
Abstract
Background
The Apiaceae family is among the most significant plant families because it contains both beneficial and poisonous plants. Due to their morphological similarity, these harmless and lethal species are frequently confounded. Cumin, fennel, and anise are the most prevalent members of the family Apiaceae in Egypt. Members of this family are routinely used as medical surrogates, so it is crucial that they are correctly identified and distinguished. DNA barcoding is a molecular technique used for identifying species and reconstructing phylogenetic trees.
Results
Six plants from this family were chosen for this study due to their medicinal importance, and four DNA barcoding loci (rbcL, matK, trnH-psaA, and ITS) were used to identify them. The amplicons were sequenced, and the comparative analysis was conducted between the sequences evaluated and the most significant Blast results. The DNA rbcL, trnH-psaA, and ITS barcodes exhibited similar amplicons among the six species of Apiaceae, while the trnH-psaA barcode exhibited different amplicons among the Apiaceae. Maximum likelihood approach was used to calculate the genetic distance between the sex species of Apiaceae. The most significant findings were that the one from four DNA barcoding was able to distinguish between distinct species and confirm their evolutionary belonging to this family.
Conclusions
The current study concludes that trnH-psbA and ITS DNA identifiers can be used to accurately identify, differentiate, and record Apiaceae species, while the rbcl DNA barcode appears to have fallen short of its intended purpose. So, the data that come from DNA barcodes could be used for the biodiversity assessment and the similarities between hazardous and commercial plants to resolve some of these deficiencies.
Similar content being viewed by others
1 Background
To keep the world’s healthcare system running, we need medicinal plants. Herbal remedies have been shown to cure a wide range of illnesses and disorders, sometimes with fewer side effects and at a lower cost than pharmaceutical options [26]. It is estimated that there are between 3600 and 3751 different species of plants in the Apiaceae family [24]. Many important phytochemicals, including phenolic compounds and flavonoids, are found in the Apiaceae family. Flavonoids’ antiviral, anticancer, antioxidant, and anti-inflammatory characteristics are only a few of their many positive health effects. In addition, they shield the heart and the brain from damage. Variations in the effects of flavonoids on certain cellular activities have been reported [29] but more research is needed. Essential oils, extracted from various species in this family, have approximately 760 different chemical classes with substantial therapeutic potential. Coriander seed oil has a high concentration of petroselinic acid. The European Commission approved its sale as a novel food additive in 2014 [20] in accordance with Regulation (EC) No 258/97 of the European Parliament and Council.
This family has a lot of plants, for example, Parsley (Petroselinum crispum L.), anise (Pimpinella anisum), coriander (Coriandrum sativum), cumin (Cuminum cyminum L.), dill (Anethum graveolens Mill.), fennel (Foeniculum vulgare Mill.), and caraway (Carum carvi L.) [10]. The presence of volatile chemicals is a telltale sign of these plants, which have long been thought to have somewhat negative medicinal effects on the body and mind. However, there are some dangerous members of the Apiaceae family. Hemlock water-dropwort (Oenanthe crocata L.), fool's parsley (Aethusa cynapium L.), poison hemlock (Conium maculatum L.), and water hemlock (Cicuta virosa L.) are some of the most well-known examples of these plants. Toxic species are sometimes mistaken for fragrant food species because of their similar chemical makeup and structure [21]. Traditional approaches to biodiversity assessment are time-consuming and rely on taxonomic data, which is becoming scarcer. Recent advances like molecular methods are useful tools for identifying certain clonal variations, and establishing genetic stability [1, 2, 11,12,13,14, 23]. As reported by [6], DNA barcoding may one day offer a faster and more accurate alternative to traditional methods of estimating species diversity that rely on expert field identification personnel.
DNA barcoding has had a significant favorable effect on biodiversity identification and categorization [17]. DNA barcodes have two main uses: (1) to determine the species of an unidentified material and (2) to help researchers discover new species by screening thousands of copies of a small number of reference genes. The chloroplast genome, which includes all the DNA sequences in a single plastid, has more information than any single-locus marker for identifying and classifying plant species. DNA barcodes that make use of chloroplast genomes actively to distinguish between plant species are an important area of research and development [30]. The Plant Working Group of the Consortium for the Barcode of Life (CBOL) has proposed using ribulose-1,5-bisphosphate carboxylase/oxygenase large subunit (rbcL) and maturase K (matK), both located in the plastid genome, as a standard barcode for plants, with the option of using additional markers to fill in any gaps. It has been postulated that the trnH and photosynthetic protein II D1 (trnH-psbA) plastid intergenic spacer region is another maker [19].
The Internal Transcribed Spacer (ITS1-4) is a powerful phylogenetic marker with substantial interspecies divergence at the species level. The ITS region has been proposed as a plant barcode because to its superior selective power over plastid areas at low taxonomic levels, especially in parasitic plants for which plastid barcodes give less precision [17].
In this study, we used DNA barcodes to differentiate between six commercially relevant species of the family Apiaceae. Both DNA barcodes (ITS1-4) and three chloroplast DNA barcodes (trnH-psbA, matK, and rbcL) were used to discover the genetic diversity within this family and draw the phylogenetic tree between these species.
2 Methods
2.1 Plant material
The experimental seedlings of six Apiaceae species were planted in compost soil-filled pots. Table 1 displays six economically relevant Apiaceae plant species that were studied in this study.
2.2 DNA extraction
Using the EasyPure® Genomic DNA Kit (Beijing Trans Gen Biotechnology Co., Ltd), we isolated DNA from 100 mg of three-week-old leaves from germinated seeds in accordance with the manufacturer's instructions. The purity and concentration of the DNA were evaluated using spectrophotometry and agarose gel electrophoresis (at 0.8% concentration). The DNA was then stored at -20 degrees Celsius until needed.
2.3 PCR amplification and purification
MatK, rbcL, and trnH-psbA were amplified from the plastid genome, whereas ITS was amplified from the nuclear genome. The primers, PCR cycle, and amplicon size for each primer are listed in Table 2. Six microliters of double-distilled water were added to the PCR reaction mixture to adjust the volume of the final product. A thermal cycler (Perkin Elmer GeneAmp PCR System 2400) was used to run all PCRs alongside negative controls. 1.5% agarose gel electrophoresis with 3l of 100bp plus DNA ladder (TransGen Biotech Co., Ltd., Cat. No. BM301) was used to examine the PCR results.
2.4 Data analysis and sequencing
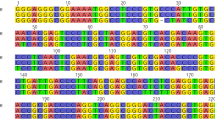
PCR products were purified and sequenced using Sanger Technology (Macrogen, Korea) for bidirectional sequencing of the rbcL, matK, trnH-psaA, and ITS barcode markers as shown in Fig. 1. The obtained sequences were assembled into contigs using BioEdit v.7.2.5 software [27]. Pairwise distance, Transition/Transversion, and Substitution Matrix were estimated with the MEGA11 software employing the Kimura-2-Parameter (K2P) model, and the contigs were aligned with ClustaW of MEGA11 to verify species identity. Relationships analysis among species were built in MEGA11 using the Maximum Likelihood (ML) approach based on the Kimura-2-Parameter (K2P) model [25]. All relationships analysis were given 500 replicates of the bootstrap test.

PCR products of different markers; (a) rbcL marker, (b) matK marker, (c) ITS marker, and (d) trnH-psbA marker for the following six plants: Parsley (pa), Khella Shaytani (Kh), Caraway (Ca), Cumin (Cu), Fennel (Fe), and Anise (An)
3 Results
3.1 PCR amplification
In this study, high-quality genomic DNA from six Apiaceae species was utilized to amplify four different barcodes. The PCR product evaluated by agarose gel (1.5%) revealed amplified amplicons ranging in size from 158 bp (exhibited by trnH-psaA) to 780 bp (exhibited by matK). These amplicons when sequenced revealed sequences ranged from 127 to 2333 bp. While rbcL, matK, and ITS exhibited one amplicon, the trnH-psaA exhibited two amplicons shown in Fig. 1 and Table 2. The size of the sequences obtained from the four DNA barcodes is shown in Table 3.
3.2 Analysis of data sequence
As presented in Tables 2 and 3, rbcL exhibited amplicon with 533 bp which revealed sequence ranged from 508 to 511 bp among the six Apiaceae species; the 508 bp was observed in Anise while the 511 bp was observed in Parsley. Similarly, ITS exhibited one amplicon with 750 bp which revealed sequences ranging between 699 and 709 bp among the Apiaceae species. While Fennel exhibited the 699 bp, Caraway exhibited the 705 bp sequence. While matK exhibited amplicon with 780 pb which revealed extreme sequences size ranged from 767 to 2333 bp among Apiaceae species, the Fennel exhibited the 767 bp sequence while Anise exhibited the 2333 bp sequence. On the other hands, the other studied species exhibited sequences ranged between 1016 and1243 bp. The trnH-psaA exhibited two amplicons with 158 and 250 bp, while the 158 bp amplicon was observed in Caraway, the 250 bp amplicon were observed in other five species. The 158 bp amplicon revealed sequence with 127 bp, while the 250 bp amplicon revealed sequences ranged from 184 to 191 bp.
The constructed sequences were compared using BLAST to check for species similarity. MatK, trnH-psbA, and ITS were all shown to be efficient DNA barcoding regions for species identification, despite the Plant Working Group of the Consortium for the Barcode of Life (PWG-CBOL) recommending matK and rbcL as core barcoding regions for plants, Khella Shaytani and Anise's species identities could not be determined using rbcL, whereas BLAST showed Pimpinella saxifrage instead of Pimpinella anisum (Accession No. of all Reference plants as stated in Table 4) and Ammi trifoliatum instead of Ammi majus. The pairwise distance values resulted for all barcodes (rbcL, matK, trnH-psaA, and ITS) being 0.01337, 0.3805, 0.3732, and 0.8018, respectively.
3.3 Relationships analysis among species
Each barcoding locus’s constructed sequence was aligned using MEGA11. Kimura’s 2-Parameter using MEGA11 was used to calculate pairwise distance and Transition/Transversion.
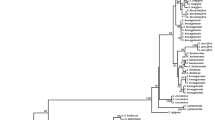
The dendrogram presented the figure of rbcL Fig. 2C that Carum carvi is more closely related to the toxic plant Oenanthe crocata than to any other species. While Fig. 2A shows that Anethum graveolens is genetically closer to the noxious plant Conium maculatum and Carum carvi is more closely related to the toxic plant Oenanthe crocata. In addition, the dendrogram presented the figure of trnH-psbA (shown in Fig. 2B) showed that Petroselinum crispum was more closely related to Ammi majus than Ammi trifoliatum (shown in Fig. 2C). However, the dendrogram presented the figure of ITS (shown in Fig. 2D) demonstrates that Carum carvi is more closely related to Cuminum cyminum than and Ammi majus was more closely Anethum foeniculum.

Phylogenetic relationships among some Apiaceae family constructed using MEGAx software by the maximum likelihood (ML) depending on four DNA barcodes, A = mat K, B = psb, C = rbcL and D = ITS
4 Discussion
The results of this study suggest that matK, trnH-psbA, and ITS are all effective DNA barcoding regions for species identification in the Apiaceae family [4, 28]. The existence of single nucleotide repeats, which generate frequent shifts in the reading frame, is responsible for the rapid pace of length divergence. There seems to be a lot of SNP repetitions in the genomes of many angiosperms. This result agrees with the conclusions made by [8], who found that, with the exception of the trnH-psbA region, the sequence of DNA coding loci has a reasonable read length in both directions, despite the PWG-CBOL recommending matK and rbcL as core barcoding regions for plants and shown that these markers are highly variable at the interspecific level, but relatively conserved at the intraspecific level.
However, rbcL was unable to distinguish between Khella Shaytani and Anise, and ITS misidentified Pimpinella saxifrage as Pimpinella anisum, and Ammi trifoliatum as Ammi majus. because rbcL was not as effective for species identification in Apiaceae. This is likely because 164 rbcL is a relatively slow-evolving gene, and therefore does not accumulate enough variation to 165 distinguish between closely related species.
The high pairwise distances and Transition/Transversion ratios between the six Apiaceae species sequenced in this study suggest that they are all distinct species. This is further supported by the fact that each species had a unique sequence for each of the three barcodes matK, trnH-psbA, and ITS. It is also worth noting that the study found that BLAST analysis of the rbcL sequences for Khella Shaytani and Anise could not determine their species identities. This suggests that there may be some taxonomic confusion surrounding these species, or that the rbcL sequences used in the study were not representative of the species.
Overall, the results of this study suggest that DNA barcoding is a powerful tool for species identification in Apiaceae, and that matK, trnH-psbA, and ITS are all good choices for DNA barcoding in Apiaceae. However, more research is needed to determine the best combination of markers to use for species identification in this family.
This finding is in line with recent studies showing that matK and rbcL are not always useful as barcodes for specific plant taxa [9, 22]. Although matK was effective in this investigation at identifying plant species and producing data, it was unable to reliably differentiate between the species. The trnH-psbA spacer, although relatively short and simple to amplify, is the plastid region with the most variability in angiosperms. Previous research [3, 5, 7, 15, 16, 18] supports our conclusion that trnH-psbA is the most effective DNA barcode for plant identification.
Furthermore, the ITS has made great strides in the identification of species. Sequences of rbcL, matK, trnH-psbA, and ITS were retrieved from the NCBI Gene repository for the following plant species: Genbank sequences for Anethum graveolens, Apium graveolens, Ammi visnaga, Cicuta maculate, Oenanthe crocata, Aethusa cynapium, Conium maculatum, Heracleum maximum, and Pastinaca sativa were used to construct phylogenetic trees using the phylogenetic maximum likelihood (ML) method. Thus, the best results were using both ITS and trnH-psbA, and the latter achieved the best results at the level of the DNA barcoding markers used.
5 Conclusion
This research clearly shows that the six Egyptian Apiaceae species can be distinguished from one another using the DNA barcodes rbcL, matK, trnH-psaA, and ITS. However, it was decided that rbcL was not enough for barcoding at the species level. Combining it with another Barcoding Loci may provide a more accurate result for members of this family. In addition, trnH-psbA and ITS did a great job of identifying species. We therefore recommend using a range of biochemical approaches to further distinguish between harmful and beneficial species, as DNA barcoding has shown a close connection between them (as evidenced by phylogenetic trees). We also recommend the combination of trnH-psbA and ITS. Table 5 also displays the nucleotide sequences that we obtained and uploaded to the NCBI database.
Availability of data and materials
The data that support the findings of this study are available from the corresponding author upon reasonable request.
References
Al-Taweel SK, Abdel-Aziz RM, Rabea K, Khaled K (2019) Studying cDNA SCoT in response to salinity stress in Stevia rebaudiana Bertoni. SABRAO J Breed Genet 51(3):281–294
Azzam C, Sultan F, Sayed M, Khaled K (2022) Gamma-rays and microwaves irradiation influence on guar (Cyamopsis Tetragonoloba): II–proteomic analysis linked to plant height and crude proteins. SABRAO J Breed Genet 54(5):1101–1112. https://doi.org/10.54910/sabrao2022.54.2.10
Bansal S, Thakur S, Mangal M, Mangal AK, Gupta R (2018) DNA barcoding for specific and sensitive detection of Cuminum cyminum adulteration in Bunium persicum. Phytomedicine 50:178–183. https://doi.org/10.1016/j.phymed.2018.04.023
CBOL Plant Working Group 1, Hollingsworth PM, Forrest LL, Spouge JL, Hajibabaei M, Ratnasingham S, Little DP (2009) A DNA barcode for land plants. Proc Natl Acad Sci 106(31):12794–12797. https://doi.org/10.1073/pnas.0905845106
Chen S, Yao H, Han J, Liu C, Song J, Shi L, Pang X (2010) Validation of the ITS2 region as a novel DNA barcode for identifying medicinal plant species. PLoS ONE 5(1):e8613. https://doi.org/10.1371/journal.pone.0008613
Costion C, Ford A, Cross H, Crayn D, Harrington M, Lowe A (2011) Plant DNA barcodes can accurately estimate species richness in poorly known floras. PLoS ONE 6(11):e26841. https://doi.org/10.1371/journal.pone.0026841
De Mattia F, Bruni I, Galimberti A, Cattaneo F, Casiraghi M, Labra M (2011) A comparative study of different DNA barcoding markers for the identification of some members of Lamiacaea. Food Res Int 44(3):693–702. https://doi.org/10.1016/j.foodres.2010.12.032
Fazekas AJ, Steeves R, Newmaster SG (2010) Improving sequencing quality from PCR products containing long mononucleotide repeats. Biotechniques 48(4):277–285. https://doi.org/10.2144/000113369
Fu YM, Jiang WM, Fu CX (2011) Identification of species within Tetrastigma (Miq) Planch (Vitaceae) based on DNA barcoding techniques. J Syst Evol 49(3):237–245. https://doi.org/10.1111/j.1759-6831.2011.00126.x
Jamalova DN, Gad HA, Akramov DK, Tojibaev KS, Musayeib NMA, Ashour ML, Mamadalieva NZ (2021) Discrimination of the essential oils obtained from four Apiaceae species using multivariate analysis based on the chemical compositions and their biological activity. Plants 10(8):1529. https://doi.org/10.3390/plants10081529
Khaled KA, El-Arabi NI, Sabry NM, El-Sherbiny S (2018) Sugarcane genotypes assessment under drought conditions using amplified fragment length polymorphism. Biotechnology 17:120–127. https://doi.org/10.3923/biotech.2018.120.127
Khaled KA, El-Demardash IS, Amer EAM (2015) Genetic polymorphism among some sugarcane germplasm collections as revealed by RAPD and ISSR analyses. Life Sci J 12(3):159–167
Khaled KAM, Sultan FM, Azzam CR (2022) Gamma-rays and microwave irradiation influence on guar (Cyamopsis tetragonoloba): I—markers assisted selection for responding to mutagen agents. SABRAO J Breed Genet 54(2):331–349. https://doi.org/10.54910/sabrao2022.54.2.10
Khaled KA, Habiba RM, Bashasha JA, Azzam CR, MH AA (2023) In silico and genetic analysis related to tillering ability in maize. SABRAO J Breed Genet 55(1):156–162. https://doi.org/10.54910/sabrao2023.55.1.15
Kress WJ, Erickson DL (2007) A two-locus global DNA barcode for land plants: the coding rbcL gene complements the non-coding trnH-psbA spacer region. PLoS ONE 2(6):e508. https://doi.org/10.1371/journal.pone.0000508
Kress WJ, Wurdack KJ, Zimmer EA, Weigt LA, Janzen DH (2005) Use of DNA barcodes to identify flowering plants. Proc Natl Acad Sci 102(23):8369–8374. https://doi.org/10.1073/pnas.050312310
Li X, Yang Y, Henry RJ, Rossetto M, Wang Y, Chen S (2015) Plant DNA barcoding: from gene to genome. Biol Rev 90(1):157–166. https://doi.org/10.1111/brv.12104
Liu J, Shi L, Han J, Li G, Lu H, Hou J, Downie SR (2014) Identification of species in the angiosperm family Apiaceae using DNA barcodes. Mol Ecol Resour 14(6):1231–1238. https://doi.org/10.1111/1755-0998.12262
Nazar N, Howard C, Slater A, Sgamma T (2022) Challenges in medicinal and aromatic plants DNA barcoding—lessons from the Lamiaceae. Plants 11(1):137. https://doi.org/10.3390/plants11010137
Nguyen Q-H, Talou T, Cerny M, Evon P, Merah O (2015) Oil and fatty acid accumulation during coriander (Coriandrum sativum L.) fruit ripening under organic cultivation. Crop J 3(4):366–369. https://doi.org/10.1016/j.cj.2015.05.002
Parveen I, Techen N, Khan IA (2019) Identification of species in the aromatic spice family Apiaceae using DNA mini-barcodes. Planta Med 85(02):139–144. https://doi.org/10.1055/a-0664-0947
Roy S, Tyagi A, Shukla V, Kumar A, Singh UM, Chaudhary LB, Nair NK (2010) Universal plant DNA barcode loci may not work in complex groups: a case study with Indian Berberis species. PLoS ONE 5(10):e13674. https://doi.org/10.1371/journal.pone.0013674
Salah HA, Temerk HA, Salah NA, Alshehri SRZ, Al- Harbi JA, Mawad AMM, Khaled KAM, Hesham AEL, Amein KA (2021) Production and optimization of xylanase and (alpha)- amylase from non-saccharomyces yeasts (Pichia membranifaciens). J Pure Appl Microbiol 15(1):452–462. https://doi.org/10.22207/JPAM.15.1.43
Sonigra P, Meena M (2021) Metabolic profile, bioactivities, and variations in the chemical constituents of essential oils of the Ferula genus (Apiaceae). Front Pharmacol 11:608649. https://doi.org/10.3389/fphar.2020.608649
Tamura K, Stecher G, Kumar S (2021) MEGA11: Molecular evolutionary genetics analysis version 11. Mol Biol Evol 38:3022–3027. https://doi.org/10.1093/molbev/msab120
Ullah R, Alqahtani AS, Noman OM, Alqahtani AM, Ibenmoussa S, Bourhia M (2020) A review on ethno-medicinal plants used in traditional medicine in the Kingdom of Saudi Arabia. Saudi J Biol Sci 27(10):2706–2718. https://doi.org/10.1016/j.sjbs.2020.06.020
Wang H, Shi LL, Zhou J, Zhu GP (2018) DNA barcoding identification of Dendrobium huoshanense and its adulterants. Zhongguo Zhong Yao Za Zhi 43(20):4055–4061. https://doi.org/10.19540/j.cnki.cjcmm.2018.0107
Whitlock BA, Hale AM, Groff PA (2010) Intraspecific inversions pose a challenge for the trnH-psbA plant DNA barcode. PLoS ONE 5(7):e11533. https://doi.org/10.1371/journal.pone.0011533
Youssef D, El-Bakatoushi R, Elframawy A, El-Sadek L, Badan GE (2023) Molecular phylogenetic study of flavonoids in medicinal plants: a case study family Apiaceae. J Plant Res 136(3):305–322. https://doi.org/10.1007/s10265-023-01442-y
Yu J, Wu X, Liu C, Newmaster S, Ragupathy S, Kress WJ (2021) Progress in the use of DNA barcodes in the identification and classification of medicinal plants. Ecotoxicol Environ Saf 208:111691. https://doi.org/10.1016/j.ecoenv.2020.111691
Acknowledgements
We would like to express our gratitude to Ain Shams University and Beni-Suef University for all the support we have received, as well as to the staff of the Molecular Cytogenetics Laboratory for creating an ideal setting in which to conduct our studies. Hadeer M. A. Mansour and Hasnaa Yehia, Teaching Assistants. The Microbiological Resources Center “Cairo Mircen” also merits recognition for doing some of the PCR experiments. Dr. Hala Zoghly deserves special recognition for her help in uploading the sequencing data to the NCBI database.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
FME and RAAY with MAA contributed to conceptualization, SAA collected, prepared, and drafted data. SAA, KAMK and MMM contributed to writing and approval of the contents. RAAY and KAMK did the work of paraphrasing and measuring the plagiarism ratio. MMM and KAMK generated the numbers and did the final revision. FME, RAAY and MAA supervised all stages of manuscript preparation. MMM and KAMK have read, reviewed, and approved the content of the final version of this review.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Abdelaziz, S.A., Khaled, K.A.M., Younis, R.A.A. et al. Comparison of four DNA barcoding loci to distinguish between some Apiaceae family species. Beni-Suef Univ J Basic Appl Sci 13, 12 (2024). https://doi.org/10.1186/s43088-023-00457-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43088-023-00457-7