
Abstract
Experiments 1A and 1B used a taste-aversion procedure with rats to demonstrate that exposure to easily discriminated flavors along a dimension (1 % and 10 % sucrose) can facilitate learning a subsequent hard discrimination (4 % and 7 % sucrose) when one of those flavors is paired with illness. Experiment 1A compared the effects of preexposure to the easily discriminated flavors against exposure to the same stimuli used in the discrimination training or no exposure at all. Experiment 1B replicated the conditions in Experiment 1A, with 2 additional days of training and unrestricted access to the flavors on CS+/CS– trials in discrimination training. Contrary to findings with multidimensional stimuli (Scahill & Mackintosh, Journal of Experimental Psychology: Animal Behavior Processes, 30, 96–103, 2004; Suret & McLaren, The Quarterly Journal of Experimental Psychology, 56B, 30–42, 2003), we found that preexposure to the easily discriminable stimuli varying along a single dimension of sweetness facilitated subsequent discrimination training over the other conditions in each experiment. We discuss the results in terms of the ideas presented by Gibson (1969) and Mackintosh (Psychological Review, 82, 276–298, 1975) and in terms of hedonic variables not considered by theories of perceptual learning.
Similar content being viewed by others

Lawrence (1952) was the first to clearly show the “easy-to-hard effect” (ETH), an effect that was indicated by work as early as Pavlov (1927). The essentials of the effect are demonstrated most clearly by two of the four groups in Lawrence’s work. In a test phase, both groups of rats were required to discriminate between two similarly shaded gray goal boxes. In one box, rats would receive food reinforcement, but not in the other. The critical manipulation occurred prior to the test. One group had received training on the discrimination with the same gray goal boxes while the other had been trained on an easier version in which the two shades of gray were further apart in brightness. The rats that first received training on the easy discrimination performed better on the subsequent hard discrimination than did those that had received training with the hard discrimination throughout. Since Lawrence’s demonstration, the ETH effect has been widely replicated (see Hall, 1991, for a review).
The phenomenon has been effectively explained in associative terms in two ways. First, the effect is expected simply because of less overlap of the excitatory and inhibitory gradients around the reinforced and nonreinforced stimuli, generalizing to the hard discrimination. Second, attention is assumed to play a role. Attention to the relevant dimensions can increase during pretraining, and elements of the relevant dimension are more prevalent in the easy condition, ensuring they are already attended to at the onset of the hard discrimination (for a review, see Hall, 1991; Suret & McLaren, 2003).
According to Gibson (1969), changes in the representations of stimuli underlie any behavioral changes, and exposure to stimuli should make the representations become more detailed regardless of the presence of external reinforcement. Learning to differentiate stimuli, according to Gibson, is reinforcing in and of itself, because it reduces uncertainty about the environment. For Gibson, exposure to stimuli allows organisms to ignore features that are not useful in distinguishing stimuli, and to abstract those features that are distinctive.
Gibson proposed that the mechanism whereby the distinctive features are abstracted can be facilitated by experiencing the stimuli with the unique features enhanced or exaggerated. One way of doing this is to maximize the contrast between the distinctive features. Therefore, experiencing the stimuli with more salient distinctive elements, as when subjects experience the easy version of an ETH discrimination, should improve subsequent discriminations.
Though Gibson offered no mechanistic account of how these processes operate, Saksida (1999) offers one approach that can describe these processes. Saksida also pointed out that the ETH effect can be considered to be a case of perceptual learning, because it reflects an increase in a subject’s ability to distinguish similar stimuli. As such, the effect should appear if subjects are simply preexposed to the stimuli from the easy discrimination; no explicit pretraining on a discrimination involving reinforcement should be required. These issues are well discussed in Scahill and Mackintosh (2004).
A perceptual learning approach can explain the ETH effect, and it predicts that such an effect should emerge after simple preexposure. However, this prediction has not been confirmed (Scahill & Mackintosh, 2004; Suret & McLaren, 2003). In the experiments of Scahill and Mackintosh, the stimuli were flavor compounds of saline and lemon or a sweet taste (sucrose or saccharine) and lemon. Varying by experiment, the discrimination difficulty was altered by changing the intensity of the saline and sweet tastes, or manipulating the presence/absence of the lemon. An ETH effect was obtained when the easy discrimination was pretrained. However, simple exposure to the easy discrimination was no more effective than preexposure to the hard discrimination. A similar pattern of results was obtained by Suret and McLaren; in their study, humans were required to categorize faces. There, the stimuli employed were two relatively distinct photographs that had been progressively morphed together, from which easy- or hard-to-discriminate stimulus pairs were selected.
The present experiments investigated whether an ETH effect would emerge in rats as a function of simple preexposure using a flavor-aversion procedure conceptually more analogous to other demonstrations of the ETH effect; see, for example, Lawrence (1952), where the stimuli varied along a single dimension. In the present experiment, the stimuli varied along a single continuum of sweetness. In the work described earlier, the stimuli were more complex and multifaceted, varying along dimensions in proportion to each other. For example, in the work of Scahill and Mackintosh (2004), sweet and salty tastes varied in proportion to sourness. The stimuli in the work of Suret and McLaren (2003) are arguably considerably more complex, making it difficult to isolate the degree to which various potential elements changed individually.
Experiments 1A and 1B used a flavor-aversion procedure in which rats received a difficult discrimination between sucrose concentrations of 4 % and 7 %. One of these flavors was paired with lithium chloride–induced illness and the other was not. Prior to the discrimination, Group Easy received four preexposures to 1 % and 10 % sucrose concentrations, in alternation. Group Hard received four alternated preexposures to the same 4 % and 7 % sucrose concentrations as were used in the discrimination. Group No Exposure received no preexposures. Experiment 1B replicated the conditions in Experiment 1A, with minor differences noted in the Method section below.
In addition to an ETH effect, comparisons of both preexposed groups to Group No Exposure allow for an assessment of whether the preexposure could produce any perceptual learning effect. Exposure to the stimuli, either the hard or easy versions, should allow latent inhibition to especially accrue to the elements they hold in common (see McLaren & Mackintosh, 2000). Thus, this latent inhibition should lead to more conditioning of the unique elements of the reinforced stimulus, reducing generalization to the other. As observed by Scahill and Mackintosh (2004), this effect should be equally prevalent in Groups Easy and Hard in the absence of an ETH effect. The ETH effect would appear as a better discrimination in Group Easy than in Group Hard.
Method
Subjects and apparatus
Subjects were male Wistar rats in both experiments (Ns = 18 and 24 in Experiments 1A and 1B, respectively), with average weights of 375 g and 339 g in Experiments 1A and 1B, respectively. Rats in Experiment 1A had previously served in an experiment using conditioned bar-press suppression, with stimuli and treatments unrelated to the present experiment. Rats in Experiment 1B were experimentally naive. All rats were housed individually under constant environmental conditions (temperature 23 ° C, 50 % humidity) on a 12/12 light–dark cycle, with the light cycle beginning at 8 a.m. Food was available ad libitum. Water access was restricted during experimental sessions. The experiment was conducted in the home cages. The stimuli employed were solutions of 1 %, 4 %, 7 %, and 10 % sucrose (weight to volume).
Procedure
Water deprivation and preexposure
Access to water was restricted to two 30-min sessions per day for the duration of the experiment. The first session was at 10:00 a.m. and the second at 4:00 p.m. During the first 6 days, water was provided on each daily session. Groups Easy, Hard, and No Exposure (ns = 6 in Experiment 1A, ns = 8 in Experiment 1B) were then formed by matching subjects on consumption during these first 6 days.
On the next 8 days, experimental treatments were delivered in the morning sessions and water was available in the afternoon. During each session, 10 ml of fluid was available. In the morning sessions, Group Easy received solutions of 1 % and 10 % sucrose on alternate days, whereas Group Hard received solutions of 4 % and 7 % sucrose (solution orders were counterbalanced). Group No Exposure received water.
Discrimination training
All rats received discrimination training between 4 % and 7 % sucrose solutions, where one of the solutions (counterbalanced) served as CS+ and was paired with intraperitoneal injections of lithium chloride (LiCL) at .3 M per 10 ml/kg of body weight and the other (CS–) was not. This dose was chosen because it was used by Symonds and Hall (1995) in their work demonstrating perceptual learning in taste aversion. In the morning sessions, CS+ or CS– was available and the injection was administered on CS+ trials immediately at the end of the 30-min session. In Experiment 1A, 10 ml of fluid was available on each session. In Experiment 1B, fluid access was unrestricted during the 30-min morning session in this phase.
Discrimination training lasted 42 days, in 3-day sequences. Each sequence began with a reinforced trial followed by a nonreinforced trial and then a recovery day, on successive days, for a total of 14 trials with each stimulus. In Experiment 1B, discrimination training lasted 48 days, for a total of 16 trials with each stimulus.
Data and analysis
Data were the number of milliliters of CS+ and CS– consumed, determined by weighing the drinking tubes before and after the sessions. The data were analyzed with mixed factorial analysis of variance (ANOVA) using standard Type III sums of squares. The rejection criterion was p < .05. ANOVA was also used for pairwise comparisons using error terms and degrees of freedom appropriately derived from the omnibus analysis, as described by Howell (1987, pp. 432–435). To add protection to what is afforded by the omnibus ANOVAs, alpha on pairwise comparisons was adjusted to control the false-discovery rate using the Benjamini–Hochberg method (Benjamini & Hochberg, 1995). Effect sizes are reported as partial eta squared for analyses involving more than two means, and as Cohen’s d for pairwise comparisons.
Results
Although the rats received several injections of LiCL, each injection was separated by 2 days. The rats’ body weights were maintained throughout the experiment, and there were no obvious behavioral complications as a result of the injections.
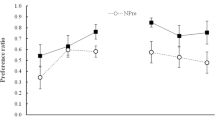
In both experiments, rats consumed the entire 10 ml of fluid available during each preexposure session. Data from discrimination training are shown in Fig. 1. Experiment 1A is shown at left and 1B at right. In both experiments, a perceptual learning effect may have been observed early in training, but an ETH effect was clearly observed later in training. Early in training, consumption of CS– (each of which followed a CS+ trial) was less than that of CS+, with less generalization (more consumption) in the preexposed conditions. Later in training, there was less generalization to CS– in Group Easy than in the other groups. A successful discrimination was obtained in Group Easy in both experiments. No discrimination was evident in the other groups in Experiment 1A, and the discrimination in the other groups was smaller than that of Group Easy in Experiment 1B. The analyses below support these conclusions.

Consumption of CS+ (solid symbols) and CS– (open symbols) during discrimination training with 4 % and 7 % sucrose solutions in Experiments 1A (left) and 1B (right). Prior to discrimination training, Group Easy (circles) received exposure to 1 % and 10 % sucrose solutions, Group Hard (squares) received exposure to the 4 % and 7 % solutions used in training, and Group No Exposure (triangles) received exposure to water. Fluid access in discrimination training was restricted to 10 ml on conditioning sessions in Experiment 1A and was unrestricted in 1B.
Experiment 1A
A Group × Stimulus × Trials ANOVA revealed a three-way interaction [F(26,195) = 3.54, p < .0001, η2 p = .32]. Stimulus × Trials ANOVAS within the groups revealed interactions in each [Fs(13,65) > 7.03, ps < .0001, η2 p > .58]. Similarly, Group × Trials ANOVAs on each stimulus revealed interactions on both CS+ and CS– [Fs(26,195) > 3.16, ps < .0001,η2 p > .36]. One-way ANOVAs on each trial within each stimulus showed effects of group on both CS+ (Trials 2, 3, and 10–14) and CS– (Trials 1 and 8–14) [Fs(2,33) > 3.65, ps < .037, η2 p min-max = .18–.57].
Between subjects there are 21 meaningful pairwise comparisons among the 7 trials where CS+ differences were indicated, and 24 such comparisons among the 8 trials where differences in CS– were indicated. Within subjects, there are 14 possible comparisons of CS+ to CS– on each trial in each group, for a total of 42 comparisons. Alpha on the following analyses was therefore adjusted for 87 comparisons. Thirty-eight comparisons were significant.
In the first half of discrimination training, the preexposed groups did not differ in consumption of CS+ and each drank more than Group No Exposure on Trials 2 and 3 [Fs(1,132) > 5.57, ps < .018, d min-max = 1.23–2.36]. The two preexposed groups did not differ in consumption of CS–, and each consumed more than Group No Exposure on Trial 1 [Fs(1,132) > 15.22, ps < .0002, d min-max = 1.97–2.54]. Consistent with those patterns, there was reliably less consumption of CS– than CS+ on the first three trials in Groups Easy and Hard, and on the first two trials in Group No Exposure [Fs(1,310) > 6.42, ps < .01, d min-max = 1.48–4.98].
Group Easy consumed more CS– than did Group No Exposure on Trials 8–14, and more than Group Hard on Trials 9–14 [Fs(1,132) > 5.57, ps < .02, d min-max = .93–5.28]. Group Hard differed from Group No Exposure only on the final trial [F(1,132) = 5.5, p = .02, d = 1.34]. Group Easy consumed more CS+ than Group Hard on Trials 10, 11, and 14, and more than Group No Exposure on Trials 11–14 [Fs(1,132) > 6.29, ps < .01, d min-max = 1.07–2.55]. Groups Hard and No Exposure never differed. Though consumption of both CS+ and CS– differed between the groups, the difference in consumption between the stimuli was reliable only in Group Easy, with reliably less consumption of CS– than CS+ on Trials 10 and 13 [Fs(1,310) > 5.50, ps < .02, d = .81–1.02]. Consumption of the stimuli was equivalent on every trial in Groups Hard and No Exposure in the latter half of discrimination training. As indicated by the pattern of significant differences above, the size of the discrimination (CS– minus CS+) was greater in Group Easy than in Group Hard in the latter half of training, as confirmed by a Trials × Group interaction [F(6,60) = 2.32, p = .04, η2 p = .19] on the CS–/CS+ difference with these groups.
The average within-group variance on a trial was 20.84. The average within-subject variance was 3.36.
Experiment 1B
A Group × Stimulus × Trials ANOVA revealed a three-way interaction [F(30,315) = 2.17, p = .0006, η2 p = .17]. Stimulus × Trials ANOVAS within each group revealed interactions in each [Fs(15,105) > 16.92, ps < .0001, η2 p min-max = .70–.75]. Group × Trials ANOVAs on each stimulus type revealed Group × Trials interactions on CS– [F(30,315) = 2.39, p = .0001, η2 p = .19] and only an effect of trials on CS+ [F(15,315) = 124.72, p < .0001, η2 p =.86]. One-way ANOVAs on CS– on each trial showed effects of group on Trials 1 and 10–16 [Fs(1,150) > 6.60, ps < .002, η2 p min-max = .13–.35].
Between subjects, there are 24 meaningful pairwise comparisons among the 8 trials where differences in consumption of CS– were indicated. Within subjects, there are 16 possible comparisons of CS+ to CS– on each trial for each group, resulting in 48 possible comparisons. Alpha on subsequent analyses, described below, was therefore adjusted for 72 comparisons. Thirty-five comparisons were significant.
On Trial 1, the preexposed groups did not differ among themselves, and each consumed more CS– than did Group No Exposure [Fs(1, 150) > 8.1, ps < .005, d min-max 1.5–1.67]. In the first half of training, the difference in consumption between CS+ and CS– was significant on the first 3 trials in all three groups [Fs(1, 368) > 9.35, ps < .01, d min-max = 1.17–3.19].
On Trials 9–16, Groups No Exposure and Hard did not differ. Group Easy consumed more CS– than did Group Hard on Trials 12–16, and more than Group No Exposure on Trials 10–16 [Fs(1,150) > 6.4, ps < .012, d min-max = .83–1.31]. In the latter half of training, the difference in consumption between CS+ and CS– was significant on Trials 10–16 in Group Easy, Trials 10, 14, and 15 in Group Hard, and Trials 15–16 in Group No Exposure [Fs(1,368) > 7.57, ps < .018, d min-max = .94–2.55]. The size of the discrimination (CS– minus CS+) was greater in Group Easy than in Group Hard. Though there was no Trials × Group interaction when comparing Group Easy to Group Hard on the CS–/CS+ difference, as in Experiment 1A, the main effect of group was reliable [F(1,14) = 5.47, p = .035, η2 p = .28] on this difference score.
The average variance within a group on a trial was 18.22. The average within-subject variance was 12.42.
Discussion
Experiments 1A and 1B were designed to assess whether an ETH effect could be observed after exposure to easily discriminable stimuli that varied along a single dimension. The common result between both experiments was that preexposure to 1 % and 10 % solutions of sucrose facilitated discriminating between 4 % and 7 % more than preexposure to the 4 % and 7 % solutions themselves. The experiments reported here are the first to show an ETH effect following simple preexposure.
A perceptual learning effect may have been observed in Group Hard. Exposure to the stimuli should allow latent inhibition to form to the common elements, reducing the extent to which they can be conditioned and thereby reducing generalization. Early in training, there was less generalization to CS– in Group Hard than in Group No Exposure in both experiments. In Experiment 1A, there was a reliable difference in consumption of CS+ early in training, but not in Experiment 1B. Whether the differences in CS– were due to true changes in generalization in both experiments or because consumption of CS– simply provided a more sensitive assessment of latent inhibition’s interference with the conditioning of CS+, cannot be determined. The possibility that the perceptual learning–like effect we observed simply reflects differential rates of conditioning to the preexposed versus novel flavors is strengthened by the fact that the apparent facilitation observed early in training in the animals exposed to the hard stimuli was not present later in training.
The present findings are at odds with those of Scahill and Mackintosh (2004) and Suret and McLaren (2003), who both failed to observe any benefit of exposure to easily discriminated stimuli over exposure to hard-to-discriminate stimuli. In Scahill and Mackintosh, the study that is procedurally more similar to the present experiments, easy discriminations were produced by using higher concentrations of salt and sweet tastes relative to lemon, compared with those in the hard discrimination, while holding the lemon taste constant. In their Experiment 1B, an additional easy condition was created by using low concentrations of salt and sweet tastes without lemon. In no case was preexposure to the easy discrimination more effective than preexposure to the hard discrimination. In the case in which preexposure to the easy discrimination was produced by removing lemon, preexposure was less effective than that to the hard stimuli. In each experiment there appeared to be a perceptual learning effect. Groups receiving preexposure learned the hard discrimination more rapidly, at least in comparison with the group that received training with the hard discrimination at an earlier point in the experiment without preexposure.
Scahill and Mackintosh (2004) concluded that latent inhibition was likely to be the only mechanism operating during preexposure. Latent inhibition would accrue equally to lemon in the compounds containing it, regardless of the concentrations of the unique elements, allowing the compounds to be more easily discriminated after preexposure. Consistent with their analysis, there was less benefit of preexposure in easily discriminated flavors that did not contain an explicit common element than in groups for which lemon was present. They did not, however, apply their analysis of latent inhibition to the changes in the intensity of the salt and sour tastes. For example, they gave no indication how latent inhibition to a more salty flavor encountered in preexposure may affect a less salty flavor on the discrimination test.
Because latent inhibition might explain the null effect Scahill and Mackintosh (2004) obtained with their multicomponent stimuli, it cannot be expected to explain the positive result obtained here. Moreover, an analysis based only on latent inhibition could be expected to predict the opposite of the results we obtained. Viewing the stimuli as existing along a dimension of components (i.e., s1–s10), the middle stimuli (i.e., s5–s6) will be sampled less frequently with exposure to the easy stimuli (s1, s10) than exposure to the difficult stimuli. Thus, after exposure to s1 and s10, there will be less latent inhibition of the middle stimuli, those between s4 and s7, than exposure to s4 and s7 would produce. Because it is conditioning of those middle stimuli that would make evidence of a discrimination difficult to observe, preexposure to the easy stimuli should result in a hard discrimination being more difficult than after preexposure to the hard stimuli themselves.
These results appear to require an enhancement of attention to the unique, differentiating aspects of the stimuli, as Scahill and Mackintosh (2004) and Gibson (1969) suggest is responsible for an ETH effect. We believe that there are two differences between our study and theirs that could account for why we obtained an effect. First, we used a stimulus that varied along a single dimension. Second, we used a sweet stimulus with a clear hedonic value. The use of a stimulus that varies along a single dimension might be more effective in activating attention mechanisms. It is a reasonable assumption that with fewer dimensions along which to detect differences, such differences are more difficult to detect and may require heightened attention to detect the different dimension.
There is also reason to expect that varying the stimulus’s intensity along a hedonic dimension may have been important, because it could allow a role for differential reinforcement. The use of easily discriminable stimuli of high hedonic value could produce an anticipatory contrast effect (see, e.g., Flaherty & Largen, 1975). As described by Flaherty and Checke (1982), the taste of one stimulus serves as a “retrieval cue” for the memory of the other, and directly affects the hedonic value of the stimulus. Presentations of less sweet would remind the animal of more sweet, which could produce frustration on less-sweet trials, even when the presentations are separated by 24 h (Flaherty, Lombardi, Wrightson, & Deptula, 1980). Such a contrast could also serve to make sweet trials particularly reinforcing (see, e.g., Capaldi, Sheffer, & Pulley, 1989). To the extent that the preexposure phase could produce such a contrast effect, the 4 % and 7 % solutions in the hard discrimination, like the weak concentrations used by Scahill and Mackintosh, are unlikely sufficiently hedonically different to do so. Because the easy stimuli could produce different reactions that would be heightened with preexposure, attention to the single varying dimension could have been heightened by a mechanism such as that described by Mackintosh (1975).
Based on the analysis above, the present experiments cannot differentiate between Gibson’s (1969) account, whereby detecting differences in stimuli is sufficiently reinforcing to enhance their unique features, or whether the stimuli must be paired with differential outcomes to produce the effect, as Mackintosh (1975) would anticipate. The present experiments do demonstrate a clear ETH effect following simple preexposure to stimuli that vary in a single dimension of sweetness intensity. It is an important demonstration, because such an effect appears to depend on an enhancement in the processing of the unique features of the stimuli.
References
Benjamini, Y., & Hochberg, Y. (1995). Controlling the false discovery rate: A practical and powerful approach to multiple testing. Journal of the Royal Statistical Society Series, 57, 289–300.
Capaldi, E., Sheffer, J. D., & Pulley, R. J. (1989). Contrast effects in flavor preference learning. The Quarterly Journal of Experimental Psychology, 41B, 307–323.
Flaherty, C. F., & Checke, S. (1982). Anticipation of incentive gain. Animal Learning & Behavior, 10, 177–182.
Flaherty, C. F., & Largen, J. (1975). Within subjects positive and negative contrast effects in rats. Journal of Comparative and Physiological Psychology, 88, 653–664.
Flaherty, C. F., Lombardi, B. R., Wrightson, J., & Deptula, D. (1980). Conditions under which chlordizepoxide influences gustatory contrast. Psychopharmacology, 67, 269–277.
Gibson, E. J. (1969). Principles of perceptual learning and development. New York: Appleton-Century –Crofts.
Hall, G. (1991). Perceptual and associative learning. Oxford, UK: Clarendon Press.
Howell, D. (1987). Statistical Methods for Psychology (2nd ed.). Boston, Massachusetts: PWS-KENT Publishing Company.
Lawrence, D. H. (1952). The transfer of a discrimination along a continuum. Journal of Comparative Psychology, 45, 511–516.
Mackintosh, N. J. (1975). A theory of attention: Variations in the associability of stimuli with reinforcement. Psychological Review, 82, 276–298.
McLaren, I. P. L., & Mackintosh, N. J. (2000). An elemental model of associative learning: I. Latent inhibition and perceptual learning. Animal Learning & Behavior, 28, 211–246.
Pavlov, I. P. (1927). Conditioned reflexes: An investigation of the physiological activity of the cerebral cortex. (Trans./Ed. G. V. Anrep). London: Oxford University Press.
Saksida, L. M. (1999). Effects of similarity and experience on discrimination learning: A nonassociative connectionist model of perceptual learning. Journal of Experimental Psychology: Animal Behavior Processes, 25, 308–323.
Scahill, V. L., & Mackintosh, N. J. (2004). The easy to hard effect and perceptual learning in flavor aversion conditioning. Journal of Experimental Psychology: Animal Behavior Processes, 30, 96–103.
Suret, M. B., & McLaren, I. P. L. (2003). Representation and discrimination along an artificial dimension. The Quarterly Journal of Experimental Psychology, 56B, 30–42.
Symonds, M., & Hall, G. (1995). Perceptual learning in flavor aversion learning: Roles of stimulus comparison and latent inhibition of common elements. Learning and Motivation, 26, 203–219.
Author note
The experiments presented here were conducted as part of M.C.S.’s dissertation and were supported by a predoctoral fellowship from the Universidad del País Vasco (UPV/EHU) awarded to her, as well as by Grant PI97/35, from the Gobierno Vasco, and Grant PB98-0230, from the Ministerio de Ciencia y Tecnología de España, awarded to G.A. Preparation of the manuscript was supported by Grant PSI2011-2431 from the Ministerio Español de Economía y Competitividad and Grant IT-694-13 from the Gobierno Vasco. We also express gratitude to Ana Yolanda Medina for her assistance, despite her untimely passing.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Sanjuán, M.C., Nelson, J.B. & Alonso, G. An easy-to-hard effect after nonreinforced preexposure in a sweetness discrimination. Learn Behav 42, 209–214 (2014). https://doi.org/10.3758/s13420-014-0139-9
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13420-014-0139-9