
Abstract
Background
Pink stem borer, Sesamia inferens (Walker), a major rice pest, has rarely been reported to cause serious damage to sorghum in China. During a survey of insect pests in glutinous sorghum for Chinese liquor production areas in Guizhou Province, it was found that S. inferens has caused serious damage to local sorghum in recent years.
Methods
In this context, we assessed the adaptation of S. inferens to three gramineous crop host plants (maize, rice and sorghum) based on age-stage, two-sex life table theory.
Results
Sesamia inferens had the highest development time, survival rate and total longevity on rice, followed by on sorghum and maize. There was no significant difference in intrinsic rate of increase and finite rate of increase among maize (r = 0.0702 day−1, λ = 1.0727 day−1), sorghum (r = 0.0681 day−1, λ = 1.0705 day−1) and rice (r = 0.0645 day−1, λ = 1.0666 day−1), but the mean generation period (T) was significantly higher for populations reared on rice (53.84 days) than on sorghum (49.53 days) and maize (47.31 days).
Conclusions
The results of our study indicate that S. inferens was able to complete the full developmental cycle on all three host plants. This study further supports that glutinous sorghum is an adaptable host plant for S. inferens.
Similar content being viewed by others


Background
Pink stem borer, Sesamia inferens (Walker) (Lepidoptera: Noctuidae), is a polyphagous pest attacking various gramineous crops like rice, maize, pearl millet, finger millet, wheat, oats, barley, sugarcane, and some grasses (Banerjee et al. 1967; Khan et al. 1991), and complete generations can be achieved on rice, water bamboo, and maize (Chen et al. 2017). It is a serious pest of almost all cereals by attacking the crops at various stages and causing severe damage and yield losses (Hashmi 1994; Baladhiya et al. 2018). Sesamia inferens was first reported in rice, which later migrated to other gramineous crops (Pathak and Khan 1994).
When S. inferens infests crops in the field, the larvae emerge in groups within or behind the leaf sheath and feed on the epidermal layer of the leaf sheath (Jeevanandham et al. 2020). As a result of eating, gummy oozes with water-soaked lesions appears on the affected leaf sheaths that were externally visible in the initial stages of eating (Chen 2019). The larvae then reach the central growing point by boring through the sheaths (Hashmi 1994). Then feed on unopened leaves in the whorl, resulting in the formation of oblong (oblong or oval, slightly larger than pin size) holes of 2–3 mm in size in parallel rows in the unfolded leaves (Jeevanandham et al. 2020). As the plant grows, these holes widened and became slits and streaks. In severe cases, tunneling of the midrib was visible on the leaf blades and the plants had a frayed appearance (Pathak and Khan 1994). Attack of the shoot growing point eventually lead to desiccation, resulting in the formation of a “dead heart” at the seedling stage in cereals and “white ears” at the ear head stage in sorghum (Sau et al. 2022). Before the 3rd instar, the larvae usually gather in the form of dozens of heads, feed first on the inner layer of the leaf sheath, then bore into the stem of the plant to continue feeding, resulting in a “withered heart” (Baladhiya et al. 2018). The insects feed on the stems, causing hollow stems, the plant collapses and the rate of course reduces (Hashmi 1994).
In China, S. inferens, known as a major pest in rice, has rarely been reported to cause serious damage to sorghum, but our research team unexpectedly found a high number of S. inferens infestations on glutinous sorghum in the Guizhou region in 2021. The pests have resulted in the withering of young sorghum seedlings, large areas of fallen or lopsided plants, and reduced spike rates, and have led to a significant reduction in sorghum yield and quality. Sorghum is a gramineous crop that has many uses such as food, feed, and brewing (Aruna 2018). With the adjustment of the agricultural industrial structure, sorghum has huge development space and industrial advantages in agricultural production (Zhang et al. 2014). Among them, the winemaking industry is one of the important industries in China’s economic development, and also one of the industries with the most benefits and development potential in China’s sorghum industry (Gao et al. 2020). Since 2012, China’s liquor industry has entered a deep adjustment period. As the main upstream raw material of liquor companies, plant structure has changed markedly (Fan et al. 2022; Yan et al. 2017). In this context, Southwest China region of high-grade glutinous sorghum for wine with first-class quality, is leading the rapid development of high-quality grain development and use as a model (Yan et al. 2017). Guizhou province is the birthplace and main production area of Moutai flavored liquors, and the liquor industry is inseparable from the production of sorghum (Ding et al. 2019). The occurrence of S. inferens in sorghum production areas in Guizhou Province is increasing year by year, requiring widespread attention. Sorghum was its host plant at the time of its discovery, and S. inferens is widely regarded as an important rice pest in China and has once been reported as a pest on maize too (Chen 2019; Baladhiya et al. 2018; Dhaliwal et al. 2010; Chen et al. 2017). While rice and maize are major food crops, sorghum is of great economic value in the liquor industry. It is important to ensure the quality of the yields of these three crops. Maize and sorghum are both gramineous crops and have similar growth stages and morphological structures (Lozano et al. 2021). So three host plants, maize, sorghum, and rice, were chosen for this study.
Phytophagous insects are dependent on host plants and host plant be selected is important for their survival and reproduction, and insects have different adaptations to different host plants (Bernays 1994). Different host plants had a significant effect on the bioenergetic, development, and oviposition preference of fall armyworm, and the results of this study are important for understanding the adaptability of fall armyworm on different host plants (Banerjee and Ray 1995; Veenstra et al. 1995; Guo et al. 2020). Agrawal (2000) found that mites can adapt to different host plants and that there is a trade-off between adaptability and fitness on different hosts. On some hosts, the mites can achieve higher reproductive success, but may face higher survival risks on other hosts. Therefore, the research on the adaptation of insects to different host plants is important for understanding the ecological characteristics of insects and assessing crops at potential risk.
The life table is an important tool in population ecology, it can clearly show the demographic parameters such as survival rate, development time, longevity and fecundity (Chi et al. 2022a, b). Various analyses, such as population extinction estimation, population growth capacity and pest outbreak prediction, can be measured based on the demographic parameters (Amiri et al. 2010). Therefore, the construction of life table contributes to the understanding of insect population dynamics, which plays a vital role in integrated pest management (IPM) strategy (Caldirola and Southwood 1968). In this study, we compare and analyze the statistical population parameters of S. inferens on maize and rice compared to sorghum (the native host in the sorghum-producing area of Guizhou) using the computer program TWOSEX-MS Chart (Chi 2021) based on age-stage, two-sex life table theory (Chi et al. 2020; Saska et al. 2021). The fitness of S. inferens on three host plants was investigated to provide the theoretical basis for their prevention and control measures in the future.
Methods
Plants
Plants of maize (variety: Zhuoyu 9), sorghum (variety: Jinnuoliang 8), and rice (variety: Taiyou 808), all obtained from GuiZhou Zhuohao Agricultural Science, Guiyang, China, were grown in a greenhouse at 27 ± 3 °C with 70 ± 10% RH (relative humidity) and a photoperiod of 14L:10D (light:dark) in Guizhou University, China. Healthy and undamaged plants were collected for experiments. No pesticides were applied to any of the three crops and no infestations were observed. For the experiments, we used plants that were at different growth stages, including the seedling stage, the jointing stage/tillering stage, and the heading stage.
The seedlings of the three host plants were selected at a height of about 40 cm for testing. Maize plants were with 6–9 leaves, a height of about 60–70 cm, and at the beginning to reach the jointing stage. Sorghum plants were with 8–12 leaves, a height of about 60–70 cm, and to reach the jointing stage. The selection of plants to be tested has been carried out within these criteria. The criterion of the shoot stage was that the first shoot bud germinated and protruded 1–2 cm from basal leaf axils. Rice plants with primary shoots were used in this study, and their height was about 50–60 cm. Heading refers to the stage or condition in which the fully developed young panicle of cereal crops emerges from the sheath of the flag leaf. The maize and sorghum plants selected for this study were about 1 m in high, and the rice plants were about 60–70 cm high.
Insects
The original colony of S. inferens was collected in June 2020 from Sorghum in Renhuai county (106°44′ E, 27°67′ N), Guizhou province, China. It was identified as S. inferens by comparison with the morphological descriptions of S. inferens as reported in Zhu et al. (1963) and by molecular biology depending on CO I gene (accession number: GenBank OR676825). Larvae were grown on zizania in plastic cages of 1500 mL until they became pupa. After emergence, the adults were placed into a cage (80 cm × 100 cm × 120 cm) and provided with a cotton-wool ball soaked with 10% honey water solution as food. In this experiment, the test insects were cultured in the laboratory (temperature: 26 °C ± 1 °C, relative humidity: 70 ± 5%, light/dark cycle: 14L:10D) with zizania for more than three generations.
Demography and fitness of S. inferens feeding on three host plants
The experiment was performed under greenhouse conditions at 27 ± 3 °C with 70 ± 10% RH and a photoperiod of 14L:10D. Taking the S. inferens life table experiment on the sorghum plant as an example, the methods are as follows. Ninety eggs were taken and the time of egg laying and hatching was recorded. We used 30 eggs in a group for this test. After hatching, the larvae were transferred with a hairbrush to the leaf sheaths and core leaves of the sorghum seedlings (3 plants in 1 pot, and 1 pot provided). The sorghum seedlings were placed in an insect cage of 80 cm × 100 cm × 120 cm, and the larvae were allowed to feed freely inside the plant. Three days later, we observed the traces of insect infestation, if it is clear that there are traces of insect infestation, then we continued to test; but if not, then we followed the experimental procedure to repeat a series of experiments. In addition, new sorghum seedlings (3 plants in 1 pot, and 5 pots provided) were placed into the insect cage. We daily observed the sorghum plants eaten by the borers for a period of 10 days, and we timely provided new sorghum plants to the cage. Here there was no longer a need to provide sorghum plants at the seedling stage, but we could provide plants at the jointing stage and heading stage until the larvae became pupa. This is also because the older S. inferens larvae prefer to feed on stout plant stems. Newly potted plants were placed just next to old plants to ensure that their leaves touched each other so that the larvae could independently move to new plants (Marina et al. 2019).
After adult emergence, they were introduced into an insect cage (80 cm × 100 cm × 120 cm) including new sorghum plants at different growth stages (seedling, jointing and heading stages) as oviposition places, and provided with 10% water honey solution as diet. The process of insects breeding and experiments referred to Fig. 1. The sorghum plants and honey solution were changed daily. The total number of eggs was counted daily until all adults were died and we also recorded the sex ratio.
The method with maize as host plant is similar to that of sorghum as a host plant, while in the experiment with rice as host plant we only needed to replace the plants at the jointing stage with those at the tillering stage.

Insects breeding and experiment processing
Life table analysis
The computer program TWOSEX-MSChart, based on the age-stage, two-sex life table theory, was used to calculate the demographic parameters such as development time, survival rate, longevity, and fecundity. The formulas and definitions used in this study are shown in Additional file 1: Table S1 (Definitions and formulas of population parameters estimated in the present study). TWOSEX-MSChart 2022 software was used to calculate each parameter. The standard errors were determined by bootstrap with 100,000 repetitions, and evaluated the statistical significance of the observed differences using paired bootstrap, where P < 0.05 was considered statistically significant (Shahidi-Noghabi et al. 2009; Wang et al. 2016; Ding et al. 2021; Zhao et al. 2021; Amir-Maafi et al. 2022). All graphs were created using SigmaPlot 12.5 (Systat Software, San Jose, CA, USA, https://www.systatsoftware.com).
Results
Development time, longevity, and the sex ratio of S. inferens
The development time, longevity, and proportion of adult females of S. inferens from the different host plants are shown in Table 1. The preadult longevity of S. inferens on rice (49.34 d) was longer than on sorghum (45.33 d) and maize (42.68 d). The highest preadult survival rate (sa) of S. inferens on rice (0.5556) was significantly higher than on maize (0.3778). Preadult longevity of female and male on rice (48.76 d, 50.14 d) was longer than on sorghum (45.12 d, 45.69 d) and maize (41.91 d, 44.27 d). The total longevity on rice (55.69 d) was the longest, followed by sorghum (50.79 d) and maize (49.16 d). Of the three host plants, the total longevity of females and males on rice is the longest (55.90 d, 57.38 d). There were significant differences among the three host plants in the total longevity of the females (rice, 55.9 d; sorghum, 51.88 d; maize, 49.87 d). In contrast, no significant differences were observed for the S. inferens male and female adult longevity between maize (7.96 d, 7.27 d) and rice (7.14 d, 7.24 d), the fecundity (number of eggs per female) (maize, 108.13; sorghum, 100.88; rice, 99.93), and the proportion of female adults (maize, 0.2556; sorghum, 0.2889; rice, 0.3222).
Population statistics parameters
The net reproduction rate (R0), intrinsic rate of increase (r), finite rate of increase (λ), mean generation time (T), and gross reproduction rate (GRR) of S. inferens on the three host plants are shown in Table 2. There were no significant differences between r, R0, and λ on the three hosts. The r and λ values of S. inferens populations on the three host plants were > 0 and > 1, respectively (in detail for maize, 0.0702, 1.0727; sorghum, 0.0681, 1.0705; rice, 0.0645, 1.0666, Table 2), indicating that S. inferens is able to survive on the three hosts. The highest T of S. inferens on rice (53.84 d) was significantly longer than on sorghum (49.53 d) and maize (47.31 d). There were no significant differences in GRR.
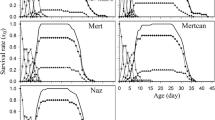
Survival rates of each insect stage of S. inferens feeding on different host plants
The age-stage survival rates (sxj) of S. inferens on maize, sorghum and rice are shown in Fig. 2. The sxj differed between developmental stages, which was due to variable growth rates between individuals. Sesamia inferens completed the preadult stage and adults emerged after 39 days on maize, 43 days on sorghum and 47 days on rice. There were differences in the survival on the three hosts. Female adults survived on maize (17 d) longer than those on sorghum (12 d) and rice (12 d). While, male adults survived longer on rice (13 d) and maize (13 d) than on sorghum (11 d). The peak of sxj in the female adult stage at age of 48 d was higher on rice (32.22%) than at 48 d on sorghum (28.89%) and at 43 d on maize (22.22%). For male adults, the peak of the survival rate (sxj) on rice (23.33%) at 54 d was higher than on sorghum (17.78%, 47 d) and maize (12.22%, 48 d).
Population survival rates and fecundity of S. inferens feeding on different host plants
The age-specific survival rate (lx) on maize, sorghum and rice showed a downward trend with the increase of age are shown in Fig. 3. Based on the estimated values, the last adult fed on rice died at day 59, while this was at day 54 on sorghum and day 55 on maize. On sorghum the age-specific female fecundity (fx2) at 50 d (39.4 offspring/female) was higher than on rice at 56 d (38.0) and maize at 46 d (30.8). Similarly, the age-specific net maternity (lxmx) on sorghum (7.4 offspring/female, 50 d) was higher than on rice (5.6, 52 d) and maize (5.13, 46 d). The highest age-specific population fecundity (mx) was achieved on maize (27 offspring/female, 55 d) and this was significantly greater than on sorghum (23.1, 55 d) and rice (14.9, 56 d).
Life expectancy of each insect stage of S. inferens feeding on different host plants
The value of exj showed a downward trend for maize, sorghum and rice, with maximum average longevity values of 49.16, 50.79, and 55.69 days, respectively (Fig. 4). The exj was highest for females on maize (10.74 d) compared to rice (8.90 d) and sorghum (8.88 d). Also for males, the highest exj was achieved on maize (10.55 d), while for rice it was 10.38 d and 8.81 d for sorghum.
Reproductive values of each insect stage of S. inferens feeding on different host plants
The reproductive values (vxj) of S. inferens feeding on maize, sorghum and rice at age zero (v0,1) were 1.0727, 1.0705, and 1.0666, respectively (Fig. 5). The vxj on sorghum reached a peak of 88.39 d−1 at age 46 d and this was higher than on maize (85.39d−1, 41 d) and rice (81.77 d−1,49 d). The highest vxj was in female adults fed on sorghum.

Survival rates of each insect stage of S. inferens on different host plants

Population survival rates and fecundity of S. inferens on different host plants. lx: population survival rate; fx2: female fecundity; mx: population fecundity; lxmx: population maternity

Life expectancy of each insect stage of S. inferens on different host plants

Reproductive values of each insect stage of S. inferens on different host plants
Discussion
The age-stage, two-sex life table can accurately describe the population variability and adaptability of insects (Zhang et al. 2019; Chi et al. 2020; Ding et al. 2021). Here we applied the age-stage, two-sex life table to describe the population variation of the pink stem borer, S. inferens, known as a major pest in rice, when fed on three different hosts, specifically rice, maize and sorghum. In this study, we found that the adaptability of S. inferens was influenced by the host species.
The difference in nutrient content of host plants has a major effect on the life cycle of herbivorous insects and influences the development of their population (Liu et al. 2018; Alami et al. 2014). Insect development generally depends on the quality of the diet in the larval stage, which was different among the host plants (Barros et al. 2010). However, a longer larva-to-adult period is thought to reflect a compensatory response of larvae to a low-quality diet (Silva et al. 2017). In this study, there were significant differences in the preadult period between the three host plants (rice, 49.34 d; sorghum, 45.33 d; maize, 42.68 d). The reason for this may be the difference in the main nutrients in different host plants (Liu et al. 2009). A similar study showed that larval stage development time differed significantly when S. frugiperda was fed maize, rice, wheat and sorghum, with the longest larval development being on sorghum (19.4 d), while the shortest was on maize (15.4 d) (He et al. 2021).
In general, individuals which develop faster at a certain stage have a lower survival rate at this stage (Ban 1995; Bou Sleiman et al. 2022). One study pointed to the low survival rate of S. inferens in the preadult stage due to the short preadult stage on maize (Feng 2011). In this study, in which S. inferens has a short preadult stage on maize and the survival rate is low, is consistent with previous findings. In addition, the sex ratio of the population is influenced by the nutritional status of the host plant (Wei 2021; Li et al. 2020; Gong et al. 2016). In this study the proportion of adult females of S. inferens that fed on rice was highest. The reason may be that the mortality of S. inferens feeding on rice is lowest in the preadult stage.
The r and λ values of S. inferens populations on maize, rice, and sorghum were > 0 and > 1, respectively, indicating that S. inferens is able to survive on the three hosts. The r represents the growth potential of insect populations; a larger value reflects faster development (Hu et al. 2014; Qin et al. 2017). The low r values of S. inferens populations on all three host plants and the excessive mortality of their larvae to the adult stage can be explained by the fact that S. inferens is subject to translocations in the field and there is no restriction on its feeding by the greater amount of crops in the field (Baladhiya et al. 2018). In contrast, in the cages that we used in the current experiments, while adequate food is continuously provided, the abundance of host resources in the field has not yet been achieved, and in the cage of the life table experiment, there is competition for both food resources and survival space in the limited space available to S. inferens.
Sesamia inferens was originally a major pest on rice, which later migrated to other gramineous crops (Pathak et al. 1994). Rice and wheat are the same major cropping system, and later S. inferens started to emerge as a serious pest in wheat. It was first recorded as a pest in maize by Fletcher in 1914 (Fletcher 1914). It was also widely reported to damage maize in Hebei, China, in 2012 (Chen et al. 2017). But now it has been found to be seriously damaging to sorghum in Guizhou province. The reasons are as follows. First, S. inferens is a typical omnivorous insect with a wide range of hosts (Feng 2012); second, there are many kinds of rice pests, such as Chilo suppressalis (Lepidoptera, Pyralidae) and Scirpophaga incertulas (Lepidoptera, Pyralidae), and S. inferens is competing for niche (Yang et al. 2019); thirdly, maize, rice and sorghum are major crops of Gramineae, and their morphological structure is similar; and last but not least that the prospect of glutinous sorghum for Chinese liquors in Guizhou province is so broad that the local planting structure is changed and the glutinous sorghum planting industry is increasing (Ding et al. 2019), to increase the transmission of S. inferens from rice to sorghum. The results of this study showed that different host plants (sorghum, rice and maize) had effects on the S. inferens population parameters, but the overall difference was not significant. Sesamia inferens is highly adaptable in these three host plants.
The dynamics and growth and development of insect populations are influenced not only by the host (Liang et al. 2019), but also by natural enemies (Zheng et al. 2019), insecticides (Desneux et al. 2007; Guedes et al. 2016; Wang et al. 2022), environmental factors (Lu and Wu 2011; Wang and Ma 2022), and other biotic and abiotic factors (Barteková and Praslička 2018).
The results of our study indicate that S. inferens was able to complete the full developmental cycle on all three host plants. The populations of S. inferens fed on sorghum were more stable compared to those fed on rice and maize. This study further supports that glutinous sorghum is an adaptable host plant for S. inferens. It should be remarked here that the current experiment was performed under specific greenhouse conditions for S. inferens, while the growth, development and reproduction parameters of S. inferens under natural field conditions are influenced by temperature, humidity and natural enemies. In the future, further research is needed in the natural field to better understand the occurrence pattern and to provide a theoretical basis for the integrated prevention and treatment of S. inferens.
Availability of data and materials
All data sets presented in this study are included in the article and can be made available by the authors upon reasonable request.
References
Agrawal AA. Host-range evolution: adaptation and trade-offs in fitness of mites on alternative hosts. Ecology. 2000;81(2):500–8.
Alami S, Naseri B, Golizadeh A, Razmjou J. Age-stage, two-sex life table of the tomato looper, Chrysodeixis chalcites (Lepidoptera: Noctuidae), on different bean cultivars. Arthropod-Plant Interact. 2014;8:475–84. https://doi.org/10.1007/s11829-014-9330-3.
Amir-Maafi M, Chi H, Chen ZZ, Xu YY. Innovative bootstrap-match technique for life table set up. Entomol Generalis. 2022;42(4):597–609. https://doi.org/10.1127/entomologia/2022/1334.
Amiri A, Talebi AA, Zamani AA, Kamali K. Effect of temperature on demographic parameters of the hawthorn red midget moth, Phyllonorycter corylifoliella, on apple. J Insect Sci. 2010;10(1):134. https://doi.org/10.1673/031.010.13401.
Aruna C. Breeding Sorghum for Diverse End Uses. Food Science, Technology, and Nutrition Book. 2018.
Baladhiya HC, Sisodiya DB, Pathan NP. A review on pink stem borer, Sesamia inferens Walker: a threat to cereals. J Entomol Zool Stud. 2018;6(3):1235–9.
Ban Y. Evolution of plant life-history strategies. J Ecol. 1995;(03):33–9.
Banerjee SN, Pramanik LM. The lepidopterous stalk borers of rice and their life cycles in the tropics. In: The major insect pests of the rice plant. 1967. p. 103–24.
Banerjee TC, Ray D. Bioenergetics and growth of the fall armyworm, Spodoptera litura (F.) larvae reared on four host plants. Int J Trop Insect Sci. 1995;16(3/4):317–24.
Barros EM, Torres JB, Ruberson JR, Oliveira MD. Development of Spodoptera frugiperda on different hosts and damage to reproductive structures in cotton. Entomol Exp Appl. 2010;137(3):237–45. https://doi.org/10.1111/j.1570-7458.2010.01058.x.
Barteková A, Praslička J. The effect of ambient temperature on the development of cotton bollworm (Helicoverpa armigera Hübner, 1808). Plant Prot Sci. 2018;42:135–8. https://doi.org/10.17221/2768-PPS.
Bernays EA, Chapman RF. Host-plant selection by phytophagous insects. London: Chapman and Hall; 1994.
Bou Sleiman M, Roy S, Gao AW, Sadler MC, Alvensleben GV, Li H, Auwerx J. Sex-and age-dependent genetics of longevity in a heterogeneous mouse population. Science. 2022;377(6614): eabo3191. https://doi.org/10.1126/science.abo3191.
Caldirola P, Southwood TRE. Ecological methods with particular reference to the study of insect populations. Scientia. 1968;62(103):415.
Chen DF. Investigation of the types of pests and the green control technology of main pests of wine-making sorghum in Jinsha County. Guiyang: Guizhou University; 2019.
Chen LT, Wang YF, Liu L. The first report of Sesamia inferens on summer maize in Guantao County, Hebei Province. J Hebei Agric Sci. 2017;21(6):53–5. https://doi.org/10.16318/j.cnki.hbnykx.2017.06.012.
Chi H. TWOSEX-MSChart: a computer program for the age-stage, two-sex life table analysis. http://140.120.197.173/Ecology/. 2021.
Chi H, You MS, Atlihan R, Smith CL, Kavousi A, Ozgokçe MS, Güncan A, Tuan SJ, Fu JW, Xu YY. Age-stage, two-sex life table: an introduction to theory, data analysis, and application. Entomol Generalis. 2020;40:103–24.
Chi H, Kara H, Özgökçe MS, Atlihan R, Güncan A, Rişvanl MR. Innovative application of set theory, cartesian product, and multinomial theorem in demographic research. Entomol Generalis. 2022a;42(6):863–74.
Chi H, Güncan A, Kavousi A, Gharakhani G, Atlihan R, Özgökçe MS. WOSEX-MSChart: the key tool for life table research and education. Entomol Generalis. 2022b;42(6):845–9.
Desneux N, Decourtye A, Delpuech JM. The sublethal effects of pesticides on beneficial arthropods. Ann Rev Entomol. 2007;52:81–106. https://doi.org/10.1146/annurev.ento.52.110405.091440.
Dhaliwal GS, Jindal V, Dhawan AK. Insect pest problems and crop losses: changing trends. Indian J Ecol. 2010;37(1):1–7. https://doi.org/10.13140/RG.2.2.25753.47201.
Ding YQ, Zhou LB, Wang C, Cao N, Cheng B, Gao X, Zhang LY. Research advance in glutinous sorghum for making sauce-flavor liquor in China. Biotechnol Bull. 2019;35(5):28. https://doi.org/10.13560/j.cnki.biotech.bull.1985.2019-0080.
Ding HY, Lin YY, Tuan SJ, Tang LC, Ch H, Atlıhan R, Güncan A. Integrating demography, predation rate, and computer simulation for evaluation of Orius strigicollis as biological control agent against Frankliniella intonsa. Entomol Generalis. 2021;41:179–96. https://doi.org/10.1127/entomologia/2020/1082.
Fan QG, Lin H, Zhang J. Development status of scientific and technological innovation in Baijiu industry. Liquor-Mak Sci Technol. 2022;11:82–8.
Feng XH. Studies on the developmental characteristics and fungicides of the Sesamia inferens (Walker) on different host plants. Nanjing: Nanjing Agricultural University; 2011.
Feng CY. Research progress on occurrence rules and control technique of Sesamia inferens Walker in rice field in China. Agric Eng. 2012;2(10):62–5.
Fletcher TB. Some south Indian insects and other animals of importance: considered especially from an economic point of view/by T. Bainbrigge Fletcher. Dehra Dun: Bishen Singh Mahendra Pal Singh; 1914.
Gao J, Feng GC, Li XR. Phenotypic diversity and cluster analysis of sorghum germplasm resources in different regions of Guizhou Province. Crop J. 2020;2(6):54–60.
Gong Q, Geng J, Zhang K, Wu H, Liu X, Sun R. Phenotypes and sex ratio changes of Harmonia axyridis (Pallas) in Taian. J Environ Entomol. 2016;38(2):313–23.
Guedes RNC, Smagghe G, Stark JD, Desneux N. Pesticide-induced stress in arthropod pests for optimized integrated pest management programs. Ann Rev Entomol. 2016;61:43–62.
Guo JF, Zhang MD, Gao ZP. Comparison of larval performance and oviposition preference of Spodoptera frugiperda among three host plants: potential risks to potato and Tobacco crops. Insect Sci. 2020. https://doi.org/10.1111/1744-7917-12830.
Hashmi A. Insect pest management, vol. 1. Islamabad: Pakistan Agricultural Research Council; 1994. p. 14–7.
He LM, Wang TL, Chen YC, Ge SS, Wyckhuys KA, Wu KM. Larval diet affects development and reproduction of east Asian strain of the fall armyworm, Spodoptera frugiperda. J Integr Agric. 2021;20(3):736–44. https://doi.org/10.1016/s2095-3119(19)62879-0.
Hu L, He Z, Zhang X. Age-stage two-sex life tables of the experimental population of Problepsis superans (Lepidoptera: Geometridae) on three Ligustrum species. Acta Entomol Sin. 2014;57(12):1408–17.
Jeevanandham N, Ramiah N, Chockalingam V, Jegadeesan R. An overview of the bionomics, host plant resistance and molecular perspectives of Sesamia inferens Walker in cereals and millets. Agronomy. 2020;10(11): 1705.
Khan ZR, Litsinger JA, Barrion AT. World bibliography of rice stem borers. Los Banos: International Rice Research Institute; 1991. p. 1–426.
Li WJ, Li XC, Li X, Cui J, Xu W, Zhang JP, Shi SS. Effect of sex ratio on fecundity and lifetime of Leguminivora glycinivorella adults. Soybean Sci. 2020;39(3):451–7.
Liang HY, Zhang HY. Effects of resistant and susceptible varieties of mungbean on life parameters of aphid population. Chin J Plant Prot. 2019;39(10):5–9.
Liu J, Li JW, Lan H. Analysis of the nutritional composition in different plant parts of yacon. Acta Pratacult Sin. 2009;18(4):236–40.
Liu Y, Li G, Yang L. Demography and mass rearing of the medicinal blister beetle Epicauta impressicornis (Pic) (Coleoptera: Meloidae) at different temperatures. J Econ Entomol. 2018;111:2364–74.
Lozano R, Gazave E, Dos Santos JPR, et al. Comparative evolutionary genetics of deleterious load in sorghum and maize. Nat Plants. 2021;7(1):17–24. https://doi.org/10.1038/s41477-020-00834-5.
Lu YH, Wu KM. Effect of relative humidity on population growth of Apolygus lucorum (Heteroptera: Miridae). Appl Entomol Zool. 2011;46(3):421–7. https://doi.org/10.1007/s13355-011-0058-6.
Marina R, Nur Azura A, Lau WH, Yaakop S. Life table and demographic parameters of rice leaf folder, Cnaphalocrocis medinalis Guenee (Lepidoptera: Pyralidae). 2019.
Pathak MD, Khan ZR. Insect pests of rice. Los Banos: International Rice Research Institute; 1994. p. 5–17.
Qin JY, Zhang L, Chen YX. Age-stage two-sex life table for laboratory populations of oriental armyworm Mythimna separata (Walker) under different temperatures. J Plant Prot. 2017;44(5):729–36.
Saska P, Skuhrovec JAR, Ozgokçe MS, Güncan A, Zamani AA, Tuan SJ. Bias introduced by the simplified method for the estimation of the intrinsic rate of increase of aphid populations: a meta-analysis. Entomol Generalis. 2021;41:305–16.
Sau AK. Aspects of host plant resistance with reference to Sesamia inferens (Walker): a review. Pharma Innov J. 2022;11(2):1006–10.
Shahidi-Noghabi S, Van Damme EJM, Smagghe G. Expression of Sambucus nigra agglutinin (SNA-I′) from elderberry bark in transgenic tobacco plants results in enhanced resistance to different insect species. Transgenic Res. 2009;18:249–59.
Silva DM, Bueno AF, Andrade K. Biology and nutrition of Spodoptera frugiperda (Lepidoptera: Noctuidae) fed on different food sources. Sci Agric. 2017;74(1):18–31. https://doi.org/10.1590/1678-992x-2015-0160.
Veenstra KH, Pashley DP, Ottea JA. Host-plant adaptation in fall armyworm host strains: comparison of food consumption, utilization, and detoxication enzyme activities. Ann Entomol Soc Am. 1995;88(1):80–91.
Wang XJ, Ma CS. Challenge generality of prediction based on Jensen’s inequality: moderate and large temperature fluctuations can lead to opposite performance deviation at high mean temperature. Entomol Generalis. 2022;42(5):681–9.
Wang SY, Chi H, Liu TX. Demography and parasitic effectiveness of Aphelinus asychis reared from Sitobion avenae as a biological control agent of Myzus persicae reared on Chili pepper and cabbage. Biol Control. 2016;92:111–9.
Wang WQ, Zheng YQ, Chen B. Effects of different hosts on growth and development and fecundity of potato tuber moths based on age-stage hermaphroditic life table. J Plant Prot. 2022;47(3):488–96.
Wei P, Li Y, Zhang W. Factors affecting the ratio of female to male of pinewood nematodes Bursaphelenchus xylophilus (Steiner and Buhrer) Nickle. Forests. 2021;12(6): 763.
Yan SX, Lu YH, Wang L. Germplasm formation and development of Baijiu-making sorghum in the southwest of China. Forum Summ. 2017;36(5):17–21.
Yang SW, Zhang XM, Guo HY. Dynamics and temporal niches of major pest insects and their natural enemies in perennial rice fields. Chin J Appl Entomol. 2019;56(6):1370–81.
Zhang S, Zhang HB, Dong WJ. The main uses and development prospects of sorghum. Agric Technol. 2014;34(7):111.
Zhang Y, Guo L, Atlihan R, Chi H, Chu D. Demographic analysis of progeny fitness and timing of resurgence of Laodelphax striatellus after insecticides exposure. Entomol Generalis. 2019;39(3–4):221–30. https://doi.org/10.1127/entomologia/2019/0816.
Zhao Y, Zhao CL, Yang X, Chi H, Dai P, Desneux N, Zang LS. Yacon as an alternative host plant for Encarsia formosa mass-rearing: validating a multinomial theorem for bootstrap technique in life table research. Pest Manag Sci. 2021;77(5):2324–36. https://doi.org/10.1002/ps.6259.
Zheng LL, Chen JX, Wang KK. Study on the life table of experimental populations of hooded gourd blunt Sui mite predation on tea yellow thrips. Guangdong Agric Sci. 2019;46(6):93–8.
Zhu HF, Fang CL, Wang LY. The economic insects of China, vol. 7. Lepidoptera, Noctuidae. Beijing: Science Press; 1963.
Acknowledgements
We are grateful to Hsin Chi, National Chung Hsing University, for the fruitful discussions and helpful comments on an earlier draft and instruction of his ecology software based on MSChart.
Funding
This research was funded by the National Key R&D Program of China (2023YFE0104800), the Natural Science Research Program of Guizhou University (202202), and Program of Introducing Talents to Guizhou University (600183233301).
Author information
Authors and Affiliations
Contributions
Conceptualization, L-SZ and M-YM; methodology, CL, L-DT and L-SZ; investigation, CL, and Q-YH; validation, CL and ZS; writing—original draft preparation, CL, GS and Y-FZ; writing—review and editing, ZS, L-DT and L-SZ; project administration, M-YM and L-SZ; funding acquisition, L-SZ and L-DT. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Table S1.
Definitions and formulas of population parameters estimated in the present study.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Li, C., Shen, Z., Tang, LD. et al. Demography and fitness of Sesamia inferens Walker (Lepidoptera: Noctuidae) on three important gramineous crops. CABI Agric Biosci 4, 49 (2023). https://doi.org/10.1186/s43170-023-00191-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43170-023-00191-1