
Abstract
Background
Glutathione s-transferases (GSTs) perform an essential role in detoxification of xenobiotics and endogenous compounds via their conjugation to reduce glutathione.
Results
A GST enzyme, designated tick larvae glutathione S transferase (TLGST), was purified from larvae of the camel tick Hyalomma dromedarii via ammonium sulfate precipitation, glutathione-Sepharose affinity column and Sephacryl S-300 chromatography. TLGST-specific activity was found to be 1.56 Umg−1 which represents 39 folds and 32.2% recovery. The molecular weight of TLGST purified from camel tick larvae was found as 42 kDa by gel filtration. TLGST has a pI value of 6.9 and was found a heterodimeric protein of 28 and 14 kDa subunits as detected on SDS-PAGE. The Lineweaver–Burk plot calculated the km for CDNB to be 0.43 mM with Vmax value of 9.2 Umg−1. TLGST exhibited its optimal activity at pH 7.9. Co2+, Ni2+ and Mn2+ increased the activity of TLGST while Ca2+, Cu2+, Fe2+ and Zn2+ inhibited it. TLGST was inhibited by cumene hydroperoxide, p-hydroxymercuribenzoate, lithocholic acid, hematin, triphenyltin chloride, p-chloromercuribenzoic acid (pCMB), N-p-Tosyl-L-phenylalanine chloromethyl ketone (TPCK), iodoacetamide, EDTA and quercetin. pCMB inhibited TLGST competitively with Ki value of 0.3 mM.
Conclusions
These findings will help to understand the various physiologic conditions of ticks and targeting TLGST could be significant tool for development of prospective vaccines against ticks as a bio-control strategy to overcome the rapid grows in pesticide-resistant tick populations.
Graphical Abstract

Similar content being viewed by others


Background
Camels produce meat, milk, wool, hair and hide and are utilized in transport and most of which are infected with the external parasites ticks [4, 39]. Ticks are blood-sucking parasitic arachnids infesting humans and all animals of economic importance leading to big losses in livestock production through transmission of various pathogenic organisms comprising rickettsiae, bacteria, protozoa, spirochetes and viruses. Ticks cling to animal hides and then introduce the toxins into them and transmit the diseases, which reduce their productivity [14, 32]. The ticks Hyalomma dromedarii are specific on camels and infecting them with the spring–summer encephalitis virus [3]. Studies about tick behaviour, ecology and physiology provide well understanding about these species, making them important tools for developing new biocontrol methods. Studying of tick physiology has acquired growing significance regarding the mechanisms implicated in toxins detoxification [15, 20, 27]. Ticks are exposed to various oxidative stress causing agents like exposure to acaricides and the ability for ingesting large quantities of blood from their hosts. Various potential toxic molecules are included in blood which have the ability to elevate the output of various free radicals such as reactive oxygen species (ROS) causing oxidative stress [16, 48]. Ticks must counteract the ROS produced by uptaking and digesting blood meals. Accordingly, ticks utilize various antioxidant molecules like superoxide dismutase (SOD) [25], catalase (CAT) [23, 28], glutathione peroxidase (GPx) [24] and glutathione S-transferases (GSTs) to overcome oxidative stress [21].
Glutathione S-transferase (GST) [GSTs, EC 2.5.1.18] is an enzyme that acts to excrete physiologic and xenobiotic molecules for cells protection from chemical toxins and stresses [9, 15]. GSTs are ubiquitous enzymes of multiple functions exist in microbial, plant and animal sources. The primary function of GSTs is detoxifying different toxins (insecticides, herbicides, drugs) via catalyzing the conjugation to glutathione (GSH). GSH is a fundamental tri-peptide sulfhydryl antioxidant found in plant and animal tissues in very low concentrations for performing the function of detoxification of xenobiotics and endogenous oxidants [17, 33, 34]. This takes place by the nucleophilic attack of the GSH sulfur atom on electrophilic substrates,decreasing their reactivity with cellular macromolecules and forming an important line of defence for cell components from reactive molecules [22]. GSTs are divided into soluble cytosolic (alpha, mu, Pi, omega, theta, delta, sigma and zeta), mitochondrial GST (alpha, mu, Pi and kappa), and membrane bound microsomal GST [7, 49]. A special property of all cytosolic GSTs is being homo- or heterodimers with two substrate binding sites: a highly specific and conserved G-site (GSH binding site) and H-site (hydrophobic binding site) for electrophilic substrates [30]. The inhibition mechanisms of GST by different inhibitor compounds like quercetin, hematin, ethacrynic acid and triphenyltin chloride have been broadly studied as a diagnostic way for resistance mechanisms [2, 11, 40, 46]. GSTs of various tick species were identified and characterized; Rhipicephalus microplus [38], R. appendiculatus [8], Dermacentor variabilis [10], R. Sanguineus [12] and H. longicornis [21]. The aim of this study is purification and characterization of GST enzyme from H. Dromedarii larvae and inspection of the effect of some chemical inhibitors on the tick GST activity which will be helpful for development of prospective vaccines against ticks.
Methods
Tick material
The engorging females of camel tick H. dromedarii were gathered from a market for camels near Cairo and incubated at 28 ºC and 85% relative humidity. Eggs were daily collected from oviposited females, immediately frozen (− 40 °C) or either held at same conditions till the required age then frozen and stored at 3 days interval (0, 3, 6, 9 etc.). The larvae hatched after 27 days.
Chemicals
1-chloro-2, 4-dinitrobenzene (CDNB), phenylmethylsulfonylfluoride (PMSF), reduced glutathione (GSH), glutathione sepharose 4 fast flow, sephacryl S-300, cumene hydroperoxide, p-hydroxymercuribenzoate, lithocholic acid, hematin, p-chloromercuribenzoic acid (pCMB), N-p-tosyl-l-phenylalanine chloromethyl ketone (TPCK), iodoacetamide, gel filtration molecular weight markers, and triphenyltin chloride were purchased from Sigma Chemical Co. All other chemicals were of analytical grade.
Assay of GST activity
The GST assay reaction mixture contained in 1 ml total volume of 0.1 M K-phosphate buffer, pH 6.5, 1 mM 1-chloro-2,4-dinitrobenzene (CDNB), 1 mM GSH, and enzyme solution. The increase in absorbance was monitored at 340 nm for 3 min at 25 °C. The product extinction coefficient was considered as 9.6 mM−1 cm−1. One GST unit is equivalent to the enzyme amount which conjugates 1 μmole CDNB in 1 min [19].
Protein determination
Protein concentrations were estimated via the dye binding procedure [6] using albumin from bovine serum (BSA) as a standard.
Purification of camel tick larvae glutathione S-transferase
Preparation of crude extract
All experimental procedures were carried out at 4 °C. Homogenization of camel tick larvae was taken place in 0.02 M Na-phosphate buffer pH 7.0, using a Teflon pestled homogenizer. Cell residues and indissoluble substances were brought out via centrifugation (12,000 × g, 30 min) and supernatant was kept as crude extract.
Ammonium sulfate treatment
Crude extract was gradually saturated with 40% (NH4)2SO4 with continuous stirring (30 min, 4 °C) followed by centrifugation (8000 × g, 20 min). The precipitate was ignored and the supernatant was raised to 80% (NH4)2SO4 saturation, obtaining the pellet via centrifugation (12,000 × g, 30 min) and dissolving in 0.02 M Na-phosphate buffer pH 7.0 followed by inclusive dialysis in this buffer.
Glutathione sepharose affinity chromatography
The dialyzed 40 − 80% ammonium sulfate sample was applied onto a GSH-sepharose 4 fast flow column (8 × 1.4 cm) pre-equilibrated with 0.02 M Na-phosphate buffer pH 7.0 and set at a flow rate of 30 ml/h. Washing the column with 100 m1 equilibration buffer and elution of the adsorbed proteins was achieved with 10 mM GSH contained in 0.05 M Tris–HCl buffer pH 8.0. Collection of 3 ml fractions and those with GST activity were combined and concentrated via lyophilization.
Sephacryl S-300 size exclusion chromatography
Fractioning the GST containing concentrate on a Sephacryl S-300 column (142 cm × 1.75 cm) pre-equilibrated and run with 0.02 M Na-phosphate buffer pH 7.0 at 30 ml/h flow rate with collection of 2 ml fractions. For estimation of the native GST molecular weight, Sephacryl S-300 column was calibrated with 440 kDa ferritin, 240 kDa catalase, 150 kDa alcohol dehydrogenase, 67 kDa bovine serum albumin, and 17 kDa myoglobin as standard markers.
Electrophoretic analysis
Twelve percent SDS PAGE was done for estimation of the purified GST subunits [29, 42]. Isoelectrofocusing PAGE was done for estimation of the pI values [35, 41]. Staining of proteins was achieved with 0.25% Coomassie Brilliant Blue R-250.
Optimum pH determination
For estimation of the action of pH on GST activity, the enzyme was assayed utilizing two buffers; 0.1 M potassium phosphate (5.7 − 8.0) and Tris–HCl (8.0 − 9.2) at 25 °C.
Enzyme kinetics
Michalis-Menten constant (Km) was deduced from the Linewear-Burk plot construction utilizing (0.1 − 2 mM) CDNB as substrate at 25 °C.
Effect of divalent cations and inhibitors on GST activity
Effect of various cations and inhibitors on GST activity was achieved via preincubation of the purified enzyme with each molecule at 25 °C prior to assaying activity.
Results
Purification of tick larvae GST enzyme
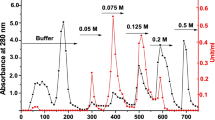
Typical GST enzyme purification scheme from larvae of H. dromedarii tick (Table 1) showed the primary GST-specific activity in larvae crude extract as 0.04 Umg−1. Forty to 80% ammonium sulfate precipitated most of GST enzyme and achieved 82.9% recovery. The glutathione sepharose affinity chromatography of 40–80% ammonium sulfate fraction (Fig. 1a) appeared one GST activity peak eluted with 10 mM GSH and designated TLGST. The TLGST concentrate fractionation on Sephacryl S-300 column (Fig. 1b) gives one GST peak with an increase in TLGST-specific activity to 1.56 Umg−1, 39 folds and 32.5% yield (Table 1). Apparent molecular weight of H. dromedarii TLGST was derived from its elution volume from the Sephacryl column as 42 kDa (Fig. 1b).

a The chromatographic pattern of 40–80% ammonium sulfate fraction of tick larvae crude extract on GSH-Sepharose column (8 × 1.4 cm) previously equilibrated and washed with 0.02 M Na-phosphate buffer pH 7.0. b The chromatography of tick larvae TLGST on Sephacryl S-300 column (142 cm × 2.4 cm) formerly equilibrated with 0.02 M Na-phosphate buffer pH 7.0
Electrophoretic analysis of TLGST
SDS-PAGE of denatured H. dromedarii TLGST enzyme (Fig. 2a) displayed residues of the native intact protein (42 kDa) and two protein bands of 28 kDa and 14 kDa representing the TLGST subunits. The TLGST pI value was seen as single species on isoelectrofocusing PAGE at pH 6.9 (Fig. 2b).

a Analysis of TLGST on 12% SDS PAGE; (1) molecular weight marker proteins, (2) TLGST. b Analysis of TLGST on isoelectric focusing PAGE: (1) pI marker proteins, (2) TLGST
Determination of optimum pH and Km value of TLGST
The impact of pH on TLGST activity was tested using K-phosphate and Tris–HCl buffers which revealed the highest activity of the enzyme at pH 7.9 (Fig. 3a). Construction of Lineweaver–Burk plot of reaction speed and substrate concentration reciprocals showed the Km value of TLGST as 0.43 mM CDNB with a corresponding Vmax of 9.2 Umg−1 (Fig. 3b).

a The pattern of TLGST pH profile utilizing 0.1 M potassium phosphate (5.7–8.0) and Tris–HCl (8.0–9.2). b Lineweaver–Burk plot of TLGST reaction speed in response to CDNB concentrations
Effect of divalent cations and inhibitors on TLGST
The data present in Table 2 showed TLGST activity with 2 mM and 5 mM of various cations. CoCl2, NiCl2 and MnCl2 led to increasing of TLGST activity while CaCl2, CuCl2, ZnCl2 and FeCl2 caused an activity decrease. The influence of various inhibitors on the purified camel tick larvae TLGST is presented in Table 3. N-p-Tosyl-L-phenylalanine, Quercetin and p-chloromercuribenzoic acid (pCMB) were found the most potent inhibitors of TLGST.
Inhibition kinetic of TLGST by pCMB
The effect of p-Chloromercuribenzoic acid (pCMB) concentrations on the purified camel tick larvae TLGST indicted that I50 = 0.7 mM pCMB with a maximum inhibition of 97.2% was carried out by 1 mM pCMB (Fig. 4a). A linear relation was monitored via construction of the Hill plot for TLGST inhibition with pCMB giving a slope of 0.88 (Fig. 4b). The type of inhibition of TLGST by pCMB was constructed (Fig. 4c) and Ki value of TLGST inhibition is determined as 0.3 mM (Fig. 4d).

a TLGST Inhibition with pCMB various concentrations. b Hill plot of TLGST inhibition with pCMB. c Lineweaver–Burk plots showing TLGST inhibition type with pCMB. d TLGST inhibition constant (Ki) value for pCMB
Discussion
Glutathione S-transferase is the major detoxifying enzyme that implicated in toxins and insecticide resistance in various insects [13]. Our purpose here was to purify and characterize GST enzyme from larvae of the camel tick H. dromedarii. We purified the GST from H. dromedarii tick larvae via ammonium sulfate precipitation, GSH-sepharose affinity chromatography and size exclusion chromatography. Similar purification procedures of GSTs were reported, GST from German cockroach [46], from non-biting midges Chironomidae larvae [47] from filarial worms [2], from Liposcelis insects [43] and from blueberry fruits [5]. In this study, the affinity chromatographic pattern of tick larvae GST on GSH-Sepharose column showed one GST activity peak eluted with 10 mM GSH and named TLGST (Fig. 1a). A successive chromatographic step of TLGST via Sephacryl S-300 column (Fig. 1b) gave one GST activity peak and raised TLGST-specific activity to 1.56 Umg−1 which correspond 39 folds and 32.5% yield (Table 1). A broad purification folds and recovery percentages for GST from various sources were stated from German cockroach (52.8 folds, 33% yield) [46], from Atactodea striata snail (43.2 folds) [45], from Apis mellifera macedonica bee (51.9 folds, 54.5% yield) [37], from filarial worms (43.2 folds, 11.1% yield) [2], from Liposcelis insects (32.2, 99.8 and 42.5 folds and 41.7%, 120.8% and 79.2% yields) [43] and from blueberry fruits (189 fold, 23.3% yield) [5].
The elution volume of TLGST from the size exclusion column calculated its native molecular mass as 42 kDa (Fig. 1b). This was assured via SDS-PAGE of denatured purified TLGST (Fig. 2a) which showed ruminants of the native intact protein (42 kDa) and two subunits of 28 kDa and 14 kDa protein bands denoting that TLGST is heterodimeric protein. Various structures were stated for GSTs; monomeric GSTs from the spider mite with Mwt of 22.1 kDa [26], the Aedes aegypti mosquito with Mwt of 25 kDa [31] and rice moth with Mwt of 23 kDa [18]. Homodimeric GSTs as that of German cockroach with 25.5 kDa subunit [46], Atactodea striata snail with 24 kDa subunit [45], grasshopper insect with 19 kDa subunit [1], filarial worms with 24.6 kDa subunit [2] and teleost Monopterus albus fish with 26 kDa subunit [22]. Heterodimeric GSTs as that of Apis mellifera macedonica bee with 29 and 25 kDa subunits [37] and Rhipicephalus microplus tick with 30.7 and 28.4 kDa [38]. In this present study, TLGST purity was inspected through analysis on isoelectrofocusing PAGE which appeared as one species (Fig. 2b) with pI value of 6.9 denoting an alkaline isoenzyme. Various isoelectric points (pI) values were reported for GSTs; non-biting midges Chironomidae larvae and Atactodea striata snail GSTs pI value of 5.5 [45, 47], Rhipicephalus microplus tick GST pI value was 8.6 [38], R. Appendiculatus tick GST pI values were 7.67 and 8.51 [8], Apis mellifera macedonica bee GSTs pI values were 7.4 and 4.58 [37], silkmoth insect GST pI value was 6.01 [44] and H. longicornis tick GST pI value was 7.67 [21].
The camel tick larvae TLGST displayed its optimum pH at 7.9 (Fig. 3a) confirming its alkaline property. The optimum pH of the H. longicornis tick GST was around 7.5–8.0 [21], at pH 8.0 in non-biting midges Chironomidae larvae and Atactodea striata snail GSTs [45, 47], around 6.5–7.5 in filarial worms GST [2] and 7.2 for blueberry fruits GST [5]. TLGST showed Km value of 0.43 mM CDNB and has Vmax of 9.2 units/mg protein (Fig. 3b) indicating its high ability to conjugate with CDNB. The GSTs Km values for CDNB were 0.82 and 0.64 mM in H. longicornis tick HlGST and HlGST2 [21], 0.16 mM in German cockroach [46], 0.62 mM in Apis mellifera macedonica bee larvae [37], 2.5 mM in filarial worms [2] and 5.68 mM in blueberry fruits CDNB [5]. In the current study, Co2+, Ni2+ and Mn2+ cause increases in TLGST activity while Ca2+, Cu2+, Fe2+ and Zn2+ decreased it (Table 2). The inhibition of TLGST by Cu2+ and Zn2+ ions was consistent with that of Van fish GST [36]. The effect of various inhibitors on the purified TLGST (Table 3) showed slight inhibition of TLGST with DTT, β-Mercaptoethanol, Cumene hydroperoxide and p-Hydroxymercuribenzoate (pHMB). It was moderately inhibited with Lithocholic acid, Hematin and Triphenyltin chloride. TLGST was potently inhibited with the cysteinyl protease inhibitors p-chloromercuribenzoic acid (pCMB) and N-p-Tosyl-L-phenylalanine chloromethyl ketone (TPCK) which revealed its dependence on thiol reactivity and suggested it to have a cysteine protease activity. TLGST was inhibited with iodoacetamide indicated that residues of cysteine, histidine and methionine have significant impacts on structure and effectiveness of TLGST. Inhibition of TLGST with the metal chelating agent EDTA suggested it as a metalloenzyme. Triphenyltin chloride inhibited GSTs of German cockroach, Apis mellifera macedonica bee and equine [37, 40, 46], hematin inhibited that of filarial worm [2] while quercetin inhibited that of cockroach [46].
Here, pCMB was found the most potent inhibitor of TLGST since 1 mM inhibited 97.2% of its activity with an I50 = 0.7 mM (Fig. 4a). A straight line was achieved on plotting the Hill plot of TLGST inhibition with pCMB with a slope of 0.88 which refers to one binding site for pCMB on TLGST (Fig. 4b). A competitive inhibition of TLGST with pCMB was achieved due to absence of change in Vmax value while Km value was increased (Fig. 4c). The Ki value for inhibition of TLGST with pCMB is 0.3 mM (Fig. 4d) which confirms the potency of pCMB as TLGST inhibitor.
Conclusion
In conclusion, this is the first report on purification and characterization of GST from larvae of the camel tick H. dromedarii. This TLGST might have an essential role in avoiding oxidative stress produced either by ingestion of large blood meals or via the broad utilization of pesticides. Targeting TLGST could be significant tool for development of prospective vaccines against ticks as a bio-control strategy to overcome the rapid grows in pesticide-resistant tick populations.
Availability of data and materials
All data and materials are available.
Abbreviations
- GST:
-
Glutathione S transferase
- TLGST :
-
Tick larvae glutathione S transferase
- pCMB:
-
p-Chloromercuribenzoic acid
- BSA:
-
Bovine serum albumin
- PAGE:
-
Polyacrylamide gel electrophoresis
- ROS:
-
Reactive oxygen species
- CDNB:
-
1-Chloro-2,4-dinitrobenzene
References
Adewale IO, Afolayan A (2006) Studies on glutathione transferase from grasshopper (Zonocerus variegatus). Pestic Biochem Phys 85:52–59
Ahmad R, Srivastava AK, Walter RD (2008) Purification and biochemical characterization of cytosolic glutathione-S-transferase from filarial worms Setaria cervi. Comp Biochem Physiol B 151:237–245
Alanazi AD, Nguyen VL, Alyousif MS, Manoj RRS, Alouffi AS, Donato R, Sazmand A, Mendoza-Roldan JA, Dantas-Torres F, Otranto D (2020) Ticks and associated pathogens in camels (Camelus dromedarius) from Riyadh Province. Saudi Arabia Parasit Vector 13(1):110
Azawi SM, Carter SD, Woldehiwet Z, Wernery U (1996) Serology of orthopoxvirus cameli infection in dromedary camels. Analysis be ELISA and western blotting. Comp Immun Microbiol Infect Dis 19:65–78
Balc N, Turkan F, Sakiroglu H, Aygun A, Sen F (2019) Purification and characterization of glutathione S-transferase from blueberry fruits (Vaccinium arctostaphylos L.) and investigated of some pesticide inhibition effects on enzyme activity. Heliyon 5:e01422
Bradford MM (1976) A rapid and sensitive method for quantization of microgram quantities of protein utilizing the principle of protein-dye-binding. Anal Biochem 72:248–254
Chatzikonstantinou M, Vlachakis D, Chronopoulou E, Papageorgiou L, Papageorgiou AC, Labrou NE (2017) The glutathione transferase family of Chlamydomonas reinhardtii: identification and characterization of novel sigma class-like enzymes. Algal Res 24:237–250
da Silva Vaz Jnr I, Imamura S, Ohashi K, Onuma M (2004) Cloning, expression and partial characterization of a Haemaphysalis longicornis and a Rhipicephalus appendiculatus glutathione S-transferase. Insect Mol Biol 13:329–335
Dasari S, Gonuguntla S, Yellanurkonda P, Nagarajan P, Meriga B (2019) Sensitivity of glutathione S-transferases to high doses of acrylamide in albino wistar rats: affinity purification, biochemical characterization and expression analysis. Ecotoxicol Environ Saf 182:109416
Dreher-Lesnick SM, Mulenga A, Simser JA, Azad AF (2006) Differential expression of two glutathione S-transferases identified from the American dog tick, Dermacentor variabilis. Insect Mol Biol 15:445–453
Dücker R, Parcharidou E, Beffa R (2020) Flufenacet activity is affected by GST inhibitors in blackgrass (Alopecurus myosuroides) populations with reduced flufenacet sensitivity and higher expression levels of GSTs. Weed Sci 68:451–459
Duscher GG, Galindo RC, Tichy A, Hummel K, Kocan KM, dela Fuente J, (2014) Glutathione S-transferase affects permethrin detoxification in the brown dog tick, Rhipicephalus sanguineus. Ticks Tick Borne Dis 5:225–233
Erban T, Stara J (2014) Methodology for glutathione S-transferase purification and localization in two-dimensional gel electrophoresis performed on the pollen beetle, Meligethes aeneus (Coleoptera: Nitidulidae). J Asia Pac Entomol 17:369–373
Fernandez-Ruiz N, Estrada-Pena A (2022) Scenes from tick physiology: proteins of sialome talk about their biological processes. Front Cell Infect Microbiol 11:767845
Freitas DRJ, Rosa RM, Moraes J, Campos E, Logullo C, Da Silva Vaz I Jr, Masuda A (2007) Relationship between glutathione S-transferase, catalase, oxygen consumption, lipid peroxidation and oxidative stress in eggs and larvae of Boophilus microplus (Acarina: Ixodidae). Comp Biochem Physiol A 146:688–694
Galay RL, Umemiya-Shirafuji R, Mochizuki M, Fujisaki K, Tanaka T (2015) Iron metabolism in hard ticks (Acari: Ixodidae): the antidote to their toxic diet. Parasitol Int 64:182–189
Ghani M, Zayeri Z, Maleki B (2021) Glutathione stabilized Fe3O4 nanoparticles as the sorbent for magnetic solid-phase extraction of diazepam and sertraline from urine samples through quantitation via high-performance liquid chromatography. J Sep Sci 44:1195–1202
Gullipalli D, Arif A, Aparoy P, Svenson GJ, Whiting MF, Reddanna P, Dutta-Gupta A (2010) Identification of a developmentally and hormonally regulated Delta-Class glutathione S-transferase in rice moth Corcyra cephalonica. Comp Biochem Physiol B 156:33–39
Habig WH, Pabst MJ, Jakoby WB (1974) Glutathione S-transferases: the first enzymatic step in mercapturic acid formation. J Biol Chem 249:7130–7139
Hemingway J (2000) The molecular basis of two contrasting metabolic mechanisms of insecticide resistance. Insect Biochem Mol Biol 30:1009–1015
Hernandez EP, Kusakisako K, Talactac MR, Galay RL, Hatta T, Matsuo T, Fujisaki K, Tsuji N, Tanaka T (2018) Characterization and expression analysis of a newly identified glutathione S-transferase of the hard tick Haemaphysalis longicornis during blood-feeding. Parasite Vector 11:91
Huang Q, Liang L, Wei T, Zhang D, Zeng QY (2008) Purification and partial characterization of glutathione transferase from the teleost Monopterus albus. Comp Biochem Physiol C 147:96–100
Ibrahim MA, Ghazy AM, Masoud HMM (2015) Catalase from larvae of the camel tick Hyalomma dromedarii. Biochem Biophys Reports 4:411–416
Ibrahim MA, Mohamed MM, Ghazy AM, El-Mogy M, Masoud HMM (2016) Purification and characterization of two glutathione peroxidases from embryo of the camel tick Hyalomma dromedarii. Russ J Bioorg Chem 42(3):272–281
Ibrahim MA, Mohamed MM, Ghazy AM, Masoud HMM (2013) Superoxide dismutases from larvae of the camel tick Hyalomma dromedarii. Comp Biochem Physiol B 164:221–228
Konanz S, Nauen R (2004) Purification and partial characterization of a glutathione S-transferase from the two-spotted spider mite, Tetranychus urticae. Pestic Biochem Phys 79:49–57
Kostaropoulos I, Papadopoulos AI, Metaxakis A, Boukouvala E, Papadopoulou-Mourkidou E (2001) Glutathione S-transferase in the defence against pyrethroids in insects. Insect Biochem Mol Biol 31:313–319
Kumar D, Budachetri K, Meyers VC, Karim S (2016) Assessment of tick antioxidant responses to exogenous oxidative stressors and insight into the role of catalase in the reproductive fitness of the Gulf Coast tick, Amblyomma maculatum. Insect Mol Biol 25(3):283–294
Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of Bacteriophage T4. Nature 227:680–685
Lallement PA, Brouwer B, Keech O, Hecker A, Rouhier N (2014) The still mysterious roles of cysteine-containing glutathione transferases in plants. Front Pharmacol 5:192
Lumjuana N, Stevensona BJ, Prapanthadara L, Somboon P, Brophy PM, Loftus BJ, Severson DW, Ranson H (2007) The Aedes aegypti glutathione transferase family. Ins Biochem Molecul Biol 37:1026–1035
Madison-Antenucci S, Kramer LD, Gebhardt LL, Kauffman E (2020) Emerging tick-borne diseases. Clin Microbiol Rev 33(2):e00083-e18
Maleki B, Atharifar H, Reiser O, Sabbaghzadeh R (2019) Glutathione-coated magnetic nanoparticles for one-pot synthesis of 1,4-dihydropyridine derivatives. Polycycl Aromat Compd 41(4):1–14
Maleki B, Esmaeilnezhad E, Choi HJ, Koushki E, Aliabad HAR, Esmaeili M (2020) Glutathione-capped core-shell structured magnetite nanoparticles: fabrication and their nonlinear optical characteristics. Curr Appl Phys 20:822–827
O’Farrell PH (1975) High resolution two-dimentional electrophoresis of proteins. J Bio Chem 250(10):4007–4021
Ozaslan MS, Ciftci M (2022) Impacts of some metal ions on glutathione s-transferase in the liver of Chalcalburnus tarichi: an endemic species of Lake Van. Turk J Chem 46:1324–1331
Papadopoulos AI, Polemitou I, Laifi P, Yiangou A, Tananaki C (2004) Glutathione S-transferase in the insect Apis mellifera macedonica Kinetic characteristics and effect of stress on the expression of GST isoenzymes in the adult worker bee. Comp Biochem Physiol C 139:93–97
Rosa de Lima MF, Sanchez Ferreira CA, Joaquim de Freitas DR, Valenzuela JG, Masuda A (2002) Cloning and partial characterization of a Boophilus microplus (Acari: Ixodidae) glutathione S-transferase. Insect Biochem Mol Biol 32:747–754
Sazmand A, Joachim A, Otranto D (2019) Zoonotic parasites of dromedary camels: so important, so ignored. Parasite Vector 12:610
Skerlov J, Ismail A, Lindstrom H, Sjodin B, Mannervik B, Stenmark P (2021) Structural and functional analysis of the inhibition of equine glutathione transferase A3–3 by organotin endocrine disrupting pollutants. Environ Pollut 268:115960
Ubuka T, Masuoka N, Yoshida S, Ishino K (1987) Determination of isoelectric point value of 3-mercaptopyruvate sulfurtransferase by isoelectric focusing using ribonuclease A-glutathione mixed disulfides as standards. Anal Biochem 167:284–289
Weber K, Osborn M (1969) The reliability of molecular weight determinations by dodecyl sulfate polyacrylamide gel electrophoresis. J Biol Chem 244:4406–4412
Wu S, Dou W, Wu J, Wang J (2009) Purification and partial characterization of glutathione S-transferase from insecticide-resistant field populations of Liposcelis paeta Pearman (Psocoptera: Liposcelididae). Arch Insect Biochem Physiol 70(2):136–150
Yamamoto K, Nagaoka S, Banno Y, Aso Y (2009) Biochemical properties of an omega-class glutathione S-transferase of the silkmoth, Bombyx mori. Comp Biochem Physiol C 149:461–467
Yang H, Zeng Q, Nie L, Zhu S, Zhou X (2003) Purification and characterization of a novel glutathione S-transferase from Atactodea striata. Biochem Biophys Res Com 307:626–631
Yu SJ, Huang SW (2000) Purification and characterization of glutathione S-transferases from the German cockroach, Blattella germanica (L.). Pestic Biochem Phys 67:36–45
Yuen WK, Ho JW (2001) Purification and characterization of multiple glutathione S-transferase isozymes from Chironomidae larvae. Comp Biochem Physiol A 129:631–640
Zheng H, Li AY, Teel PD, Pérez de León AA, Seshu J, Liu J (2015) Biological and physiological characterization of in vitro blood-feeding in nymph and adult stages of Ornithodoros turicata (Acari: Argasidae). J Insect Physiol 75:73–79
Zhuge X-L, Xu H, Xiu Z-J, Yang H-L (2020) Biochemical functions of glutathione S-transferase family of salix babylonica. Front Plant Sci 11:364
Acknowledgements
This study was supported and funded by National Research Centre, Egypt.
Funding
This study was funded by National Research Centre, Egypt.
Author information
Authors and Affiliations
Contributions
All authors contributed to the experimental design, hands on work, discussions, and commented on the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Masoud, H.M.M., Helmy, M.S., Darwish, D.A. et al. Purification, characterization, and enzyme kinetics of a glutathione S transferase from larvae of the camel tick Hyalomma dromedarii. J Genet Eng Biotechnol 21, 28 (2023). https://doi.org/10.1186/s43141-023-00486-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43141-023-00486-w