
Abstract
Background
Thermostable cellulases are in constant demand for several biotechnological applications. Two thermophilic bacterial strains PW1 and PW2 isolated from Tattapani hot spring were found to have cellulolytic activity. Subsequently, PW1 and PW2 were identified and mined for genes encoding cellulase activity.
Results
Sequencing of the 16S rDNA of PW1 and PW2 identified them as Bacillus sp. PW1 (Acc no. KU711837) and Bacillus sp. PW2 (Acc no. KU711838), respectively, which clustered in the clades containing thermophilic members of Bacillus sp. and Geobacillus species. Phylogenetic analysis revealed that despite the morphological and sequence identities, Bacillus sp. PW1 and Bacillus sp. PW2 are different at the genetic level. The cellulase genes (~ 1.1 kb) of the two bacterial strains were amplified using primers designed against related thermophilic cellulases. Sequencing of the cellulase gene amplicons of PW1 and PW2 revealed that they encode proteins of 280 and 206 amino acid residues, respectively. Sequence and domain analysis of the protein products of PW1 and PW2 revealed that they belong to M42 family of aminopeptidase/endoglucanase. The PW2 endoglucanase coding sequence was submitted to Genbank under accession no. MH049504. The structures of putative endoglucanases of PW1 and PW2 were generated using 1VHE.A as template, which showed the presence of vast proportion of random coils. Molecular docking of the modeled endoglucanase proteins with various substrates and products of cellulases showed that carboxymethyl cellulose and maltose exhibit the highest binding affinity, while xylan and glucose the least.
Conclusions
The two thermophilic bacteria PW1 and PW2 and their endoglucanase gene can be further utilized for recombinant production of thermostable cellulases for their application in industries.
Similar content being viewed by others


Background
Degradation of cellulose by cellulolytic enzymes is mediated by differential cleavage of the glycosidic bonds linking the monomers/chains [1]. Several bacteria produce a group of enzymes known as cellulases, which act in a concerted manner to hydrolyze the β-1, 4-d-glycosidic bonds within the cellulose molecules [2]. Cellulases comprise three types of activities: endoglucanases or carboxymethylcellulases (CMCases; endo-β-1, 4-glucanases; EC 3.2.1.4), exoglucanases or cellobiohydrolases (CBHases; exo-β-1,4-glucanase; EC 3.2.1.91), and β-glucosidases (β-d-glucoside glucohydrolase; EC 3.2.1.21) [3,4,5]. Endoglucanases hydrolyze internal β-1, 4-D-glycosidic bonds in cellulose [6]. As a result, the length of the polymer decreases rapidly, and the concentration of reducing sugar increases slowly. Exoglucanases hydrolyze cellulose by detaching the cellobiose units from the non-reducing ends of cellulose [2, 7]. Cellulases are produced by several organisms, including bacteria and fungi [8, 9]. Thermophilic microbes are utilized for their cellulolytic potential due to fast growth rates at high temperatures, presence of complex multienzymes, and their occurrence in diverse environmental niches. There are a few thermophilic bacteria that are potent producers of cellulases, including Bacillus subtilis, Geobacillus pallidus, Bacillus licheniformis, Bacillus pumilis, Aneurinibacillus thermoaerophilus, and Clostridium thermocellum [10]. Thermophilic enzymes are ideal biocatalysts for the present day biotechnology because of their thermo-stability [11] and better yields under intense operational conditions [12].
Thermostable cellulases are gaining wide industrial and biotechnological significance due to their amenable use in harsh industrial processes like paper and textile production [13,14,15,16,17]. Moreover, the use of elevated temperatures in bioconversions minimizes the risk of contamination by common mesophiles and enhances the bioavailability and solubility of organic compounds [18]. Other values of elevated process temperatures include higher reaction rates due to a decline in viscosity and an increase in the diffusion coefficient of substrates and higher process yield due to enhanced solubility of substrates and products and favorable equilibrium displacement in endothermic reactions [19, 20]. Such cellulase enzymes can also be used as models for understanding thermo-stability and thermo-activity, which is beneficial for protein engineering. The hot springs of north-west Himalayas harbor a plethora of thermophilic microbes [21]. One such hot spring is located in Tattapani, Himachal Pradesh, India [21]. Diverse thermophiles were isolated from Tattapani hot spring possessing a variety of hydrolytic enzyme activity [21]. Previously, we reported the features of extracellular thermophilic cellulase produced by Geobacillus sp. of Tattapani hot spring [21]. In view of the importance of thermophilic cellulases for biotechnological applications, the present study was undertaken to screen thermophiles for cellulase activity. Two thermophilic bacterial strains PW1 and PW2 of Tattapani hot spring were screened and found to have cellulolytic activity. Towards overexpression of the thermophilic cellulase in recombinant system, genes encoding the cellulase activity of the two bacterial strains were mined and characterized by in silico studies.
Methods
Strains used in the study
Two thermophilic bacterial strains PW1 and PW2 were isolated from water samples of hot spring (water temperature ~ 70 °C) located in Tattapani, district Mandi, Himachal Pradesh, India [21]. The bacterial strains were cultured in nutrient broth (NB; Himedia Labs, India) and stored on nutrient agar at 4 °C and as 50% glycerol stocks at − 80 °C.
Biochemical analyses of the bacterial isolates
Biochemical tests such as catalase, urease, nitrate reduction, and oxidase tests were carried out to study the biochemical characteristics of thermophilic isolates PW1 and PW2 [22].
Optimization of physical parameters for growth of PW1 and PW2
Bacterial isolates were grown in nutrient broth adjusted to different pH ranging from 4 to 10, and incubated at 60 °C for 24 h with shaking at 200 rpm. Effect of temperature was measured by culturing the bacterial isolates in nutrient broth at different incubation temperatures (30–80 °C) for 24 h. Cell density was determined by measuring absorbance at 600 nm. The growth profile at higher than 80 °C could not be studied due to evaporation of media at high temperature.
Screening of chemical parameters for growth of PW1 and PW2
Bacterial isolates were grown in minimal salt medium (M9) containing NH4Cl as nitrogen source supplemented with 1% of different carbon sources such as glucose, starch, sucrose, fructose, lactose, raffinose, galactose, glycerol, and sorbitol. Minimal salt medium (M9) containing glucose as carbon source complemented with 0.25% of different nitrogen sources such as yeast extract, peptone, tryptone, beef extract, casein hydrolysate, urea, and NH4Cl was used to study the role of nitrogen source on growth. The bacterial cultures of PW1 and PW2 were incubated at 60 °C with shaking at 200 rpm for 24 h. The effect on growth of PW1 and PW2 was studied by measuring cell density at 600 nm. Due to evaporation of media at high temperature, we could not study growth profile at temperatures higher than 80 °C.
Screening and estimation of cellulase activity of thermophilic bacterial isolates PW1 and PW2
The thermophilic bacterial isolates (PW1 and PW2) were grown at 60 °C, and an equal number of cells was spotted on NB agar medium supplemented with 1% carboxymethyl cellulose (CMC) to screen for cellulase activity and incubated at 60 °C for 24 h. The CMC agar plate was stained with Gram’s iodine solution (1%). The appearance of a clear zone around the bacterial growth indicated the utilization of CMC, thus indicative of cellulase activity. Cellulase activity was measured by the DNS method [23], through the determination of the amount of reducing sugars liberated from carboxymethyl cellulose (CMC) at 540 nm. Twenty micrograms of total protein as crude cell-free enzyme was added to 0.2 ml of 1% CMC. The reaction mixture was incubated at 60 °C for 30 min, and the reaction was stopped by the addition of 2.0 ml DNS reagent. Enzyme activity was calculated. One unit of enzyme activity is defined as the amount of enzyme liberating 1 μg of glucose. Enzyme blank and substrate blank were performed in the assays with only added enzyme source or substrate, respectively.
Molecular identification of cellulolytic thermophilic bacterial isolates PW1 and PW2 by 16S rDNA sequencing
The thermophilic bacterial isolates were cultured at 60 °C to an A600 of ~ 1.0, and the cells were harvested by centrifugation at 12,000 rpm for 10 min. Genomic DNA from each of the bacterial cell pellet was isolated as described [24]. For identification of thermophilic bacterial isolates, 100 ng of genomic DNA was subjected to PCR amplification of 16S rDNA gene using 27F and 1492R primers (Additional file 1: Table S1) [25]. The PCR thermal cycling conditions were as follows: initial denaturation at 94 °C for 2 min followed by 35 cycles of denaturation (94 °C, 30 s), annealing (45 °C, 30 s), and extension (72 °C, 2 min), with a final extension of 10 min at 72 °C. The PCR products were resolved on 1% agarose gel. The gel-purified PCR products of 16S rDNA gene were sequenced on both strands using the primers 27F and 1492R at Eurofins, Bangalore, India (https://www.eurofins.com). The nucleotide sequences were manually analyzed, overlapping sequences were removed, and the complete 16S rDNA sequence for both the bacterial strains was generated. The nucleotide sequences were analyzed by BLAST (blastn) search and compared against bacterial 16S rDNA sequences available in the Genbank database [26]. The phylogenetic tree was constructed using MEGA4 (http://www.megasoftware.net) [27]. The nucleotide sequences were submitted in the GenBank database (https://www.ncbi.nlm.nih.gov/genbank/).
Isolation of gene encoding cellulase from Bacillus sp. PW1 and Bacillus sp. PW2
To amplify the cellulase gene from the thermophiles PW1 and PW2, gene-specific primers were designed using cellulase gene of Geobacillus sp. Y412MC52 [GYMC52-2749]. The nucleotide sequence of the primers used for PCR amplification of cellulase gene is listed in Additional file 1: Table S1. The following PCR conditions were used for amplification of cellulase gene from genomic DNA of Bacillus sp. PW1 and Bacillus sp. PW2: initial denaturation at 94οC, 2 min, and 35 cycles of the following steps: denaturation at 94οC, 30 s; annealing at 48οC, 30 s; extension at 72οC, 2 min; and final extension at 72οC, 10 min. The amplified PCR products were resolved on 1% agarose gel, visualized under UV gel documentation system (Alpha Innotech, USA), and then purified with a gel extraction kit (Thermo Scientific Inc., USA). The gel-purified PCR products were subjected to sequencing on both the strands using the same primers. The complete cellulase gene sequence for each bacterial isolate was generated manually by removing overlapping sequences. The nucleotide sequence was converted into protein sequence using the Expasy translation tool (https://web.expasy.org/translate). The protein sequence obtained for each bacterial isolate was compared against other known bacterial protein sequences available in the Genbank database using BLAST (BLASTP) search [26]. The phylogenetic tree was constructed by neighbor joining using MEGA4 [27].
Multiple sequence alignment of predicted PW1 endoglucanase and PW2 endoglucanase protein sequences with their homologs
The amino acid sequences showing more than 90% similarity from BLAST hits against PW1 endoglucanase and PW2 endoglucanase were selected and subjected to sequence alignment using PRALINE software [28] (www.genome.jp/tools/clustalw/).
Conserved domain analysis and hydropathy plots of predicted PW1 endoglucanase and PW2 endoglucanase
The protein sequences of PW1 endoglucanase and PW2 endoglucanase were subjected to conserved domain analysis using Conserved Domain Database tool of NCBI (http://www.ncbi.nlm.nih.gov/Structure/cdd/). Hydropathy plots were generated using Expasy-Protscale (https://web.expasy.org/protscale/).
Molecular modeling and in silico docking of PW1 and PW2 endoglucanases with the substrates and reaction products involved in cellulase-mediated catalysis
In order to study enzyme/substrate interaction, homology models of PW1 endoglucanase and PW2 endoglucanase were generated using the Swiss model (https://swissmodel.expasy.org/). The template selected by the Swiss model was aminopeptidase/glucanase homolog of Bacillus subtilis strain168(PDB id. 1VHE.A Chain A), which showed 80.71% sequence identity with Bacillus sp. PW1 endoglucanase and 76.70% sequence identity with Bacillus sp. PW2 endoglucanase, respectively. The ligands selected for docking include CMC, cellulose, xylan, glucose, maltose, and dextrin. The structure of ligands was procured from ChemSpider database (http://www.chemspider.com) in .mol format and then converted to .pdb format using open Babel software (http://openbabel.org/). Docking of the ligands was performed with the surface models of PW1 and PW2 endoglucanases using Hex software (http://hex.loria.fr/). The molecular visualization of structural interactions between PW1 and PW2 endoglucanases and different ligands were studied using Pymol software (https://www.pymol.org/). Amino acids involved in protein-ligand interaction were retrieved using PDB sum software (www.ebi.ac.uk/thornton-srv/databases/pdbsum/Generate.html :).
Results and discussion
Thermophilic bacterial isolates PW1 and PW2 possess cellulolytic activity and identified as Bacillus sp. PW1 and Bacillus sp. PW2
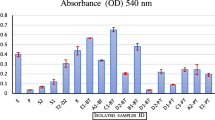
As shown in Fig. 1b, both PW1 and PW2 bacterial isolates produced clear zones (22 ± 0.1 mm and 20 ± 0.1 mm, respectively) on CMC agar at 60 °C, indicating the presence of cellulase enzyme. Quantification of CMC cellulase activity for both the bacterial species at 60 °C indicated the presence of predominantly extracellular cellulase activity (Fig. 1e). The thermophilic extracellular cellulase activities of PW1 and PW2 were found to be 1015 U/mg/min and 994 U/mg/min, respectively. For molecular identification of PW1 and PW2 isolates, total genomic DNA was isolated and used for amplification of 16S rDNA. An amplicon of 16S rDNA of ~ 1500 bp was observed for both PW1 and PW2 (Fig. 1c). The amplified 16S rDNA products were sequenced on both the strands, and a nucleotide sequence of 1435 bp and 1433 bp was obtained for PW1 and PW2, respectively. Analysis of the sequences by BLAST showed that the cellulolytic isolates PW1 and PW2 belong to Bacillus sp. showing 99% similarity to Bacillus sp. SP76. The 16S rDNA nucleotide sequences of the two bacterial isolates have been submitted to the NCBI GenBank database under the accession numbers KU711837 (Bacillus sp. strain PW1) and KU711838 (Bacillus sp. strain PW2), respectively. Phylogenetic analysis of 16S rDNA sequences of PW1 and PW2 with their close homologs revealed that Bacillus sp. PW2 clustered in the clade containing members of Bacillus and Geobacillus genera (Fig. 1d). On the other hand, Bacillus sp. PW1 emerged as an independent clade with no related members. It is interesting to note that despite the morphological and sequence identity, Bacillus sp. PW1 and Bacillus sp. PW2 are different at the genetic level.

a Growth of thermophilic bacterial isolates PW1 and PW2 on nutrient agar at 60 °C. b Detection of thermophilic cellulase activity of PW1 and PW2 bacterial isolates. Purified bacterial isolates of PW1 and PW2 were spotted on NA medium supplemented with 1% CMC and incubated at 60 °C for 24 h and flooded with 1% Gram’s iodine. The appearance of clear halo zones around bacterial growth indicates cellulase production by PW1 and PW2. c Amplification of 16S rDNA of PW1 and PW2 by PCR. Lanes 1 and 2: PCR products showing 16S rDNA amplification of PW1 and PW2, respectively. The size of 16S rDNA amplicon (~ 1500 bp) is indicated. M size marker (in kb). d Phylogenetic analysis of 16S rDNA sequences of Bacillus sp. PW1 and Bacillus sp. PW2. Dendrogram of PW1 and PW2 with related thermophiles (> 99% homology) is shown. The Jaccard’s coefficient scale for genetic relatedness is indicated below phylogenetic tree. e Quantitative cellulase assay of bacterial isolates PW1 and PW2
Bacillus sp. PW1 and Bacillus sp. PW2 exhibit differences in morphological, biochemical, and growth characteristics
At the morphological level, Bacillus sp. PW1 appear small rod shaped, forming creamish colonies, while Bacillus sp. PW2 are long rod shaped and form whitish colonies (Fig. 1a; Table 1). Both the isolates were positive for Gram’s staining, nitrate utilization, catalase, and oxidase reactions. However, Bacillus sp. PW1 was urease negative and glutaminase positive, and Bacillus sp. PW2 was urease positive and glutaminase negative (Table 1). Both PW1 and PW2 were found to be strict thermophiles, with no significant growth below 50 °C (Additional file 1: Figure S1). Optimum pH and temperature for growth of both the bacterial isolates were pH 8 and 80 °C, respectively (Additional file 1: Figure S1; Table 1). Among different carbon sources tested, galactose and glucose were the best carbon source for Bacillus sp. PW1 and Bacillus sp. PW2, respectively (Additional file 1: Figure S2; Table 1). Although all the carbon sources supported the growth of bacteria, sorbitol was found to be a poor carbon source (Additional file 1: Figure S2). Among different nitrogen sources studied, ammonium chloride was the best nitrogen source for the growth of Bacillus sp. PW1, while tryptone was the best nitrogen source for the growth of Bacillus sp. PW2 (Additional file 1: Figure S2). However, urea and casein hydrolysate were the least effective for the growth of both PW1 and PW2 isolates (Additional file 1: Figure S2). Thus, the two bacterial isolates exhibit differences in enzymatic and growth characteristics, despite being similar at the sequence level.
Bacillus sp. PW1 and Bacillus sp. PW2 possess genes encoding endoglucanase of M42 aminopeptidase/endoglucanase family
To mine the genes encoding cellulase activity from the two thermophiles, primers were designed based on the available gene sequences from closely related thermophiles. An amplicon of ~ 1.1 kb was observed for both PW1 and PW2 (Fig. 2a, b). The amplified fragments were sequenced on both the strands, and nucleotide sequences obtained were translated into amino acid sequences, yielding ORFs of 280 and 206 amino acids for PW1 and PW2, respectively (Additional file 1: Figure S3 and Figure S4). The translated ORFs were subjected to BLASTp analysis. Interestingly, the ORFs of both Bacillus sp. PW1 and Bacillus sp. PW2 showed 100% similarity with endoglucanase M of Geobacillus sp.WSUCF1 (EPR27003.1), as well as M42 family aminopeptidase of Geobacillus sp. WSUCF-018B (WP_100663940.1). Subsequently, the PW2 endoglucanase coding sequence was submitted to Genbank under accession no. MH049504. To validate the BLASTp results, phylogenetic analysis of the putative ORFs of PW1 and PW2 with their related hits was carried out. The ORFs of both PW1 and PW2 grouped with the members of M42 and M28 family peptidase of different Geobacillus sp. as well as endoglucanase/cellulase of thermophilic bacteria (Fig. 2c). The endoglucanase from Bacillus sp. PW1 formed a distinct clade, containing endoglucanase M, M42 peptidase endoglucanase, and M28 peptidase of Geobacillus sp., while PW2 endoglucanase coevolved with all the related members in the tree. It is rather intriguing that the PW1 and PW2 ORFs are similar to a peptidase as well as an endoglucanase, indicating the possibility for a dual enzyme activity of the putative ORFs. Therefore, in silico analysis of the two ORFs of PW1 and PW2 was undertaken to gain insights into the nature of the two proteins.

Isolation of genes encoding cellulase from the thermophiles Bacillus sp. PW1 and Bacillus sp. PW2. a Analysis of genomic DNA of PW1 (lane 1) and PW2 (lane 2) by agarose gel electrophoresis. b PCR amplification of cellulase gene from genomic DNA of PW1 and PW2. Lanes 1 and 2: PCR products showing amplification of cellulase gene of Bacillus sp. PW1 and Bacillus sp. PW2 (~ 1.1 kb), respectively. The size marker (M) bands are indicated (kb). c Dendrogram showing phylogenetic relationship of PW1 and PW2 endoglucanase protein sequences with their representatives showing > 99% homology. The Jaccard’s coefficient scale for genetic relatedness is indicated below phylogenetic tree
PW1 and PW2 endoglucanases share sequence identities with representative members of thermophilic cellulase as well as peptidases
The protein sequences of PW1 and PW2 endoglucanases were subjected to multiple sequence alignment with related cellulase sequences as well as peptidase [M42 family peptidases and YsdC [29]. Based on the sequence analysis, PW2 endoglucanase was found to be a truncated form of PW1 endoglucanase (Fig. 3). Interestingly, both the putative ORFs of PW1 and PW2 exhibited moderate to high degree of conservation with canonical endoglucanases as well as the two amino peptidases studied (Fig. 3). These results raise the possibility that the putative ORFs encode for dual enzyme activities. Although there are not many such reports, Maiti et al. [30] reported the dual activity of a protein isolated from Brevibacillus agri, which exhibits gelatinase as well as cellulase activities in vitro.

Sequence alignment of PW1 and PW2 endoglucanases with representative members of endoglucanases, cellulases, and aminopeptidase proteins. The protein sequences corresponding to M42 family peptidase of Geobacillus sp. WSUCF-018B (WP_100663940.1), endoglucanase M of Geobacillus sp. WSUCF1 (EPR27003.1), cellulase of thermophilic bacterium enrichment culture clone XM70 (AHN98139.1), cellulase of Geobacillus kaustophilus (GAD11976.1), cellulase of Bacillaceae thermophilic enrichment clone (ALY06177.1), aminopeptidase/glucanase homolog of Bacillus subtilis strain168 (pdb_1VHE.A Chain A), endoglucanase PW1 (Bacillus sp. PW1), and endoglucanase PW2 (Bacillus sp. PW2) were selected, and alignment was performed using PRALINE. The conservation scale is shown on the top
PW1 and PW2 endoglucanases possess domains of M42 peptidase/endoglucanase family
Conserved domain analysis of PW1 and PW2 endoglucanase proteins revealed the presence of M42 peptidase/endoglucanase domain of Zinc peptidase super family in both the proteins (Fig. 4a, b). These results are consistent with the phylogenetic analysis of the putative endoglucanases of PW1 and PW2 (Fig. 2c). Further, hydropathy plots indicate that both the proteins are hydrophilic in nature with hydropathy score of − 2.4 for PW1 endoglucanase and − 2.2 for PW2 endoglucanase, respectively (Fig. 4c, d). The presence of domain with endoglucanase activity in the M42 peptidase family has been challenged by Dutoit et al. [29], wherein CelM cellulase and TmPep1050 endoglucanase were shown to be an aminopeptidase. Therefore, the identity of the putative PW1 and PW2 endoglucanase proteins needs to be validated by functional analysis of the endoglucanase and peptidase activities.

Domain analysis and hydropathy analysis of PW1 and PW2 endoglucanases. Conserved domain analyses of endoglucanase of PW1 (a) and PW2 (b). Hydropathy plots for PW1 endoglucanase protein (c) and PW2 endoglucanase protein (d) were generated using Protscale
In silico molecular docking reveals that PW1 and PW2 endoglucanases exhibit the strongest interaction with CMC, substrate of cellulase enzyme
To gain insights into the structure of the putative endoglucanases, and their interaction with the substrates/products of cellulase, homology models were generated by the Swiss model based on the structure of 1VHE.A aminopeptidase/endoglucanase (Fig. 5). PW1 and PW2 endoglucanase sequences exhibited protein coverage of 100% and identity of 80.71% and 76.70% with the template, respectively. PW1 endoglucanase showed the presence of 32% alpha helices and 23% beta strands (Fig. 5a, b). As shown in Fig. 5c, d, PW2 endoglucanase contained 33% alpha helices and 21% beta strands. Both PW1 and PW2 endoglucanases had a large proportion (~ 50%) of the unstructured region, as indicated by the presence of linkers in the structures. Similar studies on cellulases from different Pseudomonas sp. indicated the predominance of random coils [31].

The three-dimensional molecular structures of PW1 and PW2 endoglucanases generated by homology modeling. Surface models (a, c) and ribbon models (b, d) of PW1 and PW2 endoglucanases were generated using aminopeptidase/glucanase homolog of Bacillus subtilis strain168 (pdb_1VHE.A) as a template. Alpha helices are colored green, beta strands are in blue, and linkers are shown in red
Subsequently, the 3D surface models were used for docking enzyme-substrate interaction with the substrates namely CMC, cellulose, xylan, and products/intermediates namely, glucose, maltose, and dextrin (Additional file 1: Figure S5 and Figure S6). The binding energy for the interactions [Etotal (kcal/mol)] was calculated (Tables 2 and 3). CMC and maltose were found to be the best interacting ligands for both PW1 and PW2 endoglucanases. As expected, xylan, a hemi-cellulose, and the products like glucose and dextrin exhibited lower binding energies for both the endoglucanases. However, maltose showed strong affinity for binding, indicating an alternate binding mechanism with the enzymes. Thus, molecular docking studies favor the possible function of the putative proteins as endoglucanases, despite belonging to M42 peptidase/endoglucanase family.
Conclusion
In the present study, we have explored two strict thermophiles of Tattapani hot spring (Himachal Pradesh, India), namely PW1 and PW2 for their cellulolytic potential. Both the bacterial strains exhibited profound thermophilic cellulase activity. The two isolates were identified as members of Bacillus genera by 16S rDNA sequencing, with high sequence similarities, yet distinct growth features and phylogenetic positions. The two thermophiles were mined for genes encoding cellulase activity and found to encode putative endoglucanases of a rather contentious dual family of enzymes namely M42 peptidase/endoglucanase. The putative endoglucanases of PW1 and PW2 were studied by in silico structural modeling and molecular docking with substrates of cellulases. Both of the putative proteins showed strongest binding with carboxymethyl cellulose and maltose. Biochemical studies of the putative endoglucanases in a recombinant expression system are required to validate their cellulase activities for potential industrial applications.
Availability of data and materials
Supporting data is provided in the supplementary material.
Abbreviations
- CBHases:
-
Cellobiohydrolases
- CMCases:
-
Carboxymethylcellulases
- CMC:
-
Carboxymethyl cellulose
- NB:
-
Nutrient broth
References
Onsori H, Zamani MR, Motallebi M, Zarghami N (2005) Identification of over producer strain of endo-β-1,4-glucanase in Aspergillus Species: Characterization of crude carboxymethyl cellulase. Afr. J. Biotechnol 4:26–30
Akiba S, Kimura Y, Yamamoto K, Kumagai H (1995) Purification and characterization of a protease-resistant cellulase from Aspergillus niger. J. Ferment. Bioeng 79:125–130
Parry JB, Stewart JC, Heptinstall J (1983) Purification of the major endoglucanase from Aspergillus fumigates Freseius. Biochem J. 213:437–444
Gielkens MMMC, Dekkers E, Visser J, Graaff LH (1999) Two cellubiohydrolase-encoding genes from Aspergillus niger require D-Xylose and the xylanolytic transcriptional activator XlnR for their expression. Appl Environ Microbiol. 65:4340–4345
Kang SW, Ko EH, Lee JS, Kim SW (1999) Over production of β-glucosidase by Aspergillus niger mutant from lignocellulosic biomass. Biotechnol Lett. 21(8):647–650
Siddiqui KS, Shemsi AM, Anwar MA, Rashid MH, Rajoka MI (1999) Partial and complete alteration of surface charges of carboxymethylcellulose by chemical modification: thermos stabilization in water-miscible organic solvent. Enzyme Microb Technol 24:599–608
Han SJ, Yoo YJ, Kang HS (1995) Characterization of a bifunctional cellulase and Its structural gene. J Biol Chem. 270:26012–26019
Moussa TAA, Tharwat NA (2007) Optimization of cellulase and β-glucosidase induction by sugarbeet pathogen Sclerotium rolfsii. Nagwa. Afr. J Biotechnol. 6(8):1048–1054
Suto M, Tomita F (2001) Induction and catabolite repression mechanisms of cellulase in fungi. J Biosci Bioeng. 92(4):305–311
Sharada R, Venkateswarlu G, Venkateshwar S, Rao MA (2013) Production of cellulase – a review. IJPCBS 3:1070–1090
Haki GD, Rakshit SK (2003) Developments in industrially important thermostable enzymes: a review. Bioresour Technol. 89:17–34
Andrade CMMC, Nei P Jr, G Antranikian (1999) Extremely thermophilic microorganisms and their polymerhydrolytic enzymes, Braz. J. Microbiol 30:287–298
Fredrich A, Antrakian G (1996) Keratin degradation by Fervidobacterium pennavorans, a novel thermophilic anaerobic species of the order Thermotogales. Appl Environ Microbiol. 62:2875–2882
Leuschner C, Antranikan G (1995) Heat stable enzymes from extremely thermophilic and hyper thermophilic microorganisms. World J Microbiol Biotechnol. 11(1):95–114
Zeikus J, Lee C, Lee Y, Saha B (1998a) Thermostable saccharides: new sources, uses and biodesigns. In: Kalegoris, E., Christakopoulos D., Kekos, D., Macris, B. (Eds.), Studies on the Solid-State Production of Thermostable Endoxylanases from Thermoascusaurantiacus: Characterization of Two Isoenzymes. J Biotechnol. l. 60:155–163
Zeikus J, Vielle C, Savachenko A (1998b) Thermozymes: biotechnology and structure-function relationships. Extremophiles 2(3):179–183
Wilkinson D, Reuter S, Zimmermann W (1998) Application of thermostable enzymes for carbohydrate modification. In: Contribution of the Fourth International Workshop on Carbohydrate as Organic Raw Materials, WUV- Universitatverlag, Vienna, p 176–184
Becker P, Abu-Reesh I, Markossian S, Antranikian G, Märkl H (1997) Determination of the kinetic parameters during continuous cultivation of the lipase producing thermophile Bacillus sp. IHI-91 on olive oil. Appl MicrobiolBiotechnol 48:184–190
Krahe M, Antranikian G, Markel H (1996) Fermentation of extremophilic microorganisms. FEMS Microbiol Rev. 18:271–285
Mozhaev VV (1993) Mechanism-based strategies for protein thermos stabilization. Trends Biotechnol. 11(3):88–95
Sharma P, Gupta S, Sourirajan A, Dev K (2015) Characterization of extracellular thermophilic cellulase from thermophilic Geobacillus sp. isolated from Tattapani hot spring of Himachal Pradesh, India. Int J Adv Biotechnol Res., 6:433–442
Smibert RM, Krieg NR (1994) American Society for Microbiology, Washington DC. Phenotypic characterization. In: Gerhardt P, Murray RG, Wood WA, Krieg NR (eds), Methods for General and Molecular Bacteriology., p611–654
Miller GL (1959) Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal.Chem. 31:426–428
Sambrook J, Fritsch EF, Maniatis TI (2012) Molecular cloning: a laboratory manual Spring Harbor Laboratory Press, New York, p19–25
Lane DJ (1991) Nucleic acid techniques in bacterial systematics. In: Stackebrandt E, Goodfellow M (eds) . Wiley, Chichester, UK, p115–175
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic local alignment search tool. J Mol Biol 215(3):403–410
Tamura K, Dudley J, Nei M, Kumar S (2007) MEGA 4: molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol Biol Evol. 24(8):1596–1599
Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 22(22):4673–4680
Dutoit R, Brandt N, Legrain C, Bauvois C (2012) Functional characterization of two M42 aminopeptidases erroneously annotated as cellulases. PloS one 7(11):e50639
Maiti S, Samanta T, Sahoo S, Roy S (2017) The dual carboxymethyl cellulase and gelatinase activities of a newly isolated protein from Brevibacillus agri ST15c10 confer reciprocal regulations in substrate utilization. J Mol Microbiol Biotechnol. 27:319–331
Pradeep NV, Anupama A, Vidyashree KG, Lakshmi P (2012) In silico characterization of industrial important cellulases using computational tools. Advances in life science and technology, vol 4, pp 2224–7181
Acknowledgements
The authors are thankful to Shoolini University, Solan, for the financial and infrastructural support provided for this study.
Funding
Shoolini University, Solan, HP, India provided infrastructural and consumables support for the study.
Author information
Authors and Affiliations
Contributions
DS performed all the experiments; PS isolated the bacterial strains; KD and AS conceived the study and participated in its design and coordination and helped to draft the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional file
Additional file 1:
Supplementary material. (DOCX 1660 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Sharma, D., Sharma, P., Dev, K. et al. Endoglucanase gene of M42 aminopeptidase/endoglucanase family from thermophilic Bacillus sp. PW1 and PW2 isolated from Tattapani hot spring, Himachal Pradesh, India. J Genet Eng Biotechnol 17, 4 (2019). https://doi.org/10.1186/s43141-019-0001-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43141-019-0001-8