to learn about the consequences of behavior. If an animal experiences that the performance of a certain response (e.g., a lever press) leads to a desirable outcome (e.g., food pellets), the probability of that response increases. Contextual changes conducted after the acquisition of an instrumental response were found to disrupt acquisition performance (e.g., Bouton, Todd, Vurbic & Winterbauer, 2011; Todd, 2013). In instrumental extinction, reinforced responses are decreased by withdrawal of the reinforcing outcome. However, extinguished instrumental responses can recover when tested outside the context of extinction. For instance, when animals acquire an instrumental response in Context A that is subsequently extinguished in a second Context B, the extinguished response recovers when tested in the initial acquisition Context A (ABA renewal; e.g., Nakajima, Tanaka, Urushihara & Imada, 2000) or in a novel Context C (ABC renewal; e.g., Todd, 2013). Extinguished instrumental responses were also found to recover when acquisition and extinction take place in the same context, but testing in a second one (AAB renewal; e.g., Bouton et al., 2011).
Research on contextual control of instrumental behavior is consistent with the principle that generalization of acquisition and extinction depend on a match between conditions present during learning and conditions present during testing. However, little is known about the rules governing generalization of instrumental learning across contexts. This is in part because research focusing on context-switch effects typically involved massive contextual manipulations in order to provide optimal conditions for such effects. One exception is provided by Todd, Winterbauer, and Bouton (2012), who investigated the role of context similarity on renewal of instrumental responding. They used three operant chambers as contexts with two of the chambers being relatively similar to each other (chambers were modified versions of the same manufactured model), while the third chamber was more distinct from the others (a different chamber model). Todd et al. found stronger ABC renewal when the pair of similar chambers served as the acquisition and test contexts, and the distinct chamber as the extinction context, compared to a condition in which the similar chambers were used as the extinction and test contexts, and the distinct chamber as the acquisition context—demonstrating the importance of context similarity for the generalization of instrumental performance. However, a more systematic manipulation of contextual stimuli would allow further insights into the mechanisms of generalization, which was the aim of the present study.
Contemporary theories of associative learning propose different rules governing generalization. Purely elemental theories such as the Rescorla–Wagner (Rescorla & Wagner, 1972) theory assume that any stimulus is represented as an array of individual elements, each encoding a specific feature of the stimulus and possessing the capacity to acquire associative strength. According to this view, generalization across stimuli depends on the number of elements of the training stimulus that is shared by the test stimulus. For instance, if a test stimulus is created by adding novel elements to the training stimulus (testing with Stimulus AB after training with Stimulus A), generalization should be perfect as all the elements that were learned about during training are present during testing. In contrast, if testing occurs with a stimulus that is created by removing elements of the training stimulus (testing with Stimulus A after training with Stimulus AB), generalization should be impaired because the contributions of the removed elements to performance are lost during testing.
Alternatively, Pearce’s (1987, 1994) configural theory assumes that the entire pattern of stimulation provoked by a stimulus results in one unitary representation that can develop associative connections. Generalization within the model is governed by a rule according to which the representation of a stimulus can be partially activated by other stimuli based on a measure of similarity that puts the number of common elements shared by two stimuli in relation to the total number of elements comprising each stimulus. Accordingly, generalization depends on the similarity between the training and test stimuli regardless of whether the test stimulus is created by adding elements to or removing elements from the training stimulus (testing with A after training with AB should impair generalization in the same way as testing with AB after training with A).
A third account is provided by the replaced elements conception (e.g., Brandon, Vogel, & Wagner, 2000), according to which the internal representation of a stimulus depends on the presence or absence of other stimuli. In keeping with an elemental approach, the model assumes that any stimulus is represented by an array of elements. However, when two stimuli are presented together, some of the elements of each stimulus are replaced by elements that encode the unique conjunction of stimuli. According to the model, any stimulus change, be it by adding or by removing elements, will inactivate a proportion of elements representing the stimulus. However, the proportion of inactivated elements will be greater when elements are removed than when they are added. Therefore, testing with A after training with AB and testing with AB after training with A should each result in impaired generalization, but the degree of impairment should be stronger in the former than the latter case.
The theoretical accounts summarized in the preceding paragraphs have been originally proposed to account for Pavlovian conditioning, where responding toward a conditioned stimulus (CS; e.g., a tone) is established by repeatedly pairing the CS with an unconditioned stimulus (US; e.g., shock). In Pavlovian extinction, acquired responding is decreased by presentations of the CS without the US (for a review, see Bouton & Woods, 2008; Delamater & Westbrook, 2014; Todd et al., 2014). As for instrumental extinction, extinguished Pavlovian responses were found to recover when tested outside the extinction context (e.g., Bernal-Gamboa et al., 2012; Bouton & Bolles, 1979; Bouton & Peck, 1989; Bouton & Ricker, 1994; Rescorla, 2008; Thomas, Larsen, & Ayres, 2003). However, in contrast to instrumental conditioning, Pavlovian acquisition performance was often found to generalize perfectly across contexts (e.g., Bouton & King, 1983; Bouton & Peck, 1989; Thomas et al., 2003). Nevertheless, under certain circumstances, contextual changes can also disrupt Pavlovian acquisition (e.g., Bonardi, Honey, & Hall, 1990; Hall & Honey, 1990). For instance, González, Quinn, and Fanselow (2003) investigated the generalization of contextual fear conditioning by systematically manipulating context stimuli. In one of their experiments, rats were conditioned with unsignalled footshocks in a context comprising two distinctive elements (e.g., a tone and an odor). Following contextual fear conditioning, rats were tested for generalization of fear either in a context in which one of the elements of the training context was removed or in a context in which a further element (e.g., a light) was added to the training context. González et al. found that acquisition perfectly generalized to the context with an added element, whereas testing in the context with a removed element resulted in a loss of fear. The asymmetrical generalization effect reported by González et al. is consistent with the view that contexts are represented in an elemental way (e.g., Rescorla & Wagner, 1972). However, whether the contextual control of instrumental behavior is also based on context representations according to elemental theories is unclear up to now.
The aim of the present experiments was to explore whether adding or removing elements of a context differentially affects generalization of instrumental behavior. Experiments 1 and 2 investigated generalization following free operant acquisition learning, while Experiment 3 explored whether generalization of instrumental extinction is affected by the same principles.
Experiment 1
The top panel of Table 1 presents the design of the two groups of Experiment 1. Rats were trained to press a lever for food in a particular context, followed by a generalization test, in which lever pressing was not rewarded, conducted in a changed environment. For one group of rats (Group A-AB), initial acquisition took place in a context containing one distinctive feature (Context A), while testing occurred in a context with an additional element (Context AB). For a second group (Group AB-A), the initial acquisition context comprised two distinctive features (Context AB), while one of these context elements was removed for generalization testing (Context A).
If generalization of instrumental behavior follows the principles advocated by the Rescorla–Wagner (1972) theory or the replaced elements model (Brandon et al., 2000), responding during the test should be stronger in Group A-AB than in Group AB-A. In contrast, according to Pearce’s (1987, 1994) configural theory, we should observe no difference in responding during the test across the groups.
Method
Subjects
Twenty-four female Wistar rats were used. They were about 3 months old and experimentally naïve at the beginning of the experiment. They were housed individually in methacrylate cages (21 × 24 × 46 cm, H × W × D) inside a room maintained on a 12–12 hr light–dark cycle. All subjects were maintained with ad libitum access to water but were food deprived to 83% of their initial body weights throughout the experiment, with an average maintenance weight of 223.5 g.
Apparatus
Eight identical chambers manufactured by MED Associates (Model ENV-008) measuring 29 cm height × 22 cm long × 24 cm wide were used. Each chamber was enclosed in a sound attenuating chamber equipped with an exhaust fan that produced background noise of 60 dB. The side walls and ceiling were made of clear acrylic plastic, while the front and rear walls were made of stainless steel. The floor of the chamber consisted of 16 0.5-cm diameter stainless steel rods spaced 1.5 cm apart. A recessed 5 cm × 5 cm food magazine in which 45 mg Noyes A/I pellets could be delivered was centered on the front wall. Each chamber had one retractable lever, which was positioned to the right of the food tray. Each lever was 4.8 cm long and positioned 6.8 cm above the floor. A 28 Vdc bulb was placed 4.2 cm above the lever, which served as a general house light. The chambers were connected to a PC that controlled and recorded the events.
We used three different additional stimuli in order to control the removing–adding manipulation. For a visual modality, the walls of the chambers were covered with a white sheet with red circles. The odor modality consisted of a vinegar scent provided with a dish containing 5 ml of white vinegar (Clemente Jacques, Sabormex S.A. de C.V., México, DF) placed outside each chamber near the front wall. For a tactile modality, we covered the floor with a sandpaper sheet (Number 10). The additional stimuli of different sensory modalities and their possible combinations were counterbalanced across rats.
Procedure
Sessions were conducted on successive days at the same time each day. The experiment was composed of two phases: Acquisition and test. During the first day all rats received acclimatization to the context. During this session, food pellets were delivered approximately 70 times on a variable time (VT) 30-s schedule. No lever was presented. The session lasted 30 min.
Acquisition
During the next 4 days, all rats were trained to press a left lever for food on a variable interval (VI) 30-s schedule. Rats in Group AB-A experienced this phase in a context with two of the three additional stimuli of different sensory modalities (visual, olfactory or tactile counterbalanced) presented throughout the session. For rats in Group A-AB, acquisition took place in a context with one of the three additional stimuli (visual, olfactory, or tactile counterbalanced). Each session lasted 30 min.
Test
On the following day, rats received a single 10 min test session for lever pressing. For rats in Group AB-A testing was conducted in a context with one element of the original context removed (visual, olfactory or tactile stimuli counterbalanced). Rats in the Group A-AB received this phase in a context with one additional element (visual, olfactory or tactile stimuli counterbalanced). In both groups, lever-pressing was not reinforced during the session.
Statistical analysis
For this and the following experiments, mean responses per minute were compared using analyses of variance (ANOVA). For all experiments, the rejection criterion was set at p < .05, and effect sizes were reported using partial eta-squared (ηp2).
Results and discussion
Acquisition
The left panel of Fig. 1 shows the mean responses per minute during each session of acquisition for both groups. The figure indicates that the response was acquired similarly by all rats and that the rate of responding increased as acquisition progressed.

Mean responses per minute during acquisition and test for Groups A-AB and AB-A in Experiment 1. Error bars denote standard errors of the mean
A 2 (group) × 4 (session) ANOVA conducted on the acquisition data showed a significant main effect of session, F(3, 66) = 66.75, mean square error (MSE) = 13.18, p = .001, ηp2 = .75. The ANOVA found no main effect of group, F < 1. The Group × Session interaction was not significant either, F(3, 66) = 1.21, MSE = 13.18, p = .31.
Test
The right panel of Fig. 1 shows the mean responses per minute for Groups A-AB and AB-A during testing. As can be seen, lever pressing was higher in Group A-AB than in Group AB-A. This was confirmed by a one-way ANOVA conducted with the test data revealing significant differences in responding between the groups, F(1, 22) = 11.63, MSE = 9.38, p = .002, ηp2 = .36.
In the present experiment, acquisition performance was weaker when tested in a context generated by removing one of the elements of the original acquisition context compared to a context created by adding one element to the context of initial acquisition. Our results are consistent with those reported by González et al. (2003) and extend their findings to an appetitive instrumental preparation.
Our finding that adding and removing context elements produced asymmetrical generalization of instrumental responding is consistent with the Rescorla–Wagner (1972) theory or the replaced elements model (Brandon et al., 2000), but contradictory to the predictions of Pearce’s (1987, 1994) configural theory. However, with Experiment 1 it was not possible to differentiate between the Rescorla–Wagner theory and the replaced elements model as the experiment is silent about the question whether or not the addition of context elements disrupted acquisition performance. Therefore, the purpose of the following experiment was to assess the individual impacts of adding and removing context elements on generalization of instrumental behavior.
Experiment 2
Experiment 2 was similar to Experiment 1, but comprised a third group of rats for which testing was conducted in the same context in which acquisition had taken place. Furthermore, we equated the conditions of acquisition across groups in Experiment 2. Three groups of rats were trained to press a lever for food in a context comprising two distinctive features, A and B. Following acquisition training, one group of rats was tested for generalization in the original acquisition context (Group AB-AB). For a second group, the acquisition context was changed for testing by removing one of its elements (Group AB-A), while for a third group, an element was added to the original acquisition context (Group AB-ABC).
For the test, the Rescorla–Wagner (1972) theory predicts that responding in Group AB-AB should be stronger than in Group AB-A, while there should be no difference in lever pressing between the Groups AB-AB and AB-ABC. However, according to Pearce’s (1987, 1994) configural theory or the replaced elements model (Brandon et al., 2000), responding during the test in Group AB-AB should be stronger compared to each of the Groups AB-A and AB-ABC.
Method
Subjects
Thirty-six female Wistar rats, weighing on average 241 g, were used. They were about three months old and experimentally naïve at the beginning of the experiment. They were maintained under the same conditions as in Experiment 1.
Apparatus and procedure
We used the same chambers and context stimuli of different sensory modalities as in Experiment 1. Sessions were conducted on successive days at the same time each day. The experiment was composed of two phases: Acquisition and test (see middle part of Table 1). During the first day all rats received a similar acclimatization to the context as in the previous experiment.
Acquisition
During the next four days, all rats were trained to press a left lever for food on a VI 30-s schedule. All rats experienced this phase in a context with two of the three additional stimuli of differing sensory modalities (visual, olfactory, or tactile counterbalanced) presented throughout the session. Each session lasted 30 min.
Test
On the following day, rats received a single 10-min test session for lever pressing. For rats in Group AB-A testing was conducted in a context with one element of the original context removed (visual, olfactory, or tactile stimuli counterbalanced). Rats in the Group AB-ABC received this phase in a context with one additional element (visual, olfactory, or tactile stimuli counterbalanced). Rats in Group AB-AB experienced this session in the same context of acquisition. In each group, lever pressing was not rewarded during the session.
Results and discussion
Acquisition
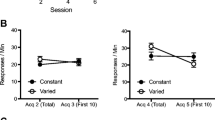
The left panel of Fig. 2 shows the mean responses per minute during each session of acquisition for all groups. The figure indicates that lever pressing was acquired similarly by all groups and that the rate of responding increased as this phase progressed.

Mean responses per minute during acquisition and test for Groups AB-ABC, AB-A, and AB-AB in Experiment 2. Error bars denote standard errors of the mean
A 3 (group) × 4 (session) ANOVA conducted on the acquisition data found a significant main effect of session, F(3, 99) = 99.21, MSE = 10.65, p = .001, ηp2 = .75. The main effect of group was not significant, F < 1. Also, the Group × Session interaction was not significant, F(3, 99) = 1, MSE = 10.65, p = .42.
Test
Mean responses per minute for all groups during testing are depicted in the right panel of Fig. 2. A one-way ANOVA conducted with the test data found that groups performed different levels of responding during the session, F(2, 33) = 5.45, MSE = 13.26, p = .008, ηp2 = .25. Follow-up comparisons indicated that rats in Group AB-ABC showed higher levels of responding than rats in Group AB-A, F(1, 33) = 9.46, MSE = 13.26, p = .004. The planned comparisons also found lower levels of responding in Group AB-A than in Group AB-AB, F(1, 33) = 6.64, MSE = 13.26, p = .01. Additionally, no difference was found between rats in Group AB-AB and rats in Group AB-ABC, F < 1.
Consistent with Experiment 1 and González et al. (2003), the present results revealed asymmetrical generalization of instrumental behavior—removing but not adding context elements produced a loss of lever pressing. Following acquisition in Context AB, responding was weaker when tested in a context created by removing one element from the acquisition context compared to a condition with unaltered contexts. However, when testing took place in a context generated by adding one element to the acquisition context, performance was similar to that in the condition with unaltered contexts. This pattern of results is consistent only with the Rescorla–Wagner (1972) theory. According to either the replaced elements model (Brandon et al., 2000) or Pearce’s (1987, 1994) configural theory, adding elements to the context of acquisition should have impaired generalization, which was not evident in the present results.
Experiment 3
The purpose of Experiment 3 was to investigate the impacts of adding and removing context elements on the generalization of instrumental extinction performance (see, bottom panel of Table 1). Following acquisition of an instrumental lever-pressing response in Context AB, lever pressing underwent extinction in the same Context AB. Finally, three groups of rats received a generalization test either in the same context (Group AB-AB), in a context created by adding one element to the original context (Group AB-ABC), or in a context generated by removing one of the original context’s elements (Group AB-A).
Note that the context manipulation in each of the Groups AB-ABC and AB-A corresponds to an AAB renewal protocol, with Group AB-AB being the AAA control condition. If instrumental extinction is controlled by context stimuli that are represented according to the Rescorla–Wagner (1972) theory, the context change following extinction in Group AB-A should disrupt extinction performance (lever pressing should recover compared to Group AB-AB), while there should be no difference in extinction performance between the Groups AB-ABC and AB-AB. However, if context stimuli are represented according to Pearce’s (1987, 1994) configural theory or the replaced elements model (Brandon et al., 2000), extinguished lever pressing should recover during the test in each of the Groups AB-ABC and AB-A.
Method
Subjects
Thirty-six female Wistar rats weighting in average 244.5 g were used. They were about three months old and experimentally naïve at the beginning of the experiment. They were maintained under the same conditions as described for Experiment 1.
Apparatus and procedure
We used the same chambers and the same context stimuli of different sensory modalities as described for Experiment 1. Sessions were conducted on successive days at the same time each day. The experiment was composed of three phases: acquisition, extinction and test (see lower part of Table 1). During the first day, all rats received a similar acclimatization to the context as in the previous experiments.
Acquisition
During the next 4 days, all rats were trained to press a left lever for food on a VI 30s schedule. All rats experienced this phase in a context with two of the three additional stimuli of different sensory modalities (visual, olfactory, or tactile counterbalanced) presented throughout the session. Each session lasted 30 min.
Extinction
On each of the next four days, all rats received one daily response extinction session. All rats received extinction training in the same context as acquisition. Each session lasted 30 min.
Test
On the following day, rats received a single 10-min test session for lever pressing. For rats in Group AB-A testing was conducted in a context with one element of the extinction context removed (visual, olfactory, or tactile stimuli counterbalanced). Rats in Group AB-ABC received this phase in a context with one additional element (visual, olfactory, or tactile stimuli counterbalanced). Group AB-AB experienced this session in the same context as extinction. In each group, lever pressing was not reinforced during the session.
Results and discussion
Acquisition
Mean responses per minute during acquisition for Groups AB-ABC, AB-A, and AB-AB are shown in the left panel of Fig. 3. As in the previous experiments no differences in acquisition rates were found across groups. And, as acquisition training progressed lever pressing increased.

Mean responses per minute during acquisition and extinction for Groups AB-ABC, AB-A, and AB-AB in Experiment 3. Error bars denote standard errors of the mean
A 3 (group) × 4 (session) ANOVA found a significant main effect of session, F(3, 99) = 106.00, MSE = 10.38, p = .001, ηp2 = .77. The main effect of group was not significant, F < 1. The ANOVA also found no Group × Session interaction, F(3, 99) = 1.22, MSE = 10.38, p = .30.
Extinction
The right panel of Fig. 3 depicts the mean responses per minute during each session of the extinction phase. A similar decrement in lever pressing was observed in the three groups. A 3 (group) × 4 (session) ANOVA conducted with the extinction data found a significant main effect of session, F(3, 99) = 122.64, MSE = 1.77, p = .001, ηp2 = .79. Both the main effect of group, F(2, 33) = 1.36, MSE = 4.07, p = .26, and the Group × Session interaction, F < 1, were not significant.
Test
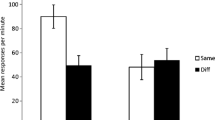
Figure 4 shows mean responses per minute for Groups AB-ABC, AB-A, and AB-AB during the test session. Figure 4 depicts that rats showed lower response rates when testing took place in a context with removed elements (Group AB-A).

Mean responses per minute during test for Groups AB-ABC, AB-A, and AB-AB in Experiment 3. Error bars denote standard errors of the mean
A one-way ANOVA indicated differences in levels of responding across groups, F(2, 33) = 3.46, MSE = .44, p = .04, ηp2 = .18. Follow-up comparisons confirmed that rats in Group AB-ABC performed higher levels of responding than those in Group AB-A, F(1, 33) = 3.81, MSE = .44, p = .05. And, rats in Group AB-AB pressed the lever more frequently than rats in Group AB-A, F(1, 33) = 6.26, MSE = .44, p = .01. Moreover, the planned comparisons showed no difference in performance between Groups AB-ABC and AB-AB, F < 1.
Following acquisition and extinction treatment in the same context, rats performed similar levels of responding when they were tested in the same extinction context (Group AB-AB) or in a context with an additional element (Group AB-ABC). However, when tested in a context created by removing one element from the extinction context (Group AB-A), rats showed increased extinction performance (less responding) compared to the condition with unaltered contexts.
Consistent with the results of the previous experiments and in accord with the Rescorla–Wagner (1972) theory, Experiment 3 provided no evidence that a context change achieved by the addition of context elements affected instrumental behavior. However, on the basis of the Rescorla–Wagner theory, we expected that the removal of context elements following extinction should disrupt extinction performance inducing AAB renewal, but in contrast, we observed the opposite result (for a further discussion, see the General Discussion section).
General discussion
Three experiments with rats in a free operant procedure explored generalization across contexts with added or removed elements. Experiments 1 and 2 replicated and extended findings by González et al. (2003). Following acquisition of instrumental responding, changing the acquisition context by removing context elements disrupted acquisition performance, while adding context elements to the original context did not affect acquired behavior. Experiment 3 revealed asymmetrical generalization following extinction treatment. Changing the extinction context by removing context elements increased extinction performance, while adding elements to the extinction context was without influence on behavior.
The results from Experiments 1 and 2 are consistent with an elemental view of context representation (e.g., Rescorla & Wagner, 1972). According to this view each context element gains control over instrumental acquisition behavior. Therefore, the removal of elements from the acquisition context disrupts acquisition behavior as the contributions of the removed elements to performance are lost. However, the addition of elements to the context of acquisition leaves acquisition performance unaffected because all the elements that comprise the acquisition context are still available. The results of the two experiments are not in support of Pearce’s (1987, 1994) configural theory or the replaced elements model (Brandon et al., 2000). According to either view, changing the context of acquisition by adding or removing context elements results in an internal context representation that differs from the representation encoding the acquisition context. Therefore, these accounts predict that the addition of context elements will impair acquisition performance, which was not evident in the present study.
The present results are consistent with previous studies that revealed similar asymmetrical generalization effects after removing elements from a CS (compared to adding CS elements) in nonhuman and human animals (e. g., Brandon et al., 2000; Glautier, 2004; Rescorla, 1999; Wheeler, Amundson & Miller, 2006). This consistency indicates parallels between processing a “simpler” stimulus compound (i.e., a brief CS) and a larger “complex” stimulus compound like a context (stimuli of long duration).
For Experiment 3, we expected AAB renewal of instrumental behavior, at least in the group in which the context of extinction was changed by removing context elements. We assumed that the extinction context acquires the ability to inhibit the instrumental response in some form (see below) and that any contextual manipulation that impairs this inhibitory control will induce a recovery of instrumental responding. In contrast to our prediction, we observed the opposite result. Rats that were tested in a context created by removing elements from the extinction context responded less compared to a condition with unaltered contexts. One way to reconcile the results from Experiment 3 with the Rescorla–Wagner (1972) theory is by assuming that our extinction treatment left the contextual stimuli with some excitatory strength toward the instrumental response. The context’s excitatory strength that may have survived our extinction treatment would generalize perfectly to the context with added elements, while it would be disrupted by removing elements from the extinction context.
The finding in Experiment 3 that responding was stronger in the test context created by adding context elements than in the test context created by removing elements could be explained by disinhibition. This phenomenon refers to response recovery that is produced by presenting a novel stimulus after experimental extinction (i.e., Pavlov, 1927). From this perspective, it could be assumed that changing the extinction context by adding a novel element would produce disinhibition inducing more responding. However, a disinhibition account would also predict stronger responding in the test context created by adding context elements compared to the condition with unaltered contexts, which was not evident in Experiment 3.
Bouton, Doyle-Burr, and Vurbic (2012) investigated generalization of extinction between compound CS and their elements. Using an appetitive conditioning procedure, rats were initially trained with A+ and AB+ trials, followed by extinction with either A or AB. During a subsequent test, Bouton et al. observed that animals responded more strongly to AB than A when they had received extinction with A, while animals that had been extinguished with AB responded equally to AB and A during the test. The authors concluded that extinction of AB generalized more to A than extinction of A generalized to AB. However, as noted by the authors, responding during the test may also reflect generalization of acquisition learning. During acquisition training with A+ and AB+, element B from the AB compound may have acquired excitatory strength. Following extinction with A, presentations of the excitatory element B in compound with A during the test may have increased responding to the compound. Future research is required to better understand the generalization of extinction across CS and across contexts.
The present study supports the view that the contextual control of instrumental behavior is based on context representations according to elemental theories (e.g., Rescorla & Wagner, 1972). However, the present experiments are silent about the associative structure that underlies the acquisition and extinction of instrumental responding. The representations of contextual elements may (a) enter into direct associations with the reinforcer, (b) develop direct connections to the response, or (c) modulate response-reinforcer associations. Previous studies revealed that direct context–response associations and context modulated response–reinforcer associations play an important role in instrumental learning (e.g., Bouton & Todd, 2014), however, future research is required to uncover the associative structures that underlie the behavioral effects induced by adding and removing context elements.
The results of the present experiments revealed strong parallels between processing of CS compounds and more complex context compositions. We found asymmetrical generalization following both acquisition and extinction learning. Removing elements of a context impaired generalization of acquisition, whereas removing contextual elements improved extinction performance. In either case, adding elements to the original context was without influence on behavior. Given the similarity with results reported in Pavlovian preparations, the present findings suggest a common mechanism that underlies context processing in Pavlovian and instrumental learning.
References
Bernal-Gamboa, R., Juárez, Y., González-Martín, G., Carranza, R., Sánchez-Carrasco, L., & Nieto, J. (2012). ABA, AAB and ABC renewal in taste aversion learning. Psicológica, 33, 1–13.
Bonardi, C., Honey, R. C., & Hall, G. (1990). Context specificity of conditioning in flavor aversion learning: Extinction and blocking tests. Animal Learning & Behavior, 18, 229–237. https://doi.org/10.3758/BF03205280
Bouton, M. E. (2010). The multiple forms of context in associative learning. In B. Mesquita, L. Feldman Barret, & E. Smith (Eds.) The mind in context (pp. 233–258) New York: Guilford Press.
Bouton, M. E., & Bolles, R. C. (1979). Contextual control of the extinction of conditioned fear. Learning and Motivation, 10, 445–466. https://doi.org/10.1016/0023-9690(79)90057-2
Bouton, M.E., & Todd, T.P. (2014). A fundamental role for context in instrumental learning and extinction. Behavioural Processes, 104, 13–19. https://doi.org/10.1016/j.beproc.2014.02.012
Bouton, M. E., Doyle-Burr, C., & Vurbic, D. (2012). Asymmetrical generalization of conditioning and extinction from compound to element and element to compound. Journal of Experimental Psychology: Animal Behavior Processes, 38(4), 381–393. https://doi.org/10.1037/a0029726
Bouton, M. E., & King, D. A. (1983). Contextual control of the extinction of conditioned fear: Tests for the associative value of the context. Journal of Experimental Psychology: Animal Behavior Processes, 9(3), 248–265. https://doi.org/10.1037/0097-7403.9.3.248
Bouton, M. E., & Peck, C. A. (1989). Context effects on conditioning, extinction, and reinstatement in an appetitive conditioning preparation. Animal Learning and Behavior, 17, 188–198. https://doi.org/10.3758/BF03207634
Bouton, M. E., & Ricker, S. T. (1994). Renewal of extinguished responding in a second context. Animal Learning and Behavior, 22, 317–324. https://doi.org/10.3758/BF03209840
Bouton, M. E., Todd, T. P., Vurbic, D. & Winterbauer, N. (2011). Renewal after the extinction of free operant behavior. Learning and Behavior, 39, 57–67. https://doi.org/10.3758/s13420-011-0018-6
Bouton, M. E., & Woods, A. M. (2008). Extinction: Behavioral mechanisms and their implications. In J. H. Byrne, D. Sweatt, R. Menzel, H. Eichenbaum, & H. Roediger (Eds.), Learning and memory: A comprehensive reference: Vol. 1, Learning theory and behaviour (pp. 151–171). Oxford: Elsevier.
Brandon, S. E., Vogel, E. H., & Wagner, A. R. (2000). A componential view of configural cues in generalization and discrimination in Pavlovian conditioning. Behavioural Brain Research, 110, 67–72. https://doi.org/10.1016/S0166-4328(99)00185-0
Delamater, A. R., & Westbrook, R. F. (2014). Psychological and neural mechanisms of experimental extinction: A selective review. Neurobiology of Learning and Memory, 108, 38–51. https://doi.org/10.1016/j.nlm.2013.09.016
Glautier, S. (2004). Asymmetry of generalization decrement in causal learning. The Quarterly Journal of Experimental Psychology B: Comparative and Physiological Psychology, 57, 315–329. https://doi.org/10.1080/02724990344000169
Godden, D. R., & Baddeley, A. D. (1975). Context-dependent memory in two natural environments: On land and underwater. British Journal of Psychology, 66, 325–331. https://doi.org/10.1111/j.2044-8295.1975.tb01468.x
González, F., Quinn, J. J., & Fanselow, M. S. (2003). Differential effects of adding and removing components of a context on the generalization of conditional freezing. Journal of Experimental Psychology: Animal Behavior Processes, 29, 78–83. https://doi.org/10.1037/0097-7403.29.1.78
Hall, G., & Honey, R. C. (1990). Context-specific conditioning in the conditioned-emotional-response procedure. Journal of Experimental Psychology: Animal Behavior Processes, 16, 271–278. https://doi.org/10.1037/0097-7403.16.3.271
Nakajima, S., Tanaka, S., Urushihara, K., & Imada, H. (2000). Renewal of extinguished lever-press responses upon return to the training context. Learning and Motivation, 31, 416–431. https://doi.org/10.1006/lmot.2000.1064
Pavlov, I. P. (1927). Conditioned reflex. London: Oxford University Press.
Pearce, J. M. (1987). A model for stimulus generalization in Pavlovian conditioning. Psychological Review, 94, 61–73.
Pearce, J. M. (1994). Similarity and discrimination: A selective review and a connectionist model. Psychological Review, 101, 587–607.
Rescorla, R. A. (1999). Associative changes in elements and compounds when the other is reinforced. Journal of Experimental Psychology: Animal Behavior Processes, 25, 247–255. https://doi.org/10.1037/0097-7403.25.2.247
Rescorla, R. A. (2008). Within-subject renewal in sign tracking. The Quarterly Journal of Experimental Psychology, 61, 1793–1802. https://doi.org/10.1080/17470210701790099.
Rescorla, R. A., & Wagner, A. R. (1972). A theory of Pavlovian conditioning: Variations in the effectiveness of reinforcement and nonreinforcement. In A. H. Black & W. F. Prokasy (Eds.), Classical conditioning II: Current research and theory (pp. 64–99), New York: Appleton-Century-Crofts.
Smith, S. M. (2007). Context: A reference for local experience. In H. L. Roediger, Y. Dudai, & S. M. Fitzpatrick (Eds.), Science of memory: Concepts (pp. 111–114). Oxford: Oxford University Press.
Smith, S. M., & Vela, E. (2001). Environmental context-dependent memory. In G. M. Davies & D. M. Thomson (Eds.), Memory in context: Context in memory (pp. 13–34). New York, Wiley.
Thomas, B. L., Larsen, N., & Ayres, J. B. (2003). Role of context similarity in ABA, ABC and AAB renewal paradigms: Implications for theories of renewal and for treating human phobias. Learning and Motivation, 34, 410–436. https://doi.org/10.1016/S0023-9690(03)00037-7
Todd, T. P. (2013). Mechanisms of renewal after the extinction of instrumental behavior. Journal of Experimental Psychology: Animal Behavior Processes, 39, 193–207. https://doi.org/10.1037/a0032236
Todd, T. P., Vurbic, D., & Bouton, M. E. (2014). Behavioral and neurobiological mechanisms of extinction in Pavlovian and instrumental learning. Neurobiology of Learning and Memory, 108, 52–64. https://doi.org/10.1016/j.nlm.2013.08.012
Todd, T. P., Winterbauer, N. E., & Bouton, M. E. (2012). Effects of the amount of acquisition and contextual generalization on the renewal of instrumental behavior after extinction. Learning & Behavior, 40(2), 145–157. https://doi.org/10.3758/s13420-011-0051-5
Wheeler, D. S., Amundson, J. C., & Miller, R. R. (2006). Generalization decrement in human contingency learning. Quarterly Journal of Experimental Psychology, 59, 1212–1223. https://doi.org/10.1080/17470210600576342
Acknowledgements
This research was funded by UNAM-DGAPA through grant project PAPIIT IN306817. Contributions by Metin Uengoer to this research were supported by the German Research Foundation (DFG) through FOR 1581. Correspondence concerning this article should be addressed to Metin Uengoer (uengoer@staff.uni-marburg.de).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Bernal-Gamboa, R., Nieto, J. & Uengoer, M. Removing but not adding elements of a context affects generalization of instrumental responses. Learn Behav 46, 256–264 (2018). https://doi.org/10.3758/s13420-017-0307-9
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13420-017-0307-9