
Abstract
Introduction
Mesenchymal stem cells have been increasingly used for cell-based therapies. Adipose-derived stem/stromal cells (ASCs) from the stromal vascular fraction (SVF) of fat tissue are a particularly attractive option for cell based therapy given their accessibility and relative abundance. However, their application in both clinical and basic science investigations is complicated by the isolation of differentiable cells within the SVF. Current enrichment strategies, such as monolayer passaging and surface marker-based sorting, can be time-consuming or overly stringent. Ideally, a population of cells with great regenerative capacity could be isolated with high yields so that extensive in vitro manipulation is not necessary. The objective of this study was to determine whether SVF cells sorted based on expression of alkaline phosphatase liver/bone/kidney (ALPL) resulted in populations with increased osteogenic differentiation potential.
Methods
SVF samples were obtained from four, human donors and processed to isolate initial, heterogeneous cell populations. These SVF cells underwent a four day osteogenic priming period, after which they were treated with a fluorescent, oligodeoxynucleotide molecular beacon probe specific for ALPL mRNA. Cells were separated into positive and negative groups using fluorescence-activated cell sorting (FACS) then differentiated down the osteogenic lineage. Differentiation was assessed by measuring calcified matrix production in each sample.
Results
Cells positive for ALPL expression (ALPL+) represented approximately 34% of the gated population, while cells negative for ALPL expression (ALPL-) represented approximately 18%. ALPL+ cells produced 3.7-fold and 2.1-fold more calcified matrix than ALPL- and unsorted SVF cells, respectively, indicating a significant improvement in osteogenic differentiation. Further, ALPL+ cells showed increases in metabolite production for both adipogenesis and chondrogenesis, suggesting that the enrichment process yields an enhanced multipotent phenotype. Osteogenic differentiation response and cell yields for ALPL+ cells were markedly improved over surface marker-sorted samples.
Conclusion
This study demonstrates a novel method to enrich heterogeneous SVF cells for increased osteogenic potential. The procedure requires less time and results in higher yields of therapeutically useful cells than other existing approaches. Gene expression-based sorting of MSCs is a potentially paradigm-shifting approach that could benefit applications spanning from basic science to clinical therapy.
Similar content being viewed by others

Introduction
Cell enrichment or purification is often a necessary first step for clinical, cell-based therapies as well as basic science investigations into homogeneous subpopulations. Adult mesenchymal stem cells (MSCs) are one type of cell for which this is of great importance. As our understanding of MSCs improves, their use in regenerative medicine becomes ever more promising. This has been especially true for musculoskeletal tissues, with researchers proposing many possibilities using MSCs for the treatment of orthopedic ailments [1–7]. Autologous stem cell transplantation therapies have been proposed for their potential therapeutic versatility and low immunogenicity [8, 9]. However, many of these proposed therapies rely on prior in vitro expansion of the cell populations, which is a slow process and can negatively affect cell phenotype [10]. Single-surgery therapies, where autologous MSCs are isolated and reintroduced into the site of injury in short succession, have the potential to save on both costs and rehabilitation time. Human adipose-derived stem/stromal cells (ASCs), isolated from the stromal vascular fraction (SVF) of lipoaspirate, may be particularly well suited for such single-surgery strategies due to their accessibility and relative abundance in fat tissue, as well as their ability to differentiate down the osteogenic, adipogenic, and chondrogenic lineages [11].
While prolonged culture and passaging is an effective method for isolating ASCs from SVF populations, this process can take weeks to complete. A more rapid approach for the isolation of regeneratively advantageous cells from other cell types contained within the SVF would be enormously beneficial. Traditional cell enrichment strategies have found limited success purifying MSCs due to the lack of a universal surface marker profile [12]. This approach is further complicated by the temporal variability of surface antigens, which can change over time with passaging [13]. Additionally, reported surface marker profiles often result in very low cell yields, necessitating post-sort expansion to obtain sufficient cell numbers for practical applications [14]. Discarded SVF cells can include mesenchymal and endothelial progenitors that may have the potential to differentiate down a subset of lineages [15, 16]. Rigorous surface marker definitions that have been proposed in the literature or by organizations such as the International Federation for Adipose Therapeutics and the International Society for Cellular Therapy may be unnecessarily restrictive for therapeutic applications since they exclude a large fraction of cells with regenerative potential [17].
Alternative enrichment strategies are needed to sort large numbers of therapeutically beneficial cells from the SVF. We thus propose a cell sorting scheme based on expression of mRNA, using molecular beacons as fluorescent reporters. A molecular beacon is an oligodeoxynucleotide, hairpin-shaped, hybridization probe with a fluorophore on the 5′ end and a quencher on the 3′ end [18]. The probe fluorescence is quenched in the absence of target oligonucleotide but is unquenched when the loop region binds to its target mRNA. Other groups have used molecular beacons in conjunction with fluorescence-activated cell sorting (FACS) to sort embryonic stem cells based on OCT4 and SOX2 expression for stemness and pluripotent stem cells based on MHC and TNNT gene families for cardiomyogenesis [19–21]. Given previous successes using molecular beacons with pluripotent stem cells, we apply them here to obtain therapeutically useful cells from a heterogeneous mesenchymal cell population, the SVF.
The objective of this study was to sort cells derived from the SVF of adipose tissue based on expression of alkaline phosphatase liver/bone/kidney (ALPL) to obtain subpopulations of cells capable of enhanced osteogenesis. To do this, we employed a custom-designed molecular beacon for ALPL in combination with FACS [22]. This approach produced high-yield isolations of cellular subpopulations capable of significantly enhanced osteogenesis compared with both unsorted SVF cells and surface-marker sorted ASCs, suggesting increased therapeutic potential for bone regeneration therapies.
Materials and Methods
Cell isolation, culture, and multipotency assessment
Media compositions
Cells were cultured in defined media that served to either maintain stemness, act as a control condition, or induce a differentiation response. Stromal medium, which acted as a control condition, contained Dulbecco’s modified Eagle’s medium (DMEM) with Ham’s F12 salt solution in a 1:1 ratio, 10% fetal bovine serum (FBS) (Zen-Bio, Research Triangle Park, NC, USA), and 1% antibiotic/antimycotic (Fisher Scientific, Pittsburgh, PA, USA). Expansion medium contained stromal medium, with the addition of 5 ng/ml epidermal growth factor, 1 ng/ml fibroblast growth factor, and 0.25 ng/ml transforming growth factor beta-1 (R&D Systems, Minneapolis, MN, USA) to maintain cellular proliferation and multipotency characteristics [10]. Osteogenic differentiation medium contained DMEM with high glucose (4.5 g/l), 10% FBS, 1% antibiotic/antimycotic, 1 nM dexamethasone, 21.6 mg/ml β-glycerophosphate, 50 μg/ml ascorbate-2-phosphate, and 10 μg/ml vitamin D3 (Sigma-Aldrich, St. Louis, MO, USA) [23]. Osteogenic differentiation medium also acted as the priming medium to induce ALPL expression prior to sorts. Adipogenic differentiation medium contained DMEM/F-12, 10% FBS, 1% antibiotic/antimycotic, 10 μg/ml insulin, 0.39 μg/ml dexamethasone, 55.6 μg/ml isobutyl-1-methylxanthine (Sigma-Aldrich), and 17.5 μg/ml indomethacin (Cayman Chemical, Ann Arbor, MI, USA) [23]. Chondrogenic differentiation medium contained DMEM with high glucose, 10% FBS, 1% antibiotic/antimycotic, 10 ng/ml transforming growth factor beta-1, 50 μg/ml ascorbate-2-phosphate, 39.0 ng/ml dexamethasone, and 1% insulin –transferrin – selenium + premix (BD Biosciences, San Diego, CA, USA) [24].
Adipose-derived stromal cell isolation
All procedures involving human donors were approved by the institutional review board of Rhode Island Hospital. Donors provided consent to use surgical waste material for research purposes. SVF cells were isolated from the subcutaneous adipose tissue of human, female donors (N = 4) following established protocols [24]. Briefly, 250 ml liposuction waste tissue was washed with warm phosphate-buffered saline at pH 7.4 and digested with a solution of 0.1% w/v collagenase solution in 1% v/v bovine serum albumin fraction V (Invitrogen, Grand Island, NY, USA) and 2 mM calcium chloride for 60 minutes. Released cells were washed four times with stromal medium, and then incubated for 10 minutes in red blood cell lysis buffer containing 155 mM ammonium chloride, 10 mM potassium carbonate, and 0.1 mM ethylenediamine tetraacetic acid. The resultant cells were then stained with trypan blue and counted using a hemocytometer to determine viability and cell yield. Isolated cells were cryogenically stored in freezing medium containing 10% dimethylsulfoxide, 10% DMEM:Ham’s F12 salt solution, and 80% FBS at a concentration of 5 × 106 to 6 × 106 cells/ml. For designated preliminary/pilot studies, an ASC superlot containing cells from seven nondiabetic donors between the ages of 18 and 60 was purchased commercially and grown to passage 4 before use in experiments (Zen-Bio).
Multilineage differentiation
To determine general multipotency of donor cells, primary SVF cells were seeded in 96-well plates (Genesee Biomedical, Denver, CO, USA) at 8,000 to 10,000 cells/well and differentiated down the osteogenic and adipogenic lineages using the differentiation media described previously (n = 4 for each lineage and corresponding control). For chondrogenic differentiation, 50,000 cells/well were seeded in a V-bottomed 96-well plate and centrifuged at 400 × g to form cell pellets [24]. The cell pellets were then given chondrogenic differentiation medium to induce chondrogenesis or stromal medium to act as a control (n = 4). Samples were cultured for either 2 weeks (adipogenic) or 3 weeks (osteogenic, chondrogenic) before being assessed for lineage-specific metabolites as described below.
Assessment of osteogenesis, adipogenesis, and chondrogenesis
For osteogenesis, samples at 21 days were fixed with 4% paraformaldehyde and stained with alizarin red S, which binds to calcified matrix and is indicative of bone formation (Sigma-Aldrich). For quantification, the dye was eluted using 10% cetylpyridinium chloride, and the absorbance of the eluent was measured at 540 nm. For adipogenesis, samples at 14 days were fixed and stained with oil red O (Sigma-Aldrich), a dye that binds intracellular lipids indicative of fat formation. For quantification, the stain was eluted from fixed cells using 100% isopropanol, and the absorbance of the eluents was measured at 500 nm. To report elution data on a per-cell basis, cell numbers in each sample were quantified by counting Hoechst 33342-stained nuclei per sample using either Gen5 (BioTek U.S., Winooski, VT, USA) or CellProfiler software [25]. For chondrogenesis, samples at 21 days were digested with 125 μg/ml papain at 65°C and pH 6.5 for 24 hours (Sigma-Aldrich). The sulfated glycosaminoglycan (sGAG) content of each digested pellet was quantified using the dimethylmethylene blue assay, modified from established protocols [26, 27]. Briefly, 2.1 mg dimethylmethylene blue was dissolved in 1 ml of 100% ethanol and 10 ml of 0.3 M HCl containing 304 mg glycine and 237 mg sodium chloride. The resulting solution was brought to a volume of 100 ml with deionized water, and the pH of the dimethylmethylene blue dye solution was adjusted to 1.5 using 6 M HCl to account for nonstandard DNA content contributions across samples [28]. Then 200 μl dye was added to 50 μl digest solution, and the absorbance of the resulting mixture was measured at 525 nm. The PicoGreen assay (Invitrogen, Carlsbad, CA, USA) was used to quantify DNA amounts using 100 μl each digest following the manufacturer’s protocol (480 nm excitation, 520 nm emission). A standard curve was used to calculate the total sGAG amount in each pellet, which was then normalized on a per-DNA basis.
Beacon design, gene expression-based sorting, and differentiation of sorted cells
ALPL molecular beacon design, osteogenic priming, and molecular beacon treatment
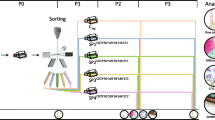
A custom-designed molecular beacon targeting human ALPL, an early marker of osteogenesis [29], was developed as part of our previous work [22]. The beacon sequence was (stems italicized): 5′-(6-carboxyfluorescein) CGCTCC AGAGTGTCTTCCGAGGAGGTCAA GGAGCG (Black Hole Quencher 1)-3′ (melting temperature, 69.4°C; Eurofins MWG Operon, Huntsville, AL, USA). Freshly thawed, primary SVF cells (and passage 4 superlot ASCs, for pilot studies) were seeded in monolayer at 33,000 cells/cm2 and given either osteogenic differentiation medium to prime osteogenic gene expression (primed cells) or expansion medium to maintain their undifferentiated state (nonprimed cells). After 4 days, primed and nonprimed cells were trypsinized using 0.25% trypsin–ethylenediamine tetraacetic acid (Fisher Scientific) and separately resuspended in nonsupplemented base medium (DMEM:Ham’s F12 salt solution) at a concentration of 1 × 106 cells/100 μl. ALPL molecular beacons were added to both cell suspensions at a final concentration of 1 μM immediately prior to electroporation. Beacon-treated cells were electroporated using an Amaxa Nucleofector according to manufacturer’s instructions (program U-23; Lonza AG, Basel, Switzerland). When the process was complete, the cuvette was removed and gently rinsed with 500 μl stromal medium three times to collect all cells in a total of 1.5 ml medium. The cells were allowed to recover for 60 minutes in a humidified 37°C, 5% carbon dioxide incubator. Cells were then pelleted by centrifugation at 400 × g for 5 minutes and subsequently resuspended at a concentration of 10 × 106 cells/ml in warm Hank’s Buffered Salt Solution (Fisher Scientific) for FACS. Samples were protected from light prior to sorting. All sorts were initiated within 1 hour of electroporation. The overall experimental design for the study is illustrated in Figure 1.

Alkaline phosphatase liver/bone/kidney-based sorting method. Gene expression-based sorting of stromal vascular fraction (SVF) cells involved a 4-day osteogenic priming period prior to sorting. Next, cells were treated by electroporation with molecular beacons targeting alkaline phosphatase liver/bone/kidney (ALPL) mRNA. The beacon-treated cells were sorted using fluorescence-activated cell sorting (FACS) into ALPL+ and ALPL– populations. A subset of the initial, primed, SVF cells was left unsorted. The ALPL+, ALPL–, and unsorted cells were seeded onto tissue culture plates, differentiated down the osteogenic lineage, and assessed for production of calcified matrix.
Gene expression-based fluorescence-activated cell sorting
SVF cells were sorted based on positive versus negative signals for ALPL using the previously mentioned molecular beacon. All gene expression-based sorts were performed on a BD FACSAria IIu instrument (BD Biosciences). Cell samples (primed and nonprimed) treated with ALPL beacon were sorted following standard FACS protocols. The instrument was outfitted with an extra-wide 100 μm nozzle to minimize cellular shear stress during the sorting procedure. The forward scatter threshold was set at 5,000 units. Cells were sorted by FACS into positive (ALPL+) and negative (ALPL–) populations using a 488 nm laser and a 530/30 bandpass filter for detection. In every sorting experiment, cells that were not treated with any molecular beacons were used to set gates defining threshold fluorescence levels (that is, all untreated cells were negative, and all intensities above that level were positive). Primed, unsorted cells from the initial SVF population that were not treated with beacons were used as controls for the study. Nonprimed, unsorted cells were not analyzed as part of the main study; however, that condition was analogous to the general multipotency tests conducted for each donor. A mock sort was also conducted with Donor 4 SVF cells using only forward and side scatter parameters to demonstrate that the initial gating process had no influence on osteogenesis (see Additional file 1). Sort data were analyzed using FlowJo FACS analysis software (TreeStar, Inc., Portland, OR, USA).
Cell seeding and differentiation
Following FACS, ALPL+/− cells and unsorted cells were seeded in 96-well plates at 8,000 to 10,000 cells/well and differentiated down the osteogenic lineage using the differentiation medium described previously (N = 4, n = 16 each for osteogenic and control conditions for primed ALPL+ cells, primed ALPL– cells, nonprimed ALPL– cells, and unsorted cells; N = 2, n = 8 for nonprimed ALPL+ cells). After 21 days, samples were fixed, stained, and quantified as described above. Calcified matrix deposition was determined per sample, and then normalized within donor groups to allow for relative comparisons among sorted cell populations. Specifically, raw absorbance values for samples within each donor were normalized to the absorbance of their corresponding unsorted cells. When not noted otherwise, results in this study are reported using these per-sample, donor-normalized values. For a subset of analyses, the raw absorbance values were also normalized on a per-cell basis by counting the number of Hoechst-stained nuclei per sample. In an additional experiment, primed ALPL+/− cells and unsorted cells from a representative donor (Donor 1) were differentiated down the osteogenic, adipogenic, and chondrogenic lineages to assess multipotency of the sorted populations (n = 4 per subpopulation for osteogenesis, adipogenesis, and corresponding control; n = 3 per subpopulation for chondrogenesis and corresponding control).
Surface marker-based sorting comparison
Surface marker-based fluorescence-activated cell sorting
SVF cells from a representative donor (Donor 1) were freshly thawed, stained with trypan blue, and counted with a hemocytometer to determine viability. Cells were then washed twice in 4°C wash buffer (1× PBS, 1% bovine serum albumin), resuspended, and incubated in 4°C blocking buffer (1× PBS, 3% bovine serum albumin) for 10 minutes. Following a wash, cells were aliquoted into separate tubes at a concentration of 105 cells/100 μl for single color controls, negative controls, and sorting. Preconjugated antibodies from BD Pharmingen against human CD34-FITC (#560942), human CD31-PE (#560983), and human CD45-PE-Cy5 (#560974) were used to target the ASC subpopulation [30, 31]. The cells were incubated with antibodies on ice for 20 minutes, followed by a wash. The sort was performed on a BD FACS Influx, and cytometry data were analyzed using FlowJo. Cells were collected into a tube containing expansion medium with 20% FBS.
Differentiation of surface marker-sorted cells
Sorted CD34+/CD31−/CD45− cells were plated at a density of 10,000 cells/well in a tissue culture-treated 96-well plate. The cells were differentiated down the osteogenic and adipogenic lineages (n = 4 for each lineage and corresponding controls), and differentiation was quantified as described previously. Results were compared with ALPL-based sorting to evaluate the relative effectiveness of the two enrichment techniques.
Statistical analysis
Significance in multipotency assessments for all four donors was assessed using Student’s t test (P <0.05; differentiated vs. control in osteogenic, adipogenic, and chondrogenic conditions). Donor-specific experiments were done iteratively to incorporate any systematic, run-to-run error that might be present. Experiments involving ALPL-sorted cells differentiated down the osteogenic, adipogenic, and chondrogenic lineages were assessed using two-way analysis of variance (P <0.05; ALPL expression and differentiation condition) with Holm–Sidak post hoc analyses on SigmaPlot software (SYSTAT Software, San Diego, CA, USA). Lineage-specific differentiation response for surface marker-sorted cells was assessed using Student’s t test (P <0.05; differentiated vs. control in osteogenic and adipogenic conditions). Data are shown as mean ± standard deviation.
Results
Stromal vascular fraction isolation, cell yield, viability, and multipotency
SVF cells were isolated from the fat tissue of four human female donors with high yields and viabilities (Table 1). SVF cells from all four donors exhibited multipotency down at least two different lineages. Donors 1 to 3 could successfully differentiate down the osteogenic, adipogenic, and chondrogenic lineages, whereas Donor 4 did not successfully undergo chondrogenesis.
ALPL-based sorting of stromal vascular fraction cells
SVF cells were successfully sorted based on ALPL gene expression using a custom designed molecular beacon, resulting in an average cell yield for primed ALPL+ cells of 9 ± 3% of the input population (see Additional file 2). Following standard FACS procedures, the input cell population was gated using forward and side scatter parameters, which eliminated aberrantly large or small events (debris particles and cell aggregates; Figure 2). For this gated population, sort data showed that, on average, 34 ± 12% of cells displayed a positive beacon signal (ALPL+) while 18 ± 7% had low/no signal (ALPL–; see Additional file 2). To obtain distinct sort populations, a gap was inserted between positive and negative groups (Figure 2), which represented 48 ± 9% of the gated cells (see Additional file 2). Decreasing the size of this gap would improve cell yields but might result in lower purity for ALPL+/− sorted populations.

Alkaline phosphatase liver/bone/kidney sort yields. Stromal vascular fraction cells treated with alkaline phosphatase liver/bone/kidney (ALPL) molecular beacon were sorted based on gene expression signals. (A) Cells were first gated using forward and side scatter measurements to eliminate debris and cellular aggregates. (B) Nonprimed cells were largely ALPL–, displaying a unimodal distribution overlapping with the primed ALPL– peak. (C) Sorting was determined using the bimodal distribution of primed SVF cells, with clear peaks existing for positive expressers (ALPL+) and negative expressers (ALPL–). A representative set of data is shown here for a single sorting experiment. 6-FAM, 6-carboxyfluorescein.
Osteogenic differentiation across ALPL-sorted donors
ALPL+ cells deposited dramatically more calcified matrix than unsorted or ALPL– cells under osteogenic conditions (Figure 3A). In particular, primed ALPL+ cells induced for osteogenesis displayed a strong propensity for osteogenic matrix formation, showing 2.1-fold more calcified matrix deposition than unsorted cells (P <0.05) and 3.7-fold more than primed ALPL– cells (P <0.05). While significant variation existed among the four donors, these trends were generally consistent, with ALPL+ samples exhibiting more robust osteogenic responses (Figure 3B; see Additional file 3). Unsorted samples did successfully differentiate and produce calcified matrix, but the extent of matrix formation on a donor-normalized basis was much less than that of the primed ALPL+ samples (Figure 4). Unsorted cells deposited 0.9-fold more matrix than their matched, undifferentiated controls, whereas primed ALPL+ cells deposited 5.1-fold more matrix than their undifferentiated controls. Normalizing matrix deposition on a per-cell basis revealed the same trends, suggesting that the sorting process was successfully isolating individual cells with increased synthetic capacity (Table 2).

Osteogenic differentiation of alkaline phosphatase liver/bone/kidney-sorted stromal vascular fraction cells. (A) Osteogenic ALPL+ cells produced significantly more matrix than any other group. Raw absorbance values for all groups were normalized within donors to their respective osteogenic, unsorted samples to allow for relative comparisons. (B) While trends among sorted groups remained the same when separating data by donor, extensive variability in matrix production was observed, with some donor cell populations being especially productive and others relatively dormant. Two-way analysis of variance with Holm–Sidak post hoc test determined significance among sorted groups (*P <0.05). ALPL, alkaline phosphatase liver/bone/kidney.

Alizarin red S staining of osteogenically primed, sorted stromal vascular fraction cells. Primed, sorted SVF cells were differentiated down the osteogenic lineage for 3 weeks and stained for calcified matrix deposition (control medium conditions shown in insets). ALPL+ cells visually produced the most matrix, with very dense, consistent staining across the entire sample. ALPL– cells produced the least matrix, with the stain appearing as a slight wash of orange–red across the culture surface. Unsorted cells produced some matrix, but deposition was scattered, with some areas appearing very red and others unstained. Scale bar: 100 μm. ALPL, alkaline phosphatase liver/bone/kidney.
ALPL-based sorting and differentiation of passaged human adipose-derived stem/stromal cells
Monolayer-expanded, passage 4 superlot ASCs were sorted based on ALPL expression and differentiated down the osteogenic lineage as described previously. The yield of ALPL+ cells with respect to overall cell numbers was 6.7%, with the forward and side scatter gated population being comprised of 81% ALPL+ cells and 12% ALPL– cells. Following osteogenic differentiation, primed ALPL+ cells displayed a 1.2-fold increase in matrix deposition over unsorted cells (P <0.05); however, raw absorbance values reflecting total calcified matrix amounts per sample were on average one-tenth that observed for freshly isolated SVF cells across all sorted groups and donors.
Multipotency assessment of ALPL-sorted cells
To determine whether ALPL sorting isolated a unipotent or multipotent cellular phenotype, additional lineages beyond osteogenesis were assessed for primed ALPL+/− and unsorted groups. ALPL+ cells were the only population capable of adipogenesis (P <0.05; Figure 5A), with lipid accumulation in unsorted and ALPL– samples not changing significantly (P = 0.63 and P = 0.053, respectively). ALPL+ cells showed a 1.3-fold increase in lipid formation over unsorted cells (P <0.05). Chondrogenesis was assessed by quantifying sGAG content normalized to DNA. Both ALPL+ and unsorted samples showed successful differentiation responses (P <0.05), whereas ALPL– samples did not (P = 0.79; Figure 5B). ALPL+ cells produced 1.2-fold and 0.9-fold more sGAG than unsorted cells and ALPL– cells, respectively (P <0.05).

Multilineage differentiation of alkaline phosphatase liver/bone/kidney-sorted stromal vascular fraction cells. Primed, sorted stromal vascular fraction subpopulations were differentiated down the adipogenic and chondrogenic lineages to determine multipotency. ALPL+ samples showed significant increases in metabolite production over ALPL– and unsorted samples for both (A) adipogenic and (B) chondrogenic conditions. Of particular note, only ALPL+ samples showed a significant differentiation response for adipogenesis. ALPL– samples showed no response for either lineage, and unsorted samples only differentiated for chondrogenesis. Two-way analysis of variance with Holm–Sidak post hoc tests determined significance (*or nonmatching letters, P <0.05). ALPL, alkaline phosphatase liver/bone/kidney; sGAG, sulfated glycosaminoglycan.
Surface marker-based sorting and differentiation of stromal vascular fraction cells
SVF cells were sorted using a traditional, surface marker-based approach targeting the CD34+/31−/45− ASC subpopulation and then differentiated down the osteogenic and adipogenic lineages. CD34+/31−/45− cells represented 4% of the overall SVF population and 14% of the gated cell population (see Additional file 4). Following differentiation, the CD34+/31−/45− osteogenic samples produced significantly less matrix than control samples on a per-sample basis, indicating an unsuccessful differentiation response (absorbance: 0.19 ± 0.02 vs. 0.23 ± 0.02, P <0.02; see Additional file 4). On a per-cell basis, osteogenic samples had more matrix deposition than controls, but this increase did not achieve significance (absorbance: 0.22 ± 0.09 vs. 0.14 ± 0.02, P = 0.16). CD34+/31−/45− adipogenic samples produced significantly more lipids than control samples, indicating successful adipogenesis (absorbance: 1.82 ± 0.31 vs. 0.58 ± 0.13, P <0.05), and when normalized on a per-cell basis the relationship remained the same, with adipogenic cells producing significantly more matrix than control cells (absorbance/cell: 4.47 ± 0.90 vs. 0.39 ± 0.11, P <0.05).
Discussion
The results of this study showed that ALPL+ cell populations undergoing osteogenesis could produce up to 4.9 times the calcified matrix of unsorted samples, while also exhibiting increased differentiation potential for adipogenic and chondrogenic lineages. We predicted that, given the heterogeneous nature of SVF cells, only a small subset would express ALPL in response to osteogenic priming. However, results showed that, on average, 34% of gated cells in the SVF were capable of expressing the early osteogenic marker, indicated by a positive ALPL molecular beacon fluorescence. Calcified matrix deposition was significantly increased in ALPL+ cells compared with both unsorted and surface marker-sorted stromal cells, and these trends remained the same whether they were assessed on a per-sample or per-cell basis. ALPL+ cells showed increases in adipogenic and chondrogenic potential as well, indicating that they are not only multipotent but represent a beneficial subpopulation for lineages beyond osteogenesis. Further, this ALPL+ subpopulation was isolated with significantly higher yields than traditional stem cell sorting approaches, representing a potentially transformative method of cell enrichment for MSCs.
Fluorescent tagging of live-cell gene expression in this study allowed for enrichment of SVF cells in a manner similar to surface marker-based sorting. Gene expression-based sorting is an advantageous method for cell enrichment for several reasons. Because gene expression occurs before protein translation, we are able to target cells earlier in the differentiation process than is feasible with protein labeling. This paradigm-shifting approach had no negative effect on cellular growth and differentiation, and the ALPL-targeting probe is safely degraded by natural processes in the cells [22]. Most importantly, ALPL+ cells were found to be a highly productive subpopulation within the SVF and could be an excellent cell source to target for regenerative therapies and basic research. To gain further insight into the abilities and cellular composition of ALPL+ subpopulations, we tested the multilineage differentiation capability of primed, sorted samples. Presumably, sorting based on osteogenic gene expression after a 4-day priming period would result in a population of cells geared towards osteogenic differentiation. As such, we hypothesized that ALPL+ subpopulations would have diminished multilineage differentiation capabilities. Interestingly, ALPL+ subpopulations showed significant increases not only in osteogenic metabolite production but also in characteristic chondrogenic and adipogenic molecules production. When subjected to a standard, 2-week adipogenic differentiation protocol, ALPL+ samples produced 2.3 times the amount of lipid compared with unsorted samples. For chondrogenesis, ALPL+ samples possessed one-half the DNA content of unsorted samples while producing the same amount of sGAG, suggesting that ALPL+ cells were potentially producing double the sGAG per cell than unsorted cells. These findings demonstrated that ALPL+ subpopulations have increased multilineage differentiation capabilities, although the most notable improvement was still for osteogenesis. Further, we found that a 4-day priming period did not fully commit cells to the osteogenic lineage, which is consistent with previous reports [32].
Although ALPL-based sorting reliably isolates cells capable of increased calcified matrix deposition, it is unclear whether the procedure captures cells with the greatest synthetic potential or simply removes an inhibitory population. If we isolated maximally synthetic cells, then an additive relationship should be apparent among sorted groups, with the output of the unsorted population approximately equaling the sum of its contributing subpopulations, primed ALPL+/− cells. Surprisingly, increases in matrix production in ALPL+ cells did not linearly relate to the percentage of cells removed from the initial population. In ALPL+ samples, removing nonexpressing cells, which represented about 18% of the SVF cells on average, resulted in a 210% increase in matrix production. This disproportionate increase may be due to beneficial intercellular communication among cells capable of differentiation, which are enriched in ALPL+ cell populations compared with ALPL– and unsorted samples. This hypothesis is consistent with studies showing that osteogenic differentiation in MSCs is dependent on intercellular communication [33–35]. In comparison, matrix deposition by ALPL– cells was less than that by unsorted cells, but this difference was not statistically significant, indicating the cells still had some osteogenic capacity and were probably not actively inhibiting the process. The specific role of intercellular communication within the sorted populations is yet to be examined.
Donor-to-donor variability is a continual obstacle in assessing the robustness of a treatment or experimental finding [36, 37], so in this study we sorted cells from four different donors to demonstrate the broad applicability of the enrichment procedure. As expected, each donor population deposited variable amounts of calcified matrix following osteogenic differentiation, with Donor 4 cells being the highest producers overall. Interestingly, unsorted Donor 3 samples were unable to osteogenically differentiate on a per-sample basis, but the ALPL+ subpopulation overcame this lack of response with a robust, successful differentiation. Similarly, while unsorted Donor 1 samples were unable to differentiate significantly down the adipogenic lineage, contrary to results from initial multipotency testing on this donor, the ALPL+ subpopulation exhibited a positive differentiation response, indicating superior differentiation capabilities of these sorted cells. Despite differences among donors, gene expression-based sorting consistently isolated highly productive subpopulations of cells (ALPL+). The versatility of this procedure is apparent regardless of innate donor differentiation ability.
Two of the donor SVF populations (Donors 3 and 4) contained a subpopulation of cells that inherently expressed ALPL, without any presort priming. Interestingly, nonprimed ALPL + cells performed similarly to primed ALPL+ cells in terms of matrix deposition, suggesting that some donors may have a ready population of high-potential osteogenic cells that can be isolated directly without need for priming. Another contributor to variability may be the morbidities associated with each donor. Notably, Donors 1, 2, and 3 had previous breast cancer diagnoses, while Donor 4 did not. Donor 4 cells displayed more robust osteogenic differentiation than the others, perhaps due to being from a noncancerous donor. Regardless of prior morbidity, all of the donors were found capable of multilineage differentiation. This is especially encouraging, because it suggests that even prior pathology and treatment regimens did not limit the differentiation ability of ALPL+ subpopulations. That said, more detailed medical histories would be necessary to make any specific hypotheses as to a donor-specific cause of any differentiation differences. Future studies focusing on donor population characteristics may help answer some of these questions.
Surface marker-based sorting strategies are the gold standard technique for cell enrichment. However, these approaches typically produce cell yields significantly lower than those obtained via our gene expression-based sorting strategy. While the highest, reported MSC yields for antigen-based sorting are ~30% of the gated population, some of the more specific surface marker profiles limit yields to <0.1% [17, 31, 38–41]. Also, in many cases the assessed population may only represent a small portion of the initial cell harvest because of restrictive gating for size and granularity. In the current study, we also included a gap region between positive and negative ALPL population peaks to minimize contamination from the other group. Because the peaks are often relatively near each other, a large proportion of cells are sacrificed to the gap (24 to 62% of cells). Refinements to the procedure, such as using a more intense fluorophore with better quenching, can further separate positive and negative peaks, lessening the number of cells present in the gap region. Even with these limitations, the ALPL- based sorting approach produced an overall 9% yield for ALPL+ cells (34% of gated events). We also performed a representative surface marker-based sorting experiment, which isolated cells using a broad CD34+/CD31−/CD45− immunophenotype profile for ASCs [30]. Using this loose definition, ASCs represented only 4% of the population, which is two-fifths of the yield we obtained via gene expression-based sorting. In addition to limited yield, CD34+/CD31−/CD45− cells displayed no increase in calcified matrix deposition when compared with unsorted cells. We hypothesized that surface marker-based sorting would isolate tightly defined, highly differentiable subsets of the larger SVF population. Surprisingly, we found that these cells demonstrated a more limited ability to differentiate down the osteogenic lineage than ALPL+ or unsorted cells. Because of the rigorous, immunophenotype definitions characteristic of surface marker-based sorting, it is possible that other subpopulations of cells capable of differentiation are excluded, resulting in diminished osteogenic capabilities [13]. The broad inclusiveness of our gene expression-based sorting procedure ensures that all cells capable of osteogenic gene expression are captured, rather than a single immunophenotype. Others have shown that a mixed population of cells may be more beneficial to osteogenesis than a highly purified stem-like population [42–44]. These results bolster the notion that stem cell-specific sorting, which aims to eliminate nonstem immunophenotypes, may actually impair the overall regenerative potential of heterogeneous MSC populations.
In addition to experiments using primary cells, gene expression-based sorting was also performed with highly enriched, passaged ASCs comprised of a superlot of donors. Researchers have shown that passaging freshly isolated SVF cells results in enrichment for highly adherent stem-like cells, since less adherent cell types, such as endothelial and circulating hematopoietic cells, remain detached from the culture surface [9, 13]. Based on the reported homogeneity of these passaged ASC populations, we hypothesized that the majority of these cells would express ALPL in response to osteogenic growth factors and that increases in osteogenic matrix deposition in ALPL+ cells compared with unsorted cells would be minimal. As predicted, a larger proportion of cells exhibited a positive beacon signal (81%) based on flow cytometry. Interestingly, a significant increase in matrix deposition was still observed for ALPL+ cells versus unsorted samples. This observation was surprising because it indicated that even supposedly pure ASC populations could be further enriched for cells with enhanced osteogenic capabilities. Despite the positive trends, however, superlot ASCs differentiated poorly in comparison with freshly isolated cells, producing 10 times less matrix than either sorted or unsorted primary SVF cells. Other researchers have also acknowledged the drawbacks of using passaged stromal cells for differentiation [45–47]. While the gene expression-based sorting procedure does benefit the overall differentiation response of passaged ASCs, our best results were achieved using primary SVF-derived cells.
The gene expression-based sorting strategy demonstrated in this study requires a 4-day osteogenic priming period, which is nonideal for single-surgery therapeutic applications. However, priming has been shown to be an effective approach for inducing the expression of characteristic molecules [48]. That said, refinements to our strategy, such as targeting mRNA transcripts earlier in the gene expression pathway, should be considered to minimize or eliminate the priming period. However, the increase in matrix production currently afforded to sorted cells may offset drawbacks associated with cell priming, and some donors may not even require this if ALPL+ cells are inherently present at sufficient numbers. Another potential drawback of the sorting procedure is the presence of false positive cells in the ALPL+ population, given that nonspecific fluorescence is a known limitation of molecular beacons [22, 49, 50]. Despite this potential source of error, ALPL + populations still produced significantly more matrix than any other population of SVF-derived cells, suggesting minimal negative influence from the inclusion of some false-positive cells. Improvements in beacon technology have the potential to further improve outcomes by minimizing this source of error. Lastly, the sorting procedure functions best when targeting abundant mRNA molecules, because the high concentration of target transcripts yields an easily defined positive signal. To target genes that are expressed in low copy numbers, a different strategy must be employed, with one possibility being binding multiple probes to a single transcript to amplify positive signals within the cell [51].
Conclusion
We have demonstrated that sorting stromal cells based on expression of ALPL mRNA isolated a highly synthetic ALPL + subpopulation with significantly improved osteogenesis capabilities. These cells produced more calcified matrix than unsorted stromal cells and were collected in higher yields than existing cell enrichment strategies. ALPL+ cells also showed improved multilineage differentiation capability over unsorted samples, indicating that they are most probably a highly responsive subpopulation within the SVF and not merely osteoprogenitors. Furthermore, since this method improves the cell source at the front end of tissue engineering and cell-based therapies, downstream outcomes have the possibility to see even greater gains. The significance of this work lies in a potential paradigm shift in how cells can be identified and sorted using gene expression-based markers with existing flow cytometry infrastructure. Both basic science investigations and cell-based, clinical therapies could realize significant improvements for applications involving heterogeneous samples.
Note
This article is part of an ‘Emerging Investigators’ collection showcasing the work of early career investigators who have demonstrated growing leadership in the field of stem cells and regenerative medicine. Other articles in the series can be found online at http://stemcellres.com/series/emerginginvestigators.

Box 1. About Eric Darling
EMD is the Manning Assistant Professor of Medical Science, Orthopaedics, and Engineering in the Department of Molecular Pharmacology, Physiology, & Biotechnology at Brown University. He also currently serves as the Graduate Program Director for the Center for Biomedical Engineering. He received a BS in engineering from Harvey Mudd College, a PhD in bioengineering from Rice University, and postdoctoral training in orthopedic research at Duke University. His research area focuses on understanding the relationship between the mechanical and biological characteristics of cells and tissues, with emphasis on the musculoskeletal system. He is specifically interested in understanding heterogeneity in adult stem cell populations and developing approaches to identify tissue-specific cells for regenerative medicine and disease diagnostics. Recent work in his group has focused on two, novel cellular characteristics: single-cell mechanical biomarkers and live-cell gene expressions.
Abbreviations
- ALPL :
-
alkaline phosphatase liver/bone/kidney
- ASC:
-
human adipose-derived stem/stromal cell
- DMEM:
-
Dulbecco’s modified Eagle’s medium
- FACS:
-
fluorescence-activated cell sorting
- FBS:
-
fetal bovine serum
- MSC:
-
mesenchymal stem cell
- sGAG:
-
sulfated glycosaminoglycan
- SVF:
-
stromal vascular fraction.
References
Valarmathi MT, Yost MJ, Goodwin RL, Potts JD: A three-dimensional tubular scaffold that modulates the osteogenic and vasculogenic differentiation of rat bone marrow stromal cells. Tissue Eng A. 2008, 14: 491-504. 10.1089/tea.2007.0235.
Arzi B, Verstraete FJ, Huey DJ, Cissell DD, Athanasiou KA: Regenerating mandibular bone using rhBMP-2: Part 1 – immediate reconstruction of segmental mandibulectomies. Vet Surg. 2014, [Epub ahead of print]
Diekman BO, Rowland CR, Lennon DP, Caplan AI, Guilak F: Chondrogenesis of adult stem cells from adipose tissue and bone marrow: induction by growth factors and cartilage-derived matrix. Tissue Eng A. 2010, 16: 523-533.
Turgeman G, Pittman DD, Muller R, Kurkalli BG, Zhou S, Pelled G, Peyser A, Zilberman Y, Moutsatsos IK, Gazit D: Engineered human mesenchymal stem cells: a novel platform for skeletal cell mediated gene therapy. J Gene Med. 2001, 3: 240-251. 10.1002/1521-2254(200105/06)3:3<240::AID-JGM181>3.0.CO;2-A.
Bruder SP, Kurth AA, Shea M, Hayes WC, Jaiswal N, Kadiyala S: Bone regeneration by implantation of purified, culture-expanded human mesenchymal stem cells. J Orthop Res. 1998, 16: 155-162. 10.1002/jor.1100160202.
Parrilla C, Saulnier N, Bernardini C, Patti R, Tartaglione T, Fetoni AR, Pola E, Paludetti G, Michetti F, Lattanzi W: Undifferentiated human adipose tissue-derived stromal cells induce mandibular bone healing in rats. Arch Otolaryngol Head Neck Surg. 2011, 137: 463-470. 10.1001/archoto.2011.61.
Wang CZ, Chen SM, Chen CH, Wang CK, Wang GJ, Chang JK, Ho ML: The effect of the local delivery of alendronate on human adipose-derived stem cell-based bone regeneration. Biomaterials. 2010, 31: 8674-8683. 10.1016/j.biomaterials.2010.07.096.
McIntosh KR, Lopez MJ, Borneman JN, Spencer ND, Anderson PA, Gimble JM: Immunogenicity of allogeneic adipose-derived stem cells in a rat spinal fusion model. Tissue Eng A. 2009, 15: 2677-2686. 10.1089/ten.tea.2008.0566.
McIntosh K, Zvonic S, Garrett S, Mitchell JB, Floyd ZE, Hammill L, Kloster A, Di Halvorsen Y, Ting JP, Storms RW, Goh B, Kilroy G, Wu X, Gimble JM: The immunogenicity of human adipose-derived cells: temporal changes in vitro. Stem Cells. 2006, 24: 1246-1253. 10.1634/stemcells.2005-0235.
Estes BT, Diekman BO, Guilak F: Monolayer cell expansion conditions affect the chondrogenic potential of adipose-derived stem cells. Biotechnol Bioeng. 2008, 99: 986-995. 10.1002/bit.21662.
Zuk PA, Zhu M, Mizuno H, Huang J, Futrell JW, Katz AJ, Benhaim P, Lorenz HP, Hedrick MH: Multilineage cells from human adipose tissue: implications for cell-based therapies. Tissue Eng. 2001, 7: 211-228. 10.1089/107632701300062859.
Tuan RS, Boland G, Tuli R: Adult mesenchymal stem cells and cell-based tissue engineering. Arthritis Res Ther. 2003, 5: 32-45.
Mitchell JB, McIntosh K, Zvonic S, Garretta S, Floyd ZE, Kloster A, Di Halvorsen Y, Storms RW, Goh B, Kilroy G, Wu X, Gimble JM: Immunophenotype of human adipose-derived cells: temporal changes in stromal-associated and stem cell-associated markers. Stem Cells. 2006, 24: 376-385. 10.1634/stemcells.2005-0234.
Mesimaki K, Lindroos B, Tornwall J, Mauno J, Lindqvist C, Kontio R, Miettinen S, Suuronen R: Novel maxillary reconstruction with ectopic bone formation by GMP adipose stem cells. Int J Oral Maxillofac Surg. 2009, 38: 201-209. 10.1016/j.ijom.2009.01.001.
Zimmerlin L, Donnenberg VS, Pfeifer ME, Meyer EM, Peault B, Rubin JP, Donnenberg AD: Stromal vascular progenitors in adult human adipose tissue. Cytometry A. 2010, 77: 22-30.
Lin G, Garcia M, Ning H, Banie L, Guo YL, Lue TF, Lin CS: Defining stem and progenitor cells within adipose tissue. Stem Cells Dev. 2008, 17: 1053-1063. 10.1089/scd.2008.0117.
Bourin P, Bunnell BA, Casteilla L, Dominici M, Katz AJ, March KL, Redl H, Rubin JP, Yoshimura K, Gimble JM: Stromal cells from the adipose tissue-derived stromal vascular fraction and culture expanded adipose tissue-derived stromal/stem cells: a joint statement of the International Federation for Adipose Therapeutics and Science (IFATS) and the International Society for Cellular Therapy (ISCT). Cytotherapy. 2013, 15: 641-648. 10.1016/j.jcyt.2013.02.006.
Tyagi S, Kramer FR: Molecular beacons: probes that fluoresce upon hybridization. Nat Biotechnol. 1996, 14: 303-308. 10.1038/nbt0396-303.
King FW, Liszewski W, Ritner C, Bernstein HS: High-throughput tracking of pluripotent human embryonic stem cells with dual fluorescence resonance energy transfer molecular beacons. Stem Cells Dev. 2011, 20: 475-484. 10.1089/scd.2010.0219.
Ban K, Wile B, Kim S, Park HJ, Byun J, Cho KW, Saafir T, Song MK, Yu SP, Wagner M, Bao G, Yoon YS: Purification of cardiomyocytes from differentiating pluripotent stem cells using molecular beacons that target cardiomyocyte-specific mRNA. Circulation. 2013, 128: 1897-1909. 10.1161/CIRCULATIONAHA.113.004228.
Larsson HM, Lee ST, Roccio M, Velluto D, Lutolf MP, Frey P, Hubbell JA: Sorting live stem cells based on Sox2 mRNA expression. PLoS One. 2012, 7: e49874-10.1371/journal.pone.0049874.
Desai HV, Voruganti IS, Jayasuriya C, Chen Q, Darling EM: Live-cell, temporal gene expression analysis of osteogenic differentiation in adipose-derived stem cells. Tissue Eng A. 2014, 20: 899-907. 10.1089/ten.tea.2013.0761.
Gonzalez-Cruz RD, Fonseca VC, Darling EM: Cellular mechanical properties reflect the differentiation potential of adipose-derived mesenchymal stem cells. Proc Natl Acad Sci U S A. 2012, 109: E1523-E1529. 10.1073/pnas.1120349109.
Estes BT, Diekman BO, Gimble JM, Guilak F: Isolation of adipose-derived stem cells and their induction to a chondrogenic phenotype. Nat Protoc. 2010, 5: 1294-1311. 10.1038/nprot.2010.81.
Carpenter AE, Jones TR, Lamprecht MR, Clarke C, Kang IH, Friman O, Guertin DA, Chang JH, Lindquist RA, Moffat J, Golland P, Sabatini DM: Cell Profiler: image analysis software for identifying and quantifying cell phenotypes. Genome Biol. 2006, 7: R100-10.1186/gb-2006-7-10-r100.
Guilak F, Lott KE, Awad HA, Cao Q, Hicok KC, Fermor B, Gimble JM: Clonal analysis of the differentiation potential of human adipose-derived adult stem cells. J Cell Physiol. 2006, 206: 229-237. 10.1002/jcp.20463.
Awad HA, Halvorsen YD, Gimble JM, Guilak F: Effects of transforming growth factor beta1 and dexamethasone on the growth and chondrogenic differentiation of adipose-derived stromal cells. Tissue Eng. 2003, 9: 1301-1312. 10.1089/10763270360728215.
Enobakhare BO, Bader DL, Lee DA: Quantification of sulfated glycosaminoglycans in chondrocyte/alginate cultures, by use of 1,9-dimethylmethylene blue. Anal Biochem. 1996, 243: 189-191. 10.1006/abio.1996.0502.
Kim EK, Lim S, Park JM, Seo JK, Kim JH, Kim KT, Ryu SH, Suh PG: Human mesenchymal stem cell differentiation to the osteogenic or adipogenic lineage is regulated by AMP-activated protein kinase. J Cell Physiol. 2012, 227: 1680-1687. 10.1002/jcp.22892.
Oedayrajsingh-Varma MJ, van Ham SM, Knippenberg M, Helder MN, Klein-Nulend J, Schouten TE, Ritt MJ, van Milligen FJ: Adipose tissue-derived mesenchymal stem cell yield and growth characteristics are affected by the tissue-harvesting procedure. Cytotherapy. 2006, 8: 166-177. 10.1080/14653240600621125.
Kanthilal M, Darling EM: Characterization of mechanical and regenerative properties of human, adipose stromal cells. Cell Mol Bioeng. 2014, doi:10.1007/s12195-014-0350-y
Engler AJ, Sen S, Sweeney HL, Discher DE: Matrix elasticity directs stem cell lineage specification. Cell. 2006, 126: 677-689. 10.1016/j.cell.2006.06.044.
Ishikawa M, Iwamoto T, Nakamura T, Doyle A, Fukumoto S, Yamada Y: Pannexin 3 functions as an ER Ca(2+) channel, hemichannel, and gap junction to promote osteoblast differentiation. J Cell Biol. 2011, 193: 1257-1274. 10.1083/jcb.201101050.
Nakahama K: Cellular communications in bone homeostasis and repair. Cell Mol Life Sci. 2010, 67: 4001-4009. 10.1007/s00018-010-0479-3.
Chang JC, Fujita S, Tonami H, Kato K, Iwata H, Hsu SH: Cell orientation and regulation of cell-cell communication in human mesenchymal stem cells on different patterns of electrospun fibers. Biomed Mater. 2013, 8: 055002-10.1088/1748-6041/8/5/055002.
Bodle JC, Teeter SD, Hluck BH, Hardin JW, Bernacki SH, Loboa EG: Age-related effects on the potency of human adipose-derived stem cells: creation and evaluation of superlots and implications for musculoskeletal tissue engineering applications. Tissue Eng Part C. 2014, doi:10.1089/ten.tec.2013.0683
Baer PC, Kuci S, Krause M, Kuci Z, Zielen S, Geiger H, Bader P, Schubert R: Comprehensive phenotypic characterization of human adipose-derived stromal/stem cells and their subsets by a high throughput technology. Stem Cells Dev. 2013, 22: 330-339. 10.1089/scd.2012.0346.
Varma MJ, Breuls RG, Schouten TE, Jurgens WJ, Bontkes HJ, Schuurhuis GJ, van Ham SM, van Milligen FJ: Phenotypical and functional characterization of freshly isolated adipose tissue-derived stem cells. Stem Cells Dev. 2007, 16: 91-104. 10.1089/scd.2006.0026.
Li H, Zimmerlin L, Marra KG, Donnenberg VS, Donnenberg AD, Rubin JP: Adipogenic potential of adipose stem cell subpopulations. Plast Reconstr Surg. 2011, 128: 663-672. 10.1097/PRS.0b013e318221db33.
Zannettino AC, Paton S, Arthur A, Khor F, Itescu S, Gimble JM, Gronthos S: Multipotential human adipose-derived stromal stem cells exhibit a perivascular phenotype in vitro and in vivo. J Cell Physiol. 2008, 214: 413-421. 10.1002/jcp.21210.
Gronthos S, Zannettino AC: A method to isolate and purify human bone marrow stromal stem cells. Methods Mol Biol. 2008, 449: 45-57.
Stains JP, Civitelli R: Cell–cell interactions in regulating osteogenesis and osteoblast function. Birth Defects Res C Embryo Today. 2005, 75: 72-80. 10.1002/bdrc.20034.
Forriol F, Shapiro F: Bone development: interaction of molecular components and biophysical forces. Clin Orthop Relat Res. 2005, 432: 14-33.
Wang IE, Shan J, Choi R, Oh S, Kepler CK, Chen FH, Lu HH: Role of osteoblast–fibroblast interactions in the formation of the ligament-to-bone interface. J Orthop Res. 2007, 25: 1609-1620. 10.1002/jor.20475.
Qureshi AT, Chen C, Shah F, Thomas-Porch C, Gimble JM, Hayes DJ: Human adipose-derived stromal/stem cell isolation, culture, and osteogenic differentiation. Methods Enzymol. 2014, 538: 67-88.
Schellenberg A, Lin Q, Schuler H, Koch CM, Joussen S, Denecke B, Walenda G, Pallua N, Suschek CV, Zenke M, Wagner W: Replicative senescence of mesenchymal stem cells causes DNA-methylation changes which correlate with repressive histone marks. Aging. 2011, 3: 873-888.
Dao LT, Park EY, Hwang OK, Cha JY, Jun HS: Differentiation potential and profile of nuclear receptor expression during expanded culture of human adipose tissue-derived stem cells reveals PPARγ as an important regulator of Oct4 expression. Stem Cells Dev. 2014, 23: 24-33. 10.1089/scd.2013.0137.
Ochiai-Shino H, Kato H, Sawada T, Onodera S, Saito A, Takato T, Shibahara T, Muramatsu T, Azuma T: A novel strategy for enrichment and isolation of osteoprogenitor cells from induced pluripotent stem cells based on surface marker combination. PLoS One. 2014, 9: e99534-10.1371/journal.pone.0099534.
Tsourkas A, Behlke MA, Bao G: Hybridization of 2′-O-methyl and 2′-deoxy molecular beacons to RNA and DNA targets. Nucleic Acids Res. 2003, 31: 5168-5174.
Santangelo PJ, Nix B, Tsourkas A, Bao G: Dual FRET molecular beacons for mRNA detection in living cells. Nucleic Acids Res. 2004, 32: e57-10.1093/nar/gnh062.
Bratu DP, Catrina IE, Marras SA: Tiny molecular beacons for in vivo mRNA detection. Methods Mol Biol. 2011, 714: 141-157. 10.1007/978-1-61779-005-8_9.
Acknowledgements
The authors would like to thank Dr Paul Liu and Dr Mikki Ciombor from Rhode Island Hospital for providing human lipoaspirate. This work was supported by awards from the National Science Foundation (EMD; CAREER Award, CBET1253189), the National Institute of Arthritis and Musculoskeletal and Skin Diseases (EMD; R01 AR063642), the National Institute of General Medical Sciences (EMD; P20 GM104937), and the United States Department of Education (HDM; GAANN fellowship P200A120064). The content of this article is solely the responsibility of the authors and does not necessarily represent the official views of the National Science Foundation, National Institutes of Health, or US Department of Education.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
HDM and EMD are inventors on a provisional patent application involving this work (US Provisional Application Serial No. 61/874,070, ‘Methods of Identifying and Isolating Cells Using Molecular Beacons’).
Authors’ contributions
HDM and EMD designed the study. HDM and BAS performed gene expression-based sorting experiments. HDM and MK performed surface marker-based sorting experiments. HDM, VCF, and BAS performed staining and elution of differentiated samples. HDM, BAS, and EMD analyzed the data and wrote the manuscript. All authors read and approved the final manuscript.
Electronic supplementary material
13287_2014_410_MOESM1_ESM.pdf
Additional file 1: Figure S1: Showing a mock sort of gated SVF cells. Donor 4 SVF cells were gated and sorted based on just forward (FSC) and side (SSC) scatter parameters, to ensure that the initial gating process alone was not enriching the cells. Gated osteogenic samples did not show any change in calcified matrix production compared with unsorted osteogenic samples, providing evidence that simply putting cells through the flow cytometer did not affect osteogenic response to any significant extent (P = 0.89). (PDF 275 KB)
13287_2014_410_MOESM3_ESM.pdf
Additional file 3: Figure S2: Showing donor-specific calcified matrix production for sorted SVF cells. SVF cells from four, distinct donors were sorted based on expression of ALPL mRNA. Absorbance values of eluted alizarin red S, indicative of calcified matrix production, were normalized on a per-cell basis by counting Hoechst 33342-stained nuclei in each sample. In all donors, ALPL+ groups consistently produced more calcified matrix on a per-cell basis than any other group. However, in Donor 4 the overall matrix production was higher, and so primed ALPL+ cells were not significantly more productive than unsorted cells (P = 0.97). Sample groups with nonmatching letters are significantly different (P <0.05). (PDF 333 KB)
13287_2014_410_MOESM4_ESM.pdf
Additional file 4: Figure S3: Showing surface marker-based sorting of SVF cells. Freshly thawed SVF cells were labeled with fluorescent antibodies for CD34, CD31, and CD45 and sorted using a BD FACS Influx. (A) Gated forward and side scatter cells were (B) 23% positive for CD34 surface antigen expression. (C) Of the CD34+ cells, 58% were also CD31− and CD45−. Overall yield for CD34+/31−/45− cells was 4%, and these cells displayed only a limited ability to differentiate down the osteogenic lineage. (D) In comparison, primed, ALPL+ cells produced 25-fold more calcified matrix than CD34+/31−/45− cells. Sample groups with nonmatching letters are significantly different (P <0.05). (PDF 411 KB)
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Marble, H.D., Sutermaster, B.A., Kanthilal, M. et al. Gene expression-based enrichment of live cells from adipose tissue produces subpopulations with improved osteogenic potential. Stem Cell Res Ther 5, 145 (2014). https://doi.org/10.1186/scrt502
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1186/scrt502