
Abstract
Motion is an important factor in visual information processing. Studies have shown that global optic flow guides attention, but it remains unclear whether this attentional guidance occurs regardless of top-down attentional control settings for another endogenous cue. To address this issue, we developed a visual search paradigm in which a task-irrelevant optic flow starts and stops prior to a visual search task itself. Participants first observed an initial optic flow motion pattern for a brief period; next, they searched a static display for a target amongst multiple distractors. Results showed faster target detection when a target’s locus coincided with the implied focus of expansion (FOE) from the preceding optic flow (vs. other loci). Eye-movement analyses revealed that initial saccades were drawn to the FOE during optic flow exposures and that relatively few saccades were needed to find targets contingent to the preceding FOE. The advantage of FOE for finding target occurred even when a salient feature singleton captured attention or when a task-relevant feature singleton was prioritized. Results of six experiments suggest that attentional control settings for a feature singleton do not over-ride a sustained influence of optic flow on attentional guidance.
Similar content being viewed by others


Humans are surrounded by enormous amounts of information in our visual world. Since the visual system operates with a limited capacity, humans selectively process the information relevant to a current behavioral goal. Visual attention is known to selectively engage relevant information in the visual world. Although mechanisms of visual attention are well documented for static visual scenes, they have not been fully examined for dynamic scenes. When we move, the visual world shifts dynamically depending on our movements. Typically, our goal when we move about in this world is to arrive at a destination, so it is critical to know whether these changing experiences of motion are consistent with this destination. Optic flow is a global dynamic pattern of apparent motion in a visual scene caused by the relative motion between a moving observer and an environmental scene. It is recognized as a source of information that affords perception of one’s heading direction (Gibson, 1950; Warren, Morris, & Kalish, 1988). For example, when we move forward, an expanding optic flow occurs and the focus of expansion (FOE) of optic flow indicates our heading direction. Therefore, it is reasonable to hypothesize that attention is directed to the FOE of optic flow in the visual field as we move to the destination.
It has been shown that motion is one of the factors that can guide visual attention in a bottom-up manner. For example, in visual search tasks, target detection is facilitated when the target is moving relative to when it is not moving (Abrams & Christ, 2003; Al-Aidroos, Guo, & Pratt, 2010; Franconeri & Simons, 2003; Hillstrom & Yantis, 1994; Takeuchi, 1997). Various types of motion, such as scintillation, revolving dots, streaming texture, oscillation, and looming, are shown to capture attention (Hillstrom & Yantis, 1994), although the strength of attentional capture can vary depending on the motion type, e.g., jitter motion, looming, receding, and so on (Franconeri & Simons, 2003; Takeuchi, 1997).
Others have shown that not only do local motions (motions of single object) impact attention, but so too do global motion patterns (motion spreading throughout the visual field). Global motions are evident, for instance, in optic flow, where changing whole flow patterns appear to guide attention (Kawahara, Yanase, & Kitazaki, 2012; Shirai & Imura, 2016; von Mühlenen & Lleras, 2007; Wang, Fukuchi, Koch, & Tsuchiya, 2012; Watanabe, 2001). von Mühlenen and Lleras (2007) have shown that coherent looming motion presented on the left or right hemi-field of the display guides attention, relative to random motion on the other hemi-field of the display. In their study, the participants’ task was to detect the onset of a target appearing at either the left or right hemi-field of the display. The looming and random motion started prior to the target presentation, and continued until participants’ response to the target. Results showed that target detection was faster when the target appeared following the looming-motion hemi-field than when it followed the presentation of a random-motion hemi-field. In contrast, target detection was not facilitated when the target was presented following a receding-motion hemi-field. Therefore, the authors concluded that looming motion plays a unique role in guiding spatial attention.
However, it seems that motion does not always attract attention in a bottom-up manner. Some studies suggest that top-down attentional control settings over-ride attentional shift caused by motion. The attentional control settings are defined as high-level and task-induced attentional processes for the endogenous cue relevant to the behavioral goals (Folk, Remington, & Johnston, 1992; Folk, Remington, & Wright, 1994). The study has shown that a moving object attracts attention when it is relevant to target locations in visual search display, but it does not draw attention when it is irrelevant to target locations (Hillstrom & Yantis, 1994). Similarly, the study of global optic flow also suggests that attentional control settings for the other endogenous cue eliminates attentional guidance induced by optic flow (von Mühlenen & Lleras, 2007, Experiment 6). Using a task that requires detecting a target at either the random-motion or looming-motion hemi-field, the authors showed that the cueing advantage for the looming motion disappeared when attention was drawn to the target location by a task-relevant arrow (endogenous cue) that always pointed to the side where the target was going to appear. This finding is important, however, because von Mühlenen and Lleras (2007) focused on the facilitation observed in initial target detection. Specifically, they examined this facilitation in relation to the early process of attention where it remains unclear whether attentional guidance, presumably associated with global motion, also disappears in the later process after attentional shift by an endogenous cue. Attention may be directed toward the location pointed to an endogenous cue in the early period, but global motion might later guide attention when this cue turns out to be invalid. In fact, Schreij, Los, Theeuwes, Enns, and Olivers (2014) demonstrated that the abrupt onset at the search display captures attention following capture by the color pre-cue. The task used in von Mühlenen and Lleras (2007) is insufficient for investigating such a “second attentional capture,” because this task focuses on the initial shift of attention. A visual search task is a suitable candidate, because this task sometimes involves the multiple attentional shifts after directing attention to the location indicated by the other cue (Schreij et al., 2014).
Accordingly, to address the issue of whether attentional control settings over-ride attentional guidance induced by optic flow, it is useful to combine the optic flow with a visual search task, rather than with von Mühlenen and Lleras (2007)’s task. However, if an optic flow pattern occurs simultaneously with a target search, this could decrease the visibility of search display containing distractors and a target. Therefore, we developed a visual search paradigm in which a task-irrelevant optic flow precedes the onset of a search display. That is, it supplies a dynamic visual context that starts and stops prior to the search display, hence permitting a test of the lingering impact of this optic flow context upon an individual’s later search performance. The question posed here, then, concerns whether or not the preceding optic flow “shapes” the guidance of future attending after the cessation of the flow pattern. This question is closely related to some situations in our real world. For example, when we stop our car on a road at a traffic signal or a stop sign, the optical flow disappears. Subsequently, when we are in the motionless car, people or bikes crossing the road may capture our attention. In this type of situation, does the optic flow that has already stopped still affect attentional guidance?
The current study uses this visual search task to explore three issues not fully addressed in previous studies. First, we investigate whether an optic flow pattern that has already ceased continues to guide attention. One study has revealed that the continuous optic flow does engage attention (von Mühlenen & Lleras, 2007); nevertheless, it remains unclear as to whether attentional guidance persists following the cessation of a preceding optic flow stimulation. Optic flow appears to guide attention in synchrony as long as the flow pattern is visible, but it may not guide attention after the offset of motion. Perhaps people immediately reorient their attention when the optic flow disappears. Alternatively, perhaps the impact of optic flow continues to guide attention even in the subsequent display where this motion has disappeared, since it is known that expanding (or contracting) optic flow affords information for perceiving a particular heading direction when people are moving about in a static visual scene. Moreover, even when we stop moving, the prevailing scene structure and the heading direction do not change, hence it would be adaptive to retain the heading direction in order to restart the movement in the visual environment, while stopping. That is, high attentional priority for the heading direction may be retained while stopping the global optic flow.
The second issue to be clarified concerns how optic flow may guide attention following the offset of a dynamic pattern. Does the guidance of attention by a preceding optic flow pattern occur regardless of participants’ attentional control settings? Some studies have shown that behavioral goals restrict stimulus-driven attentional capture; that is, stimulus-driven attentional capture is more evident when a task-irrelevant distractor shares a feature relevant to current task demands than when it does not contain a task-relevant feature (Becker, Folk, & Remington, 2010; Folk & Annett, 1994; Folk et al., 1992, 1994). Thus, Folk et al. (1994) employed an experimental setting involving a local motion cue, i.e., small circles move and stop before visual search. They found that such a motion cue does not affect search performance when attention is directed to another feature singleton like color or onset. This implies it may be possible that attentional control settings can over-ride the attentional guidance, caused by the persisting influence of a prior motion, in search of a static display.
The third issue concerns how optic flow, as well as other features, determines priority during attending when participants’ attentional control settings are varied. More specifically, we examine whether effects of optic flow are eliminated in a subsequent target search of a display, when display features such as target saliency or task relevance are manipulated. Eye-movement data may be useful for investigating these effects (Schreij et al., 2014). If the attentional control settings for a particular feature in a visual display completely cancel out the attentional guidance of optic flow, an individual’s eyes would always shift to this more salient feature, and thus would not be driven by optic flow. However, if optic flow continues to guide attention, a multiple-step pattern of eye movements would reflect how both the optic flow pattern and other features figure into determining the attentional priority. In this case, for example, eyes that instantly shift to another feature might then shift to the next location to which the optic flow has guided.
In six experiments, we used the following visual search task. Participants first observed an optic flow pattern; then when this motion stopped, they searched for a target amongst a static visual display with multiple distractors. Based on the findings in Franconeri and Simons (2003), we expected that an expanding motion pattern would guide a viewer’s attention toward the FOE. If the implicated FOE continues to attract attention in the search task (i.e., following offset of optic flow), then reaction times (RTs) to targets in this task are predicted to be faster when a target appears at the implied FOE than when it appears at other locations. In subsequent experiments, we presented color singleton in the static visual search display in order to test whether attentional control settings for a feature singleton over-ride attentional guidance instilled by the previous optic flow pattern. The color singleton was either task-irrelevant (Experiment 2) or task-relevant (Experiments 3–6) to show the effect of attentional control settings. In addition, we measured eye movements for the optic flow and visual search displays to investigate the overt attentional process. These investigations would help us understand how attentional guidance of global motion surrounding us interacts with top-down and bottom-up visual information processing.
Experiment 1
The first experiment examined whether task-irrelevant global optic flow guides attention even after the optic flow has ceased in a visual search paradigm. Participants were asked to find a target, i.e., a Gabor patch with an orientation different from distractors, and to report the orientation of the grating of the patch. Thus, task-relevant search information concerns patch orientation whereas task-irrelevant information involves the optic flow pattern, which was presented for 1, 3, or 5 s. Following this presentation, a static visual search display was immediately presented. We expected that the task-irrelevant information based on visible dot movements of global optic flow would guide attention toward the FOE. Moreover, if this optic flow instills a persisting continuation of attention after the offset of this initial display, then RTs in the following (static) search display should be faster when the target appears at the FOE of original optic flow than when it appears at other locations. In addition, a viewer’s eyes should move to FOE, reflecting its high priority.
Methods
This study was approved by the institutional review boards of Kyoto University (protocol number: 25-P-28, title: Studies for development of driving support system by understanding of implicit cognitive mechanisms).
Participants
We conducted a pilot experiment (N = 5) and estimated sample size by a power analysis using G Power version 3.1.9.2 (Faul, Erdfelder, Lang, & Buchner, 2007) with alpha = .05 and power = .95 for the result of the pilot experiment (\( {\upeta}_{\mathrm{p}}^2 \) = .586, f = 1.19). Although the estimated sample size was N = 5, studies on motion and attention using a visual search task have usually recruited more than ten participants (e.g., Abrams & Christ, 2003; Franconeri & Simons, 2003; Hillstrom & Yantis, 1994). We also considered the calibration error in eye recording, and determined the sample size as N = 18. Eighteen students attending Kyoto University (three women and 15 men, mean age = 21.8 years, SD = 1.68) participated in Experiment 1. All participants had normal or corrected-to-normal vision and were naïve concerning the purpose of the experiment. Written informed consent was obtained from all participants prior to participation in the experiment. The experiment lasted about 90 min. The participants received JPN\ 1,500 (approximately equal to US$15).
Apparatus
Stimulus presentation was controlled by MATLAB (MathWorks, Inc., Natick, MA, USA) with Psychtoolbox (Brainard, 1997; Pelli, 1997). The visual stimuli were presented on a 19-in. CRT monitor with a resolution of 1,024 × 768 pixels and a refresh rate of 60 Hz. The position of the participant’s head was fixed with a chin rest; the visual distance of the head from the CRT was 75 cm. Responses were made on a standard computer keyboard. Eye movements were recorded using an eye tracker (Eyelink1000, SR Research) with a sampling rate of 500 Hz. We conducted a 9-point eye movement calibration and validation prior to the start of the experiment and after breaks.
Stimuli
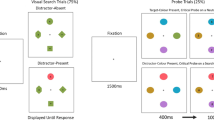
Figure 1 shows example displays. All stimuli were presented on a gray background. The optic flow display consisted of light-gray location placeholders and moving white dots. Eight location placeholders were distributed evenly around the circumferences of circle with a diameter 10.4° of visual angle. The size of each placeholder was 2.1°. We randomly located white dots with a density of 0.35 dot/deg2. The dot area fully covered the whole display and approximately 190 dots were presented. Individual dot size was 0.2°. One of the placeholders was chosen for the FOE of optic flow, and each dot moved every 16.7 ms (60 frames/s) along a radial trajectory from the center of the placeholder. Dot speed was 6.0 deg/s at the focus of radial optic flow and 2% increased in accordance with the distance, d, from the FOE toward the periphery (deg/s = 6.0 + 0.02d). In each frame, 5% of the dots were randomly chosen and re-plotted at a random location on the display. Similarly, the dots that moved out of the display were re-plotted at a random location. Due to this manipulation, dots never gathered around the FOE, and the dot density was approximately equal throughout the display.

Examples of optic flow display (left) and visual search display (right). The movie is available on the author’s website. http://yokohiguchi.net/stimuli/. FOE focus of expansion
The visual search display consisted of location placeholders, static white dots, and Gabor patches for a target and seven distractors. The location placeholders were identical to the optic flow display and Gabor patches appeared inside the placeholders. The white dots continued to be presented at the same location with the last frame of the optic flow display. The cycle/deg of each Gabor was 5.3, and Gaussian standard deviation was 0.3° of visual angle. The target was a Gabor patch that rotated 45° either to the right or to the left of the vertical axis. The distractors were vertical Gabor patches.
Design and procedure
The task was to find the target, with the Gabor patch rotated either to the right or to the left; the viewer had to report the direction of the target rotation as quickly and accurately as possible by pressing a right or left arrow key with the corresponding finger (middle or index finger) of the right hand. Figure 2 shows a trial sequence. A fixation cross (0.5° × 0.5°) appeared at the center of the display, and the optic flow display was presented 1,000 ms after the fixation cross. The optic flow display was presented for 1, 3, or 5 s. After the optic flow display, the visual search display appeared. The static dots in the visual search display were presented at the same locations as in the last frame of the optic flow display; thus, this display looked like the visual search started when the moving dots stopped. RTs for the target were calculated from the onset of the visual search display. The visual search display remained visible until the participants’ response. Feedback displays and a low-pitched buzz were provided only for incorrect responses.

The experiment began with ten practice trials, followed by a 384-trial search task. Participants were able to take a break every 64 trials. A two-factor within-participants design was used. Independent variables were target location condition and optic flow duration. Location condition had two levels, match and non-match. In the match condition, the target appeared at the location placeholder determined as the FOE of optic flow. In the non-match condition, the target appeared at the different placeholder from the FOE of optic flow. To rule out location probability effects, the target and FOE appeared equally often at eight possible location placeholders. Therefore, the number of the trials was 48 and 336 for the match and non-match conditions, respectively. The second independent variable, optic flow duration, had three levels, 1, 3, and 5 s for the exposure duration of the optic flow display. For each optic flow duration, the number of trials was 16 in the match condition, and 112 in the non-match condition. These trials of different conditions were presented in a random order. We randomly determined the direction of the target rotation for each trial, but each condition contained an equal number of trials for the right and left directions.
Results and discussion
RT data in the visual search display
We excluded incorrect trials (0.78% of the total data) and trials with RTs outside 3 SD of the mean in each condition (0.59% of the total data).
Figure 3 shows the RT results. A repeated-measures analysis of variance (ANOVA) was conducted with the factors of the target location (match or non-match) and optic flow duration (1, 3, or 5 s). An ANOVA on the RTs showed a significant main effect of the location condition, F (1, 17) = 21.41, p < .001, \( {\upeta}_{\mathrm{p}}^2 \) = .557, indicating that RTs were faster when the target was presented at the FOE of optic flow (match condition) than when the target appeared at a different location from the FOE (non-match condition). The marginal RT means (the mean for one factor averaged across all levels of the other factor) were 1,174 ms and 1,461 ms for the match and non-match conditions, respectively. Thus, this result suggests that the task-irrelevant optic flow guides attention to the FOE of optic flow. The main effect of optic flow duration was also significant, F (2, 34) = 6.33, p = .005, \( {\upeta}_{\mathrm{p}}^2 \) = .271. The marginal RT means were 1,297 ms, 1,292 ms, and 1,364 ms in each of the 1-, 3-, and 5-s duration conditions, respectively. The interaction of location condition and optic flow duration was also significant, F (2, 34) = 10.81, p < .001, \( {\upeta}_{\mathrm{p}}^2 \) = .388. A simple effect of target location was significant for all the optic flow durations, 1s: F (1, 17) = 24.96, p < .001, \( {\upeta}_{\mathrm{p}}^2 \) = .595; 3s: F (1, 17) = 28.10, p < .001, \( {\upeta}_{\mathrm{p}}^2 \) = .623; 5s: F (1, 17) = 7.59, p = .014, \( {\upeta}_{\mathrm{p}}^2 \) = .309. These results suggest that attention was guided to the FOE regardless of the optic flow duration.

The reaction times in Experiment 1 as a function of optic flow duration, shown for match and non-match target location conditions. Error bars indicate standard errors. Asterisks indicate significant differences at *p < .05 and **p < .001
Eye-movement data in the optic flow display
We detected saccades and fixations using the saccade detection algorithm supplied by SR Research. If attention is promptly guided by the optic flow, the initial saccades may be directed to the FOE of optic flow in the optic flow display. Therefore, we focused on initial saccades in the optic flow display and calculated the angular difference between the initial saccade and the mathematically straight line to the FOE from the initial fixation point (Fig. 4). Given that initial saccades were directly sent to the FOE, the angular difference between the second fixation and the straight line to the FOE, measured from the location of the initial fixation, should be close to 0°. Figure 5 summarizes the angular difference for all participants. As can be seen in Fig. 5 (the left panel), the mode of the distribution was located in the 0° to 10° bin (mode = 0.0007), indicating that initial saccades were most often directed to the FOE of optic flow. This result shows that attention is initially guided toward the FOE of optic flow when the optic flow display appears.

We calculated the angular difference θ between the initial saccade and the mathematically straight direction to the focus of expansion (FOE) from the initial fixation

Eye-movement data in the visual search display
As shown in the RT data, responses were faster when the target appeared at the FOE of optic flow than when the target appeared at other locations. Therefore, we expected that, in the visual search display, the placeholder presented at FOE would be more frequently fixated first, thus preceding other placeholders. For the investigation of initial fixation to the FOE, we defined an area of each placeholder as a circle with a radius of 2.1° of visual angle around the center, considering 1.1° for the radius of the placeholder and 1.0° for the calibration error in eye recording. Since we were interested in which of the placeholders was fixated first, in cases where the initial fixation went to the non-placeholder locations, such as around the fixation cross, we looked at the second or later fixations that landed on any of the placeholders. Table 1 shows the probability that participants initially looked at the FOE or other (non-FOE) placeholders. An ANOVA on probability of looking at the FOE with a factor of optic flow duration (26.4%, 24.2%, and 20.4% for the 1-, 3-, and 5-s optic flow duration, respectively) revealed that the probabilities were different between the optic flow duration conditions, F (2, 34) = 9.55, p < .001, \( {\upeta}_{\mathrm{p}}^2 \) = .360. A multiple comparison revealed that the probability that participants looked at the FOE was higher in the 1- and 3-s optic flow durations than in the 5-s optic flow duration, t (17) = 4.52, p < .001 and t (17) = 3.32, p = .004. There was no difference between the 1- and 3-s optic flow durations, t (17) = 1.34, p = .198. We also compared the probability with chance levels (12.5%) separately for each condition. The probability was significantly higher than the chance level in all the conditions, 1 s: t (17) = 8.10, p < .001, d = 1.91, 3 s: t (17) = 7.76, p < .001, d = 1.82, 5s: t (17) = 5.85, p < .001, d = 1.37. Participants initially looked at the FOE, and that optic flow guided attention more to the FOE in the 1- and 3-s optic flow duration than in the 5-s optic flow duration.
Experiment 2
Results of Experiment 1 showed that the task-irrelevant optic flow continued to guide attention toward the FOE even after the offset of this flow. Therefore, in subsequent experiments, we sought to further investigate whether task-irrelevant optic flow guides attention under different conditions manipulating physical salience or current selection goals (i.e., attentional control settings). In Experiment 2, a placeholder with task-irrelevant color (in green) was presented in the visual search display. Although the color singleton did not correspond to (hence cue) a future target’s location, according to theories of stimulus-driven attention this singleton should draw attention from any concurrently presented with a target (Theeuwes, 1991). We tested whether optic flow that has already stopped still guides attention to the FOE when the salient feature singleton captures attention in the visual search display.
Methods
Eighteen students who did not participate in Experiment 1 were recruited (seven women and 11 men, mean age = 21.8 years, SD = 1.59). The apparatus, stimuli, and procedure were identical to those in Experiment 1 with the exception below.
We only used a 1-s duration for the optic flow display since we confirmed that 1-s exposure was sufficient for guiding attention. Importantly, one of the placeholders was colored green in the visual search display (see Fig. 2). The target, FOE, and color placeholders appeared equally often at eight possible location placeholders. There were four conditions: both-match, FOE-match, color-match, and non-match conditions. In the both-match condition, both the FOE of optic flow and color placeholder were the same location as the target. In the FOE-match condition, the FOE occupied the same location as the target, but the color placeholder appeared at a different location. In the color-match condition, the color placeholder was presented at the same location with the target. In the non-match condition, neither the FOE nor color placeholder was presented at the target location. The numbers of trials were eight, 56, 56, and 392 for the both-match, FOE-match, color-match, and non-match conditions, respectively. Participants completed 512 trials in total.
Results and discussions
RT data in the visual search display
We excluded incorrect trials (0.67% of the total data) and trials with RTs outside 3 SD of the mean in each condition (0.74% of the total data).
Figure 6 shows RT results of Experiment 2. An ANOVA on the RTs with the factor location condition (both-match, FOE-match, color-match, and non-match) showed a significant main effect of location condition, F (3, 51) = 17.82, p < .001, \( {\upeta}_{\mathrm{p}}^2 \) = .512. A multiple comparison using Shaffer’s method (Shaffer1; Holland & Copenhaver, 1988; Shaffer, 1986; alpha level is modified to be 0.05) revealed that RTs in the both-match, FOE-match, and color-match conditions were faster than RTs in the non-match condition, t (17) = 5.44, t (17) = 5.56, and t (17) = 4.58 (all ps < .001). The difference between the FOE-match and non-match conditions suggests that optic flow guided attention toward the FOE even when the color placeholder captured attention in the visual search display. In addition, RTs in the both-match condition was faster than RTs in the color-match condition, t (17) = 2.96, p = .027, indicating the additive effects of optic flow and color singleton. There were no differences between the both-match and FOE-match conditions, t (17) = 1.05, p = .307, and between the FOE-match and color-match conditions, t (17) = 2.00, p = .124.

The reaction times in Experiment 2. Error bars indicate standard errors. Asterisks indicate significant differences at *p < .05 and **p < .001
Eye-movement data in the optic flow display
As in Experiment 1, we calculated the angular difference between the initial saccade and the straight line to the FOE from the initial fixation. As evident in Fig. 5 (the center panel), mode of the distribution was located in the 0° to 10° bin (mode = 0.0032), indicating that the task-irrelevant optic flow influenced initial saccades in the optic flow display, even when the colored placeholder captured attention in the visual search display.
Eye-movement data in the visual search display
Table 1 shows the probability that participants initially looked at the FOE, color, and other placeholders. Here, to investigate the effect of color and FOE separately, we excluded the trials in which both the FOE and color placeholders were presented at the same location. We compared the probability that participants looked at the color placeholder (38.6%) and the FOE placeholder (9.2%) with chance levels (12.5%). The probability that the color placeholder was initially fixated was significantly higher than chance level, t (17) = 7.40, p < .001, d = 1.74. In contrast, the probability that the FOE was initially fixated was significantly lower than chance level, t (17) = 5.14, p < .001, d = 1.21. This result, unlike Experiment 1, indicates that participants initially looked at the color placeholder rather than the FOE. However, as shown in RT data, attention continues to be oriented toward the FOE after the initial fixation to the color placeholder.
Experiment 3
In Experiment 2, the optic flow guided attention toward the FOE even when the task-irrelevant color singleton captured attention in the visual search display. In Experiment 3, we examined whether bias toward FOE is observed even when participants have attentional control settings for an endogenous cue in a search display. In this experiment, a color placeholder is likely to appear at the same location with the target in the visual search display. Participants were able to use the color singleton as a cue for finding a target quickly. Therefore, not only the bottom-up salience but also the top-down control settings for the cue (modulated by the statistical probability) should influence attentional process.
Methods
Eighteen students who did not participate in the previous experiments were recruited (four women and 14 men, mean age =21.5 years, SD = 2.23). The apparatus, stimuli, and procedure were identical to those in Experiment 2 with the exception below.
The color placeholder was associated with the target location in 448 trials out of 576 trials (77.8% of all trials), thus participants were able to use the color singleton for searching a target. The number of trials for the both-match, FOE-match, color-match, and non-match conditions were 56, 16, 392, and 112, respectively.
Results and discussion
RT data in the visual search display
We excluded incorrect trials (0.66% of the total data) and trials with RTs outside 3 SD of the mean in each condition (1.31% of the total data).
Figure 7 shows the RT results of Experiment 3. An ANOVA on the RTs with the factor location condition (both-match, FOE-match, color-match, and non-match) showed a significant main effect of location condition, F (3, 51) = 135.20, p < .001, \( {\upeta}_{\mathrm{p}}^2 \) = .888. A multiple comparison using Shaffer’s method (alpha level is modified to be 0.05) revealed that RTs in the both-match, FOE-match, and color-match conditions were faster than RTs in the non-match condition, t (17) = 19.28, t (17) = 7.05, and t (17) = 20.59 (all ps < .001). RTs in the both-match and color-match condition were reliably faster than RTs in the FOE-match condition, t (17) = 8.70 and t (17) = 6.04 (ps < .001). In addition, RTs in the both-match condition were faster than RTs in the color-match condition, t (17) = 5.36, p < .001. These results suggest that the optic flow still guided attention toward the FOE even when the color singleton strongly captured attention. We also found that cueing effects of color and FOE are additive, suggesting that the two forms of attentional guidance operate independently.

The reaction times in Experiment 3. Error bars indicate standard errors. Asterisks indicate significant differences at **p < .001
Eye-movement data in the optic flow display
Consistent with the analyses in the previous experiments, we calculated the angular difference between the initial saccade and the straight line to the FOE from the initial fixation (Fig. 5, the right panel). Mode of the distribution was located in the 0° to 10° bin (mode = 0.0014). This result indicates that the task-irrelevant optic flow still influenced initial saccades even when the color placeholder strongly captured attention in the visual search display.
Eye-movement data in the visual search display
Table 1 shows the probability that participants initially looked at the color, FOE, or other placeholders. To investigate the effect of color and FOE separately, we excluded the trials in which both the FOE and color placeholder were presented at the same location. The probability that the color placeholder was initially fixated was high (85.0%). Since we found that participants initially looked at the color placeholder in most of the trials, we further investigated which of the placeholders was fixated next after the color placeholder when the target was not presented at the color placeholder (color-invalid trials). Participants sometimes made multiple fixations to the color placeholder, but we were interested in which of the other placeholders was fixated after a color placeholder fixation. Therefore, we looked at later fixations that moved from the color placeholder in the cases where multiple fixations occurred for the color placeholder. We found that, when the color placeholder was invalid, the second fixation then went to the FOE of optic flow in 17.8% (SD = 4.98) of trials (Fig. 8). This probability was significantly higher than the chance levels (14.3%), t (17) = 3.02, p = .008, d = 0.711, suggesting that the FOE of optic flow retained cueing attention after the initial fixation. The participants looked at all other (non-FOE) placeholders after the color placeholder in 79.5% (SD = 4.98) of trials. There were few cases in which participants made the response before looking at the target (2.7% of trials). These trials included both correct and incorrect responses, and a few cases of calibration errors in eye recording.

Probabilities (%) of initial and second fixations to each location: color, focus of expansion (FOE) (focus of contraction [FOC] or onset cue), and other placeholders. We excluded the trials in which both the FOE (FOC or onset cue) and color placeholder were presented at the same location. Participants initially looked at the color placeholder in most of the trials. When the target was not presented at the color placeholder (color-invalid trials), the second fixation then went to the FOE (FOC or onset cue). For the sake of simplicity, Fig. 8 does not illustrate the second fixation following initial fixation to other placeholders. There were a few cases in which participants made a response before looking at the target (“response” in the figure). These trials included both correct and incorrect responses, and a few cases of calibration errors in eye recording
Taken together, we found bias toward the FOE of optic flow even when participants have attentional control settings for a feature singleton in a search display. Participants were likely to look at the color placeholder first, but when the color cue turned out to be invalid, they then looked at the FOE. These results show that attention is guided toward the FOE after the initial fixation is directed to the color placeholder in a top-down control manner. In other words, attentional guidance to FOE survives when participants have attentional control settings for other features.
Experiment 4
Results of Experiment 3 revealed that the task-irrelevant optic flow guided attention toward the FOE even when the presence of a single colored placeholder predicted a different target location. That is, when participants had attentional control settings for a feature other than optic flow, which should capture attention, attention was nonetheless attracted to the FOE locus. It is possible that this FOE bias is attributable to eye movements during the optic flow display where participants were able to move their eyes freely. Perhaps eye position hovered near the FOE during the optic flow display thus leading to facilitation of responses whenever a target appeared at the FOE. To prevent the effect of eye position during the exposure to the optic flow for searching a target in the search display, in Experiment 4, we instructed participants to maintain fixation on the center of the display during the optic flow display. The other aspects of procedure were kept the same as Experiment 3.
Methods
Twenty-four students who did not participate in the previous experiments were recruited. We excluded six participants who moved their eyes 3° away from the fixation cross before the onset of visual search display in more than 10% of trials, and analyzed the data of the remaining 18 participants (seven women and 11 men, mean age = 21.3 years, SD = 1.91). The apparatus, stimuli, and procedure were identical to those in Experiment 3 with the exception that the fixation cross appeared in the optic flow display to help maintain fixations to the center of the display. The experimenter instructed participants to keep looking at the fixation cross during the optic flow display. The participants were able to move their eyes when the fixation cross disappeared and the visual search started.
Results and discussions
RT data in the visual search display
We excluded incorrect trials (0.87% of the total data) as well as trials with RTs outside 3 SD of the mean in each condition (1.05% of the total data). In addition, we removed trials when participants moved their eyes more than 3° away from the fixation cross (2.97% of the total data).
Figure 9 shows the RT result in Experiment 4. An ANOVA on the RTs with the factor location condition (both-match, FOE-match, color-match, and non-match) showed a significant main effect of location condition, F(3, 51) = 107.88, p < .001, \( {\upeta}_{\mathrm{p}}^2 \) = .864 A multiple comparison using Shaffer’s method (alpha level is modified to be 0.05) revealed that RTs in the both-match, FOE-match, and color-match conditions were faster than RTs in the non-match condition, t(17) = 10.88, t(17) = 6.51, and t(17) = 10.98 (all ps < .001). RTs in the both-match and color-match condition were faster than RTs in the FOE-match condition, t (17) = 9.87 and t (17) = 9.93 (ps < .001). There was no difference between both-match and color-match conditions, t (17) = 1.12, p = .278. These results suggest that the optic flow guides attention toward the FOE even when eye movements to the FOE are prohibited during the exposure to the optic flow. The difference between the both-match and color-match conditions, observed in Experiments 2 and 3, disappeared here. This suggests that eye movements to the FOE may have caused the additive cueing effect of FOE and color singleton.

The reaction times in Experiment 4. Error bars indicate standard errors. Asterisks indicate significant differences at **p < .001
Eye-movement data in the visual search display
We instructed participants not to make any eye movements in the optic flow display, and removed trials in which participants moved their eyes more than 3° away from the fixation cross. Therefore, in the current and later experiments, we focused only on the eye-movement data in the visual search display.
As evident in Table 1, participants initially looked at the color placeholder in the visual search display in most of the trials (96.6%). As in Experiment 3, we investigated which placeholder was fixated after the initial fixation to the color placeholder when the target did not appear at the color placeholder (Fig. 8). We found that, when the color placeholder was invalid, fixations went to the FOE in 24.5% of trials (SD = 5.13); the probability of this was significantly greater than chance (14.3%), t (17) = 8.45, p < .001, d = 1.99. This result suggests that the FOE of optic flow still guided attention after the initial fixation to the color placeholder. The participants looked at the overall other placeholder after the color placeholder in 74.6% of trials (SD = 4.99). There were a few cases in which participants made the response before looking at the target (0.9% of trials).
Thus, the results in Experiment 4 showed that FOE bias was not due to the eye position during the optic flow display. Even when the eye movements were prohibited during the exposure to the optic flow, attention is still guided toward the FOE.
Experiment 5
In the previous experiments, we demonstrated that expanding visual motion guides attention toward an FOE regardless of attentional control settings. The purpose of Experiment 5 was to investigate whether this finding can be extended to optical flow contexts characterized by contracting motion. Some studies have demonstrated the difference between expanding and contracting visual motion patterns in guiding attention (Franconeri & Simons, 2003; Takeuchi, 1997; von Mühlenen & Lleras, 2007). For example, von Mühlenen and Lleras (2007) showed that expanding motion attracts attention while contracting motion does not draw attention, suggesting that expansion plays a unique role in guiding attention. In Experiment 5, we tested whether the contracting motion guides attention toward the focus of contraction (FOC) in a similar manner to the expanding motion.
Methods
Eighteen students who did not participate in the previous experiments were recruited (four women and 14 men, mean age = 19.9 years, SD = 1.68). No participant moved their eyes 3° away from the fixation cross before the onset of visual search display in more than 10% of trials, so we analyzed all the data.
The apparatus, stimuli, and procedure were identical to those in Experiment 4 with the following exception. The contracting motion was presented instead of the expanding motion. We first randomly located white dots with a density of 0.35 dot/deg2, and one of the placeholders was chosen for the FOC of optic flow. Each dot moved every 10 ms along a radial trajectory to the center of the placeholder. Dot speed was 6.0 deg/s at the focus of radial optic flow and 2% decreased in accordance with the distance d from the FOC toward the periphery (deg/s = 6.0 - 0.02d). In each frame, 5% of the dots were randomly chosen and re-plotted at the random location on the display. Similarly, the dots that moved into the FOC were re-plotted at the random location.
Results and discussions
RT data in the visual search display
We excluded incorrect trials (0.91% of the total data) and trials with RTs outside 3 SD of the mean in each condition (1.23% of the total data). In addition, we removed trials in which participants moved their eyes more than 3° away from the fixation cross (2.31% of the total data).
Figure 10 shows the RT result in Experiment 5. An ANOVA on the RTs with the factor location condition (both-match, FOC-match, color-match, and non-match) showed a significant main effect of location condition, F (3, 51) = 324.07, p < .001, \( {\upeta}_{\mathrm{p}}^2 \) = .950. A multiple comparison using Shaffer’s method (alpha level is modified to be 0.05) revealed that RTs in the both-match, FOC-match, and color-match conditions were faster than RTs in the non-match condition, t(17) = 20.03, t(17) = 5.03, and t(17) = 20.88 (all ps < .001). Also, the RTs in the both-match and color-match condition were faster than RTs in the FOC-match condition, t (17) = 17.91 and t (17) = 18.71 (ps < .001). There was no difference between the both-match and color-match conditions, t (17) = 0.87, p = .398. These results suggest that the optic flow guides attention toward.

The reaction times in Experiment 5. Error bars indicate standard errors. Asterisks indicate significant differences at **p < .001
Eye-movement data in the visual search display
We excluded trials in which participants’ eye movement exceeded 3° from the fixation cross (2.31% of the total data). Consistent with the previous experiments, participants initially looked at the color placeholder in the visual search display in most of the trials (93.0%, Table 1). We found that, when the color placeholder was invalid, the fixations went to the FOC of optic flow in 30.9% of trials (SD = 10.9, Fig. 8); moreover, this probability was significantly higher than chance level (14.3%), t (17) = 6.47, p < .001, d = 1.52. These results suggest that the FOC of optic flow attracts attention after the initial fixation to the color placeholder. Participants looked at the other placeholder after the color placeholder in 68.0% of trials (SD = 10.6). There were few cases where participants made the response before looking at the target (1.6% of trials).
These results suggest that attention is guided toward the FOC of optic flow even when the color singleton captured attention. Not only the expanding motion but also the contracting motion can influence attentional process.
Experiment 6
Experiment 5 demonstrated that attention is guided not only toward the FOE of expanding motion but also toward the FOC of contracting motion regardless of attentional control settings. The remaining question is whether optic flow has a unique role in guiding attention. Is it possible that another cue, such as an abrupt onset, induces a similar effect in a visual search? The purpose of Experiment 6 was to investigate whether attention is captured by the task-irrelevant onset cue under the manipulation of attentional control settings. Studies have shown that an abrupt onset in visual display is likely to capture attention in a bottom-up process (Yantis & Jonides, 1984, 1990). However, Folk et al. (1992) showed that attentional control settings for an endogenous cue over-rides capture of an abrupt onset. Based on this finding, an abrupt onset might not attract attention when participants’ attention is directed to the task-relevant color singleton. We used an onset cue instead of the optic flow pattern, and tested whether the search performance is facilitated when the target in a static visual display appeared at the location of the abrupt onset cue compared to when the target appeared at the other locations, regardless of attentional control settings for a color singleton.
Methods
Twenty-two students who did not participate in the previous experiments were recruited. We excluded four participants who moved their eyes 3° away from the fixation cross before the onset of visual search display in more than 10% of trials, and analyzed the data of the remaining 18 participants (eight women and 10 men, mean age = 20.4 years, SD = 1.72). The apparatus, stimuli, and procedure were identical to those in Experiments 4 and 5 with the following exception: the task-irrelevant onset cue (white circle) was presented instead of optic flow (Fig. 1) prior to a visual search display. The static dot display was first presented for 500 ms; next the onset cue appeared at the locus of one placeholder. This onset cue appeared equally often at eight possible location placeholders, thus the onset cue did not predict a target location. The onset cue lasted 500 ms until the presentation of visual search display including a color singleton associated with the target location (visual search displays were the same as in Experiments 3–5). As in Experiments 4 and 5, the experimenter instructed participants to keep looking at the fixation cross until the fixation cross disappeared and visual search display started. The onset-match condition was used instead of the FOE/FOC-match conditions. In the onset-match condition, the onset cue occurred at the same location as the target, but the color placeholder appeared at a different location. In the both-match condition, both the onset cue and color placeholder were the same location as the target. The color-match and non-match conditions were the same as Experiments 3–5.
Results and discussions
RT data in the visual search display
We excluded incorrect trials (1.16% of the total data) and trials with RTs outside 3 SD of the mean in each condition (1.38% of the total data). In addition, we removed trials in which participants moved their eyes more than 3° from the fixation cross (3.43% of the total data).
Figure 11 shows RT results in Experiment 6. An ANOVA on the RTs with the factor location condition (both-match, onset-match, color-match, and non-match) showed a significant main effect of location condition, F(3, 51) = 104.37, p < .001, \( {\upeta}_{\mathrm{p}}^2 \) = .859 A multiple comparison using Shaffer’s method (alpha level is modified to be 0.05) revealed that RTs in the both-match and color-match conditions were faster than RTs in the non-match condition, t(17) = 10.59 and t(17) = 11.26 (ps < .001). RTs in the both-match and color-match condition were faster than RTs in the onset-match condition, t (17) = 10.14 and t (17) = 10.65 (ps < .001). Interestingly, RTs in the both-match condition were significantly slower than RTs in the color-match conditions, t (17) = 7.26, p < .001. There was no difference in RTs between the onset-match and the non-match condition, t (17) = 1.01, p = .327. These results suggest a difference in cueing effects between the abrupt onset and the optic flow. First, the lack of search facilitation at the location of onset cue suggests that attentional control setting for the color singleton weakens the attentional capture induced by abrupt onset. Second, the fact that RTs in the both-match condition were slower than the color-match condition suggests that response to the target was somehow impaired when the onset and color cue were presented at the same location compared to when only the color cue was presented at the target location.

The reaction times in Experiment 6. Error bars indicate standard errors. Asterisks indicate significant differences at **p < .001
Eye-movement data in the visual search display
We excluded trials in which participants moved their eyes more than 3° from the fixation cross. As shown in Table 1, participants initially looked at the color placeholder in the visual search display in most of the trials (94.7%). When the target did not appear at the color placeholder (color-invalid trials), the fixations went to the location of onset cue in 22.0% of trials (SD = 12.8, Fig. 8); the latter probability was significantly higher than chance level (14.3%), t(17) = 2.55, p = .021, d = 0.60, suggesting that the onset cue draws attention after a viewer’s initial fixation to the color placeholder. Usually, participants looked at the other placeholder after the color placeholder (i.e., in 76.1% of trials; SD = 12.9). In only a few cases did participants make a response before looking at the target (1.9% of trials). Although we did not find any advantage of onset cue in the RT data, participants tended to subsequently look at the location of onset cue after the color singleton. Thus, there is a trace of the onset cue attracting attention in eye-movement data, but individual differences seem to increase response variability, leading to a weaker effect size relative to the effect size obtained in the previous experiments of optic flow.
Although RTs in the both-match condition were slower than RTs in the color-match condition, the number of fixations did not differ between these conditions (see Appendix). Therefore, we further examined which component of eye-movement data is related to the slowed RTs in the both-match condition. As is shown in Table 2, only a single fixation was needed for finding a target in the both-match and color-match conditions; in addition, the RT difference was relatively small (39 ms). Therefore, we expected that the initial saccade latency or initial saccade duration would be longer when the color singleton was presented at the same location with the onset cue. We found that initial saccades latencies for the color singleton were longer when it appeared at the same location as the onset cue than when it appeared at a different location (265 ms vs. 222 ms), t (17) = 9.24, p < .001, d = 2.18; however, no such difference was observed in initial saccade durations (35 ms vs. 33 ms), t (17) = 1.59, p = .131, d = 0.37.
In summary, Experiment 6 provides a different profile of results than that observed in the previous experiments using optic flow. There was no beneficial effect in RTs when the target at the same locations the onset cue. In fact, the RTs slowed when the onset cue and the color singleton were presented at the same position. Eye-movement data showed a tendency to look at the location of onset cues after color singletons, but the number of fixations before finding a target did not decrease even when the target was presented at the location of onset cue. Furthermore, when the onset cue and the color singleton appeared at the same position, saccade latency was delayed. These results suggest that onset cues and optic flows have intrinsically different effects on attention, at least in the current experimental setting. Although both cues can instantly attract attention to their positions to some extent, the after-effect of cueing persists only for the optic flow cue.
General discussion
The current study aimed to investigate whether optic flow guides attention even when attention might be directed to a salient object or a task-relevant object. We first confirmed that a task-irrelevant expanding optic flow guides attention toward the FOE in a visual search task, even when the prior motion of optic flow has ceased (Experiment 1). Furthermore, when a color singleton was present in a static array, the impact of a preceding optic flow remained evident in a continued guiding of a viewer’s attention regardless of the stimulus-driven attention for the physical salience (Experiments 2) or viewer’s attentional control settings for the task-relevant cue (Experiments 3–4). We also found that not only might an expanding optic flow draw attending to an FOE, a contracting optic flow can guide attention an FOC (Experiment 5). These effects are not obtained when the task-irrelevant onset cue was presented instead of optic flow (Experiment 6), suggesting that optic flow governs attention in a different manner than does an abrupt onset.
The result of eye movement showed that if eye movements are allowed during the exposure to the optic flow, initial saccades frequently go to the direction of FOE of optic flow (Experiments 1–3). These results suggest that the motion is quickly processed and guides eye movements toward the FOE before the initial saccade occurrences (approximately 250 ms). Our data also showed that FOE/FOC of optic flow retains attentional priority even after the optic flow has ceased, under different types of manipulation of participants’ attentional control settings. We found that, even when a majority of initial fixations were directed straight to the color singleton, secondary fixations often moved toward the FOE/FOC of optic flow (Experiments 3–5). Note that, although the probability that the participants secondarily looked at the FOE/FOC was above chance, these secondary fixations to the FOE/FOC do not fully account for the fact that RTs for the target at the FOE/FOC were faster than for the target at the other locations. In some cases, a viewer’s eyes, which had been heading toward the FOE/FOC, suddenly averted to another location. This may be especially likely in the current displays where the placeholders for target and distractors were arranged in a circular configuration. After an initial fixation to the color singleton, a viewer’s eyes moved toward the FOE/FOC in a circular trajectory, but sometimes stopped somewhere along the way. Such eye-movement data suggest that, in either case, the processing of a color singleton is prioritized based on singleton saliency or its task relevance, and that attentional guidance based on optic flow appears to occur after these initial fixations to the color singleton.
Von Mühlenen and Lleras (2007) have demonstrated that continuous motion guides attention under conditions in which motion is completely task-irrelevant. In addition, we find that even if the opening visual motion has ceased, the primacy of this early motion continues to influence attending, guiding it toward the FOE/FOC of an optic flow pattern. Moreover, the important implication in the current study is that attention is able to return to the FOE/FOC of optic flow even after directing attention to other features (e.g., color singleton). A similar phenomenon is reported by Schreij et al. (2014) in a setting where color cue and abrupt onset appeared. They demonstrated that when either a color cue or an abrupt onset captures attention, the other one of this pair subsequently captures attention. Consistent with their findings, we demonstrated that the capture by color singleton did not eliminate the secondary capture by optic flow in spite of the fact that optic flow had already stopped. This finding suggests that the top-down attentional biases toward color singleton and the bottom-up attentional signals of optic flow are maintained simultaneously. As noted by Schreij et al. (2014), the current results can be accounted in terms of an activation map (priority map, Itti & Koch, 2001), which topographically codes for local potential areas of interest for allocation of attention. The activation map reflects both top-down and bottom-up components, and in the current experimental settings, the color singleton receives the highest accumulated activation, and the FOE/FOC of optic flow receives the second-highest activation. As a result, the initial fixations were drawn to the color singleton and following fixations were drawn to the FOE/FOC of the optic flow. Thus, optic flow appears to induce “attentional round-trip” to the original FOE/FOC from the color singleton.
One possible factor inducing such an attentional round-trip to the FOE/FOC can be the motion after-effect (Wohlgemuth, 1911; for a review, see Anstis, Verstraten, & Mather, 1998). The motion after-effect is a visual illusion of motion caused by prior exposure to motion. Even a brief exposure to a moving adaptation stimulus influences the perception of a subsequently presented ambiguous motion stimulus (Kanai & Verstraten, 2005). However, in considering recent findings that motion after-effect to global motion takes place at retinotopic levels of the visual cortex (Knapen, Rolfs, & Cavanagh, 2009), it is unlikely that motion after-effect affected attentional guidance to the location of FOE/FOC even after several saccades in visual search. Further investigation is required to clarify the contribution of relatively low-level factors on this attentional round-trip effect.
Another possible factor that may contribute to the attention return to the FOE/FOC of optic flow involves a visual memory effect. It has been suggested that visual perception, attention, and memory in humans is serially dependent on prior experience (serial dependence of visual processing, Fischer & Whitney, 2014; Kiyonaga, Scimeca, Bliss, & Whitney, 2017). Perceptual priming (Maljkovic & Nakayama, 1994, 1996) or contextual cueing (Chun & Jiang, 1998) can also be related phenomena in which exposure to a prior stimuli or configuration influences subsequent attentional process. The basic reasoning is that it would be adaptive to use the past information, when relevant, to search for an object; hence, the prior experience of optic flow may bias visual processing at the FOE/FOC for a while. Further work is necessary to understand the underlying mechanisms with respect to how attention can be returned to the source point of optic flow.
In line with results from a previous study (Folk et al., 1992), our data showed that attentional control settings for the color singleton over-rides attentional capture of an abrupt onset (Experiment 6). Moreover, when the onset cue and color singleton were presented at the same location, the response to a target was delayed compared to when only the color singleton was associated with the target location. Eye movement-data showed that this response delay was due to the longer saccade latencies to the location in which both the onset cue and color singleton appeared. The response delay to the originally cued location may be a phenomenon known as inhibition of return (IOR; Posner & Cohen, 1984; for a review, see Klein, 2000). Posner and Cohen (1984) demonstrated that facilitation or inhibition in the target detection occurs as a result of on stimulus-onset asynchrony (SOA) between the pre-cue and the target. That is, target detection at the pre-cued location is usually facilitated when the SOA was relatively short (approximately 150 ms). However, when the SOA increases to more than 300 ms (500 ms in Posner & Cohen, 1984), target detection accuracy decreases. The visual system discourages attention from returning to the originally attended location, perhaps thereby promoting more effective search behavior. In the current study, such an inhibitory effect was apparent in eye-movement data when the task-irrelevant onset cue was present. Possibly, the duration of the onset cue in Experiment 6 (500 ms) was too long for attention to still linger around the same location; an inhibitory tag could have been attached to the cued locations, rendering attention difficult to return to the cued locations upon the presentation of visual search display. Although IOR was not observed when the task-irrelevant optic flow was present, IOR might occur in a different SOA condition when optic flow guides attention. Because Experiments 1–5 presented the visual search display immediately after the offset of optic flow, attention to an FOE/FOC was still maintained even after the visual search display was presented. This may be due to the absence of a time-gap between the offset of optic flow and visual search displays. The current study did not manipulate the duration of optic flow, and further study is required to investigate whether optic flow induces IOR at FOE/FOC under various conditions of duration between the offset of optic flow and the onset of visual search display.
Interestingly, we found that both the expanding and the contracting optic flow guides attention toward the FOE/FOC. This finding is inconsistent with certain previous studies demonstrating the uniqueness of expanding motion (Franconeri & Simons, 2003; Rossini, 2014; Takeuchi, 1997; von Mühlenen & Lleras, 2007; Wang et al., 2012). The critical difference between the current study and the previous ones is that, in the current study, attentional guidance via optic flow persisted, occurring even following the intervening attraction of attention by other features. Such guidance might occur in a different mechanism to that responsible for the cuing of attention by expanding motion reported in the previous study.
The findings described here contribute to understanding of visual attention in moving space. Recent studies suggest that the vanishing point of scene structure attracts attention (Borji, Feng, & Lu, 2016; Ueda, Kamakura, & Saiki, 2017). Ueda et al. (2017) have demonstrated that vanishing points are likely to attract attention throughout a trial in a visual search; even after an initial fixation to a salient stimulus, eye movements converged on vanishing point. Not only the vanishing point but also the optic flow provides the information of scene structure, e.g., a road, is often apparent at the FOE/FOC of optic flow. Both expanding and contracting optic flow afford information of heading direction and environmental structure, and, therefore, it would be ecologically beneficial to explore FOE/FOC in order to understand how spatial information figures into efficient searchers of objects.
In conclusion, the current results suggest that optic flow guides attention regardless of attentional control setting, even when another feature captures attention. We have demonstrated that attention can return to the FOE/FOC of optic flow even after directing attention to other features. This suggestion may help expand the attentional theory in vision, but future research will be necessary to better understand the underlying mechanisms.
References
Abrams, R. A., & Christ, S. E. (2003). Motion Onset Captures Attention. Psychological Science, 14(5), 427–432. doi:https://doi.org/10.1111/1467-9280.01458
Al-Aidroos, N., Guo, R. M., & Pratt, J. (2010). You can’t stop new motion: Attentional capture despite a control set for colour. Visual Cognition, 18(6), 859–880. doi:https://doi.org/10.1080/13506280903343085
Anstis, S., Verstraten, F. A. ., & Mather, G. (1998). The motion aftereffect. Trends in Cognitive Sciences, 2(3), 111–117. doi:https://doi.org/10.1016/S1364-6613(98)01142-5
Becker, S. I., Folk, C. L., & Remington, R. W. (2010). The role of relational information in contingent capture. Journal of Experimental Psychology: Human Perception and Performance, 36(6), 1460–1476. doi:https://doi.org/10.1037/a0020370
Borji, A., Feng, M., & Lu, H. (2016). Vanishing point attracts gaze in free-viewing and visual search tasks. Journal of Vision, 16(14), 18. doi:https://doi.org/10.1167/16.14.18
Brainard, D. H. (1997). The psychophysics toolbox. Spatial Vision, 10(4), 433–436. doi:https://doi.org/10.1163/156856897X00357
Chun, M. M., & Jiang, Y. (1998). Contextual cueing: implicit learning and memory of visual context guides spatial attention. Cognitive Psychology, 36(1), 28–71. doi:https://doi.org/10.1006/cogp.1998.0681
Faul, F., Erdfelder, E., Lang, A.-G., & Buchner, A. (2007). G*Power 3: a flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behavior Research Methods, 39(2), 175–91. doi:https://doi.org/10.3758/BF03193146
Fischer, J., & Whitney, D. (2014). Serial dependence in visual perception. Nature Neuroscience, 17(5), 738–743. doi:https://doi.org/10.1038/nn.3689
Folk, C. L., & Annett, S. (1994). Do locally defined feature discontinuities capture attention? Perception & Psychophysics, 56(3), 277–287. doi:https://doi.org/10.3758/BF03209762
Folk, C. L., Remington, R. W., & Johnston, J. C. (1992). Involuntary covert orienting is contingent on attentional control settings. Journal of Experimental Psychology: Human Perception and Performance, 18(4), 1030–1044. doi:https://doi.org/10.1037/0096-1523.18.4.1030
Folk, C. L., Remington, R. W., & Wright, J. H. (1994). The structure of attentional control: Contingent attentional capture by apparent motion, abrupt onset, and color. Journal of Experimental Psychology: Human Perception and Performance, 20(2), 317–329. doi:https://doi.org/10.1037/0096-1523.20.2.317
Franconeri, S. L., & Simons, D. J. (2003). Moving and looming stimuli capture attention. Perception & Psychophysics, 65(7), 999–1010. doi:https://doi.org/10.3758/BF03194829
Gibson, J. J. (1950). The perception of the visual world. Boston: Houghton Mifflin.
Hillstrom, A. P., & Yantis, S. (1994). Visual motion and attentional capture. Perception and Psychophysics, 55(4), 399–411. doi:https://doi.org/10.3758/BF03205298
Holland, B. S., & Copenhaver, M. D. (1988). Improved Bonferroni-type multiple testing procedures. Psychological Bulletin, 104(1), 145–149. doi:https://doi.org/10.1037/0033-2909.104.1.145
Itti, L., & Koch, C. (2001). Computational modelling of visual attention. Nature Reviews Neuroscience, 2(3), 194–203. doi:https://doi.org/10.1038/35058500
Kanai, R., & Verstraten, F. A. J. (2005). Perceptual manifestations of fast neural plasticity: Motion priming, rapid motion aftereffect and perceptual sensitization. Vision Research, 45(25-26), 3109–3116. doi:https://doi.org/10.1016/j.visres.2005.05.014
Kawahara, J., Yanase, K., & Kitazaki, M. (2012). Attentional capture by the onset and offset of motion signals outside the spatial focus of attention. Journal of Vision, 12(12), 1–13. doi:https://doi.org/10.1167/12.12.10
Kiyonaga, A., Scimeca, J. M., Bliss, D. P., & Whitney, D. (2017). Serial dependence across perception, attention, and memory. Trends in Cognitive Sciences, 21(7), 493–497. doi:https://doi.org/10.1016/j.tics.2017.04.011
Klein, R. M. (2000). Inhibition of return. Trends in Cognitive Sciences, 4(4), 138–147. doi:https://doi.org/10.1016/S1364-6613(00)01452-2
Knapen, T., Rolfs, M., & Cavanagh, P. (2009). The reference frame of the motion aftereffect is retinotopic. Journal of Vision, 9(5), 1–6. doi:https://doi.org/10.1167/9.5.16
Maljkovic, V., & Nakayama, K. E. N. (1994). Priming of pop-out : I. Role of Features 22(6), 657–672. doi:https://doi.org/10.3758/BF03209251
Maljkovic, V., & Nakayama, K. E. N. (1996). Priming of pop-out : II . The role of Position, 58(7), 977–991. doi:https://doi.org/10.3758/BF03206826
Pelli, D. G. (1997). The VideoToolbox software for visual psychophysics: Transforming numbers into movies. Spatial Vision, 10(4), 437–442. doi:https://doi.org/10.1163/156856897X00366
Posner, M. I., & Cohen, Y. (1984). Components of visual orienting. Attention and Performance X: Control of Language Processes, 32, 531–556.
Rossini, J. C. (2014). Looming motion and visual attention. Psychology and Neuroscience, 7(3), 425–431. doi:https://doi.org/10.3922/j.psns.2014.042
Schreij, D., Los, S. a, Theeuwes, J., Enns, J. T., & Olivers, C. N. L. (2014). The interaction between stimulus-driven and goal-driven orienting as revealed by eye movements. Journal of Experimental Psychology: Human Perception and Performance, 40(1), 378–390. doi:https://doi.org/10.1037/a0034574
Shaffer, J. P. (1986). Modified sequentially rejective multiple test procedures. Journal of the American Statistical Association, 81(395), 826–831. doi:https://doi.org/10.1080/01621459.1986.10478341
Shirai, N., & Imura, T. (2016). Infant-specific gaze patterns in response to radial optic flow. Scientific Reports, 6, 34734. doi:https://doi.org/10.1038/srep34734
Takeuchi, T. (1997). Visual search of expansion and contraction. Vision Research, 37(15), 2083–2090. doi:https://doi.org/10.1016/S0042-6989(96)00225-8
Theeuwes, J. (1991). Cross-dimensional perceptual selectivity. Perception & Psychophysics, 50(2), 184–193. doi:https://doi.org/10.3758/BF03212219
Ueda, Y., Kamakura, Y., & Saiki, J. (2017). Eye movements converge on vanishing points during visual search. Japanese Psychological Research, 59(2), 109–121. doi:https://doi.org/10.1111/jpr.12144
von Mühlenen, A., & Lleras, A. (2007). No-onset looming motion guides spatial attention. Journal of Experimental Psychology. Human Perception and Performance, 33(6), 1297–1310. doi:https://doi.org/10.1037/0096-1523.33.6.1297
Wang, S., Fukuchi, M., Koch, C., & Tsuchiya, N. (2012). Spatial attention is attracted in a sustained fashion toward singular points in the optic flow. PLoS ONE, 7(8), e41040. doi:https://doi.org/10.1371/journal.pone.0041040
Warren, W. H., Morris, M. W., & Kalish, M. (1988). Perception of translational heading from optical flow. Journal of Experimental Psychology: Human Perception and Performance, 14(4), 646–660. doi:https://doi.org/10.1037/0096-1523.14.4.646
Watanabe, K. (2001). Modulation of spatial attention with unidirectional field motion : an implication for the shift of the OKN beating field, 41, 801–814.
Wohlgemuth, A. (1911). On the affect-effect of seen movement. British Journal of Psychology, Monograph Supplement, (1), 1–117.
Yantis, S., & Jonides, J. (1984). Abrupt visual onsets and selective attention: Evidence from visual search. Journal of Experimental Psychology: Human Perception and Performance, 10(5), 601–621. doi:https://doi.org/10.1037/0096-1523.10.5.601
Yantis, S., & Jonides, J. (1990). Abrupt visual onsets and selective attention: Voluntary versus automatic allocation. Journal of Experimental Psychology: Human Perception and Performance, 16(1), 121–134. doi:https://doi.org/10.1037/0096-1523.16.1.121
Acknowledgements
This study was conducted using the Eyelink and related facilities of Kokoro Research Center, Kyoto University. We thank Kensuke Takii and Jian Guo for the data collection. We also thank Jun Saiki, Yoshiyuki Ueda, Yasunori Kinosada, Shunsuke Kumakiri, Kansai Wakate, and Kumada lab for helpful conversations.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Appendix: The number of fixations
Appendix: The number of fixations
We computed the median number of fixations in visual search display for each participant; the mean of these scores across all participants is shown in Table 2.
The figures show the cumulative distribution functions (CDFs) for the number of fixations required for finding a target from all participants in Experiment 1 (Fig. 12) and Experiments 2–6 (Fig. 13).

The cumulative distribution functions for the number of fixations required for finding a target from all participants in Experiment 1

Rights and permissions
About this article

Cite this article
Higuchi, Y., Inoue, S., Endo, T. et al. Task-irrelevant optic flow guides attention in visual search. Atten Percept Psychophys 81, 1327–1345 (2019). https://doi.org/10.3758/s13414-018-01646-8
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-018-01646-8