
Abstract
Background
The current study aimed to investigate the oxidative stress in rat liver with diabetes mellitus (DM) as well as the protective effects of N-acetylcysteine (NAC) on irisin expression.
Methods
Twenty-eight male Wistar rats were divided into four groups, 7 rats in each group, and 30-day regimens of experimental or control groups. NAC-treated group is as follows: 100 mg/kg once daily was administered intraperitoneally (i.p.). Diabetes-induced group is as follows: single-dose intraperitoneal injection of streptozotocin (STZ) (50 mg/kg) was used to induce DM in overnight fasting Wistar rats. By determining blood glucose concentration in STZ-induced rats 72 h after injection of STZ, DM was assessed. DM + NAC group is as follows: STZ-induced DM plus NAC is described previously. On the 30th day of the experiment, liver samples were collected after fasting and anesthesia. Biochemical analyses were performed to measure total antioxidant status (TAS), total oxidant status (TOS), and malondialdehyde (MDA) levels. Each liver sample was weighed and then prepared for histopathologic evaluation by light microscopy.
Results
There was a statistically significant decrease in TAS levels and an increase in TOS and MDA levels in the DM group compared to the control group. In contrast, TOS and MDA levels were found significantly decreased, and TAS levels increased in the serum and liver tissues of the DM + NAC group compared to the DM group. Liver samples were also used for histopathological examination using hematoxylin-eosin and immunohistochemical staining. STZ-induced liver damage was detected as oxidative stress, increased irisin immunoreactivity, sinusoidal dilatation, and hepatocyte degeneration. In the DM + NAC group, it was observed that NAC significantly reduced the aforementioned histopathological changes due to STZ.
Conclusion
In the early period of diabetes, due to the antioxidant properties of irisin related to the sudden response of liver tissue to oxidative stress, it is thought that the immunoreactivity in the tissue increases in the early period. As a result, NAC in diabetic rat liver tissue was found to suppress oxidative damage and irisin immunoreactivity.
Similar content being viewed by others

Introduction
Long-term uncontrolled hyperglycemia in diabetes may cause complications by damaging organs such as kidneys, nerves, eyes, and the cardiovascular system [1, 2]. The mechanisms involved in the formation of diabetes complications are as follows: oxidative stress and increased free radicals [3]. All forms of diabetes can cause damage to the liver through oxidative stress [4].
Oxidative stress can be defined as a condition that disrupts the antioxidant defense system by increasing the production of reactive oxygen species (ROS).
By causing a toxic effect on the cell, ROS increases damage to lipids, proteins, and deoxyribonucleic acid (DNA) and may induce cell death by causing lipid peroxidation [1, 5, 6]. Regardless of the underlying etiology, the main mechanism of liver injury is oxidative stress and inflammation [7]. Malondialdehyde (MDA) is the biomarker of oxidative stress and is known to be the most frequently used biomarker [8]. The MDA production increase may cause the inactivation of many cellular proteins and receptors [1]. Irisin, a member of the myokine group, is a thermogenic protein that plays a role in energy metabolism by converting the white fat tissue to brown fat tissue [6]. Also, it is a hormone found in the skeletal muscle with autocrine, paracrine, and endocrine action [9]. Peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α) — a multi-functional regulatory transcription factor — plays a vital role in the regulation of fatty acid β-oxidation, mitochondrial biogenesis, and gluconeogenesis in the liver [10, 11]. An increase in the expression of PGC-1α, which stimulates mitochondrial biogenesis, causes an increase in irisin also known as fibronectin type 3 domain that contains 5 (FNDC5) [6]. Irisin is formed as a result of the degradation of the cell membrane protein FNDC5 and is known to be an antidiabetic hormone that plays a role in liver metabolism [10, 11].
In a previous study conducted on diabetic patients, circulating irisin level was shown to be significantly low as compared to the control group, and insulin resistance and glucose disorders were improved by FNDC5/irisin [12]. Recent studies on oxidative stress and irisin levels have reported a negative relationship between them [13]. Insulin resistance and hyperglycemia cause increased production of ROS and free radicals. In other words, oxidative stress is found to have played an essential role in the pathogenesis of diabetes-related complications [14].
Amino acid I-cysteine and N-acetylcysteine (NAC) — an acetylated precursor of glutathione (GSH) — are consumed with drug conjugation [15, 16]. NAC has an antioxidant property, and it is used as a mucolytic agent in respiratory system diseases [17]. NAC is found to have been used safely for decades in cases of paracetamol overdose. In addition, it is effective in radiocontrast nephropathy and non-paracetamol-induced acute liver failure [18]. NAC is known to regulate endothelial functions, stimulate angiogenesis, and regulate diabetes treatment [19]. In addition, it has many tasks, such as the excretion of liver toxins and providing hemostasis. NAC provides a protective effect in the liver by its antioxidant property directly or by increasing intracellular glutathione [7].
Therefore, this study aims to investigate the oxidative stress in diabetic rat liver as well as the protective effects of NAC on irisin expression.
Material and methods
Ethical statement
This study was approved by Adiyaman University Local Ethics Committee on Animal Experiments (protocol no: 2018/020). The animals used in the experimental study were obtained from the Research Center Unit of Adiyaman University. The experimental applications and maintenance were carried out there. After the experiments, the rat tissues were studied at Adiyaman University Faculty of Medicine in Histology and Embryology Department, whereas sera were studied at the Faculty of Pharmacy in the biochemistry laboratory.
Chemicals and kits
STZ was purchased from a pharmaceutical chemical company, namely Sigma Chemical Co., St. Louis, MO, USA, and NAC was purchased from Husnu Arsan pharmaceutical industry.
Irisin was purchased from another pharmaceutical company, namely irisin rabbit polyclonal H-067-17, Phoenix Pharmaceuticals, Inc., located in California, USA. Caspase-3 was purchased from Caspase-3, rabbit polyclonal IgG, Abcam, ab2302, situated in London, UK, and was measured using the immunohistochemical staining method. The highest quality solvents and chemicals available in the market were used in MDA and other measurement parameters. Total antioxidant status (TAS) and total oxidant status (TOS) activation were studied (TAS lot no: 201912, TOS lot no: 201925, Rel Assay Diagnostics, Gaziantep, Turkey).
Animals
All the experiments were carried out following the ethical standards of the institution where the study was conducted. Twenty-eight male Wistar rats weighing 220–300 g and aged 10–12 weeks purchased from the Experimental Animal Unit of Adiyaman Application and Research Center (Adiyaman, Turkey) were used in the experiments. The experimental rats were divided into four groups, each containing 7 Wistar rats. Before carrying out the investigations, all the rats were kept under the same laboratory conditions for 14 days. The laboratory conditions were as follows: 55 ± 5% relative humidity, 22 ± 3 °C temperature, 12-h day and 12-h night period, fed with standard diet (DSA Poultry, Kirikkale, Turkey), and tap water ad libitum, respectively.
Experimental design
The male Wistar rats were divided into four groups randomly. There were 7 rats in each group and 30-day regimens of experimental or control groups. Animals were untreated in the control group. The treatments applied in the experimental groups were as follows: NAC-treated group: 100 mg/kg i.p. once daily, NAC (Sigma Chemical Company, USA), was administered intraperitoneally (i.p) to animals for 30 days starting on day 1 [20] and diabetes-induced group: single-dose intraperitoneal injection of STZ (50 mg/kg B.W.) which was dissolved in buffer (pH 4.5) containing 0.1-M citrate was used to induce diabetes mellitus (DM) in overnight fasting Wistar rats [21]. By determining blood glucose concentration in STZ-induced rats 72 h after injection of STZ, diabetes was assessed. The experimental animals detected to have blood glucose levels above 250 mg/dl were used for the experimental studies [22, 23]; DM + NAC group is as follows: STZ induced DM plus NAC as described previously. The doses, application methods, and duration of treatment of STZ and antioxidant agents (NAC) were arranged according to previous studies. Blood samples were taken by cardiac puncture at the end of the study before euthanasia. Then, by performing anesthesia using ketamine overdose (Ketalar; Eczacibasi Warner-Lambert, Levent, Istanbul, Turkey), all the animals were sacrificed, and their tissue samples were obtained accordingly. Blood was collected through a microcapillary tube from all rats, and biochemical analyses were performed to measure TAS, TOS, and MDA levels. Each liver sample was weighed and then prepared for histopathologic evaluation by light microscopy.
Taking liver samples
The experimental animals were anesthetized by intraperitoneal administration of ketamine ((75 mg/kg) + xylazine (10 mg/kg)) 24 h after the last experiment. Then, the liver tissues of anesthetized animals were quickly removed and placed in a 10% formalin solution. For conducting an accurate study of TAS, TOS, and MDA parameters, some of the tissue samples were stored at −80 °C, whereas some were placed in a 10% formalin solution and evaluated for 24 h. Liver tissues were passed through a series of routine histological follow-ups and were eventually embedded in paraffin blocks. They were stored under suitable laboratory conditions for immunohistochemical and histopathological examinations.
TAS, TOS, and MDA measurements in tissue
After washing liver tissues with cold PBS buffer for tissue TAS and TOS levels, liver tissues were kept at −80 °C until analysis. Liver tissues obtained from rats were homogenized in ice-cold 0.1-M phosphate buffer with pH: 7.4 in a 1/10 ratio and were centrifuged at 1800 rpm. ELISA reader BioTek Elx800 (BioTek Instruments, USA) was used in order to provide the reading for supernatants for TAS as Trolox equivalent/L µmol at 660-nm wavelength [24] and supernatants for TOS as µmol H2O2 equiv./L at 530 nm wavelength [25].
In the experimental setup, the rat liver tissues were homogenized in the buffer solution with a homogenizer for 3 min at 16,000 rpm for MDA (Ultra Turrax type T25-B, IKA Labortechnik, Germany). Esterbauer and Cheeseman’s 1990 method was used for this purpose. MDA forms a pink-colored chromogen by reacting with thiobarbituric acid at 90–95 °C. The sample absorbance was observed to be cooled rapidly after 15 min and was read at 532 nm spectrophotometrically [26]. The obtained results were expressed in nmol/g.
Hematoxylin-eosin (H&E) staining method
Liver tissues were passed through a series of routine histological follow-ups and were eventually embedded in paraffin blocks. A total of 4–6-μm-thick sections from these blocks were prepared histopathologically after hematoxylin-eosin (H&E) staining. Using a Leica DM500 microscope (Leica DFC295), preparations were examined and photographed respectively.
Immunohistochemical staining method
A total of 4–6-μm-thick sections taken from paraffin blocks were put into polylysine slides. Initially, deparaffinized tissues were passed through graded series of alcohol and were boiled accordingly in a citrate buffer solution in order to retrieve the antigen at pH: 6 in a microwave oven (750 W) for 7 + 5 min. After boiling the tissues that were kept at room temperature for about 20 min, they were washed with phosphate-buffered saline (PBS), P4417, Sigma-Aldrich, USA) for 3 × 5 min. Then, in order to prevent endogen peroxidase activity, the tissues were incubated with a hydrogen peroxide block solution (hydrogen peroxide block, TA-125-HP, Lab Vision Corporation, USA) for 5 min. After applying Ultra V Block solution to the tissues that were washed with PBS for 5 min to prevent floor staining, primary antibodies diluted at 1/200 were incubated with irisin and caspase-3 for each section in a humid environment for 60 min. Then, after being washed with PBS for 3 × 5 min, the tissues were incubated with a secondary antibody (Biotinylated Goat anti-Polyvalent (anti-mouse/rabbit IgG), TP-125-BN, Lab Vision Corporation, USA) for 30 min at room temperature and in a humid environment. After incubating with a secondary antibody, the tissues were incubated with streptavidin peroxidase (TS-125-HR, Lab Vision Corporation, USA) for 30 min after being washed with PBS for 3 × 5 min and then taken into PBS. 3-amino-9-ethyl carbazole (AEC) Substrate and AEC Chromogen (AEC Substrate, TA-015 and HAS, AEC Chromogen, TA-002-HAC, Lab Vision Corporation, USA) solution were dripped on tissues, and the image was taken on a light microscope. After taking the image, the tissues were washed with PBS. After being passed through distilled water and PBS, the contrast-stained tissues with Mayer’s hematoxylin were closed with an appropriate closure solution (Large Volume Vision Mount, TA-125-UG, Lab Vision Corporation, USA). Using a Leica DM500 microscope (Leica DFC295), the preparations were examined and photographed respectively.
Histoscore was calculated for immunoreactivity prevalence (0.1: < 25%, 0.4: 26–50%, 0.6: 51–75%, 0.9: 76–100%) and intensity (0: non-staining, + 0.5: very less, + 1: less, + 2: median, + 3: strong) in staining. The formulation for histoscore calculation is as follows: histoscore = prevalence × intensity.
Data analysis
SPSS 15.0 program was used for the statistical analyses. The normal distributions of immune variable groups and TAS, TOS, and MDA were assessed using the Kolmogorov-Smirnov test. One-way analysis of variance (ANOVA) was performed for group comparisons of TOS, TAS, MDA, and immunity variables. For the homogeneity test of variances, Levene’s statistic was applied accordingly. In order to identify the differences between significant variable groups, Tukey’s pair-wise multiple comparison tests were conducted. The obtained results were given as mean ± standard deviation (SD). The significance level was determined to be at least 0.05 (P < 0.05).
Results
Biochemical findings
TAS, TOS, and MDA levels in liver tissue
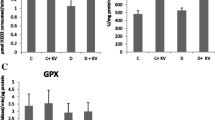
When tissue TOS and TAS levels of all groups were evaluated biochemically, their levels were observed to be similar in the control and NAC groups. There was a statistically significant decrease in TAS levels and an increase in TOS levels in the DM group compared to the control group (P < 0.05). A statistically significant increase in TAS levels and a decrease in TOS levels were observed in the DM + NAC group compared to the DM group (P < 0.05) (Fig. 1).

1 Control group; 2 NAC treated groups; 3 Diabetes-induced group; 4 DM + NAC group. TAS and TOS mean values and standard deviation rates are given. Results were given as mean ± standard deviation (n = 7 for each group)
Tissue MDA levels that were spectrophotometrically measured were similar in the control and NAC groups. There was a significant increase in MDA levels in the DM group compared to NAC and control groups. In contrast, MDA levels were found to be decreased significantly in liver tissues of the DM + NAC group when compared with the DM group (P < 0.05) (Fig. 2).

1 Control group; 2 NAC treated groups; 3 Diabetes-induced group; 4 DM + NAC group. MDA mean values and standard deviation rates are given. Results were given as mean ± standard deviation (n = 7 for each group)
Histological findings
In examining eosin and hematoxylin staining under light microscopy, liver tissues of the control group (Fig. 3a) and NAC group (Fig. 3d) were reported to have a normal histologic appearance. Significant sinusoidal dilatation (black arrow) and degeneration of hepatocytes (red arrow) were observed in the diabetic group (Fig. 3b) as compared to the control group. When compared with the diabetic group, DM + NAC (Fig. 3c) group showed a significant decrease in sinusoidal dilatation and degeneration of hepatocytes.

Representative micrographs showing the liver samples of all experimental groups. Control and NAC group (a, b) showing normal liver architecture. DM group (c) sinusoidal dilatation and hemorrhage (black arrow) and degeneration of hepatocytes (red arrow) compared to the control and NAC groups. Section from DM + NAC group showing mild sinusoidal dilatation and hemorrhage (black arrow) and degeneration of hepatocytes (red arrow) compared to DM group. ×20, H&E staining (I/R; ischemia-reperfusion)
Immunohistochemical findings
In the examination of immunohistochemical staining under light microscopy, irisin expression was detected in liver tissue hepatocytes (black arrow). Irisin expression in liver tissue was found to be similar in both control (Fig. 4a) and NAC (Fig. 4b) groups. When compared with the control group (P < 0.05), a statistically significant increase was detected in irisin immunoreactivity in the diabetic group (Fig. 4c). Irisin immunoreactivity was observed to be significantly decreased in DM + NAC (Fig. 4d) group when compared with the diabetic group (Fig. 4). In the examination of immunohistochemical staining under light microscopy, caspase-3 immunoreactivity was detected in liver tissue hepatocytes (black arrow). Caspase-3 immunoreactivity was similar in the control (Fig. 5a) and NAC (Fig. 5b) groups. It was found to be statistically significantly increased in the diabetic group (Fig. 5c) when compared with the control group (P < 0.05). A statistically significant decrease was observed in caspase-3 immunoreactivity in the DM + NAC group (P < 0.05) (Fig. 5d) when compared with diabetic group (Figs. 5 and 6).

Representative micrographs showing irisin expression for all experimental groups. Control and NAC group (a, b) showing irisin immune negativity. DM group (c) showing severe irisin expression (black arrow) compared to the control group. DM + NAC group (d) showing moderately irisin expression (black arrow) compared to control and NAC groups. ×20, immunohistochemical staining

Representative micrographs showing caspase-3 expression for all experimental groups. Control and NAC group (a, b) showing caspase-3 immune negativity. DM group (c) showing severe caspase-3 expression (black arrow) compared to the control group. DM + NAC group (d) showing moderately caspase-3 expression (black arrow) compared to control and NAC groups. ×20, immunohistochemical staining

1 Control group; 2 NAC treated groups; 3 Diabetes-induced group; 4 DM + NAC group. Irisin and caspase-3 mean values and standard deviation rates are given. Results were given as mean ± standard deviation (n = 8 for each group)
Discussion
Insulin deficiency and resistance to insulin also play a role in the formation of DM; accordingly, these can produce several changes in protein, carbohydrate, and lipid metabolism [27]. Hyperglycemia is effective in diabetes and diabetic complications. Hyperglycemia causes an increase in ROS and then causes damage to the tissues [28]. Diabetes damages the liver, primarily through oxidative stress [29]. As a result, inflammation, necrosis, fibrosis, cirrhosis, and hepatocellular carcinoma may develop [7]. It was reported that high glucose level in diabetes directly caused apoptosis and was associated with caspase-3. It was found that hyperglycemia increased caspase-3 activation by increasing cytochrome c release, thus resulting in apoptosis [30, 31]. This study investigates NAC effects on irisin immunoreactivity in diabetic rat liver tissue.
NAC has a cytoprotective effect and is a precursor of glutathione, an antioxidant. Endogenous antioxidants are depleted in cases where tissue perfusion is impaired, such as sepsis, trauma, pancreatitis, and acute myocardial infarction. NAC plays a role in the defense mechanism by removing free radicals and replacing the depleted antioxidant glutathione [32, 33]. In addition, NAC was found to have a protective role in hypoxia-related cell injury of the liver [34], ionizing radiation-induced liver failure [35], ischemic liver injury in rats [36], and toxic liver damage [37].
In a study conducted on hypothalamic obese rats by Villagarcia et al. [38], NAC significantly decreased oxidative stress in the liver and prevented inflamed-dysmetabolic liver, dyslipidemia, and the development of prediabetes. In their hepatotoxicity model with carbon tetrachloride (CCl4), Demiroren et al. [16] found that NAC provided a protective effect by decreasing oxidative stress in the liver. Mayer et al. [39] found that NAC had a protective effect against cell death due to apoptosis or oxidative stress. In the diabetes model of rats formed by Khazai et al. [40], a significant increase in TOS and MDA and decrease in TAS levels, and a significant increase in MDA level as an indicator of lipid peroxidation, were observed in liver tissue. In a study conducted on diabetic rats, Patar et al. [41] reported a significant increase in ROS, MDA, and caspase-3 levels. In another study on diabetic rats, it was found that apoptosis and lipid peroxidation increased in liver tissue [42]. In our study, we observed that NAC significantly reduced oxidative stress parameters such as TOS, MDA, and caspase-3 in the liver, strengthened the antioxidant capacity by increasing the TAS level, and also significantly improved liver damage histopathologically. In addition, we observed that NAC caused a decrease in the irisin immunoreactivity in the liver tissue by DM. Our findings related to the increased oxidative stress in liver damage due to diabetes were consistent with previous literature data, but the increased irisin immunoreactivity in tissue was noteworthy.
Irisin, a newly discovered hormone, plays a role in energy homeostasis and regulates glucose metabolism [11]. In a study, Rizk et al. [43] found that serum irisin levels were upregulated in cases diagnosed with metabolic syndrome and having elevated liver enzymes. It was also stated that the irisin clearance probably occurred in the liver. Zhang et al. [44] reported that inflammatory disorder was prominent in type 2 DM individuals with the macrovascular disease, and that irisin might be the new determinant of MDA. In a study by Moreno-Perez et al. [45], high irisin levels were found to be related to insulin resistance and nonalcoholic fatty liver disease. In the study of children with type 1 DM, Faienza et al. [46] found that a high level of irisin provided better glycemic control. In a similar studies, the increase in irisin was reported to have prevented oxidative stress by decreasing ROS production in hepatic cells and regulating inflammatory markers such as p38 MAPK, cyclooxygenase 2 (COX2), IL-6, NF-κB, and TNF. It was stated that it increases in parallel with oxidative stress [47, 48]. In their study, Liu S. et al. [49] noted that irisin might show antioxidant effects due to decreased ROS in the tissue. In another study, it was reported that exogenous irisin treatment may have an anti-apoptotic effect by reducing the expression of many apoptotic markers caused by hyperglycemia, such as caspase-3 [47]. Consistent with these findings, Bi et al. [50] reported that exogenous irisin administration alleviates damage as well as inflammatory response in the liver in ischemia-reperfusion injury. Erdogan et al. [6] found an increase in irisin immunoreactivity, which is an antioxidant in liver tissue, in case of hepatotoxicity. In addition, it was stated that irisin increases with a protective response to glycemic disorders in the early period, and the level of irisin decreases in plasma in the long term after the progression of diabetes [51]. In this study, we found that the immunoreactivity of irisin in the tissue increased due to the immediate response of the liver tissue to oxidative stress in the early stages of DM. We also found that NAC, the precursor of GSH, reduced both this oxidative damage and the immunoreactivity of irisin.
Conclusion
In this study, we examined the role of irisin in the oxidative stress caused by free radicals in liver tissue and the protective effect of NAC against it in an experimental animal model of STZ-induced DM. We found that the immunoreactivity of irisin in the tissue increased due to the immediate response of the liver tissue to oxidative stress in the early period of DM, whereas NAC, the precursor of GSH, suppressed oxidative damage and immunoreactivity of irisin.
Availability of data and materials
All data generated or analyzed during this study are included in this published article (and its supplementary information files).
Abbreviations
- NAC:
-
N-Acetylcysteine
- STZ:
-
Streptozotocin
- i.p:
-
İntraperitoneally
- TAS:
-
Total antioxidant status
- TOS:
-
Total oxidant status
- MDA:
-
Malondialdehyde
- DM:
-
Diabetes mellitus
- ROS:
-
Reactive oxygen species
- DNA:
-
Deoxyribonucleic acid
- PGC-1α:
-
Peroxisome proliferator-activated receptor gamma coactivator 1-alpha
- FNDC5:
-
Fibronectin type 3 domain that contains 5
- GSH:
-
Glutathione
- H&E:
-
Hematoxylin-eosin
- SD:
-
Standard deviation
- PBS:
-
Phosphate-buffered saline
- AEC:
-
3-Amino-9-ethyl carbazole
References
Alatawi FS, Faridi UA, Alatawi MS (2018) Effect of treatment with vitamin D plus calcium on oxidative stress in streptozotocin-induced diabetic rats. Saudi Pharm J 26(8):1208–1213. https://doi.org/10.1016/j.jsps.2018.07.012
Eva JJ, Kassab YW, Neoh CF et al (2018) Self-care and self-management among adolescent T2DM patients: a review. Front Endocrinol (Lausanne). 18(9):489. https://doi.org/10.3389/fendo.2018.00489
Darenskaya MA, Kolesnikova LI, Kolesnikov SI (2021) Oxidative stress: pathogenetic role in diabetes mellitus and its complications and therapeutic approaches to correction. Bull Exp Biol Med. 171(2):179–189. https://doi.org/10.1007/s10517-021-05191-7
Schmatz R, Perreira LB, Stefanello N et al (2012) Effects of resveratrol on biomarkers of oxidative stress and on the activity of delta aminolevulinic acid dehydratase in liver and kidney of streptozotocin-induced diabetic rats. Biochimie 94:374–383. https://doi.org/10.1016/j.biochi.2011.08.005
Celik M, Koyuncu I (2018) A comprehensive study of oxidative stress in tinnitus patients. Indian J Otolaryngol Head Neck Surg 70:521–526. https://doi.org/10.1007/s12070-018-1464-7
Erdogan MA, Yalcin A (2020) Protective effects of benfotiamine on irisin activity in methotrexate-induced liver injury in rats. Arch Med Sci 16(1):205–211. https://doi.org/10.5114/aoms.2018.80002
De Andrade KQ, Moura FA, Dos Santos JM et al (2015) Oxidative stress and inflammation in hepatic diseases: therapeutic possibilities of N-acetylcysteine. Int J Mol Sci 16(12):30269–30308. https://doi.org/10.3390/ijms161226225
Kiziler AR, Aydemir B, Gulyasar T et al (2008) Relationships among iron, protein oxidation and lipid peroxidation levels in rats with alcohol-induced acute pancreatitis. Biol Trace Elem Res 124(2):135–143. https://doi.org/10.1007/s12011-008-8127-6
Irving BA, Still CD, Argyropoulos G (2014) Does irisin have a brite future as a therapeutic agent in humans? Curr Obes Rep 3(2):235–241. https://doi.org/10.1007/s13679-014-0091-1
Martinez Munoz IY, Camarillo Romero EdS, Garduno Garcia JdJ (2018) Irisin a novel metabolic biomarker: present knowledge and future directions. Int J Endocrinol 2018:1–8. https://doi.org/10.1155/2018/7816806
Perakakis N, Triantafyllou GA, Fernández-Real JM et al (2017) Physiology and role of irisin in glucose homeostasis. Nat Rev Endocrinol 13(6):324–337. https://doi.org/10.1038/nrendo.2016.221
Kheiripour N, Karimi J, Khodadadi I et al (2019) Hepatoprotective effects of silymarin on liver injury via irisin upregulation and oxidative stress reduction in rats with type 2 diabetes. Iran J Med Sci 44(2):108–117. https://doi.org/10.1530/eje-14-0204
Zhu D, Wang H, Zhang J et al (2015) Irisin improves endothelial function in type 2 diabetes through reducing oxidative/nitrative stresses. J Mol Cell Cardiol 87:138–147. https://doi.org/10.1016/j.yjmcc.2015.07.015
Ghadermazi R, Khoshjou F, Hossini Zijoud SM et al (2018) Hepatoprotective effect of tempol on oxidative toxic stress in STZ-induced diabetic rats. Toxin reviews 37(1):82–86. https://doi.org/10.1080/15569543.2017.1313277
Rushworth GF, Megson I (2014) Existing and potential therapeutic uses for N-acetylcysteine: the need for conversion to intracellular glutathione for antioxidant benefits. Pharmacol Ther 141(2):150–159. https://doi.org/10.1016/j.pharmthera.2013.09.006
Demiroren K, Basunlu MT, Erten R et al (2018) A comparison of the effects of thymoquinone, silymarin and N-acetylcysteine in an experimental hepatotoxicity. Biomed Pharmacother 106:1705–1712. https://doi.org/10.1016/j.biopha.2018.07.125
Santus P, Corsico A, Solidoro P et al (2014) Oxidative stress and respiratory system: pharmacological and clinical reappraisal of N-acetylcysteine. COPD 11(6):705–717. https://doi.org/10.3109/15412555.2014.898040
Gu L, Shen Z, Li YL et al (2018) Oxymatrine causes hepatotoxicity by promoting the phosphorylation of JNK and induction of endoplasmic reticulum stress mediated by ROS in LO2 cells. Mol Cells 41(5):401–412. https://doi.org/10.14348/molcells.2018.2180
Zayed MA, Wei X, Park KM et al (2017) N-acetylcysteine accelerates amputation stump healing in the setting of diabetes. FASEB J 31(6):2686–2695. https://doi.org/10.1096/fj.201601348R
Rana SV, Attri S, Vaiphei K (2006) Role of N-acetylcysteine in rifampicin-induced hepatic injury of young rats. World J Gastroenterol 12(2):287–291. https://doi.org/10.3748/wjg.v12.i2.287
Baydas G, Reiter RJ, Yasar A et al (2003) Melatonin reduces glial reactivity in the hippocampus, cortex and cerebellum of streptozotocin-induced diabetic rats. Free Radical Biol Med 35:797–804. https://doi.org/10.1016/s0891-5849(03)00408-8
Pavana P, Sethupathy S, Santha K et al (2009) Effects of Tephrosia purpurea aqueous seed extract on blood glucose and antioxidant enzyme activities in streptozotocin induced diabetic rats. Afr J Tradit Complement Altern Med 6(1):78–86. https://doi.org/10.4314/ajtcam.v6i1.57077
Gozel N, Genc F, Ozdemir FA et al (2018) Effects of vitamin D on matrix metallloproteinase 9 and apoptosis in experimental diabetic rat kidney tissue. FEB 27(5 A):3766–3775
Erel O (2004) A novel automated direct measurement method for total antioxidant capacity using a new generation, more stable ABTS radical cation. Clin Biochem 37(4):277–285. https://doi.org/10.1016/j.clinbiochem.2003.11.015
Erel O (2005) A new automated colorimetric method for measuring total oxidant status. Clin Biochem 38(12):1103–1111. https://doi.org/10.1016/j.clinbiochem.2005.08.008
Esterbauer H, Cheeseman KH (1990) Determination of aldehydic lipid peroxidation products: malonaldehyde and 4-hydroxynonenal. Methods Enzymol 186:407–421. https://doi.org/10.1016/0076-6879(90)86134-h
Park SY, Gautier JF, Chon S (2021) Assessment of insulin secretion and insulin resistance in human. Diabetes Metab J. 45(5):641–654. https://doi.org/10.4093/dmj.2021.0220
Volpe CMO, Villar-Delfino PH, Dos Anjos PMF et al (2018) Cellular death, reactive oxygen species (ROS) and diabetic complications. Cell Death Dis 9(2):119. https://doi.org/10.1038/s41419-017-0135-z
Wang Q, Wei S, Zhou H et al (2019) Hyperglycemia exacerbates acetaminophen-induced acute liver injury by promoting liver-resident macrophage proinflammatory response via AMPK/PI3K/AKT-mediated oxidative stress. Cell Death Discov 5(1):1–12. https://doi.org/10.1038/s41420-019-0198-y
Cai L, Li W, Wang G et al (2002) Hyperglycemia-induced apoptosis in mouse myocardium: mitochondrial cytochrome C–mediated caspase-3 activation pathway. Diabetes 51(6):1938–1948. https://doi.org/10.2337/diabetes.51.6.1938
Francés DE, Ronco MT, Monti JA et al (2010) Hyperglycemia induces apoptosis in rat liver through the increase of hydroxyl radical: new insights into the insulin effect. J Endocrinol 205(2):187. https://doi.org/10.1677/JOE-09-0462
Pedre B, Barayeu U, Ezeriņa D, Dick TP (2021) The mechanism of action of N-acetylcysteine (NAC): the emerging role of H2S and sulfane sulfur species. Pharmacol Ther. 228:107916. https://doi.org/10.1016/j.pharmthera.2021.10791
Kavala A, Kuserli Y, Turkyilmaz S (2019) Effect of N-acetylcysteine on intimal hyperplasia and endothelial proliferation in rabbit carotid artery anastomosis. Arch Med Sci 15(6):1576–1581. https://doi.org/10.5114/aoms.2018.77769
Heil J, Schultze D, Schemmer P et al (2018) N-acetylcysteine protects hepatocytes from hypoxia-related cell injury. Clin Exp Hepatol 4(4):260–266. https://doi.org/10.5114/ceh.2018.80128
Mansour HH, Hafez HF, Fahmy NM et al (2008) Protective effect of N-acetylcysteine against radiation induced DNA damage and hepatic toxicity in rats. Biochem Pharmacol 75(3):773–780. https://doi.org/10.1016/j.bcp.2007.09.018
Kalimeris K, Briassoulis P, Ntzouvani A et al (2016) N-acetylcysteine ameliorates liver injury in a rat model of intestinal ischemia reperfusion. J Surg Res 206(2):263–272. https://doi.org/10.1016/j.jss.2016.08.049
Mendonça MCP, Ferreira LB, Rizoli C et al (2019) N-acetylcysteine reverses silver nanoparticle intoxication in rats. Nanotoxicology 13(3):326–338. https://doi.org/10.1080/17435390.2018.1544302
Villagarcía HG, Castro MC, Arbelaez LG et al (2018) N-acetyl-l-cysteine treatment efficiently prevented pre-diabetes and inflamed-dysmetabolic liver development in hypothalamic obese rats. Life sci 199:88–95. https://doi.org/10.1016/j.lfs.2018.03.008
Mayer M, Noble M (1994) N-acetyl-L-cysteine is a pluripotent protector against cell death and enhancer of trophic factor-mediated cell survival in vitro. Proc Natl Acad Sci 91(16):7496–7500. https://doi.org/10.1073/pnas.91.16.7496
Khazaei M, Karimi J, Sheikh N et al (2016) Effects of resveratrol on receptor for advanced glycation end products (RAGE) expression and oxidative stress in the liver of rats with type 2 diabetes. Phytother Res 30(1):66–71. https://doi.org/10.1002/ptr.5501
Patar AK, Sharma A, Syiem D et al (2018) Chlorophyllin supplementation modulates hyperglycemia-induced oxidative stress and apoptosis in liver of streptozotocin-administered mice. BioFactors 44(5):418–430. https://doi.org/10.1002/biof.1438
Tuncdemir M, Erturkuner S, Ozcelik D (2017) Investigation of lipid peroxidation and antiapoptotic effects of zinc aganist liver damage in diabetic rats. Hum Exp Toxicol 36(8):813–822. https://doi.org/10.1177/0960327116666619
Rizk FH, Elshweikh SA, Abd El-Naby AY (2016) Irisin levels in relation to metabolic and liver functions in Egyptian patients with metabolic syndrome. Can J Physiol Pharmacol 94(4):359–362. https://doi.org/10.1139/cjpp-2015-0371
Zhang M, Chen P, Chen S et al (2014) The association of new inflammatory markers with type 2 diabetes mellitus and macrovascular complications: a preliminary study. Eur Rev Med Pharmacol Sci 18(11):1567–1572
Moreno-Perez O, Reyes-Garcia R, Muñoz-Torres M et al (2018) High irisin levels in nondiabetic HIV-infected males are associated with insulin resistance, nonalcoholic fatty liver disease, and subclinical atherosclerosis. Clin Endocrinol 89(4):414–423. https://doi.org/10.1111/cen.13800
Faienza MF, Brunetti G, Sanesi L et al (2018) High irisin levels are associated with better glycemic control and bone health in children with type 1 diabetes. Diabetes Res Clin Pract 141:10–17. https://doi.org/10.1016/j.diabres.2018.03.046
Park MJ, Kim DI, Choi JH et al (2015) New role of irisin in hepatocytes: the protective effect of hepatic steatosis in vitro. Cell Signal 27(9):1831–1839. https://doi.org/10.1016/j.cellsig.2015.04.010
Batirel S, Bozaykut P, Altundag EM et al (2014) The effect of irisin on antioxidant system in liver. Free Radic Biol Med 75:16. https://doi.org/10.1016/j.freeradbiomed.2014.10.592
Liu S, Du F, Li X et al (2017) Effects and underlying mechanisms of irisin on the proliferation and apoptosis of pancreatic β cells. PloS one 12(4). https://doi.org/10.1371/journal.pone.0175498
Bi J, Zhang J, Ren Y et al (2019) Irisin alleviates liver ischemia-reperfusion injury by inhibiting excessive mitochondrial fission, promoting mitochondrial biogenesis and decreasing oxidative stress. Redox Biol 20:296–306. https://doi.org/10.1016/j.redox.2018.10.019
Park K, Ahn CW, Park JS et al (2020) Circulating myokine levels in different stages of glucose intolerance. Medicine 99(8):e19235. https://doi.org/10.1097/MD.0000000000019235
Acknowledgements
None.
Funding
The authors declare that no funds, grants, or other support were received during the preparation of this manuscript.
Author information
Authors and Affiliations
Contributions
MME, MAE, and SK conceived and designed research. AY and AT conducted experiments and contributed analytical tools. MME, MAE, EYA, and SK analyzed data and wrote the manuscript. All authors read and approved the manuscript. The authors declare that all data were generated in-house, and that no paper mill was used.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This study was performed in line with the principles of the Declaration of Helsinki. This study was approved by Adiyaman University Local Ethics Committee on Animal Experiments (Protocol No: 2018/020).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Erdogan, M., Erdogan, M., Koc, S. et al. Hepatoprotective effects of N-acetylcysteine on liver injury by irisin upregulation and oxidative stress reduction in diabetic rats. Egypt Liver Journal 13, 33 (2023). https://doi.org/10.1186/s43066-023-00271-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43066-023-00271-x