
Abstract
Background
The histone deacetylase (HDAC) family plays a significant role in a variety of biological processes related to cancer, such as metabolism and cancer progression, and recognized as a target of anti-cancer drugs. Nevertheless, there has been limited systematic research available regarding HDAC family in human cancers using multi-omics approaches.
Objective
To evaluate the prognostic implications and immunological features of the HDAC family across multiple cancer types through comprehensive multi-omics analysis.
Methods
We studied the associations of HDAC activity and a variety of factors, encompassing immune checkpoint genes, the tumor microenvironment (TME), instability of microsatellites (MSI), inherited mutations, variation in copy number, tumor mutation burden (TMB), and sensitivity to drugs in a variety of cancer types. Moreover, we analyzed the link between the degree of HDAC activity and the effectiveness of immunotherapy in several cohorts, providing significant details about the possible impact of HDACs on immunotherapeutic responses. Furthermore, we explored potential signaling pathways of HDACs in bladder cancer (BC) using gene set enrichment analysis (GSEA). Immune infiltration analysis in bladder cancer was performed based on HDACs expression, copy number or somatic mutations. Furthermore, the Human Protein Atlas (HPA) dataset has been used to validate the expression of the HDACs protein in BC.
Results
The HDACs expression exhibits significant heterogeneity across different cancer types and is significantly correlated with patient prognosis, clinical traits, mutations, TME, TMB, MSI, immune checkpoint genes, and objective responses in immunotherapy. The drug sensitivity of cancers was found to be correlated with the gene expression of specific HDAC familial members. In BC, GSEA demonstrates enrichment of multiple immune-related functions and pathways; moreover, there are significant associations between genomic variations in HDACs and the degree of typical immunological cell recruitment. Furthermore, findings from differential expression analysis in BC are validated by protein expression analysis obtained from the HPA database.
Conclusions
These findings reveal the significance of HDAC family in survival prognosis and treatment guidance of human cancers, and some of them have the potential to be used as immunotherapy biomarkers.
Similar content being viewed by others

Introduction
The GLOBOCAN 2020 estimates highlight cancer as a significant global public health threat, with approximately 19.3 million individuals diagnosed with new cases of cancer and nearly 10 million losing their lives due to this disease [1]. The World Health Organization (WHO) forecasts that over the following two decades, the incidence of cancer globally may rise by 60% [2]. This will increasingly burden the development of the world economy. Significant progress has been made in the detection, therapy, and management of a wide range of malignancies [3, 4]. The prognosis of cancer patients has significantly increased due to several new therapies, including immune checkpoint inhibitor therapy and personalized therapy [5, 6]. Thus, the search for effective tumor biomarkers can further promote the development of tumor therapy. Gene Expression Omnibus (GEO) and the Cancer Genome Atlas (TCGA) are substantial freely available databases that provide massive, high-throughput, multi-omics data for a wide range of cancers. These tools have significance in ensuring multiple cancers research in various omics domains executes successfully [7, 8].
The 18 members of the extensive HDAC family are categorized into classes I to IV based on their gene homology. Eleven genes of the HDAC family, known as “classics” belong to classes I, II, and IV, while the remaining class III members are known as sirtuins [9]. HDACs encoded functional proteins participate in various physiological processes, including gene expression, cellular growth, and apoptosis [10]. Previous studies have identified that aberrant expression of HDACs has been linked to diseases, such as neurodegenerative and genetic diseases [11, 12], as well as cancer [13]. Sirtuins have been previously discussed in other studies [14, 15], and won’t be covered here. Although some HDACs are aberrantly expressed in cancerous tissues [16], it is still unknown how abnormal HDAC expression leads to the pathology of tumors.
The HDAC family has not yet been thoroughly studied in human cancers. The HDAC gene members have been thoroughly studied in the present study using a range of publically available resources, such as TCGA, GEO, and other repositories. We studied the features of the tumor microenvironment, expression patterns, genetic variations, prognostic implications, and immune system infiltration in a variety of tumors. The study also analyzed how HDACs affected the responses to immunotherapy and chemotherapy, clarified the intricate biological functions of HDACs, and provide significant new data for future studies into identification, prediction, and personalized therapies for all cancer cases comprising the HDAC gene.
Materials and methods
Collecting and evaluating data
The UCSC Xena database provided the details on the expression of genes, the clinical features, rates of survival, immunophenotypic analysis, and stem index evaluation (stemness score from DNA methylation/RNA expression) of HDACs in 33 distinct tumor types. This information is accessible at https://xenabrowser.net/datapages. The mutations were downloaded by cBioPortal database, which may be accessed at http://www.cbioportal.org. We also performed external data validation with several immunotherapy datasets, such as GSE111636, GSE78220, GSE176307, GSE67501 and IMvigor210.
Furthermore, our ongoing phase II clinical trial (TRUCE-01, NCT04730219) is investigating the combination of tislelizumab and low-dose nab-paclitaxel treating bladder tumors with muscular invasion. A group of 29 patients, comprising 15 samples of tumors at baseline and 15 paired tumor samples collected after immunotherapy, provided us with mRNA sequencing data and medical details. Studying variations in HDAC activity, and evaluate any statistical differences by applying the paired Wilcoxon test.
As defined by surgical pathologists (RC-PLND or maximal TURBT), individuals undergoing immunotherapy are deemed to show a complete or partial response (CR/PR). Conversely, individuals with progressing or stable illness (SD/PD) were classified as non-responders.
Exploring the HDACs expression trends in different forms of tumors
The Strawberry Perl algorithm, which can be accessed at http://strawberryperl.com, has been used to extract and compile the HDAC gene activity data. The Wilcoxon approach was utilized to assess expression variances across HDACs in a variety of cancers. A total of 33 distinct cancer types participated in the study: ACC, BLCA, BRCA, CESC, CHOL, COAD, DLBC, ESCA, GBM, HNSC, KICH, KIRC, KIRP, LAML, LGG, LIHC, LUAD, LUSC, MESO, OV, PAAD, PCPG, PRAD, READ, SARC, SKCM, STAD, TGCT, THCA, THYM, UCEC, UCS, and UVM. Five distinct healthy samples served as standards for each tumor type to examine the 18 cancer forms. Using the 'corrupt' R package, also executed an association study on the HADC members in each matched pair. The amino acid composition of the HDAC family was confirmed by immunohistochemistry data retrieved by using Human Protein Atlas database (HPA), accessible at https://www.proteinatlas.org.
Clinical prognostic significance of HDACs
Using the average level of HDAC activity, individuals were divided into 2 categories including ones with significant expression versus those with little expression rates. Using Kaplan–Meier and Cox proportional risk regression models using 33 TCGA datasets, the association between HDAC expression and different survival rates in various cancers was studied. The survival rates were included overall survival (OS), disease-specific survival (DSS), recurrence-free survival (RFS), and progression-free survival (PFS), which were all previously defined in literature [17, 18]. The R-packages "survminer" and "survivor" has been employed to generate survival curves. The "survivor" and "forestplot" R-packages are utilized for constructing forest plots. The research employed differential analysis to identify variations in HDAC production at various stages of TNM phases.
Mutation profiles analysis
The TCGA database was used to evaluate the frequency of alterations, types of mutations, variations in copy numbers (CNVs), and single nucleotide variants (SNVs) across a variety of cancers using the cBio Cancer Genomics Portal (cBioPortal), available at http://www.cbioportal.org [19]. Using the cBioPortal and Tumor Immune Estimate Resource (TIMER) databases available at http://cistrome.dfci.harvard.edu/TIMER, predicts the consequences of HDACs expressing variations, therapeutic prognosis, and cell immune infiltration.
Association analyses of immunological variants, tumor microenvironment, stemness indices, and HDAC activity
The expression levels of HDACs were initially assessed across six distinct immunophenotypes: C1 (wound healing), C2 (INF-r dominance), C3 (inflammation), and C4 (lymphocytic). The purpose of the study was to find out the number of various immune cell types in the tumor environment [20], and TIMER dataset (http://cistrome.dfci.harvard.edu/TIMER/) was utilized. The correlation between HDACs and stromal scores (stromal cell abundance) and immune-mediated scores (immune cell proliferation) in pan-cancerous tissues was studied using the R "estimate" modules [21]. To determine whether HDAC activity and cancer stemness associations, including RNA stemness score (RNAss) and DNA stemness score (DNAss), the Spearman correlation analysis was performed.
Correlational analyses of expression of HDACs and immune-mediated checkpoints, TMB and MSI
The number of somatic mutations in tumor tissue is indicated by TMB [22], and MSI reflects the instability of genetic mutations [23], both of which are thought to largely influence tumorigenesis and progression [24]. The R program PreMSIm was implemented to assess 33 distinct cancers and calculated the MSI in patterns of gene expression [25]. The radar map has been generated by the “Fmsb” R package, while the R "pheatmap" tool was employed to show the co-expression analysis of 47 frequently reported immune-mediated checkpoints as heatmaps.
Evaluation of therapeutic sensitivity
Utilizing the National Cancer Institute (NCI)-60 server, accessed via CellMiner website at https://discover.nci.nih.gov/cellminer, the association across HDAC degree of expression and the response to treatment was studied [26].
Investigated the potential predictive capacity of HDACs expression in the context of immunotherapy
Relevant data on HDACs expression patterns and clinical information were collected from six distinct datasets, namely IMvigor210, GSE176307, GSE111636, GSE78220, GSE67501, and our TRUCE-01. Understanding the possible prognostic significance of HDAC activity related to immunotherapy was the purpose of this research. Individuals undergoing immunotherapy were separated into respondents and non-responders based on the treatment outcomes.
Assessment of Gene Set Enrichment Analysis (GSEA)
The Kyoto Encyclopedia of Genes and Genomes (KEGG) genome sets were used to conduct Gene Set Enrichment Analysis (GSEA) on bladder cancer cells to identify any prospective physiological roles for HDACs.
Association analyses between HDACs expression and immune subtype, clinical traits, stemness score, TMB, MSI, or tumor microenvironment in bladder cancer
A differential study has been conducted to investigate the correlation between the expression of HDACs and the immunologic subtype and also the clinical characteristics of bladder cancer (BC). Spearman correlation test was adopted to examine the links between HDAC activity and immunologic scores, stromal scores, tumor mutant burden (TMB), RNA methylation levels (RNAss), DNA methylation levels (DNAss), and microsatellite instability (MSI).
Immune infiltration study of HDACs expression in BC
TIMER dataset was utilized for studying the association of HDAC activity and found six major immune cells in BLCA comprising B cells, CD8+ T cells, CD4+T cells, macrophages, neutrophils, and dendritic cells (DCs). Moreover, the association among the degree of genetic alterations, copy number variations, and infiltrating immune cell concentrations has been examined.
Statistical analysis
Employing R (version 4.1.2), different R packages were used for analyzing the data gathered, including limma, ggpubr, pheatmap, ggplot2, corrplot, survival, survminer, reshape2, estimate and impute, etc. For example, box-line and heatmaps were designed using “ggpubr” and “pheatmap”, respectively. The Wilcoxon signed ranks test served for comparing two distinct groups, and over two distinct groups have been compared by the Kruskal–Wallis test. An analysis of correlations was conducted using Spearman’s correlation coefficient test. The rate of survival of the high-expression and low-expression cohorts was assessed by performing a K-M curve analysis and significant level was identified by the log-rank test. A univariate Cox regression study has been assessed to predict the potential value of HDAC genes. *, **, and *** indicate P values below 0.05, under 0.01, and less than 0.001, respectively. A probability threshold of P < 0.05 was applied to ascertain the outcomes.
Results
Expression of HDACs in pan-cancer
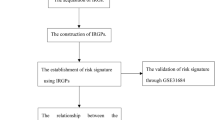
A comprehensive visual representation of the research process can be found in Fig. 1. Leveraging data from the TCGA repository, we investigated the expression levels of the HDAC family in tumor and normal samples across a variety of malignancies. Firstly, our results determined that most of the 11 members of HDACs are highly expressed in pan-cancer, especially HDAC1, while low expression was observed for HDAC9 (Fig. 2A). And, expression of each member in the 33 cancer tissues illustrated separately and ranked them from higher- lowest (Additional file 1: Figure S1). Then, results shown across 18 distinct cancer forms, there had been significant variations among HDAC family members. For example, HDAC7 was the most highly expressed in CHOL and HDAC11 was the least expressed in GBM (Fig. 2B). The level of expression of 11 HDAC family members was then found to be highly associated with one another in all types of cancers using Spearman’s correlation analysis. We learned that HDAC3 and HDAC1, HDAC6 and HDAC5 were the two most significantly positively correlated gene pairs (correlation coefficient = 0.33); HDAC5 and HDAC1 were the two most negatively correlated genes (correlation coefficient = -0.21) (Fig. 2C). Weak correlations between them can be seen, and these results suggest intrinsic differences in HDAC expression among different HDAC family members or different cancer types. Thus, a specific HDAC gene may exhibit either oncogenic or anti-oncogenic properties in different cancer types, highlighting the necessity for individual in-depth study of each HDAC family member.

Diagram showing the study’s flow regarding HDAC family genes and human cancer

Differential analysis of gene expression in the HDAC family. (A) A box plot illustrating the levels of transcriptional expression for HDACs. (B) Based on TCGA data, this heatmap illustrates the variations in HDAC activity between nearby healthy tissues and cancerous cells across a variety of cancer types. (C) Correlation coefficients by co-expression analysis between each two HDAC family genes
The levels of mRNA expression of HDACs are compared between cancer tissues and typical tissues (Additional file 2: Figure S2). Most HDAC-related genes have been discovered to be significantly expressed in a variety of tumor types, for instance HDAC1 expression was only decreased in KICH, KIRC, and KIRP; HDAC3 showed low expression in KICH, THCA; HDAC8 was down-regulated only in KIRC; and HDAC10 exhibited low expression only in KICH. However, we also found a certain degree of intra- and inter-cancer heterogeneity in the expression levels of certain HDACs, such as HDAC4 was overexpressed only in CHOL and LIHC, HDAC2 was down-regulated only in KICH and KIRC, which was in contrast to other tumor types.
Analysis of therapeutic correlation in pan-cancer
Using univariate Cox regression analysis, we studied the prognostic value of HDAC levels across different forms of cancer. As indicated in Fig. 3, the expression of some HDAC genes was related with OS, DFS, PFS and DSS in various types of cancer. The prognostic impact of HDACs exhibited some degree of variability in different cancer types. The patients were divided into groups with low or high expression following the average expression of gene values. Subsequently, we performed Kaplan–Meier analysis for the HDAC1-11 genes to assess OS, DFS, DSS, and PFS (Additional file 3: Table S1; Additional file 4: Additional Data1). Among these patients, high HDAC1 activity has been related to an unfavorable outcome in KIRC sufferers (Fig. 4A) and LGG (Fig. 4B). Conversely THYM and KIRC individuals showed less favorable results when their HDAC2 expression was decreased (Fig. 4C, D). In contrast, those having significant levels of HDAC3 in SKCM and THCA had a negative prediction (Fig. 4E and F, respectively). Besides, HDAC4 (Fig. 4G) and HDAC10 (Fig. 4K) are high-risk genes in patients with ACC; HDAC9 plays a high-risk role in THYM (Fig. 4J). Contrarily, increased expression was observed for HDAC7 in BLCA (Fig. 4H) and SKCM (P < 0.001; Fig. 4I) patients had a favorable prognosis. Similarly, the higher levels of HDAC11 was related to better prognostic results in KIRP (Fig. 4L). Significant correlation was observed between the TNM staging and the expression of HDAC family. Specifically, in KICH and TGCT, the expression level of HDAC3 showed an increase as the stage progressed (Fig. 5A). Conversely, the levels of HDAC11 expression in stage I versus stage II/III/IV were found to be the highest in KIRC and KIRP, gradually decreasing as the stage progressed (Fig. 5B). The supplementary findings of other family genes are represented in Additional file 5: Figure S3.

The forest graphs show the association between the activity of HDAC and OS (A), PFS (B), DFS (C), and DSS (D) by the univariate COX method in pan-cancer

K-M survival curves highlight the link between overall survival and the HDAC expression variances of both low and high expression cohorts. HDAC1 in KIRC (A) and in LGG (B), HDAC2 in KIRC (C) and in THYM (D), HDAC3 in THCA (E) and in SKCM (F), HDAC4 in ACC (G), HDAC7 in KIRC (H) and in SKCM (I), HDAC9 in THYM (J), HDAC10 in ACC (K), HDAC11 in KIRP (L)

The association of HDACs expression and clinicopathological stage in various cancer types. (A, B) Boxplot graph indicate the characteristics of HDAC11 and HDAC3 in relation to clinicopathological staging, respectively
Genetic alterations of HDACs in pan-cancer
We examined the situation of genetic variations in HDAC using the cBioPortal database. The HDAC family exhibited changes in 27.32% of the total 2683 cases across various tumor types, as observed from the ICGC-TCGA dataset (Fig. 6A). According to Fig. 6B, the occurrence rates of genetic mutations in HDAC varied from 1.5% to 10%. Notably, HDAC6 and HDAC9 exhibited higher mutation frequencies, with 8% and 10%, respectively, compared to other members. Amplification was the most frequent mutation type across most members in pan-cancer patients. With a frequency of mutations over 40%, the variant rate of HDACs was comparatively high in the following tumors: endometrial, head and neck, soft tissue sarcoma, lung, ovarian, esophagogastric, and bladder. In terms of amplification mutations, the incidence was higher in endometrial cancer, breast cancer, esophagogastric cancer, lung cancer, melanoma, cervical cancer (Fig. 6C). Furthermore, the more intuitive percentage of genetic changes of each HDAC family member in pan-cancer and the degree of genetic changes for each member in various cancers are detailed in Additional file 6: Figure S4. With a p-value of 0.082, the KM plotter results revealed that there was no significant difference in the OS between the groups with or without the HDAC familial gene substitution (Fig. 6D).

Alterations of the HDAC family in pan-cancer. (A) The HDACs altered in 27.32% of 2683 cases. (B, C) HDACs variant types and frequency in pan-cancer. (D) Analysis of overall survival in patients with HDACs mutation status using the cBioPortal database
In terms of CNVs, shallow deletions and gain were more prominent than deep deletion or amplification. Moreover, a statistically robust rise in the expression levels of most HDACs was observed with the rise in copy number variation (CNV). This finding aligns with the comprehensive analysis of whole genome data conducted by the ICGC/TCGA pan-cancer consortium in 2020 (Fig. 7). The study of the pan-cancer findings retrieved from the ICGC/TCGA, we also determined that the non-mutated regions accounted for a larger proportion compared to those with missense, truncating, multiple, inframe, or splice mutations. Additionally, significant differences in expression levels were observed for HDAC3/9 genes among different genomic variant types, while no statistical differences were found for other members (Additional file 7: Figure S5). These data sets demonstrated that the upregulation of HDAC mRNA in pan-cancer may mostly be attributed to an increase in the number of HDAC genomic copies.

Correlation analysis between the expression of each HDAC gene and its copy number (A-K)
Interrelation between HDAC expression and immune subtypes, tumor microenvironment, and stemness indices
To examine the correlation involving tumor immune cell infiltration and HDACs, we delved into the association between immunological cells and HDACs. A previous analysis of human malignancies identified six distinct immune subtypes: C1 (wound healing), C2 (INF-r dominant), C3 (inflammation), C4 (lymphocyte failure), C5 (Immune quiet), and C6 (TGF-β predominance), and influencing tumor progression and development [27]. It has been demonstrated that patients belonging to immune subtypes C3 and C5 exhibit significantly higher survival rates compared to those with different subtypes. However, patients classified as C4 and C6 subtypes have the lowest survival rates [27]. With a p-value less than 0.001, this study found a strong association between HDAC expression and each of the six immunological subtypes (Fig. 8A). Further analysis reveals that high expression of HDAC4/5/6/9/11 was closely correlated with C5 subtype, suggesting that these members may serve as a cancer-suppressing effect. In contrast, HDAC7 appears to function as a pro-oncogene in the C6 subtype compared to other subtypes. Similarly, high expression of HDAC1/2/3 was more relevant in C1, C2, and C6 infiltrates compared to other subtypes, illustrating that these members have a tumor-promoting role, and predicting poorer survival in these patient categories. Several studies have shown that the presence of lymphocytes infiltrating the TME is a significant indicator in predicting both tumor sentinel node status and survival duration [28]. The association between HDACs and CD8 + T cell infiltration levels determined by seven distinct algorithms has been studied using TIMER. The level of expression of HDACs proved to be positively correlated with CD8 + T cell recruitment in multiple tumor types, as depicted in Fig. 8B. Estimate method was used to examine the correlation between HDAC expression levels and stromal and immune scores in tumor samples. Most notably, the results of immune Score and stromalScore showed almost positive correlation with the expression of both HDAC7 and HDAC9 (Fig. 8C and D). Specifically, HDAC1 exhibited a strong positive correlation with the immuneScore of UVM and LGG, and HDAC4 demonstrated a similar correlation with DLBC. In the case of TGCT, the immune score had a significant unfavorable tandem with both HDAC2 and HDAC11. Regarding StromalScore, in GBM and SARC, most tumors were negatively correlated except for HDAC7, which showed a remarkably positive associated with StromalScore. Moreover, HDAC6 and HDAC10 were consistently negatively related to stromal core in almost the tumors. The association between the expression of genes and tumor stemness (RNAss and DNAss) was determined using Spearman correlation analysis (Fig. 8E and F). For RNAss, we illustrated that a vigorously positive association exist between HDAC1, HDAC2, HDAC3 and most tumors. Meanwhile, an appreciably negative correlation was seen between HDAC1 and KIRC, LGG, and PCPG. Particularly, a positive correlation was observed in HDAC1, HDAC2, HDAC3, HDAC7 and HDAC10 in the THYM. We also identified a significant negative interrelation between THYM and HDAC1/2/3/7/10 in DNAss.

Outcomes of correlation analysis of HDACs with immune subtypes, stemness index, and microenvironmental score. (A) Transcript levels of HDACs expression in multiple cancer types in C1-C6 immune subtypes. (B) Applying seven different algorithms, we examined the probable link between HDACs expression and the CD8+ T-cells infiltration. (C, D) A bubble plot illustrating the association between immunology value and stemness index with the proliferation of HDACs mRNA. (E, F) The association between stemness indices and HDAC gene expression across different cancer types is displayed by a bubble graph
Association analyses of HDACs with TMB, MSI and immune checkpoints
Given the widespread acceptance of immuno-surveillance in the prognosis of different tumor forms [29]. We studied the association between HDACs expression and 47 immune-mediated checkpoints that are frequently observed and are strongly linked to the manifestation of immunotherapy (Additional file 8: Figure S6), for instance, CTLA-4, PDL-1, and PD-1 (Fig. 9A). For instance, in LIHC, the expression of HDAC1-5 and HDAC7-9 was positively correlated with the mRNA levels of PD-L1, PD-1 and CTLA-4. HDAC1/3/8 was also identified to be positively associated with this gene expression in UVM. These results demonstrated that overexpression of HDACs may mediate immune evasion in some tumors. TMB and MSI are widely recognized as significant indicators of tumor development and advancement [24]. Additionally, investigations were conducted to explore the associations between HDAC expression, TMB, and MSI. A negative correlation was observed between HDAC1-4, HDAC6, HDAC7, and HDAC10 and TMB in THYM, whereas there was a positive association between TMB and HDAC11 in THYM, ESCA, and KIRP. It was observed that HDAC1-4, HDAC6, HDAC7, and HDAC10 are adversely associated to the TMB in THYM, while HDAC11 is positively correlated with the TMB in THYM, ESCA, and KIRP. HDAC4 was negatively associated with TMB in BLCA, BRCA, DLBC, etc., and positively correlated in ACC only (Fig. 9B, Additional file 9: Figure S7). Furthermore, HDAC1 showed an adverse correlation with MSI in CESC and a significant association with MSI in COAD, ESCA, LAML, etc. Moreover, there is merely an association between HDAC9 activity and MSI in MESO, and an unfavorable association with MSI in DLBC, ESCA, HNSC, etc. Notably, no malignancies were shown to be adversely correlated with MSI, whereas HDAC8 expression had a favorable relationship with MSI in ACC, KIRP, HNSC, etc. (Fig. 9C, Additional file 10: Figure S8).

The relationship between HDAC expression and immune checkpoint genes, such as CTLA4, PD-1, and PD-L1 (A), as well as TMB (B) and MSI (C) in 33 tumor types
The role of HDACs expression in immunotherapy of independent cohorts
A correlation analysis was conducted between HDAC expression and immunotherapy effectiveness in renal cell carcinoma, bladder cancer, and melanoma. IMvigor210, GSE111636, GSE176307, GSE78220, GSE67501, as well as our own TRUCE-01 dataset were used for this analysis. The study shows that there was a significant correlation (P < 0.09) between the expression of HDAC1/2/3 and the therapeutic objective response to anti-PD-L1. Interestingly, the GSE111636 cohort also demonstrated comparable results for HDAC3, with a statistically significant level of correlation (P = 0.017). We found that the expression levels of HDAC6 and HDAC10 had a poor correlation (P = 0.03) with the response rate to immunotherapy in the GSE111636 and TRUCE-01 datasets.
Other members should be regarded as of borderline statistical significance (0.1 ≤ P < 0.3). In summary, the findings suggest that assessing the expression of HDACs may serve as a valuable predictor for determining the responsiveness of anti-PD-1/PD-L1 immunotherapy.
Furthermore, we explored the possible mechanisms behind HDAC manifestations and immunotherapy response. In the present single-arm phase 2 clinical trial (TRUCE-01, NCT04730219), a notable increase in HDAC4 or HDAC9 expression was observed in BC patients who received tislelizumab combined with nab-paclitaxel therapy, regardless of their response to treatment. In non-responsive cases, however, there was a decrease in expression of HDAC7 after treatment (Fig. 10B–G). Therefore, these findings suggest a strong association between the HDAC gene expression and observed immunotherapy responses. This aspect will be further investigated in our future research endeavors.

The association between HDACs expression and clinical immunotherapy effect from several immunotherapy datasets. (A) Expression of HDACs in different cohorts based on their response to immunotherapy. (B-G) Changes of HDAC1/4/6/7/8/9 expression level before and after immunotherapy based on our Truce01 dataset
Drug sensitivity analysis
In order to examine whether HDAC expression may be associated with anti-cancer drug sensitivity in different types of cancer, we obtained relevant data from the CellMiner™ database. The results indicated that sensitivity to various chemotherapeutic drugs correlates significantly with HDAC expression (Additional file 11: Table S2, P < 0.05). HDAC7 expression was found to be inversely correlated and the sensitivity of Selumetinib, Cobimetinib, Trametinib and PD-98059, in contrast to a positive correlation between HDAC7 expression and the sensitivity of Everolimus, Rapamycin, Temsirolimus. The expression of HDAC4, PX-316, and chelerythrine drug sensitivity was found to be favorably correlated. Chelerythrine and Acrichine exhibited a favorable correlation with HDAC1 activity. Moreover, pharmaceutical tolerance to Ifosfamide, Oxiliplatin, Imexon, Lomustine, BN-2629, and Eribulin mesylate was inversely correlated with HDAC11 expression. Beside this, correlations results between HDAC expression and the sensitivity of commonly used chemotherapeutic agents against tumors are presented herein (Fig. 11B, P < 0.05). Paclitaxel, Vinblastine, Doxorubicin, or Docetaxel IC50 have an adverse association with HDAC7/11 activity. However, there was a beneficial association between gemcitabine's IC50 and HDAC3.

Pan-cancer drug sensitivity and HDACs. (A) We identified the top 25 drugs that were significantly correlated with HDACs expression (P < 0.01). (B) The association between the expression of HDACs and commonly employed chemotherapeutic drugs in cancer treatment is noteworthy (P < 0.05)
GSEA analysis of HDACs expression in bladder cancer
To assess the biological function and signaling pathways, we employed GSEA to identify enriched KEGG pathways within the groups with high and low levels of each HDAC gene family (Additional file 12: Table S3). High- or low-expression groups of different HDACs in bladder cancer showed significantly differential enrichment of tumor- and immune-related pathways including “CYTOKINE CYTOKINE RECEPTOR INTERACTION”, “OLFACTORY TRANSDUCTION”, “JAK STAT SIGNALING PATHWAY”, “CALCIUM SIGNALING PATHWAY”, “TGF BETA SIGNALING PATHWAY”, and “WNT SIGNALING PATHWAY”, and so on (Additional file 13: Figure S9).
Immune subgroups and clinical traits correspond to HDAC expression in BC
We compared the levels of HDACs across the five immune subtypes in BLCA. The findings highlighted significant differences in HDAC expression levels across the subtypes (C1–C4 and C6), with the exception of HDAC4/5/6 and HDAC8 (Additional file 14: Figure S10A). Furthermore, we explored the association between HDACs expression and bladder cancer clinical traits, such as stage of tumor node metastasis (TNM), tumor grade, and categorization of tumors into non-papillary or papillary subtypes. Several HDACs showed differential expression in bladder tumor tissues based on tumor grade, TNM stage, and tumor subtype, especially HDAC1, HDAC4, HDAC10, and HDAC11 (Additional file 14: Figure S10B-G).
Correlation between HDACs and stemness index, TMB, MSI, or TME in BC
As shown in Fig. 12, most HDAC family gene expression levels in BLCA patients were positively or negatively related with the RNAss/DNAss/immuneSore/stromalScore, except for HDAC7. Furthermore, the TIMER database was utilized to investigate the relationship between the expression of HDACs and the infiltration of six major immune cell types in BLCA. Findings demonstrated a substantial correlation between HDAC expression and the variety of lymphocytes penetrating BLCA (Fig. 13). The above results illustrated that HDACs might impact the survival outcomes of individuals with cancer by influencing immune cells infiltration in the TME.

Correlations between the expression of HDACs and RNAss, DNAss, StromaScore, ImmuneScore, ESTIMATEScore, TMB, and MSI in BLCA

Correlations between the expression of HDACs and immune infiltration by six frequently focused immune cells in BLCA based on TIMER database
Correlations between HDAC family CNVs, mutations, and immune infiltration in BLCA
To investigate the factors contributing to alterations in HDACs expression and the immune microenvironment, we examined the association between CNVs or mutations in HDACs and the immune microenvironment using data from the TIMER database. Figure 14A shows the relationships between changes in the copy number of HDACs and six types of immune infiltration in BLCA. In particular, copy number high-amplification or arm-level gain of HDAC3 and HDAC8 was associated with substantially lower levels of immune infiltrates (CD4 + T cells, dendritic cells, neutrophils, and CD8 + T cells) in BLCA. These findings may indicate a potential mechanism by which gene alterations in the HDAC family gene may predict responses to immunotherapy. Subsequently, all BLCA samples were classified according to the mutation status of HDACs, and difference analysis was employed to investigate their potential relationship with the level of immune cell infiltration (Fig. 14B). Mutation groups of HDAC8 or HDAC5 were enriched in NK cells and CD8 + T cells; however, mutation groups of HDAC6 or HDAC11 exhibited lower levels in CD8 + T cells compared to non-mutation groups.

The correlations between variations in the copy numbers of HDACs (A) or mutations (B), and the presence of immune cell infiltrations in BLCA
Validation of HDACs protein expression in BC from Human Protein Atlas (HPA)
The HPA database was used to compare HDAC protein levels between BC and normal bladder tissues. Compared with expression in adjacent tissues, cancer tissues exhibited an increase in the expression of HDAC1/3 and a decrease in HDAC4/5/9. These findings align with our previous analysis of transcriptome differences for these genes (Fig. 15A-I).

Verification of protein-level expression of HDACs. (A-I) Immunohistochemical staining for HDAC1-6 and HDAC8-10 protein expression based on HPA database
Discussion
Histone post-translational modifications (PTM), including lysine acetylation, are essential for controlling important genes implicated in the development and propagation of cancer [30]. Their expression of HDACs has been found to enhance the development and spread of tumors by reducing the levels of pro-apoptotic factors and facilitating angiogenesis, invasion, and migration [31, 32]. Acetylation has been validated as a key component in regulating gene expression, prompting scholars to investigate the correlation between HDACs and the aberrant gene expression commonly observed in pan-cancer [33]. The HDACs are well-acknowledged to be significant regulators of tumor differentiation, apoptosis, and proliferation in a variety of malignancies [34,35,36]. In recent years, HDAC family genes have attracted extensive scholarly attention in tumorigenesis and progression studies. Studies have reported the significant association of HDACs with tumor proliferation, differentiation, and poor prognosis [34, 37, 38], as well as the application of HDACs and HDACs inhibitors as therapeutic targets in both research and clinical treatment [39,40,41].
The present research comprised an organized study of HDACs in all types of cancer. First, we compared the abundance of HDACs between healthy tissues and 33 distinct kinds of pan-cancers. The expression of HDACs is generally high in most tumors compared to normal tissues, exhibiting high heterogeneity between tumor types. The findings revealed that HDAC1 was highly expressed in 12 tumors, while 3 tumors showed a low expression. HDAC8 exhibited significant overexpression in 14 tumors but was significantly lowly expressed in KIRC. Furthermore, HDAC10 was significantly overexpressed in 15 tumors and lowly expressed in KICH. HDAC4 displayed higher expression in LIHC and CHOL, whereas its expression was low in other 13 tumors. Thus, our study contributes to the potential application of HDACs as prognostic markers for tumors and development of HDAC-targeted therapies. In KICH and LGG, elevated levels of HDAC1, HDAC2, and HDAC3 had a strong association with a poorer prognosis, whereas elevated level of HDAC5 has been linked to an improved OS, showing that HDACs may be useful potential tumor markers. The expression of various HDACs also correlated with survival rates in various types of cancer. Studies on mutations in the HDAC family of genes in tumor tissues are rare. Our research showed that HDAC9 and HDAC6 had the highest mutation frequencies of all members, 10 and 8%, respectively. The overall mutation rate of this family gene in pan-cancer was 27.32%. However, it was shown that HDAC variants in OS individuals possessed no apparent clinical consequence. In addition, the proteomic expression data of HDACs were validated by HPA database.
Interestingly, this study also demonstrated an association between immune-mediated subtypes and all HDAC members, with the immune subtypes associated with poorer prognosis (C1, C2 and C6) [42] having the highest relative expression in HDAC1 and HDAC3. Immune infiltration is a key determinant of cancer responsiveness and prognosis [43]. To assess the association between HDAC expression and the immune microenvironment, we utilized ImmuneScore and StromalScore to calculate the ratio of HDAC expression to immune cells across pan-cancer. Two independent stemness indices (RNAss and DNAss) have been obtained by analyzing tumor transcriptome, methylome, and transcription factor binding sites in multiple platforms. They reflect gene expression and epigenetic characteristics [44]. Importantly, researchers have found that stemness indices have been linked to cancer pathology, PD-L1 levels, and immune microenvironment, as well as have been suggestive of metastatic tumors and anti-cancer drugs [45]. Our findings revealed significant positive or negative correlations between HDAC expression and stemness indices in different cancer types, particularly THYM and BLCA. Immune checkpoint inhibitors disrupt the interaction between immune checkpoints and their ligands, interfering immune tolerance, boosting the activity of immune cells, and facilitating the immune system in eliminating tumor cells, thereby impeding the initiation and progression of cancers [46]. A growing body of evidence suggests that TMB is a potential prognostic and predictive biomarker for tumors therapy [47, 48]. Several studies have reported that MSI holds promise as a predictive biomarker, offering valuable insights for guiding decisions on adjuvant chemotherapy, targeted therapy, and immunotherapy in tumor treatment [49, 50]. Our findings also suggest a significant correlation between HDACs and the tumor microenvironment, TMB, MSI and multiple immune checkpoints (such as CTLA4, PD-L1, and PD-1) in various tumors (including BLCA). This indicates that different expression levels of the HDAC family may affect TMB, MSI, and immune escape in tumors, affecting the patient sensitivity to immune checkpoint inhibition treatment, offering new evidence for the prognosis of immunotherapy.
Our single-arm phase 2 trial (NCT04730219) has demonstrated that combination of tislelizumab and nab-paclitaxel shows encouraging clinical benefits in patients with MIBC. Based on the studies mentioned earlier, we conducted further investigation into the potential prognostic value of HDACs in bladder cancer, specifically in relation to immunotherapy. Our findings demonstrated that anti-PD-L1 treatment was effective when HDAC1/HDAC8 expression levels were high in the Imvigor210 cohort. Furthermore, we observed a significant increase in HDAC4 expression among bladder cancer patients who positively responded to tislelizumab plus nab-paclitaxel therapy following treatment. Additionally, our GSEA analysis uncovered a noteworthy enrichment of pathways associated with tumors and the immunoreactive in either the high or low expression group of HDACs in BLCA. Previous studies have also indicated that the calcium signaling pathway is not only essential for the advancement and susceptibility to cellular demise but also for the formation and sustenance of resistance to multiple drugs and the TME [51]. In the TME, the Wnt protein interacts with LRP5/LRP6 co-receptors found on DC cells to initiate the classical Wnt signaling pathway. This activation leads to immune tolerance, inhibition of effector T cell functions, and alteration of antitumor effects [52]. These signaling pathways have a critical impact on various tumors to varying degrees, and may provide clues for future mechanistic studies.
Researchers have observed that a heightened TMB level and a substantial presence of CD8 + effector T cells are associated with a positive response to anti-PD-L1 treatment in metastatic urothelial cancer [53]. The primary immune markers for immunotherapy are tumor-infiltrating immune cells [54]. T cells (including CD8 + T and CD4 + T cells) have a crucial impact on the occurrence, progression, and therapy of tumors, particularly in the field of immunotherapy [55]. Simultaneously, our findings align with prior research, suggesting that HDAC9 may potentially modulate an immune response against tumors by decreasing the infiltration of CD8 + T and DCs cells [56]. Our further analysis suggested that the expression of different HDAC family genes is correlated with TMB / MSI in BLCA. Another key finding of our study revealed that HDACs levels are highly correlated with immune infiltration of six immune cells, especially CD8 + T cells in BLCA. It was previously demonstrated that increased expression of some HDACs in solid tumors including LUAD, BLCA, OV, PAD, and STAD is usually associated with poor prognosis [57]. Furthermore, OV and LUAD cellular models showed that silencing the HDAC1 gene reduced tumor cell resistance [58] and aggressiveness [59]. Moreover, research results have shown that HDAC3 exhibits high expression in STAD, PAD, and READ [60], and the overexpression of HDAC1 and 2 in READ is associated with poor prognosis [61, 62]. Overexpression of HDAC8 in KICH, NBL and UVM was associated with poor prognosis, whereas ACC, LUSC, SKCM and STAD had significantly better prognosis compared with patients with low expression [63]. It has been commonly noted in prior discussions that elevated HDAC7 expression is linked to an unfavorable prognosis in various malignant hematological diseases, including ALL and CLL [57]. It has been reported that HDAC9 expression is generally low in a variety of tumors. The downregulation of HDAC9 expression is also observed in colorectal adenocarcinoma, while low expression in ACC and CHOL is associated with significantly poor prognosis [63]. Combined with existing studies, we propose that HDAC may be responsible for a number of cancers through an inclusive study, and aberrant HDACs expression in most cancers is considered a risk factor for poor tumor prognosis. In addition, we not only demonstrated that there was a strong relationship between HDAC CNVs and immune infiltrates in BLCA, but also found that immune infiltration of different immune cells may be influenced by HDAC mutation status. These results raise the prospect that HDACs may be employed as predictive biomarkers or indicators of the effectiveness of immune checkpoint medications in BC patients. Moreover, we explored the correlation between HDAC expression and drug sensitivity in pan-cancer using the CellMiner™ database, designed for the integration and research of data derived from the NCI-60 cancer cell line on cellular and pharmacological properties [64, 65]. We found that HDAC7 expression was negatively related to drug sensitivity for Selumetinib, Cobimetinib, Trametinib and PD-98059, and positively correlated with Everolimus, Rapamycin and Temsirolimus correlation. Similarly, the sensitivity to Carmustine, Oxaliplatin, Ifosfamide, Lomustine, BN-2629 and Eribulin mesilate gradually decreased with increasing HDAC11 expression. In summary, our research revealed that the expression levels of HDAC1, HDAC3, HDAC4, HDAC7, HDAC9 and HDAC11 in tumor cells were associated with the sensitivity of certain drugs, that may serve as a basis for choosing therapeutic medications. Whether cancer cells are sensitive or resistant to drug therapy may depend on the activity of HDACs.
Although we performed a pan-cancer study to investigate the association between HDAC expression and immunotherapy, tumor microenvironment, drug sensitivity, and survival prognosis, there remain a few drawbacks. Initially, no in vivo or in vitro tests were conducted; this study solely comprised a pan-cancer multi-omics evaluation of the HDAC family based on public and our sequencing datasets. Secondly, the prognosis of tumor patients remains unclear regarding the influence of HDAC's immune-mediated infiltration.
Conclusion
In summary, we found that there are significant variations in the expression of HDACs among variety of tumors. We also discovered that these variations have strong association with aspect of the tumor microenvironment, immune subtypes, prognosis, drug sensitivity, infiltration of immune cells, immunotherapy indicators (TMB, MSI), and immune checkpoints in 33 types of tumors, including BLCA.
The present study highlights plausible pathways by which HDACs aid in the progress of malignancies, suggesting a potentially pivotal function in the progression of cancer formation, specifically in the areas of immunological infiltration, tumor environment regulation, and sensitivity to drugs. These findings have significant consequences for the development of customized therapies for cancer.
Availability of data and materials
Online repositories store the datasets used in this study. The study’s distinct findings have been included in the article and additional files. For additional queries, please contact the corresponding authors.
Abbreviations
- HDAC:
-
The histone deacetylase family
- TME:
-
Tumor micro environment
- MSI:
-
Microsatellite instability
- BC:
-
Bladder cancer
- HPA:
-
Human protein atlas
- WHO:
-
World Health Organization
- GEO:
-
Gene expression omnibus
- TCGA:
-
The cancer genome atlas
- CR:
-
Complete response
- PR:
-
Partial response
- SD:
-
Stable disease
- PD:
-
Progressive disease
- OS:
-
Overall survival
- DSS:
-
Disease specific survival
- RFS:
-
Recurrence free survival
- PFS:
-
Progression free survival
- CNVs:
-
Copy numbers
- SNVs:
-
Single nucleotide variations
- cBioPortal:
-
The cBio Cancer Genomics Portal
- CNAs:
-
Copy number alterations
- T:
-
Stage_tumor
- N:
-
Stage_node
- M:
-
Stage_metastasis
- GSEA:
-
Gene set enrichment analysis
- TIMER:
-
Tumor immune estimation resource
- HPA:
-
Human Protein Atlas
- TIMER:
-
The tumor immune estimate resource
- immune cell abundance:
-
Immune scores
- stromal cell abundance:
-
Stromal scores
- TMB:
-
Tumor mutational burden
- MSI:
-
Microsatellite instability
- RNAss:
-
RNA stemness score
- DNAss:
-
DNA stemness score
- NCI:
-
The National Cancer Institute
- KEGG:
-
The Kyoto Encyclopedia of Genes and Genomes
- TNM:
-
Tumor node metastasis
- ACC:
-
Adrenocortical cancer
- BLCA:
-
Bladder cancer
- BRCA:
-
Breast cancer
- CESC:
-
Cervical cancer
- CHOL:
-
Bile duct cancer
- COAD:
-
Colon cancer
- DLBC:
-
Large B-cell lymphoma
- ESCA:
-
Esophageal cancer
- GBM:
-
Glioblastoma
- HNSC:
-
Head and neck cancer
- KICH:
-
Kidney chromophobe
- KIRC:
-
Kidney clear cell carcinoma
- KIRP:
-
Kidney papillary cell carcinoma
- LAML:
-
Acute myeloid leukemia
- LGG:
-
Lower grade glioma
- LIHC:
-
Liver cancer
- LUAD:
-
Lung adenocarcinoma
- LUSC:
-
Lung squamous cell carcinoma
- MESO:
-
Mesothelioma
- OV:
-
Ovarian cancer
- PAAD:
-
Pancreatic cancer
- PCPG:
-
Pheochromocytoma and paraganglioma
- PRAD:
-
Prostate cancer
- READ:
-
Rectal cancer
- SARC:
-
Sarcoma
- SKCM:
-
Melanoma
- STAD:
-
Stomach cancer
- TGCT:
-
Testicular cancer
- THCA:
-
Thyroid cancer
- THYM:
-
Thymoma
- UCEC:
-
Endometrioid cancer
- UCS:
-
Uterine carcinosarcoma
- UVM:
-
Ocular melanomas
References
Ferlay J, Colombet M, Soerjomataram I et al (2021) Cancer statistics for the year 2020: an overview. Int J Cancer. 149:778–789
Cerqueira M, Belmonte-Reche E, Gallo J, Baltazar F, Bañobre-López M (2022) Magnetic solid nanoparticles and their counterparts: recent advances towards cancer. Theranostics Pharm 14(3):506
Chen W, Sun Z, Lu L (2021) Targeted engineering of medicinal chemistry for cancer therapy: recent advances and perspectives. Angew Chem Int Ed Engl 60(11):5626–5643
Graham TA, Sottoriva A (2017) Measuring cancer evolution from the genome. J Pathol 241(2):183–191
Conforti F, Pala L, Bagnardi V, De Pas T, Martinetti M, Viale G et al (2018) Cancer immunotherapy efficacy and patients’ sex: a systematic review and meta-analysis. Lancet Oncol 19(6):737–746
Printz C (2014) Targeted therapy in lung cancer: survival, quality of life improved for some patients. Cancer 120(17):2625–2626
Xiong J, Bing Z, Guo S (2019) Observed survival interval: a supplement to TCGA pan-cancer clinical data resource. Cancers 11(3):280
Lv Z, Qi L, Hu X, Mo M, Jiang H, Fan B et al (2021) Zic Family Member 2 (ZIC2): a potential diagnostic and prognostic biomarker for pan-cancer. Front Mol Biosci 8:631067
Haberland M, Montgomery RL, Olson EN (2009) The many roles of histone deacetylases in development and physiology: implications for disease and therapy. Nat Rev Genet 10(1):32–42
Yang C, Croteau S, Hardy P (2021) Histone deacetylase (HDAC) 9: versatile biological functions and emerging roles in human cancer. Cell Oncol (Dordr) 44(5):997–1017
Consalvi S, Saccone V, Giordani L, Minetti G, Mozzetta C, Puri PL (2011) Histone deacetylase inhibitors in the treatment of muscular dystrophies: epigenetic drugs for genetic diseases. Mol Med 17(5–6):457–465
Chuang DM, Leng Y, Marinova Z, Kim HJ, Chiu CT (2009) Multiple roles of HDAC inhibition in neurodegenerative conditions. Trends Neurosci 32(11):591–601
Zhu P, Martin E, Mengwasser J, Schlag P, Janssen KP, Göttlicher M (2004) Induction of HDAC2 expression upon loss of APC in colorectal tumorigenesis. Cancer Cell 5(5):455–463
Bheda P, Jing H, Wolberger C, Lin H (2016) The substrate specificity of sirtuins. Annu Rev Biochem 85:405–429
Guarente L (2006) Sirtuins as potential targets for metabolic syndrome. Nature 444(7121):868–874
Banik D, Moufarrij S, Villagra A (2019) Immunoepigenetics combination therapies: an overview of the role of HDACs in cancer immunotherapy. Int J Mol Sci. 20(9):2241
Liu J, Lichtenberg T, Hoadley KA, Poisson LM, Lazar AJ, Cherniack AD et al (2018) An integrated TCGA pan-cancer clinical data resource to drive high-quality survival outcome analytics. Cell 173(2):400–16.e11
Yang X, Miao Y, Wang J, Mi D (2021) A pan-cancer analysis of the HER family gene and their association with prognosis, tumor microenvironment, and therapeutic targets. Life Sci 273:119307
Gao J, Aksoy BA, Dogrusoz U, Dresdner G, Gross B, Sumer SO et al (2013) Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci Signal. 6(269):11
Li T, Fan J, Wang B, Traugh N, Chen Q, Liu JS et al (2017) TIMER: a web server for comprehensive analysis of tumor-infiltrating immune cells. Cancer Res. 77(21):e108–e110
Diboun I, Wernisch L, Orengo CA, Koltzenburg M (2006) Microarray analysis after RNA amplification can detect pronounced differences in gene expression using limma. BMC Genomics 7:252
Gonzalez-Pastor R, Goedegebuure PS, Curiel DT (2021) Understanding and addressing barriers to successful adenovirus-based virotherapy for ovarian cancer. Cancer Gene Ther 28(5):375–389
van Velzen M, Derks S, van Grieken N, Haj Mohammad N, van Laarhoven H (2020) MSI as a predictive factor for treatment outcome of gastroesophageal adenocarcinoma. Cancer Treat Rev 86:102024
Liu J, Zhang S, Dai W, Xie C, Li JC (2020) A comprehensive prognostic and immune analysis of SLC41A3 in pan-cancer. Front Oncol 10:586414
Li L, Feng Q, Wang X (2020) PreMSIm: an R package for predicting microsatellite instability from the expression profiling of a gene panel in cancer. Comput Struct Biotechnol J 18:668–675
Qu YY, Guo RY, Luo ML, Zhou Q (2021) Pan-Cancer Analysis of the Solute Carrier Family 39 Genes in Relation to Oncogenic, Immune Infiltrating, and Therapeutic Targets. Front Genet 12:757582
Tamborero D, Rubio-Perez C, Muiños F, Sabarinathan R, Piulats JM, Muntasell A et al (2018) A pan-cancer landscape of interactions between solid tumors and infiltrating immune cell populations. Clin Cancer Res 24(15):3717–3728
Chockley PJ, Keshamouni VG (2016) Immunological consequences of epithelial-mesenchymal transition in tumor progression. J Immunol 197(3):691–698
Xu WX, Zhang J, Hua YT, Yang SJ, Wang DD, Tang JH (2020) An integrative pan-cancer analysis revealing LCN2 as an oncogenic immune protein in tumor microenvironment. Front Oncol 10:605097
Audia JE, Campbell RM (2016) Histone modifications and cancer. Cold Spring Harb Perspect Biol 8(4):a019521
Glozak MA, Seto E (2007) Histone deacetylases and cancer. Oncogene 26(37):5420–5432
Yang XJ, Seto E (2007) HATs and HDACs: from structure, function and regulation to novel strategies for therapy and prevention. Oncogene 26(37):5310–5318
de Ruijter AJ, van Gennip AH, Caron HN, Kemp S, van Kuilenburg AB (2003) Histone deacetylases (HDACs): characterization of the classical HDAC family. Biochem J 370(Pt 3):737–749
Pouloudi D, Manou M, Sarantis P, Tsoukalas N, Tsourouflis G, Dana E et al (2021) Clinical Significance of histone deacetylase (HDAC)-1, -2, -4 and -6 expression in salivary gland tumors. Diagnostics (Basel). 11(3):517
Weichert W (2009) HDAC expression and clinical prognosis in human malignancies. Cancer Lett 280(2):168–176
Seo J, Min SK, Park HR, Kim DH, Kwon MJ, Kim LS et al (2014) Expression of histone deacetylases HDAC1, HDAC2, HDAC3, and HDAC6 in invasive ductal carcinomas of the breast. J Breast Cancer 17(4):323–331
Zhang K, Liu Z, Yao Y, Qiu Y, Li F, Chen D et al (2021) Structure-based design of a selective class I histone deacetylase (HDAC) near-infrared (NIR) probe for epigenetic regulation detection in triple-negative breast cancer (TNBC). J Med Chem 64(7):4020–4033
Lutz L, Fitzner IC, Ahrens T, Geißler AL, Makowiec F, Hopt UT et al (2016) Histone modifiers and marks define heterogeneous groups of colorectal carcinomas and affect responses to HDAC inhibitors in vitro. Am J Cancer Res 6(3):664–676
Jain S, Zain J (2011) Romidepsin in the treatment of cutaneous T-cell lymphoma. J Blood Med 2:37–47
Mann BS, Johnson JR, Cohen MH, Justice R, Pazdur R (2007) FDA approval summary: vorinostat for treatment of advanced primary cutaneous T-cell lymphoma. Oncologist 12(10):1247–1252
Edwards CM, Johnson RW (2021) Targeting histone modifications in bone and lung metastatic cancers. Curr Osteoporos Rep 19(3):230–246
Thorsson V, Gibbs DL, Brown SD, Wolf D, Bortone DS, Ou Yang TH et al (2019) The immune landscape of cancer. Immunity 51(2):411–412
McGranahan N, Furness AJ, Rosenthal R, Ramskov S, Lyngaa R, Saini SK et al (2016) Clonal neoantigens elicit T cell immunoreactivity and sensitivity to immune checkpoint blockade. Science 351(6280):1463–1469
Sokolov A, Paull EO, Stuart JM (2016) One-class detection of cell states in tumor subtypes. Pac Symp Biocomput 21:405–416
Malta TM, Sokolov A, Gentles AJ, Burzykowski T, Poisson L, Weinstein JN et al (2018) Machine learning identifies stemness features associated with oncogenic dedifferentiation. Cell 173(2):338–54.e15
Zhou M, Sun X, Zhu Y (2021) Analysis of the role of Frizzled 2 in different cancer types. FEBS Open Bio 11(4):1195–1208
Samstein RM, Lee CH, Shoushtari AN, Hellmann MD, Shen R, Janjigian YY et al (2019) Tumor mutational load predicts survival after immunotherapy across multiple cancer types. Nat Genet 51(2):202–206
Tang Y, Li J, Xie N, Yang X, Liu L, Wu H et al (2020) PIK3CA gene mutations in the helical domain correlate with high tumor mutation burden and poor prognosis in metastatic breast carcinomas with late-line therapies. Aging 12(2):1577–1590
Zhang CM, Lv JF, Gong L, Yu LY, Chen XP, Zhou HH et al (2016) Role of Deficient mismatch repair in the personalized management of colorectal cancer. Int J Environ Res Public Health. 13(9):892
Shen H, Zhong M, Wang W, Liao P, Yin X, Rotroff D et al (2017) EBV infection and MSI status significantly influence the clinical outcomes of gastric cancer patients. Clin Chim Acta 471:216–221
Monteith GR, Prevarskaya N, Roberts-Thomson SJ (2017) The calcium-cancer signalling nexus. Nat Rev Cancer 17(6):367–380
Suryawanshi A, Manoharan I, Hong Y, Swafford D, Majumdar T, Taketo MM et al (2015) Canonical wnt signaling in dendritic cells regulates Th1/Th17 responses and suppresses autoimmune neuroinflammation. J Immunol 194(7):3295–3304
Mariathasan S, Turley SJ, Nickles D, Castiglioni A, Yuen K, Wang Y et al (2018) TGFβ attenuates tumour response to PD-L1 blockade by contributing to exclusion of T cells. Nature 554(7693):544–548
Pandya PH, Murray ME, Pollok KE, Renbarger JL (2016) The immune system in cancer pathogenesis: potential therapeutic approaches. J Immunol Res 2016:4273943
Kochenderfer JN, Dudley ME, Kassim SH, Somerville RP, Carpenter RO, Stetler-Stevenson M et al (2015) Chemotherapy-refractory diffuse large B-cell lymphoma and indolent B-cell malignancies can be effectively treated with autologous T cells expressing an anti-CD19 chimeric antigen receptor. J Clin Oncol 33(6):540–549
Ning Y, Ding J, Sun X, Xie Y, Su M, Ma C, et al. HDAC9 deficiency promotes tumor progression by decreasing the CD8+ dendritic cell infiltration of the tumor microenvironment. J Immunother Cancer. 2020;8(1).
Li Y, Seto E (2016) HDACs and HDAC inhibitors in cancer development and therapy. Cold Spring Harb Perspect Med. 6(10):a026831
Liu X, Yu Y, Zhang J, Lu C, Wang L, Liu P et al (2018) HDAC1 silencing in ovarian cancer enhances the chemotherapy response. Cell Physiol Biochem 48(4):1505–1518
Zhang L, Bu L, Hu J, Xu Z, Ruan L, Fang Y et al (2018) HDAC1 knockdown inhibits invasion and induces apoptosis in non-small cell lung cancer cells. Biol Chem 399(6):603–610
Wilson AJ, Byun DS, Popova N, Murray LB, L’Italien K, Sowa Y et al (2006) Histone deacetylase 3 (HDAC3) and other class I HDACs regulate colon cell maturation and p21 expression and are deregulated in human colon cancer. J Biol Chem 281(19):13548–13558
Krishnan M, Singh AB, Smith JJ, Sharma A, Chen X, Eschrich S et al (2010) HDAC inhibitors regulate claudin-1 expression in colon cancer cells through modulation of mRNA stability. Oncogene 29(2):305–312
Weichert W, Röske A, Niesporek S, Noske A, Buckendahl AC, Dietel M et al (2008) Class I histone deacetylase expression has independent prognostic impact in human colorectal cancer: specific role of class I histone deacetylases in vitro and in vivo. Clin Cancer Res 14(6):1669–1677
Milazzo G, Mercatelli D, Di Muzio G, Triboli L, De Rosa P, Perini G et al (2020) Histone deacetylases (HDACs): evolution, specificity, role in transcriptional complexes, and pharmacological actionability. Genes (Basel). 11(5):556
Reinhold WC, Varma S, Sunshine M, Elloumi F, Ofori-Atta K, Lee S et al (2019) RNA sequencing of the NCI-60: integration into cell miner and cell miner CDB. Cancer Res 79(13):3514–3524
Reinhold WC, Sunshine M, Varma S, Doroshow JH, Pommier Y (2015) Using cell miner 16 for systems pharmacology and genomic analysis of the NCI-60. Clin Cancer Res. 21(17):3841–52
Acknowledgements
Data from the TCGA and GEO databases are greatly appreciated by the authors.
Funding
This research received support from the Tianjin Municipal Health Industry Key Project (No. TJWJ2022XK014), Technology Project of Tianjin Binhai New Area Health Commission (No. 2019BWKY026), Tianjin Institute of Urology Talent Funding Program (No. MYSRC202310) and Scientific Research Project of Tianjin Municipal Education Commission (No. 2022ZD069).
Author information
Authors and Affiliations
Contributions
The conception and design were performed by HY and HH. CS, KW, and YZ wrote and analyzed the manuscript. The literature search and data collection were conducted by ZZ and ZL-Z. HH, HY and CS interpreting data. Study supports, discussion and revision were provided by CS-C, YZ, CS, YQ, HY and HH. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
This research was performed following the instructions stated in the Declaration of Helsinki and received approval from the ethics committee at Tianjin Medical University’s 2nd Affiliated Hospital (Ethics code: KY2021K003).
Consent for publication
Informed consent forms were obtained from participants before clinical resources were used.
Competing interests
None.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1. Figure S1:
A boxplot representing the expression levels of each HDAC gene in 33 cancer tissues from TCGA database has been created (A-K).
Additional file 2. Figure S2:
The levels of expression of HDACs in cancer and healthy tissues across several cancers were visually compared with box plots (A-K). *P < 0.05, **P < 0.01, and ***P < 0.001.
Additional file 3. Table S1:
K-M analysis findings for OS, PFS, DFS and DSS were aggregated across 33 cancer types according to the HDACs expression level.
Additional file 4. Additional Data1:
The OS, PFS, DFS and DSS Kaplan‐Meier curves were plotted across 33 cancer types according to the HDACs expression level.
Additional file 5. Figure S3:
Relationship between expression levels of HDACs and cancer stage in different types of cancer.
Additional file 6. Figure S4:
Alteration frequency of each HDAC gene was separately presented in (A-K) at a pan-cancer level and different types of cancers level based on the pan-cancer datasets (2683 samples) obtained from cBioPortal, provided by ICGC/TCGA.
Additional file 7. Figure S5:
Relationship between SNVs and mRNA levels of HDAC expression based on the ICGC/TCGA pan-cancer datasets.
Additional file 8. Figure S6:
An analysis of the relationship between HDAC gene expression and 47 immune checkpoints expressed in a variety of tumors (A-K).
Additional file 9. Figure S7:
A radar chart illustrating the relationship between HDAC expression and TMB (A-K).
Additional file 10. Figure S8:
Mapping the correlation between expression of HDACs and MSI using a radar chart (A-K).
Additional file 11. Table S2:
Drug sensitivity analysis of HDACs based on the CellMiner database.
Additional file 12. Table S3:
Gene Set Enrichment Analysis was performed for each HDAC family member using the KEGG gene sets based on TCGA_BLCA datasets.
Additional file 13. Figure S9:
GSEA analysis of expression of each HDAC family member in bladder cancer (A-K).
Additional file 14. Figure S10:
Associations between the level of HDACs expression and immune subtypes (A), clinicopathological characteristics (B-G) in BC.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Chen, C., Wang, K., Zhang, Y. et al. Prognosis and immunological characteristics of HDAC family in pan-cancer through integrative multi-omic analysis. Egypt J Med Hum Genet 25, 44 (2024). https://doi.org/10.1186/s43042-024-00518-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43042-024-00518-1