
Abstract
Background
The identification of miRNAs as well as characterization of miRNA-mRNA interactions in SARS-CoV-2 infection is important to understand their role in disease pathogenesis. Therefore the aim of the present study was to measure the expression levels of hsa-mir-18a-5p in the sera of severe COVID-19 Egyptian patients admitted to ICU to investigate its roles in the pathogenesis and severity of COVID-19 disease.
Methods
A total of 180 unvaccinated severe COVID-19 patients were enrolled in our study. Besides the routine laboratory work, the expression level of hsa-mir-18a-5p was done using reverse transcription quantitative real-time PCR (RTqPCR) technique. Also, target genes of hsa-mir-18a-5p were explored by using online bioinformatics databases.
Results
The expression level of hsa-mir-18a-5p decreased in nonsurvival severe COVID-19 patients (0.38 ± 0.26) when compared to the survival ones (0.84 ± 0.23). While as a prognostic tool for the prediction of bad prognosis and mortality among severe COVID-19 patients, our results showed that the serum hsa-mir-18a-5p expression level is a good sensitive and specific marker. By using bioinformatics tools, our results revealed that the decreased hsa-mir-18a-5p expression level may have a crucial role in COVID-19 pathogenesis and severity through decreased immunological responses (interpreted as lymphopenia) or increased inflammation (interpreted as increased serum levels of IL-6, CRP, LDH).
Conclusion
Taken together, the decreased expression level of hsa-mir-18a-5p could be a bad prognostic marker and therapeutic overexpression of hsa-mir-18a-5p could be a novel approach in the treatment of COVID-19 disease.
Similar content being viewed by others

Introduction
Due to the high prevalence and long incubation periods, the severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2) has infected millions of individuals globally, causing the coronavirus disease 2019 (COVID-19) pandemic [1]. It was first emerged in Wuhan, China in December 2019 and declared a global pandemic by WHO on 11 March 2020 [2, 3]. It has become a catastrophic public health crisis affecting many people as of January 09, 2023, there have been 668,820,532 confirmed cases, and 6,714,775 deaths were reported in more than 229 countries [4]. The clinical presentation of COVID-19 varies so much from asymptomatic to milder symptoms, including dry cough, fever, myalgia, dyspnea, sore throat, and headache, or even to severe and emergent manifestations including chest pain, confusion, hypoxia, pneumonia, and other complications requiring intensive care unit (ICU) admission and mechanical ventilation [5]. The clinical guidelines of WHO define “severe COVID-19” as patients with clinical signs of pneumonia (fever, cough, dyspnea, and fast breathing) accompanied by one of the following: severe respiratory distress; O2 saturation (SpO2) ≤ 90% in room air; or respiratory rate > 30 breaths/min [6]. Till now, the precise determinants of severe COVID-19 are not known, but it primarily maybe host factors rather than viral genetic mutations [7]. The number of COVID-19 patients is continually increasing worldwide and the management in ICU has become a major challenge; therefore, early recognition of severe forms of COVID-19 is very necessary for triaging of COVID-19 patients [8].
MicroRNAs (miRNAs) are a class of highly conserved endogenous small (18–22 nt) noncoding single-stranded RNA molecules widely found in plants, animals, and some viruses. They have an essential role in post-transcriptional regulation of gene expression by targeting the mRNAs of protein-coding genes [9]. MiR-18a-5p is located at chromosome 13q31.3 and belongs to Mirc1 locus, better known as the miR-17–92 cluster, encodes six miRNAs (miR-17-5p, miR-18a-5p, miR-19a-3p, miR-19b-3p, miR-20a-5p, and miR-92a-3p); which has important roles in cell proliferation and differentiation, inflammation, immunity and immunological process [10, 11]. It was reported that miRNAs have been shown to have a role in viral infections as viruses can induce the up- or downregulation of various host miRNAs to elude the host’s immune system [12]. It was found that miR-18a-5p was reduced in bleomycin-treated pleural mesothelial cells (PMCs) which in turn contributes to epithelial-mesenchymal transition (EMT) of PMCs via upregulation of its target, TGF-β receptor II (TGF-βRII), which mediates signaling leading to sub-pleural pulmonary fibrosis [13]. Therefore the aim of the present study was to measure the expression level of hsa-mir-18a-5p in the sera of unvaccinated severe COVID-19 Egyptian patients admitted to ICU and then examine the target genes of has-mir-18a-5p using bioinformatics online tools to investigate its role in the pathogenesis and severity of COVID-19 disease also its correlations with other clinical variables in severe COVID-19 patients.
Patients and methods
Ethics statement
The current study was approved by the ethics committee of the Faculty of Medicine, Port-Said University, Egypt (ERN MED (23/04/2020)S.no(5)MED). Informed consent was obtained from all patients.
Human subjects and data collection
The current study was conducted on 180 unvaccinated severe COVID-19 patients recruited from the isolation hospitals in Port-Said, Egypt. Sputum and throat swab specimens (for qPCR for SARS-Cov-2 RNA test) and blood samples were collected from all patients. Laboratory tests were conducted at admission, including a complete blood count, liver function tests (ALT and AST), kidney function tests (urea and creatinine), CRP, ferritin, IL-6, D-dimer, PCT, and LDH. Also, chest CT scans are made for all patients. The severity of COVID-19 was graded according to Suspected COVID-19 Cases Management in Triage Hospitals by the Ministry of Health and Population of Egypt. Our Severe COVID-19 patients were defined as patients with respiratory distress, resting oxygen saturation ≤ 90%, respiratory failure requiring mechanical ventilation, or failure of other organs requiring ICU admission.
Data from severe patients were collected from the latest laboratory tests prior to physicians making the clinical diagnosis of severe disease. Demographic data, hospitalization time, medical history, oxygen saturation, respiratory rate, oxygen supply, laboratory findings, and thorax tomography of the patients were obtained from the hospital’s electronic information system retrospectively. All patients were treated by meropenem (1 gm/8 h intravenous), levofloxacin (500 mg vial/24 h, intravenous), linezolid (600 mg vial/8 h, intravenous), Enoxaparine calcium (therapeutic dose, subcutaneous), methylprednisolone sodium succinate (1 gm every 24 h for 3 days then 125 mg every 12 h), tocilizumab (if needed, 8 mg/kg), and remdesivir (loading dose 400 mg first day, then 200 mg for 5 days).
Determination of serum miR-18a expression level by RT-qPCR
MiRNA extraction and cDNA preparation
The miRNA was extracted from the sera of all patients using miRNeasy Mini kit (cat # 217004, Qiagen, USA) according to the manufacturer’s instructions. The purity and the concentration of the purified miRNA were detected using spectrophotometer nano-drop (Quawell, Q-500, Scribner, USA) and stored at − 80 °C till further assessments. To synthesize cDNA, miRNA was reverse transcribed using MiScript II reverse transcription kit (cat # 218160, Qiagen, USA) according to manufacturer’s instructions and stored at − 20 °C till performing qPCR.
Quantitative real-time PCR (qPCR)
Quantitative real-time PCR was performed using miScript primer assay (cat # 218300, Qiagen, USA) for miR-18a (Hs_miR-18a_2 miScript Primer Assay, MS00031514); the reaction was carried out using MiScript SYBR Green PCR kit (cat # 218073, Qiagen, USA). Also, RNU6–2 (Hs_RNU6-2_11 miScript Primer Assay, MS00033740) was used as an endogenous control to normalize the expression levels of the investigated miRNAs; the primer sequences are listed in Table 1. The qPCR cycling conditions were as follows: 95 °C for 10 min, followed by 40 cycles of 95 °C for 15 s, 55 °C for 30 s, and 72 °C for 30 s in which fluorescence was acquired and detected by Stratagene Real-time PCR system (Max3005P QPCR system, Stratagene, Agilent biotechnology, USA). The relative expression levels of the investigated miRNAs were evaluated using the 2−ΔΔCq method described by Livak and Schmittgen [14]. A 2-fold increased (≥ 2) or decreased (≤ 0.5) value was considered mRNA overexpression or downregulation, respectively.
Bioinformatics analysis
To examine the target genes of has-mir-18a-5p, different online databases were used, miRDB (https://mirdb.org/mirdb/index.html), TargetScan (https://www.targetscan.org/vert_80/), DIANA-TarBase(https://dianalab.e-ce.uth.gr/html/diana/web/index.php?r=tarbasev8%2Findex/), miRwalk (http://mirwalk.umm.uni-heidelberg.de/), miRNet (https://www.mirnet.ca/miRNet/home.xhtml/).
Statistical analysis
Statistical analysis was performed using IBM SPSS software (version 23.0; IBM Corp., Armonk, NY, USA), and data were presented as means ± S.D. One-way ANOVA was used to determine statistically significant differences between group’s means and Pearson’s correlation coefficient was used to determine significant correlations of serum has-mir-18a-5p expression level with other clinical parameters. The receiver operating characteristic curve (ROC curve) was used to calculate the area under the curve (AUC), sensitivity, and specificity of serum has-mir-18a-5p expression level as a biomarker for the detection of bad prognosis and deterioration of severe COVID-19 disease. The criterion for significance was p < 0.05.
Results
Demographic and biochemical data of COVID-19 patients
The present study included 180 severe COVID-19 patients; 108 males and 72 females; with mean age 67.4 ± 9.6 years; oxygen saturation 86.32 ± 4.05%; respiratory rate 26.83 ± 3.17 cycle/min; and 74 (41.1%) patients died during hospitalization; the clinical and biological data of severe COVID-19 patients are summarized in Table 1. The results of current study revealed a highly significant (p < 0.001) increase in the levels of CRP (76.29 ± 25.50), IL-6 (383.49 ± 213.84), PCT (0.82 ± 0.35), and LDH (480.43 ± 96.08) in the sera of nonsurvival severe COVID-19 patients when compared to survival severe COVID-19 patients (59.81 ± 16.69; 63.75 ± 44.71; 0.19 ± 0.14 and 273.54 ± 99.68; respectively). Also, the blood neutrophils percentage (78.61 ± 6.13) was significantly (p < 0.001) increased while lymphocytes percentage (11.63 ± 5.35) was significantly (p < 0.001) decreased among nonsurvival severe COVID-19 patients as compared with survival severe COVID-19 patients (69.12 ± 14.62 and 19.91 ± 4.94; respectively); as shown in Table 2.
Serum has-mir-18a-5p expression level and receiver operating characteristic (ROC) curves analysis
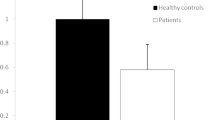
Our results showed that serum has-mir-18a-5p had a differential expression pattern, as it was found to be highly significantly (p < 0.001) decreased in nonsurvival COVID-19 patients (0.38 ± 0.26) when compared to the survival ones (0.84 ± 0.23); as shown in Fig. 1A. The sensitivity and specificity as biomarker of serum has-mir-18a-5p expression level for the prediction of bad prognosis and mortality among severe COVID-19 patients were evaluated by using ROC curve analysis. Our results showed that it is a good biomarker that could predict a bad prognosis of severe COVID-19 patients, with AUC 0.91, 92.7% sensitivity, and 84.5% specificity; as shown in Fig. 1B.

Serum hsa-mir-18a-5p expression level among severe COVID-19 patients: A relative gene expression of hsa-mir-18a-5p in nonsurvival and survival severe COVID-19 patients. B ROC curve of serum hsa-mir-18a-5p expression level in discrimination between survival and non-survival severe COVID-19 patients
Correlation of serum has-mir-18a-5p expression level with clinical variables in severe COVID-19 patients
Data recorded in Table 3 shows the correlation matrix of serum has-mir-18a-5p expression level with the different clinical parameters in this study. It was found that it was highly significantly positively correlated (p < 0.001) with lymphocyte percentage, while it was highly significantly negatively correlated (p < 0.001) with neutrophils percentage, CRP, IL-6, PCT, and LDH; as shown in Table 3.
Bioinformatics analysis
Different online databases were used to investigate the target genes of hsa-mir-18a5p. Different numbers of target genes were obtained, miRDB (382), TargetScan (321), DIANA-TarBase (1053), miRwalk (222), and miRNet (262); as shown in Fig. 2A. To visualize the target genes of has-mir-18a-5p as a figure, authors used miRwalk database (Fig. 2B), while to predict its possible roles or pathways in the COVID-19 pathogenesis and severity, miRNet database was used. According to miRNet database, our results showed that hsa-mir-18a5p has a role in the adaptive immune system by targeting CANX, FCGR2B, PSMB5, PTEN, RAP1A,UBC, EC24B, DCTN2, TNRC6B, PHLPP1, SEC61A1, DCTN5, and RICTOR genes; as shown in Fig. 3A. Also it has signaling events of B cell receptor (BCR) by targeting PSMB5, PTEN, UBC, TNRC6B, PHLPP1, and RICTOR genes; as shown in Fig. 3B. Moreover, hsa-mir-18a5p mediated Class I MHC antigen processing and presentation through CANX, PSMB5, UBC, SEC24B, and SEC61A1 genes; as shown in Fig. 3C. Furthermore, it has a crucial role in interferon gamma signaling by targeting IRF2, MID1, and SP100 genes; as shown in Fig. 3D. Other than the immunological responses, hsa-mir-18a5p has a role in inflammation (by targeting BCL2 and TXNIP genes; as shown in Fig. 4).

Network analysis of hsa-mir-18a-5p target gene networks: A numbers of target genes of hsa-mir-18a-5p. B Visualization of target genes of hsa-mir-18a-5p using miRWalk online database

Target genes of hsa-mir-18a-5p in immunological responses: A adaptive immune system. B Signaling events of B cell receptor (BCR). C Class I MHC mediated antigen processing and presentation. D Interferon gamma signaling

Target genes of hsa-mir-18a-5p in inflammation
Discussion
It is well known that miRNAs play an important role in the posttranscriptional control of gene expression that is dysregulated in different physiological pathophysiological processes, such as metabolism, growth, cell differentiation and development, apoptosis, inflammation, and cell signaling [15]. Besides, the role of miRNAs in the pathogenesis of lung disease has been recognized, as Dakhlallah et al. [16] reported that downregulation of miR-17∼92 contributes to the pathogenesis of pulmonary fibrosis, and also miR-18a levels are extraordinarily decreased in the lung of human IPF patients [16, 17]. However, the mechanisms involved in the regulatory effects of miRNA in pulmonary fibrosis have not been revealed. Therefore, we aimed to measure the expression levels of has-mir-18a-5p in the sera of unvaccinated severe COVID-19 Egyptian patients admitted to ICU to investigate its roles in the pathogenesis and severity of COVID-19 disease also its correlations with other clinical variables in severe COVID-19 patients. In our previous work, we measured the levels of serum IP-10, SAA, and sialic acid and circulating plasma has-mir-155-5p in positive COVID-19 patients to explore their clinical values and significance in discrimination between moderate and severe COVID-19 infection and predicting the severity and prognosis of COVID-19 disease [18,19,20,21]. Here, our results revealed that the expression level of serum has-mir-18a-5p was significantly decreased in nonsurvival COVID-19 patients when compared to the survival ones (Fig. 1A). As consistent with our results; Li et al. found that the level of miR-18a is down-expressed in the peripheral blood from human patients with COVID-19 [22], also other several studies reported miR-18a down expression in asthma patients [23, 24]. Moreover, Ventura et al. [25] reported that the loss-of-function of the miR-17-92 cluster resulted in smaller embryos and immediate postnatal death of all animals due to severely hypoplastic lungs of mice lacking miR-17-92, indicating the vital role of miR-18a in the proper function of lungs. Several studies reported the diagnostic and prognostic significance of serum mir-18a-5p [26,27,28]. Therefore, the current study also aimed to elaborate and assess the potential role of serum mir-18a-5p as a prognostic biomarker for the prediction of bad prognosis and mortality among severe COVID-19 patients. Our results showed that it is a good biomarker that could predict the bad prognosis of severe COVID-19 patients, with AUC 0.91, 92.7% sensitivity, and 84.5% specificity (Fig. 1B).
The innate immune response to SARS-CoV-2 Antiviral innate immunity has several humoral components, including mannose-binding lectin, interferons, chemokines, B lymphocytes, natural killer cells, and other innate lymphoid cells (ILCs) and gamma delta T cells, which generally limit the spread of viral infection by cytotoxic action on target cells, cytokine production, and induction of an adaptive response [29]. By using bioinformatics online tools, it was found that hsa-mir-18a-5p targets many genes, which may be involved in many pathways such as immunological responses (Fig. 3A–D). Therefore, as the expression level of hsa-mir-18a-5p decreased, these immunological responses such as adaptive immune responses (Fig. 3A), signaling events of B cell receptor (Fig. 3B), class I MHC mediated antigen processing and presentation (Fig. 3C) and interferon-gamma signaling (Fig. 3D) were decreased. These observations are in line with the relative lymphopenia reported in severe COVID-19 and also in our patients. It is well known that hsa-mir-18a-5p regulates the immunological responses, especially in respiratory diseases as in a study about influenza A, miR-18a-5p was found to be involved in the regulation of the pulmonary innate immune response [30]. Moreover, several viruses, including SARS-CoV-2 have been reported to enhance TGF-β signaling, which is known to induce fibrosis and suppress adaptive immunity through a modulation of TGF-β signaling, via the surface receptors and canonical SMAD and MAPK pathways regulated by hsa-mir-18a-5p regulating adaptive immune responses [31].
The pathogenesis of COVID-19 is complex, but it can be conceptually described using typical models for the three main pathological processes associated with inflammation—local manifestations of classical general (canonical) inflammation, acute systemic inflammation, and chronic systemic inflammation of low intensity [32]. Our bioinformatics analysis results showed that hsa-mir-18a5p has an inhibitory effect on inflammation (Fig. 4), which in turn may play crucial roles in COVID-19 pathogenesis and severity.
As consistent with our results, it was indicated that miR-18a-5p mimic significantly reduced inflammatory factors including IL-6, IL-8, IL-1β, and tumor necrosis factor (TNF)-α release, decreased the degranulation rate and histamine release rate of cells [33], this may explain the increased serum level of IL-6 in our severe COVID-19 patients, especially the nonsurvival ones. Moreover, Geng et al. [34] observed that has-mir-18a-5p upregulation prevents endothelial-mesenchymal transition and cardiac fibrosis induced by high glucose concentration, by targeting NOTCH2 gene, which regulates cellular phenotype. Our results also revealed that has-mir-18a-5p showed very significant correlations with the other parameters by Pearson correlation analysis, which suggested that it was a significant factor associated with the severity of patients with COVID-19.
There are some limitations that need to be addressed regarding the present study. First of all, the population was only from Egypt, which reduces the possibility of confounding by ethnicity; therefore, these results should be interpreted with caution. Second, given the limited size of the study and additional large-scale studies are needed to confirm this finding. Finally, serum levels or gene expression levels of target genes of has-mir-18a-5p should be measured concurrently with has-mir-18a-5p levels in the same patients’ samples to ensure their interactions.
Conclusion
Finally, by using survival severe COVID-19 patients as a control group our results showed that the expression level of hsa-mir-18a-5p was significantly decreased among the nonsurvival severe COVID-19 patients. While as a prognostic tool for the prediction of bad prognosis and mortality among severe COVID-19 patients, our results showed that the serum hsa-mir-18a-5p expression level is a good sensitive and specific marker. By using bioinformatics tools, our results revealed that the decreased hsa-mir-18a-5p expression level may have a crucial role in COVID-19 pathogenesis and severity through decreased immunological responses (interpreted as lymphopenia) or increased inflammation (interpreted as increased serum levels of IL-6, CRP, LDH). This provides proof of concept that the therapeutic overexpression of hsa-mir-18a-5p could be a novel approach in the treatment of COVID-19 disease.
Availability of data and materials
All data generated or analyzed during this study are included in this published article.
Abbreviations
- AUC:
-
Area under the curve
- BCL2:
-
B cell lymphoma 2
- CANX:
-
Calnexin
- COVID-19:
-
Coronavirus disease 2019
- CRP:
-
C-reactive protein
- DCTN2:
-
Dynactin subunit 2
- DCTN5:
-
Dynactin 5 (p25)
- FCGR2B:
-
Fc fragment of IgG receptor IIb
- ICU:
-
Intensive care unit
- IL-6:
-
Interleukin 6
- IRF2:
-
Interferon regulatory factor 2
- LDH:
-
Lactate dehydrogenase
- MAPK:
-
Mitogen-activated protein kinase
- MID1:
-
Midline-1
- miRNAs:
-
MicroRNAs
- NOTCH2:
-
Neurogenic locus notch homolog protein 2
- PCT:
-
Procalcitonin
- PHLPP1:
-
PH domain and leucine-rich repeat protein phosphatases
- PSMB5:
-
Proteasome subunit beta type-5
- PTEN:
-
Phosphatase and tensin homolog
- RAP1A:
-
Ras-related protein Rap-1A
- RICTOR:
-
Rapamycin-insensitive companion of mammalian target of rapamycin
- ROC curve:
-
Receiver Operating Characteristic curve
- SARS-CoV-2:
-
Severe acute respiratory syndrome coronavirus-2
- SEC24B:
-
SEC24 Homolog B
- SEC61A1:
-
SEC61 Translocon Subunit Alpha 1
- SP100:
-
Space reactor prototype
- SpO2:
-
O2 saturation
- TNRC6B:
-
Trinucleotide repeat-containing gene 6B protein
- TXNIP:
-
Thioredoxin-interacting protein
- UBC:
-
Ubiquitin C
- WHO:
-
World Health Organization
References
Chung JY, Thone MN, Kwon YJ (2021) COVID-19 vaccines: the status and perspectives in delivery points of view. Adv Drug Deliv Rev 170:1–25
WHO Director-General’s opening remarks at the media briefing on COVID-19 – 11 March 2020: World Health Organization; 2020. Available: https://www.who.int/directorgeneral/speeches/detail/who-director-general-s-opening-remarks-at-the-media-briefingon-covid-19%2D%2D-11-march-2020. [Accessed 26 Jun 2021].
Huang C, Wang Y, Li X, Ren L, Zhao J, Hu Y, Zhang L, Fan G, Xu J, Gu X, Cheng Z, Yu T, Xia J, Wei Y, Wu W, Xie X, Yin W, Li H, Liu M, Xiao Y, Gao H, Guo L, Xie J, Wang G, Jiang R, Gao Z, Jin Q, Wang J, Cao B (2020) Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 395(10223):497–506
COVID-19 coronavirus pandemic: Available: https://www.worldometers.info/coronavirus/ [Accessed January 09, 2023, 18:49 GMT].
Zaim S, Chong JH, Sankaranarayanan V, Harky A (2020) COVID-19 and Multiorgan Response. Curr Probl Cardiol 45(8):100618
Gallo Marin B, Aghagoli G, Lavine K, Yang L, Siff EJ, Chiang SS, Salazar-Mather TP, Dumenco L, Savaria MC, Aung SN, Flanigan T, Michelow IC (2021) Predictors of COVID-19 severity: A literature review. Rev Med Virol 31(1):1–10
Zhang X, Tan Y, Ling Y, Lu G, Liu F, Yi Z, Jia X, Wu M, Shi B, Xu S, Chen J, Wang W, Chen B, Jiang L, Yu S, Lu J, Wang J, Xu M, Yuan Z, Zhang Q, Zhang X, Zhao G, Wang S, Chen S, Lu H (2020) Viral and host factors related to the clinical outcome of COVID-19. Nature 583(7816):437–440
Velavan TP, Meyer CG (2020) Mild versus severe COVID-19: Laboratory markers. Int J Infect Dis 95:304–307
Fu X, Liu P, Dimopoulos G, Zhu J (2020) Dynamic miRNA-mRNA interactions coordinate gene expression in adult Anopheles gambiae. PLoS Genet 16(4):e1008765
Zhang N, Zhang H, Liu Y, Su P, Zhang J, Wang X, Sun M, Chen B, Zhao W, Wang L, Wang H, Moran MS, Haffty BG, Yang Q (2019) SREBP1, targeted by miR-18a-5p, modulates epithelial-mesenchymal transition in breast cancer via forming a co-repressor complex with Snail and HDAC1/2. Cell Death Differ 26(5):843–859
Sheng X, Yang Y, Liu J, Yu J, Guo Q, Guan W, Liu F (2022) Down-regulation of miR-18b-5p protects against splenic hemorrhagic shock by directly targeting HIF-1α/iNOS pathway. Immunobiology 227(2):152188
Barbu MG, Condrat CE, Thompson DC, Bugnar OL, Cretoiu D, Toader OD, Suciu N, Voinea SC (2020) MicroRNA involvement in signaling pathways during viral infection. Front Cell Dev Biol 8:143
Zhang Q, Ye H, Xiang F, Song LJ, Zhou LL, Cai PC, Zhang JC, Yu F, Shi HZ, Su Y, Xin JB, Ma WL (2017) miR-18a-5p inhibits sub-pleural pulmonary fibrosis by targeting TGF-β receptor II. Mol Ther 25(3):728–738
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 25:402–408
Ebrahimi A, Sadroddiny E (2015) MicroRNAs in lung diseases: Recent findings and their pathophysiological implications. Pulm Pharmacol Ther 34:55–63
Dakhlallah D, Batte K, Wang Y, Cantemir-Stone CZ, Yan P, Nuovo G, Mikhail A, Hitchcock CL, Wright VP, Nana-Sinkam SP, Piper MG, Marsh CB (2013) Epigenetic regulation of miR-17~92 contributes to the pathogenesis of pulmonary fibrosis. Am J Respir Crit Care Med 187(4):397–405
Pandit KV, Corcoran D, Yousef H, Yarlagadda M, Tzouvelekis A, Gibson KF, Konishi K, Yousem SA, Singh M, Handley D, Richards T, Selman M, Watkins SC, Pardo A, Ben-Yehudah A, Bouros D, Eickelberg O, Ray P, Benos PV, Kaminski N (2010) Inhibition and role of let-7d in idiopathic pulmonary fibrosis. Am J Respir Crit Care Med 182(2):220–229
Haroun RA, Osman WH, Eessa AM (2021) Interferon-γ-induced protein 10 (IP-10) and serum amyloid A (SAA) are excellent biomarkers for the prediction of COVID-19 progression and severity. Life Sci 15(269):119019
Haroun RA, Osman WH, Amin RE, Hassan AK, Abo-Shanab WS, Eessa AM (2022) Circulating plasma miR-155 is a potential biomarker for the detection of SARS-CoV-2 infection. Pathology 54(1):104–110
Haroun RA, Osman WH, Eessa AM (2022) Evaluation of serum total sialic acid in moderate COVID-19 patients with and without gastrointestinal tract manifestations. Tissue Cell 74:101679
Haroun RA, Osman WH, Amin RE, Eessa AM, Saad S (2023) Increased serum interleukin-6 and lactate dehydrogenase levels among nonsurvival severe COVID-19 patients when compared to survival ones. Int Immunopharmacol 15(122):110626
Li C, Hu X, Li L, Li J (2020) Differential microRNA expression in the peripheral blood from human patients with COVID-19. J Clin Lab Anal 34:e23590. https://doi.org/10.1002/jcla.23590
Suojalehto H, Lindstrom I, Majuri ML, Mitts C, Karjalainen J, Wolff H, Alenius H (2014) Altered microRNA expression of nasal mucosa in long-term asthma and allergic rhinitis. Int Arch Allergy Immunol 163:168–178
Martinez-Nunez RT, Bondanese VP, Louafi F, Francisco-Garcia AS, Rupani H, Bedke N, Holgate S, Howarth PH, Davies DE, Sanchez-Elsner T (2014) A microRNA network dysregulated in asthma controls IL-6 production in bronchial epithelial cells. PLoS ONE 9(10):e111659
Ventura A, Young AG, Winslow MM, Lintault L, Meissner A, Erkeland SJ, Newman J, Bronson RT, Crowley D, Stone JR, Jaenisch R, Sharp PA, Jacks T (2008) Targeted deletion reveals essential and overlapping functions of the miR-17 through 92 family of miRNA clusters. Cell 132(5):875–886
Morimura R, Komatsu S, Ichikawa D, Takeshita H, Tsujiura M, Nagata H, Konishi H, Shiozaki A, Ikoma H, Okamoto K, Ochiai T, Taniguchi H, Otsuji E (2011) Novel diagnostic value of circulating miR-18a in plasma of patients with pancreatic cancer. Br J Cancer 105(11):1733–1740
Teng P, Liu Y, Zhang M, Ji W (2021) Diagnostic and Prognostic Significance of serum miR-18a-5p in Patients with Atherosclerosis. Clin Appl Thromb Hemost 27:10760296211050642. https://doi.org/10.1177/10760296211050642
Zhang L, Wang D, Wang Z, Li X, Xia W, Han Y, Su L, Fan X (2021) MiR-18a-5p acts as a novel serum biomarker for venous malformation and promotes angiogenesis by regulating the thrombospondin-1/P53 signaling axis. Am J Transl Res 13(10):11271–11286
Boechat JL, Chora I, Morais A, Delgado L (2021) The immune response to SARS-CoV-2 and COVID-19 immunopathology - current perspectives. Pulmonology 27(5):423–437
Brogaard L, Larsen LE, Heegaard PMH, Anthon C, Gorodkin J, Dürrwald R, Skovgaard K (2018) IFN-λ and microRNAs are important modulators of the pulmonary innate immune response against influenza A (H1N2) infection in pigs. PLoS One. 13(4):e0194765
Sivashanmugam K, Kandasamy M, Subbiah R, Ravikumar V (2021) Repurposing of histone deacetylase inhibitors: a promising strategy to combat pulmonary fibrosis promoted by TGF-β signalling in COVID-19 survivors. Life Sci. 1(266):118883
Gusev E, Sarapultsev A, Solomatina L, Chereshnev V (2022) SARS-CoV-2-Specific Immune Response and the Pathogenesis of COVID-19. Int J Mol Sci 23(3):1716
Yijin Z, Yujian Y, Zhao L, Hui D, Ping W (2022) miR-18a-5p reduces inflammatory response and human dermal microvascular endothelial cells permeability by targeting thrombospondin 1 in chronic idiopathic urticaria. J Biomater Tissue Eng 12:1252–1259. https://doi.org/10.1166/jbt.2022.3023
Geng H, Guan J (2017) MiR-18a-5p inhibits endothelial-mesenchymal transition and cardiac fibrosis through the Notch2 pathway. Biochem Biophys Res Commun. 491(2):329–336
Acknowledgements
All authors would like to acknowledge the patients for participation in this work.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Author information
Authors and Affiliations
Contributions
All authors were involved in conceiving the study and in protocol development. A.M. Eessa was guarantor and gained ethical approval and R.A. Haroun measured miR-18a expression level. W.H. Osman and R.A. Haroun analyzed the data. R.A. Haroun wrote the first draft of the manuscript. All authors revised and approved the final version.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The current study was approved by the ethics committee of the Faculty of Medicine, Port-Said University; Egypt (ERN MED (23/04/2020)S.no(5)MED). Informed consent was obtained from all patients.
Consent for publication
This statement is to certify that all authors have seen and approved the manuscript being submitted, have contributed significantly to the work, attest to the validity and legitimacy of the data and its interpretation, and agree to its submission.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Haroun, R.AH., Osman, W.H. & Eessa, A.M. Prognostic significance of serum miR-18a-5p in severe COVID-19 Egyptian patients. J Genet Eng Biotechnol 21, 114 (2023). https://doi.org/10.1186/s43141-023-00565-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43141-023-00565-y