
Abstract
Background
Ziziphus species particularly Ziziphus mauritiana and Ziziphus nummularia constitute an important part of genetic resources in India. They contribute economically as a fruit crop with lots of morphological and pomological variability. In current study, 48 accessions belonging to two wild Ziziphus species, i.e., Z. mauritiana and Z. nummularia, were characterized using SSR markers. In addition, external features were also examined using stereomicroscope.
Results
Present investigation was done to explore the genetic structure of North Indian jujube. In total, 23 SSR markers detected 57 SSR alleles with an average of 2.47 alleles. Highest number of alleles (4) were detected by three primers, namely BFU1178, BFU479, and ZCMS14, while lowest number of alleles (2) were detected by fifteen primers. Highest Polymorphism Information Content (PIC) was 0.500 and shown by two primers, namely BFU528 and BFU1248, while lowest PIC (0.041) was observed in primers BFU286 with mean value of 0.443. Similarly, highest value of marker index (MI) was detected by primer BFU1178 i.e. 1.969, and lowest value of marker index was observed in primer BFU286 i.e. 0.021. Dendrogram generated using SSR markers data and principal component analysis showed two major groups of the analyzed germplasm with intermixing. STRUCTURE analysis also clustered all the accessions into two groups. We did not found correlation between geographic and genetic distances.
Conclusions
The preliminary results suggest that there is high level of gene pool mixing in these species which can be attributed to their cross-pollination habit. However, more such studies with large numbers of samples are required in future to gain concrete insights of the genetic structure in these species.
Similar content being viewed by others


Background
Ziziphus mauritiana and Ziziphus nummularia both jujube species are commonly known as ber (Indian jujube) in India. These are subtropical to tropical plant species, and fruits of these species are liked and eaten by large population of India. Z. mauritiana is a cultivated species, while Z. nummularia is a wild species, and both of these vary in their habits. Generally, both the species have prickly stems and branches with ovate leaves and minute flowers. The pollination is strictly cross-pollination as it also shows the protandry habit. However, plants of Z. mauritiana are larger in height having larger leaves, stems, branches and fruits, while plants of Z. nummularia show shrub-type habit with smaller leaves, profuse branching and small fruits [46, 58, 64]. Some morphological features of both of these species are shown in Fig. 1. Both of these species play important role in economy as well as in ecology. Fruits of these species contain many important constituents such as vitamins, alkaloid, and other secondary metabolites which exhibit many health benefits [34, 35, 37, 40]. However, with the introduction of new hybrid cultivars in the market, the wild genetic resources are being neglected which is the matter of concern. We do not know the type and diversity of wild genetic resources we are having at present. However, this type of information is highly required for conservation of important genetic resources. The effective conservation, management, and efficient utilization of plant genetic resources can be done if we have explored the basic knowledge about essential biological phenomena in plants and characterize them timely. An adequate knowledge regarding how to best utilize the existing genetic diversity in plant population is of fundamental interest for the efficient management of plant resources [28, 62]. Characterization of genetic resources includes many ways such as morphological traits, chemical compounds identification, genetic traits and cytological studies. To include all these techniques or methods in a single study is somewhat tedious and cumbersome and needs expertise at all these levels. The most common characterization method is morphological characterization; however, it suffers from few limitations such as varying of phenotypic traits with varying environments [50, 61] and sometimes results in wrong interpretations and conclusions. Therefore, a good alternative to this is the characterization at genetic level using DNA markers or genetic markers [39, 54, 66, 67]. These marker techniques require expertise and few advanced instrumentation, but results are reliable and free from any limitations. Genetic markers are determined by allelic forms of genes or genetic loci or polymorphic fragments of DNA and can be stably transmitted from one generation to another. Therefore, these markers can be used as experimental tags to keep track of an individual, a tissue, a cell, a nucleus, a chromosome, or a gene. Genetic markers are broadly categorized into two main categories, i.e., classical markers and DNA markers [69]. Classical markers include morphological markers, cytological markers, and biochemical markers. DNA markers are the fragments of DNA revealing polymorphism between different genotypes or individuals or alleles of a gene. The polymorphism shown by marker fragments may arise due to alteration of nucleotide or mutation in the genomic loci [20]. These fragments are associated with a defined location within the genome and may be detected by means of different molecular marker techniques such as restriction fragment length polymorphism (RFLP), randomly amplified polymorphic DNA (RAPD), amplified fragment length polymorphism (AFLP) and single-nucleotide polymorphism (SNP) [12]. Molecular makers have been established as powerful tools in the analysis and assessment of genetic variation as well as in establishing genetic relationships within and among species [9, 29, 43, 44, 50, 53, 55]. There are great advantages of molecular markers as compared to traditional morphological markers. Molecular markers exhibit high polymorphism, reproducibility, even distribution across the whole genome, and selectively neutral behavior to environmental conditions. Therefore, it is used in many different areas such as genetic mapping, diversity analysis, parentage analysis, pedigree analysis, gene identifications, fidelity checking of tissue culture raised plants, and many more areas in breeding of crops and population genetic studies [4, 7, 8, 21, 25, 27, 33, 48, 49, 51]. However, among different marker systems, simple sequence repeat (SSR) markers have become the markers of choice due to their easy availability, codominant nature, and easy detection and cross-transferring nature across species and genera [3, 24, 52]. In Indian jujube species, few studies related to morpholoical and molecular markers have been reported [2, 6, 10, 13, 19, 22, 30, 31, 36, 56,57,58,59,60, 70]. However, most studies were conducted using less number of samples and dominant markers such as RAPD and AFLP. Moreover, Indian jujube germplasm has been explored less and requires more molecular works. Therefore, in present study, we have utilized the SSR markers in Ziziphus species with specific objectives to characterize the wild and cultivated genetic resources of Ziziphus in north western Indian states and to establish genetic relationships among the analyzed accessions of both the species, i.e., Z. mauritiana and Z. nummularia.

Morphological features of Z. mauritiana and Z. nummularia. a) Mature plant of Z. mauritiana. b) Flower bud of Z. mauritiana photographed with stereomicroscope. c–g) Stereomicroscopic photographs of flower showing anthers and stigma. h) A flowering branch. i) Abaxial leaf surface under stereomicroscope. j) Adaxial leaf surface under stereomicroscope. k) Stem surface under stereomicroscope. l) Prickle under stereomicroscope. m) Z. nummularia plant-bearing fruits. n–o) Unripend and ripened fruits of Z. nummularia
Materials and methods
Plant materials and DNA extraction
In the present study, 48 Ziziphus accessions i.e. 20 accessions of Z.mauritiana and 28 accessions of Z. nummularia, were collected from different geographical locations of northwest Indian states (Punjab, Rajasthan, Haryana, and Himachal Pradesh) and were analyzed using SSR markers. Of these, thirteen samples were from Punjab, twelve from Rajasthan, twelve from Haryana, and eleven from Himachal Pradesh. A detailed description with locations of all the accessions and their altitude range is given in Table 1. Young and fresh leaf samples belonging to these plants were collected. Leaves were properly observed, and effort was made to check that samples were free of disease or any damage. Samples were put in an airtight (sterilized) plastic bag containing silica gel to prevent the moisture and subsequent degradation. DNA was extracted using CTAB method [14] and liquid nitrogen.
Simple sequence repeat reactions
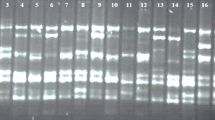
Thirty-one SSR primers which were developed by Wang et al. [65] were analyzed for polymorphism on a forty-eight selected DNA samples from Ziziphus species from various locations of northwest India (Table 1). Out of these, twenty-three primers were concluded as good reliable with unambiguous amplification and were further used for genotyping. SSR amplifications were carried in a 10 μl volume which was constituted using 4.8 μl of sterilized distilled water, 2.0 μl genomic DNA (13 ng/μl), 0.5 μl of forward and 0.5 μl of reverse primer (5 μM), 0.5 μl MgCl2 (25 mM), 1.0 μl 10 × PCR buffer (10 mM Tris-Hcl, 50 mM Kcl, pH 8.3), 0.5 μl dNTP mix (0.2 mM each of dATP, dGTP, dCTP, and dTTP), and 0.2 μl Taq polymerase (5 U/μl). The PCR conditions were as follows: 1 cycle of 5 min at 94 °C, 35 cycles of 1 min at 94 °C, 1 min at respective annealing temperature for each primer as shown in Table 2, 1 min at 72 °C, and final extension for 7 min at 72 °C. Amplification products were separated on 3% agarose gel in 1 × TBE buffer, and size of each fragment was estimated 50 bp DNA ladder (MBI Fermentas, Lithuania). Fragments were visualized by using ethidium bromide, and permanent photographs of gels were taken in gel documentation system (Bio-Rad laboratories-segrate, Milan, Italy).
Data analysis
Only unambiguously amplified alleles were scored manually and converted into binary data, i.e., 1 for the presence of band and 0 for the absence of band. Polymorphism Information Content (PIC) values were calculated using the formula given by Botstein et al. [5, 26]. Distance-based cluster analysis was performed by generating dendrogram based on Jaccard similarity coefficient and UPGMA method using DARwin [41]. The population genetic structure was elucidated using Bayesian model-based clustering method implemented in the software STRUCTURE, version: 2.3.3 [17, 42]. Ancestry model with admixture and correlated allele frequency model was set to get the estimates of posterior probability of data. Ten independent runs were given setting the value of K from 1 to 10 with 3 iterations for each value of K. Length of burn-in period was set at 100,000, and number of Markov chain Monte Carlo (MCMC) repeats after burn-in was set at 100,000. Evanno’s method [16]-based program STRUCTURE HARVESTER developed by Earl and Vonholdt [15] was utilized to find the value of estimated Ln probability of data LnP(K) and to get the best fit value of K for the data. STRUCTURE was run for all the analyzed accessions of the two species. Analysis of molecular variance (AMOVA) and Mantel test were performed using GenAlEx 6.5 version.
Results
SSR polymorphism and population structure
In the present study, twenty-three SSR primers were utilized which amplified unambiguously and produced reliable alleles. In total, 23 SSR primers amplified 57 alleles with an average of 2.47 alleles per primer. Size range of alleles varied from 80 to 500 bp. Minimum 2 alleles were amplified by fifteen primer pairs, while highest numbers of alleles were 4 and amplified by three primers as shown in Table 2. Although alleles amplified on average were not high, but considerable polymorphism was detected with these primers. Highest Polymorphism Information Content (PIC) was 0.500 and shown by two primers, namely BFU528 and BFU1248, while lowest PIC (0.021) was observed in primer BFU286 with mean value of 0.443. Similarly, highest value of marker index was detected by primer BFU1178 i.e. 1.969, and lowest value of marker index was observed in primer BFU286 i.e. 0.021. These values will be helpful in identifying suitable primers for future use in different genetic studies. STRUCTURE analysis of these two species showed two populations and the log likelihood reached a clear maximum value at K = 2 (Fig. 5a). Population structure showed that two different gene pools were contributing in the genetic makeup of analyzed accessions (Fig. 5b).
When cluster analysis of the studied species was done using dendrogram and principal component analysis (PCA), two major groups (Figs. 2 and 3) were observed. Each of this group was formed of accessions from each of the studied species. Group 1 consisted of twenty-four accessions from different geographical regions and mixed accessions of both the species under investigation. The subgroups of group 1 consisted largely on the basis of geographical locations rather than species basis. Group 2 contained accessions from different states like Himachal Pradesh, Rajasthan, Haryana, and Uttarakhand, but majority of accessions from Haryana grouped in this cluster, and out of twelve, the nine accessions from Haryana were included in this group. Two accessions, namely, Moonak 2 (Pb25) and Punjabi Uni (Pb23), remained as outlier and grouped outside the two major groups. AMOVA showed 96% variance within populations and 4% variance among population (Table 3 and Fig. 4). Mantel test showed nonsignificant correlation between geographic and genetic distance (Supplementary Fig. 1).

Dendrogram of 48 Ziziphus accessions based on 23 SSR markers data showing clustering of all accessions into two major groups

Principal coordinates analysis based on Eigenvalues calculated from 23 SSR markers. The 48 accessions were assigned into two groups

AMOVA showing genetic variance within and among populations
Discussion
SSR diversity and structure
Genetic diversity and population structure of Ziziphus germplasm from India is needed for its improvement in future and for the conservation of diverse and promising accessions. Genetic diversity gives the estimates of DNA polymorphism of the analyzed germplasm, and this polymorphism can be used in future for improving and manipulating the germplasm for various purposes. In the genus Ziziphus, the characterization of the cultivars had been largely based on morphological characters and their uses [32]. However, molecular marker studies have also been initiated at by different research groups at various places. The molecular studies in the 5 ber cultivars (Z. mauritiana) of Saudi Arabia were done using ISSR markers and it has been observed that a cultivar called Um-sulaem was paraphyletic to the other four [38] accessions analyzed. In Z. mauritiana, some other workers also conducted research using RAPD and ISSR [13, 47]. Furthermore, the two varieties of Indian jujube were also found genetically similar using RAPD markers [63]. The similar study was conducted in the same species using nr DNA and RAPD primers, and intraspecific variations were reported with about 85% polymorphism to separate delineate of the populations into 4 clusters [59]. Most recently, there is a maiden report of using SSR markers in Z. jujuba from China, and reported high genetic diversity (98.2%) in corresponding 3 clusters was observed using 31 primer pairs [65]. The present study differs from the previous as the germplasm collected is from diverse geographical locations and inclusion of two species.
To investigate the genetic relationship between the domesticated and wild jujube populations, chloroplast microsatellite markers (cpSSR) were developed by Huang et al. [23]. Using these cpSSR markers, the number of alleles per locus was found between two and four which is exactly like the alleles obtained in present study. Furthermore, the values of diversity indices were almost similar to the present study. Chaogun [68] used 24 SSR markers to explore the genetic diversity, genetic structure, and core collection of Ziziphus jujuba. STRUCTURE analysis and multivariate analyses (cluster and PcoA) were also done for the grouping of jujube accessions. Fu et al. [18] used SSR markers in Chinese jujube (Ziziphus jujuba Mill.) for population genetics, and the average number of alleles per locus was found 12.8 which was much greater than the number of allele obtained in present study. Using 11 ISSR primers to access genetic diversity within and among 34 accessions of Z. spina-christi collected from different regions of Saudi Arabia, Saleh Alans et al. (2016) obtained 109 scorable loci, of which 93.4% were found to be polymorphic. The size of amplified bands ranged between 250 and 3000 bp. Significant variability was observed in 10 selected Z. nummularia accessions based on various quantitative and qualitative characteristics of leaves, fruits, and seeds. Akhtar et al. [1] used 11 ISSR primers which showed 86.58% polymorphism. A pairwise similarity coefficient among all the 10 accessions were found to ranged from 0.45 to 0.77. Singh et al. [57] investigated genetic diversity among 47 cultivated accessions of Z. mauritiana and one wild accession of Z. nummularia using 18 ISSR markers; a total 167 products were detected, of which 89.96% were reported polymorphic. Cluster analysis based on UPGMA method and Bootstrap analysis separated all the 48 Ziziphus genotypes in 4 distinct clusters, and they were found to be divergent on the bases of Jaccard coefficient. Similar finding was observed in present study as dendrogram is showing divergent clustering as well as mixing of accessions. Saha et al. [45] studied genetic relationship among 26 fruit cultivars of Z. mauritiana which included 6 accessions of wild-type Z. mauritiana, two accessions of Z. nummularia, and 1 accession of Z. xylopyrus. The PIC values range between 0.23 to 0.46 and 0.11 to 0.36, respectively, for the RAPD and ISSR primers which are somewhat lower values as observed in present investigation using SSR markers. Thirty-eight microsatellites were isolated from Z. mauritiana to evaluate genetic diversity by Chiou et al. [11]. The PIC values ranged from 0.248 to 0.889. The number of alleles per locus ranged from 2 to 13 which were higher as compared to present study. However, most of the research work has been done in China, and Indian germplasm also requires similar works for the proper characterization, utilization, and conservation. Cluster analysis using dendrogram, principal component analysis (PCA), and STRUCTURE of the studied species was done; all of the methods showed two major groups (Figs. 2, 3 and 5). Each of this group was formed of mixed accessions from each of the studied species. Although majority of grouping was according to geographical locations, some exceptional mixing events cannot be neglected, and insights into these events are needed to make the things more clear. As both the species are cross-pollinated and at many sites both species were reported to occur in vicinity to each other, the cross-pollination may be a regular process between these two species. This mixing is also supported by Mantel test that showed nonsignificant correlation between geographic and genetic distances (Supplementary Fig. 1). On the other hand, pollens can be driven by wind to distant locations; these phenomena can be the reasons behind germplasm exchange and mixing. Furthermore, AMOVA indicate that larger portion of variance is within populations rather than among populations.

a Graph showing the best value of k at 2. b Bar plot of 48 accessions of Ziziphus showing two clusters by STRUCTURE. These clusters represent two populations with admixed individuals
Conclusion
In the present study, SSR markers showed high genetic diversity and mixing of gene pools in studied accessions of both jujube species. Results showed that there are two genetic stocks contributing in analyzed accessions. We found no specific correlation between different accessions of same species on the basis of geographical locations. The results of this research work can be useful in future research works in Ziziphus species to understand the spread of species and sharing of genomes between wild and cultivated germplasm. Furthermore, identification of diverse accessions based on minute morphological differences as well as at DNA level can be done for conservation and for initiating new breeding programs in Ziziphus species.
Availability of data and materials
Data analysis details were available from corresponding author on request.
References
Akhtar Y, Choudhary R, Sharma KC, Sharma M (2017) Genetic variability and relationship studies of Ber Ziziphus nummularia (Burm. F.) using morphological and molecular markers. Pharmacogn J. 9(3):417–424
Alansi S, Tarroum M, Al-Qurainy F, Khan S, Nadeem M (2016) Use of ISSR markers to assess the genetic diversity in wild medicinal Ziziphus spina-christi (L.) Willd. collected from different regions of Saudi Arabia, Biotechnology & Biotechnological Equipment 30:5, 942–7. https://doi.org/10.1080/13102818.2016.1199287
Bhandawat A, Sharma V, Sharma H, Sood A, Sharma RK (2014) Development and cross transferability of functionally relevant microsatellite markers in D. latiflorus and related bamboo species. J Genet 94:48–55. https://doi.org/10.1007/s12041-014-0377-9
Bhandawat A, Sharma V, Singh P, Seth R, Nag A, Kaur J, Sharma RK (2019) Discovery and utilization of EST-SSR marker resource for genetic diversity and population structure analyses of a subtropical bamboo, Dendrocalamus hamiltonii. Biochem Genet 57(5):652–672
Botstein D, White KL, Skolnick M, Davis RW (1980) Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am J Hum Genet 32:314–33l
Boussaid M, Taïbi K, Abderrahim LA, Ennajah A (2018) Genetic diversity of Ziziphus lotus natural populations from Algeria based on fruit morphological markers. Arid Land Research and Management
Chahota RK, Sharma V, Rana M, Sharma R, Choudhary S, Sharma TR, Shirasawa K, Hirakawa H, Isobe SN (2020) Construction of a framework linkage map and genetic dissection of drought- and yield-related QTLs in horsegram (Macrotylomauniflorum). Euphytica, 216, Article number: 61
Chahota RK, Shikha D, Rana M, Sharma V, Nag A, Sharma TR, Rana JC, Hiakawa H, Isobe S (2017) Development and characterisation of SSR markers to study genetic diversity and population structure of horsegram germplasm (Macrotyloma uniflorum). PlantMol Biol Rep. https://doi.org/10.1007/s11105-017-1045-z.
Chakravarthi BK, Naravaneni R (2006) SSR marker based DNA fingerprinting and diversity study in rice (Oryza sativa L.) African. J. Biotech 5:684–688
Chen W, Hou L, Zhang Z, Pang X, Li Y (2017) Genetic diversity, population structure, and linkage disequilibrium of a core collection of Ziziphus jujuba assessed with genome-wide SNPs developed by genotyping-by sequencing and SSR markers. Front Plant Sci 8:575. https://doi.org/10.3389/fpls.2017.00575.
Chiou CY, Chiang YC, Chen CH, Yen CR, Lee SR, Lin YS, Tsai CC (2012) Development and characterization of 38 polymorphic microsatellite markers from an economically important fruit tree, the Indian jujube. Am J Botany:199–202
Collard BCY, Mackill DJ (2008) Marker-assisted selection: an approach for precision plant breeding in the twenty-first century. Philos Trans Royal Society B: Biol Sci 363:557–572
Devanshi, Singh AK, Sharma P, Singh B, Singh R, Singh NK (2007) Molecular profiling and relationship among ber (Zizizphus sp.) genotypes using RAPD markers. Indian J Genet 67(2):121–7. https://doi.org/10.1080/15324982.2018.1424742
Doyle JJ, Doyle JL (1990) Isolation of plant DNA from fresh tissue. Focus 12(1):13
Earl DA, Von Holdt BM (2011) STRUCTURE HARVESTER: a website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv Genet Resour 4:359–361
Evanno G, Regnaut S, Goudet J (2005) Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Mol Ecol 14:2611–2620
Falush D, Stephens M, Pritchard JK (2007) Inference of population structure using multi locus genotype data: dominant markers and null alleles. Mol Ecol Notes 7(4):574–578
Fu PC, Zhang YZ, Ya HY, Gao QB (2016) Characterization of SSR genomic abundance and identification of SSR markers for population genetics in Chinese jujube (Ziziphus jujuba Mill.). Peer J 4:e1735. https://doi.org/10.7717/peerj.1735.
Gupta RB, Sharma S, Sharma JR, Panwar RD (2003) Study on vegetative characters of some cultivated and wild forms of ber (Ziziphus spp.). Haryana J Hort Sci 32(1/2):15–18
Hartl DL, Clark AG (1997) Principles of population genetics, 3rd edn. Sinauer Assoc, Sunderland, Massachsetts, p 163
Hartl DL, Jones EW (2005) DNA structure and DNA manipulation. In Genetics: analysis of genes and genomes, 5th edn. Ch. 2: 36-85. Sudbury: Jones and Bartlett Pub
Heikrujam M, Kumar J, Agrawal V (2015) Genetic diversity analysis among male and female Jojoba genotypes employing gene targeted molecular markers, start codon targeted (SCoT) polymorphism and CAAT box-derived polymorphism (CBDP) markers. Meta Gene 5:90–97
Huang J, Yang X, Zhang C, Yin X, Liu S, Li X (2015) Development of chloroplast microsatellite markers and analysis of chloroplast diversity in Chinese jujube (Ziziphus jujuba Mill.) and Wild jujube (Ziziphus acidojujuba Mill.). Plos One 10(9):0134519. https://doi.org/10.1371/journal.pone.0134519.
Jayaswall K, Sharma H, Bhandawat A, Sagar R, Yadav VK, Sharma V, Mahajan V, Rot J, Singh M (2019) Development of intron length polymorphic (ILP) markers in onion (Allium cepa L.), and their cross-species transferability in garlic (A. sativum L.) and wild relatives. Genet Resour Crop Evol. https://doi.org/10.1007/s10722-019-00808-3.
Kaldate R, Rana M, Sharma V, Hirakawa H, Kumar R, Singh G, Chahota RK, Isobe SN, Sharma TR (2017) Development of genome-wide SSR markers in horsegram and their utilization for genetic diversity and cross-transferability analysis. Mol Breed 37:103. https://doi.org/10.1007/s11032-017-0701-1.
Kalinowski ST, Taper ML, Marshell TC (2007) Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment. Mol Ecol 16:1099–1106
Kamunya SM, Wachira FN, Pathak RS, Korir R, Sharma V, Kumar R, Bhardwaj P, Chalo R, Ahuja PS, Sharma RK (2010) Genomic mapping and testing for quantitative trait loci in tea (Camellia sinensis (L.) O. Kuntze). Tree Genet Genomes 6:915–929
Kaur K, Sharma V, Singh V, Wani MS, Gupta RC (2016) Development of novel SSR markers for evaluation of genetic diversity and population structure in Tribulus terrestris L. (Zygophyllaceae). 3Biotech 6:156
Koul PM, Sharma V, Rana M, Chahota RK, Kumar S, SharmaTR (2017) Genetic structure and interrelationships in lentil species using morphological and SSR markers. 3Biotech 7:83
Li S, Guo M, Fu P, Liu H, Zhao X (2018) Genetic diversity and population structure of Chinese jujube (Ziziphus jujuba Mill.) and sour jujube (Ziziphus acidojujuba Mill.) using inter-simple sequence repeat (ISSR) Markers. Peers J Preprints 6:e27088v1
Liang T, Sun W, Ren H, Ahmad I, Vu N, Maryam HJ (2019) Genetic diversity of Ziziphus mauritiana germplasm based on SSR markers and ploidy level estimation. Planta 249(6):1875–1887
Liu M, Zhao J, Cai Q et al (2014) The complex jujube genome provides insights into fruit tree biology. Nature Communications 5:5315. https://doi.org/10.1038/ncomms6315.
Mehta R, Sharma V, Sood A, Sharma M, Sharma RK (2011) Induction of somatic embryogenesis and analysis of genetic fidelity of in vitro-derived plantlets of Bambusa nutans Wall., using AFLP markers. European J Foreest Res 130:729–736
Mishra T, Khullar M, Bhatia A (2011) Anticancer potential of aqueous ethanol seed extract of Ziziphus mauritiana against cancer cell lines and ehrlich ascites carcinoma. Evidence based Complementary and Alternative Medicine Vol Article ID 765029. https://doi.org/10.1155/2011/76502911.
Moustafa MF, Hesham AEL, Quraishi MS, Alrumman SA (2016) Variations in genetic and chemical constituents of Ziziphus spina-christi L. populations grown at various altitudinal zonation up to 2227 m height. J. of Gen Eng Biotechnol 14(2):349–362
Nabavi ST, Farahan F, Sheidai M, Poursakhi K, Naeini MR (2019) Population genetic study of Ziziphus jujuba Mill.: insight in to wild and cultivated plants genetic structure. Caryologia 72(4):85–92. https://doi.org/10.13128/caryologia-405.
Nyanga LK, Gadaga TH, Nout MJR, Smid EJ, Boekhout T, Zwietering MH (2013) Nutritive value of masau (Ziziphus mauritiana) fruits from Zambezi valley in Zimbabwe. Food Chem 138(1):168–172
Obeed RS, Harhash MM, Abdel-Mawgood AL (2008) Fruit properties and genetic diversity of five ber (Ziziphus mauritiana Lamk.) cultivars. Pakistan J Bio Sci 11:888–893
Patial A, Paul S, Sharma V (2017) Genetic diversity analysis using RAPD and ISSR markers revealed discrete genetic makeup in relation to fiber and oil content in Linum usitatissimum L. genotypes. The. Nucleus. https://doi.org/10.1007/s13237-017-0206-7.
Pawlowska AM, Camangi F, Bader A, Braca A (2009) Flavonoids of Zizyphus jujube L. and Zizyphus spina-christi (L.) Willd (Rhamnaceae) fruits. Food Chemistry 112:858–862
Perrier X, Jacquemoud-Collet, JP (2006) DARwin software http://darwin.cirad.fr/darwin
Pritchard JK, Stephens M, Donnelly P (2000) Inference of population structure using multi locus genotype data. Genetics 155:945–959
Raina SN, Ahuja PS, Sharma RK, Das SC, Bhardwaj P, Negi R, Sharma V et al (2012) Gen Struct Divers India Hybrid tea Genet Res CropEvol. https://doi.org/10.1007/s10722-011-9782-6.
Rana JC, Chahota RK, Sharma V, Rana M, Verma N, Verma B, Tilak R Sharma TR (2015) Genetic diversity and structure of Pyrus accessions of Indian Himalayan region based on morphological and SSR markers. TreeGenet Genomes Article ID 821
Saha D, Srivastava SC, Ramani R (2013) Genetic relationships among fruit cultivars and host plants of Indian lac insect in ber (Ziziphus mauritiana Lam.) revealed by RAPD and ISSR markers. Indian J Biotech 12:170–177
Saran PL, Godara AK, Sehrawat SK (2006) Characterization of ber (Ziziphus mauritiana Lamk.) genotypes. Haryana J Hort Sci 35(3/4):215–218
Sehrawat SK, Boora KS, Dahiya DS, Ahlawat VP, Goyal RK (2006) DNA fingerprinting of ber (Ziziphus mauritiana Lamk.) germplasm. Haryana J Horticultural Sci 35:213–214
Sharma H, Kumar P, Singh A, Aggarwal K, Roy J, Sharma V, Rawat S (2020) Development of polymorphic EST-SSR markers and their applicability in genetic diversity evaluation in Rhododendron arboreum. Mol Biol Rep. https://doi.org/10.1007/s11033-020-05300-1.
Sharma H, Kumar R, Sharma V, Kumar V, Bhardwaj P, Ahuja PS, Sharma RK (2011) Identification and cross-species transferability of 112 novel unigene-derived microsatellite markers in tea (Camelliasinensis). Am J Bot 98:e133–e138
Sharma RK, Gupta P, Sharma V, Sood A, Mohapatra T, Ahuja PS (2008) Evaluation of rice and sugarcane SSR markers for phylogenetic and genetic diversity analyses in bamboo. Genome 51:91–103
Sharma S, Sharma V, Chhabra M, Rathour R, Sharma KD, Kapila RK (2014) Characterization of novel polymorphic microsatellite markers in Dactylorhiza hatagirea: a critically endangered orchid species from Western Himalayas. Conserv Genet Resour. https://doi.org/10.1007/s12686-014-0361-y.
Sharma V, Bhardwaj P, Kumar R, Sharma RK, Sood A, Ahuja PS (2009) Identification and cross-species amplification of EST derivedSSR markers in different bamboospecies. Conserv Genet10 3:721–724
Sharma V, Rana M, Katoch M, Sharma PK, Ghani M, Rana JC, Sharma TR, Chahota RK (2015a) Development of SSR and ILP markers in horsegram (Macrotyloma uniflorum), their characterization, cross-transferability and relevance for mapping. Mol Breed 35:102
Sharma V, Wani MS, Singh V, Kaur K, Gupta RC (2017) Development and characterization of novel microsatellite markers in Trillium govanianum- a threatened plant species from north-western Himalaya. 3Biotech7:190
Sharma V, Sharma TR, Rana JC, Chahota RK (2015b) Analysis of genetic diversity and population structure in horsegram (macrotyloma uniflorum) using RAPD and ISSR markers. Agric Res 4(3):221–230
Shen J, Sun Z, Hou S, Liu R, Wang Y, Guo P (2020) Assessment of genetic diversity in Ziziphus jujube Mill. cultivars derived from northern China using inter-simple sequence repeat markers. Crop Science60:320–329
Singh AK, Devanshi SP, Singh R, Singh B, Koundal KR, Singh NK (2007) Assessment of genetic diversity in Ziziphus mauritiana using inter-simple sequence repeat markers. J. Plant Biochem Biotechn 16(1):35–40
Singh AK, Sharma RK, Singh NK, Bansal KC, Koundal KR, Mohapatra T (2006) Genetic diversity in ber (Ziziphus sp.) revealed by AFLP markers. J Hort Sci Biotechnol 81:205–210
Singh SK, Meghwal PR, Pathak R, Bhatt RK, Gautam R (2014) Assessment of genetic diversity among Indian jujube varieties based on nuclear ribosomal DNA and RAPD polymorphism. Agric Res. 3:218–228
Singh SK, Chhajer S, Pathak R, Bhatt RK, Kalia RK (2017a) Genetic diversity of Indian jujube cultivars using SCoT, ISSR, and rDNA markers. Tree Genet Genomes 13:12
Staub JC, Serquen FC, Mccreight JA (1997) Genetic diversity in cucumber (Cucumis sativus L.). An evaluation of Indian germplasm. Gen Res Crop Evolution 44:315–326
Tabin S, Kamili AN, Ganie SA, Zargar O, Sharma V, Gupta RC (2016) Genetic diversity and population structure ofRheum species in Kashmir Himalayas based on ISSR markers. Flora 223:121–128
Thanna EM, Yousif AM, Farag ME (2011) Morphlogical, physiological and biochemical genetic studies on some Indian jujube (Ziziphus mauritiana Lamak.) varieties grown at El Nubaria. Egyptian J Agricultural Res 89:1499–1525
Vashishtha BB (2001) Ber varieties: a monograph. Agrobios (India), Agro House, Behind Nasrani Cinema, Chopasani Road, Jodhpur, Rajasthan, India, p 97
Wang S, Liu Y, Ma L, Liu H, Tang Y, Wu L, Wang Z, Li Y, Wu R, Pang X (2014) Isolation and characterization of microsatellite markers and analysis of genetic diversity in Chinese Jujube (Ziziphus jujuba Mill.). Plos One 9:e99842
Wani MS, Gupta RC, Munshi AH, Sharma V (2018a) Development and characterization of SSR markers in Himalayantree species Betula utilis D. Don, J Forestry Res (Accepted)
Wani MS, Gupta RC, Munshi AH (2018b) Sharma V. Genetic diversity andstructure of Betula utilis accessions of north-western Himalaya based on RAPD and ISSR markers, The Nucleus (Accepted)
Xu C, Gao J, Du Z, Li D, Wang Z, Li Y, Pang X (2016) Identifying the genetic diversity, genetic structure and a core collection of Ziziphus jujube Mill. var. jujuba accessions using microsatellite markers. Sci Rep 6:31503. https://doi.org/10.1038/srep31503.
Xu Y (2010) Molecular plant breeding. CAB International
Zhang C, Yin X, Li X, Huang J, Wang C, Lian C (2013) Development and characterization of microsatellite markers for sour jujube (Ziziphus jujuba var. spinosa). Indian J Genet 73(3):338–341
Acknowledgement
The authors are grateful to the Department of Biotechnology (DBT) for providing financial assistance under IPLS-DBT scheme. We are also highly thankful to the Head, Department of Botany, Punjabi University Patiala for providing necessary lab facilities during the work.
Funding
We are thankful to Department of Biotechnology (DBT), Government of India, for providing facilities under DBT-IPLS programme (Reference number: BT/PR 4548/INF/22/146/2012) at Punjabi University Patiala.
Author information
Authors and Affiliations
Contributions
Manuscript writing, experimentation, and analysis of data were done by Dr. Amit Sareen. Dr. Vikas Sharma involved in technical guidance, manuscript proofreading, and data analyses and Dr. Raghbir Chand Gupta overall supervision of the study.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Supplementary Fig. 1
Showing correlation between Genetic and geographic distances of 48 samples used in present study.
Additional file 2: Supplementary Table 1
Axis wise Eigen values.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sareen, A., Sharma, V. & Gupta, R.C. Assessment of genetic diversity and population structure in wild Ziziphus species from northwest India using SSR marker technique. J Genet Eng Biotechnol 21, 4 (2023). https://doi.org/10.1186/s43141-022-00458-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43141-022-00458-6