
Abstract
Background
Tree hollows are an important habitat resource used by arboreal fauna for nesting and denning. Hollows form when trees mature and are exposed to decay and physical damage. In the absence of excavating fauna, hollow formation can take up to 200 years in Australian temperate Eucalyptus forests, making tree hollows a critical but slow forming habitat feature. The increasing frequency and severity of wildfires due to climate change has led to increased concern about the landscape-scale loss of nesting space for arboreal fauna, including endangered species such as the folivorous southern greater glider (Petauroides volans). To understand patterns of nesting resource availability, we assessed drivers of hollow occurrence in southeastern Australian mixed-species Eucalyptus forests and quantified the effects of an unprecedented large-scale wildfire, the 2019/2020 Black Summer bushfires, on hollow occurrence and abundance.
Results
Tree size and shape, as well as site productivity and topography, were important predictors for hollow occurrence both before and after the fires. The occurrence of the southern greater glider was strongly dependent on high proportions of hollow-bearing trees. While high fire severities had a negative impact on southern greater glider occurrence, the number of hollow-dependent arboreal species was not affected. While the wildfires significantly reduced hollow abundance, we did not find significant effects on hollow occurrence. Fires altered the relationship between tree size and hollow occurrence expressed as a change in the probability of hollow occurrence, with a higher likelihood at smaller tree sizes after the fires.
Conclusions
Our findings suggest that post-fire nesting space may be reduced at the tree-scale, while at the stand-scale, hollow-bearing trees persist as biological legacies. These persisting trees can support the recovery of hollow-dependent arboreal fauna, such as the endangered southern greater glider by providing denning and nesting space. Hollow-bearing trees that survived the fires have the potential to form new hollows faster compared to undisturbed mature trees.
Resumen
Antecedentes
Las cavidades de los árboles son importantes recursos de hábitats usados por la fauna arbórea para anidar y guarecerse. Estas cavidades se forman cuando los árboles maduran y se exponen al decaimiento y a los daños físicos. En ausencia de fauna excavadora, la formación de cavidades puede tardar hasta 200 años en Bosques templados de eucaliptus en Australia, haciendo que estas cavidades en los árboles sean una característica crítica, pero de formación lenta del hábitat. El incremento en la frecuencia y severidad de los incendios debido al cambio climático ha llevado a un incremento en la preocupación por la pérdida de espacios de anidamiento a escala de paisaje para las especies de fauna arbórea, incluyendo especies en peligro como el folívoro petauro gigante del sur (Petauroides volans). Para entender los patrones de disponibilidad de recursos de anidamiento, determinamos los factores conducentes que permiten la ocurrencia de estas cavidades en bosques mixtos de eucaliptus en el sudeste de Australia y cuantificamos los efectos de un incendio sin precedentes y de gran escala (el Black Summer Bushfire de 2019/2020), en la ocurrencia y abundancia de cavidades.
Resultados
El tamaño y forma de los árboles, como así también la productividad de los sitios y la topografía, fueron importantes predictores de la ocurrencia de cavidades antes y después de los incendios. La ocurrencia del petauro gigante del sur fue fuertemente dependiente de una alta proporción de árboles que tenían cavidades. Aunque las altas severidades de los fuegos tuvieron un impacto negativo en la ocurrencia del petauro gigante del sur, el número de especies de hábitos arbóreos dependientes de cavidades no fue afectada. Aunque los incendios reducen significativamente la abundancia de las cavidades, no encontramos efectos significativos en la ocurrencia de cavidades. Los incendios alteran la relación entre el tamaño del árbol y la ocurrencia de cavidades expresada como un cambio en la probabilidad de ocurrencia de una cavidad, con una alta probabilidad de ocurrencia en árboles de tamaños más pequeños luego de los incendios.
Conclusiones
Nuestros resultados sugieren que los espacios para anidamiento luego del post-fuego pueden reducirse en la escala de árbol individual, mientras que, a escala de rodal, los árboles que contienen cavidades para anidamiento persisten como legados biológicos. Los árboles persistentes en el sistema pueden sostener la recuperación de las especies de fauna arbórea dependientes de estas cavidades, como la especie en peligro petauro gigante del sur, proveyéndole de espacio para anidamiento y guarida. Los árboles con cavidades que sobrevivieron a los fuegos tienen el potencial de formar nuevas cavidades más rápidamente que aquellos árboles maduros no disturbados.
Similar content being viewed by others


Background
Tree hollows are among the most important habitat resources for many vertebrate species inhabiting forests (Gibbons and Lindenmayer 2002; Mazurek and Zielinski 2004). Hollows are needed for the denning and nesting of many arboreal species (Bennett et al. 1994; Gibbons et al. 2002; Koch et al. 2008a). Disturbances to mature forests from deforestation, land-use change, timber harvesting, planned fire, or wildfires can reduce this important resource and cause local extirpations of arboreal fauna (Diamond 1984; Lindenmayer et al. 2011; Youngentob et al. 2013; McLean et al. 2015). Natural disturbances such as wildfires are becoming more frequent, severe, and affect increasingly larger spatial extents (Collins et al. 2022). These large severe fires can impact the entire geographic distributions of many forest-dwelling fauna, as evidenced by the 2019/2020 “Black Summer” megafires in eastern Australia (Ward et al. 2020). There is now an accelerated need to assess and quantify the effects of such unprecedented events on critical habitat resources, such as tree hollows within forest landscapes, to inform and adapt forest management and conservation efforts (Woinarski et al. 2014; Davey 2018a; DELWP 2019; Geary et al. 2021).
Fauna most vulnerable to large severe landscape-scale fires are those that occupy specific niches, have specialized diets, a limited dispersal capacity, and/or small home-ranges (Powell 2000; Smith et al. 2007; Shipley et al. 2009; Bowman et al. 2014b; Davey 2018b). Therefore, arboreal folivores such as the southeastern Australian southern greater glider (Petauroides volans, SGG), are particularly vulnerable to stand- or crown-replacing disturbances (Kavanagh 2000; McLean et al. 2018). At the landscape-scale, this places such species among the most impacted by severe mega-fires (Chia et al. 2016; McLean et al. 2018; Nolan et al. 2020; Ward et al. 2020). SGGs have been listed as an endangered species as a result of the 2019/2020 Black Summer bushfires burning through large swaths of their geographic range (Ward et al. 2020; Legge et al. 2021). This development is concerning, as mega-fires have been increasing in size and severity in eastern Australia over the last 30 years (Collins et al. 2022).
Complex forested landscapes are typically characterized by multiple habitat features of varying quality (Lindenmayer et al. 1990b, 1991b; Nitschke et al. 2020). Hollow-bearing trees, required for denning and nesting, however, are considered essential for arboreal fauna to be present (Smith and Lindenmayer 1988; Goldingay 2011). While specialist arboreal species such as the SGG prefer sites where climatic conditions are within their narrow thermoregulatory capacities and that have trees of their preferred Eucalyptus species with suitable leaf chemistry (Rübsamen et al. 1984; Kavanagh and Lambert 1990; DeGabriel et al. 2009; Shipley et al. 2009; Youngentob et al. 2011; Wagner et al. 2020), they also require multiple (generally large) nesting trees within their territories (Gibbons and Lindenmayer 2002; Smith et al. 2007; Lindenmayer et al. 2022). Due to these requirements, SGGs are predominantly found in forests with cooler and wetter climates, e.g., at high elevations or in riparian areas at lower elevations (Smith and Smith 2020; Wagner et al. 2020; Lindenmayer et al. 2022). These favorable climatic conditions facilitate taller mature forests dominated by Eucalyptus species higher in foliar nitrogen (preferred for feeding) and abundant hollow-bearing trees (Smith et al. 2007; Koch et al. 2008b; Kearney et al. 2010; Youngentob et al. 2011; Smith and Smith 2020; Wagner et al. 2020, 2021a). As such, large trees likely to contain hollows were identified to be essential for SGG occurrence in wet eucalypt forests (Lindenmayer et al. 1990a, 2022), with an average of four hollow-bearing trees (HBTs) per hectare reported as a threshold for occupancy of the species (Lindenmayer et al. 1993b; Smith et al. 2007).
The development of hollows in eucalypts is dependent on tree dimensions, form, age and mechanical disturbance (Lindenmayer et al. 1991a, 1993a). Large diameters, shorter heights, and/or irregular crowns are important predictors for estimating hollow occurrence and abundance in individual trees in temperate Australian forests (Bennett et al. 1994; Gibbons et al. 2000; Fox et al. 2009). The indirect nature of hollow formation is dependent on the aging and senescing of trees and the absence of cavity excavating fauna means that hollow formation can require long time-frames and therefore mainly occurs in large, mature trees that have been exposed to different sources of damage or decay (e.g., fire or fungal and termine attacks, Fox et al. 2008; Lindenmayer et al. 2017). Formation times of a minimum of 100–200 years for different temperate species of Eucalyptus have been reported (Inions et al. 1989; Gibbons et al. 2000; Gibbons and Lindenmayer 2002). Climate also plays a role in these processes and can foster faster hollow formation in drier parts of the landscape or in certain forest types (Bennett et al. 1994; Gibbons and Lindenmayer 2002; Haslem et al. 2012; Woolley et al. 2018). Wetter climates promote the development of productive soils that can support large mature trees, more likely to contain multiple large hollows (Lindenmayer et al. 1990a, 1991a; Bennett et al. 1994; Campbell-Jones et al. 2022). These factors may lead to different patterns in hollow availability across landscapes due to the influences of climate, soils, topography, and disturbances on forest structure and composition (Turner and Gardner 2015; Lindenmayer et al. 2016).
Disturbances to landscapes, especially at the scale of 21st century megafires, are assumed to cause a major reduction in the number trees with hollows, driven by the collapse of large trees from fire damage, and therefore loss of nesting space for arboreal fauna (Inions et al. 1989; Lindenmayer and Wood 2010; Parnaby et al. 2010; Banks et al. 2011; Manning et al. 2013; Lefoe et al. 2022; Lindenmayer et al. 2022). Due to this process, surviving HBTs are regarded as a key biological legacy (Pulsford et al. 2016), important for supporting hollow-dependent species as their populations recover in disturbed forests (Mazurek and Zielinski 2004; Rainsford et al. 2020). Such legacy trees are highly vulnerable to additional, short-interval disturbance such as fires, further reducing hollow availability and therefore nesting space for arboreal fauna (Banks et al. 2011; Lindenmayer et al. 2016; Salmona et al. 2018; Gibbons et al. 2024). The vulnerability of forests to loss of HBTs in subsequent fires is shaped by patterns in fire severity. Areas impacted by different fire intensities will experience different patterns of species and size-specific tree mortality (Trouvé et al. 2020, 2021). Lower severity fires may result in tree damage rather than mortality, which may facilitate hollow development (Inions et al. 1989; Gibbons et al. 2000; Adkins 2006; Salmona et al. 2018). This process can facilitate positive biological legacy effects and reduce population recovery times of hollow-dependant fauna after disturbance.
One of the main areas of habitat for the endangered SGG and other hollow-dependent arboreal fauna are the mixed eucalypt forests of south-eastern Australia. These areas were severely affected by the 2019/2020 bushfires, which burnt large areas of the SGG’s range (Ward et al. 2020). Climate has been identified as a major landscape-scale driver of SGG occurrence in the region (Wagner et al. 2020) while nutritional quality of Eucalyptus foliage is an important factor in determining their occurrence at the stand scale (Youngentob et al. 2011; Wagner et al. 2021b). Tree hollows play a critical role in determining the likelihood of SGG occupancy at fine scales (Lindenmayer et al. 1993b; Nelson et al. 1996; Gibbons et al. 2000; Eyre 2006), in areas that have sufficient foraging resources and climatic suitability (Wagner et al. 2021b). Hollows are therefore a limiting factor that shapes SGG occupancy in space and time. In landscapes impacted by changing fire regimes (Collins et al. 2022), understanding how fire shapes this critical resource is essential for determining the ability of the region’s forests to provide habitat for the SGG, and other arboreal fauna, into the future.
In this study, we aimed to quantify the major determinants of hollow occurrence in temperate, mature mixed-species Eucalyptus forests, and examine the effects the unprecedented 2019/2020 fires have had on hollow availability. We hypothesize that the occurrence of hollows in the region is related to tree size, shape, and age as well as forest structure and disturbance history (Gibbons et al. 2000; Fox et al. 2008; Koch et al. 2008b). To address these research objectives, we test the ability of hollow-bearing trees to persist as biological legacies in the study region and if the likelihood of hollow occurrence in trees changes due to fire occurrence and severity. Quantifying major determinants of habitat availability across scales and how they are affected by disturbance is crucial for assessing and conserving habitat in a forest landscape impacted by an unprecedented mega-fire event (Ward et al. 2020; Geary et al. 2021).
Methods
We used flora and fauna surveys, soil chemistry, spatial data, and linear models to test independent covariates to estimate variability in hollow availability before and after the 2019/2020 Black Summer bushfires, as well as the impacts of these fires on tree hollow occurrence and abundance and arboreal fauna richness with a focus on the southern greater glider (SGG).
Study area
The study was conducted in the mixed Eucalyptus forest landscape of East Gippsland, Victoria, Australia (Department of Agriculture and Water Resources 2018). These forests occupy ~1.2 million hectares and range from dry open lowland forest along the coast into the mid-hills (5–400 m a.s.l) to closed tall forest at high elevation (~1000–1300 m a.s.l.). The most common eucalypt species are Eucalyptus siebieri in the lowlands, E. globoidea at mid-elevation, and E. viminalis, E. cypellocarpa, and E. denticulata at high elevations. E. obliqua is the most common species across the entire elevation gradient, with E. croajingolensis occurring frequently from mid to high elevation (Opie et al. 1990; Department of Conservation and Natural Resources 1995; Sebire and Fagg 2009; Wagner 2021; Wagner et al. 2021b). Average annual rainfall and temperature in the study region ranges from 648 to 1178 mm (average for 1981–2020, Stewart et al. 2020b) and 6–15.8 °C (average for 1981–2020, Stewart and Nitschke 2017b), respectively. Across the study area and before the megafire event in 2019/2020 (November–February), SGGs were most abundant at elevations > 700 m in cool and wet forests, where their specific thermoregulatory requirements are met (Henry 1984; Bennett et al. 1991; van der Ree et al. 2004; Wagner et al. 2020). Forty-seven percent of the study area burned at high severity in the 2019/2020 Black Summer bushfires, while 42% burned at low to medium severity. Only ~11% of the study area remained undisturbed following the fires (DAWE 2021).
Plot design
Our pre-fire survey plot network was designed to include a range of dry and wet forest types with Eucalyptus species that are utilized by SGGs as a foraging resource, as well as species that they are known to avoid (Kavanagh and Lambert 1990; Comport et al. 1996; Cunningham et al. 2004; Wagner et al. 2021b). Pre-fire study sites were established across the topographic gradient of the known distribution of SGGs in the region between 2018 and early 2019. To ensure only mature and undisturbed forests were captured in the study design, we stratified the landscape to eliminate forests that had been disturbed by fire or logging in the previous 50 years or more using publicly available spatial layers on forest fire and timber harvesting history from the Victorian State government (https://www.data.vic.gov.au/). Plots were established at least 5 km away from any previous high severity fires or previous logging areas to avoid potential impacts of nearby disturbances on undisturbed forests. We established 30 plots in three elevation bands: lowlands (0–420 m), mid-hills (420–840 m), and high elevation (840–1260 m a.s.l). Within each elevation band, we established ten plots of varying elevation, aspect, and slope. In each band, four plots were located in flat or gully locations, four in mid-slope positions (north- and south-facing) and two on the ridge, close to the maximum elevation of the respective band (Wagner et al. 2021b, Fig. 1, Appendix S1: Fig S1). In these 30 pre-fire study sites ranged in elevation from 32 to 1185 m a.s.l., covering slopes from 1 to 55% and aspects from 26 to 355°.
In late 2019 and early 2020, the Black Summer bushfires burned across this plot network, affecting 18/30 plots at varying fire severity. The fires provided a unique opportunity to study changes in forest structure and nesting space for arboreal fauna, specifically for the now endangered SGG. We re-surveyed at all burnt pre-fire sites (n = 18, and three unburnt sites that were initially mapped as having burnt) between September 2021 and May 2022 and established 19 additional sites in areas of varying fire severity, where collaborators from the Arthur Rylah Institute for Environmental Research (Heidelberg, Victoria) carried out surveys for SGGs between March and July 2020, immediately after the fires. The total number of pre- and post-fire sites was therefore 49; the post-fire site pool was 40 sites assessed for forest structure. Due to the heterogenous nature of fire impacts (i.e., fine-scale variation in fire intensity, see Price and Bradstock 2012; Tiribelli et al. 2019) we used a transect-based survey design for our post-fire surveys. At each post-fire site, we established a 500-m transect. The entire length of the transect was used for spotlighting surveys (see details below), while the 200 m of transect were used to establish a set of ten survey plots to assess forest structure using a point-centered quarter method (Appendix S1: Fig. S2). Our additional post-fire sites (n = 19) ranged in elevation from 681 to 1207 m a.s.l. Slopes of the new sites ranged from 3 to 24% and aspects from 0 to 350°.

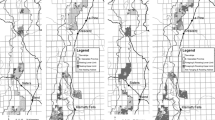
Map of the study region in East Gippsland, eastern Victoria, Australia, showing elevation gradient, pre-fire plot positions (n = 30) and fire severity at all sites (n = 49). All pre-fire sites surrounded with a square indicating a fire severity were resampled (n = 18). Any squares color-coded by fire severity without a pre-fire point contained within, are new post-fire sites (n = 19)
Assessing forest structure
In the pre-fire surveys, each plot center location was recorded using a handheld GPS. At the center of each survey plot, we recorded site attributes (elevation, slope, aspect, crown cover and basal area of dominant tree species), then established a variable radius plot using a Kramer’s dendrometer with a basal area factor (BAF) of four. This ensured the sampling would capture large, mature, and dominant trees that form the canopy, and which are the preferred nesting and feeding habitat by SGGs. For each tree in the variable radius plot sample, we recorded the species, diameter at breast height (DBH), height, crown width (north-south and east-west axis), foliage cover, vigor (health status in categories), age class, structural layer the tree belonged to, and the occurrence, number, size, and type of tree hollows using binoculars (for details on variables, see Appendix S2: Table S1).
For post-fire surveys, we used a point-centered-quarter plot design (PCQM, see, e.g., Mitchell 2010) to better capture the impacts of locally variable fire severity on forest structure. Along 200 m of the 500-m spotlighting transect, we established a survey plot every 20 m, starting at 10 m to avoid edge effects from the road (Appendix S1: Fig. S2). If a site was established where pre-fire measurements were conducted, the transect was laid out to lead through the pre-fire plot. Individual plot locations were marked during transect setup and reflective flagging tape was tied to vegetation on eye level to guide later spotlighting surveys at night. At the center of each 200-m PCQM transect, we recorded the location using a handheld GPS and noted elevation (using the GPS), slope, and aspect (using compass and clinometer) of the site, as well as average canopy crown cover (using a crown densiometer). At each PCQM plot, we located the closest canopy tree in the first plot quarter, which is the quarter bordered by the transect on the left and the second, perpendicular line on the right. Plot quarters 2, 3 and 4 were in clockwise direction from quarter 1. In each quarter, we measured the distance and bearing to the closest tree and its diameter. Trees had to have a diameter at breast height (DBH at 1.3 m) greater than 5 cm to be included. We measured height and crown widths on the north-south and east-west axis from three individuals per tree species along the PCQM transect. We assessed hollow occurrence, number and size class and noted the structural layer the tree belonged to, as well as age class and if it was reproductive or not. A total of 40 trees per PCQM transect were measured (four per plot). Dead trees were measured separately from alive trees if they were closer than the closest alive tree. Dead trees were assessed for DBH, height and hollows only.
Soil sample collection
Soil samples were collected to a depth of 10 cm from 30 locations within each pre-fire survey plot, and from one sample per PCQM plot during post-fire surveys (ten locations) using a tube corer. One composite sample per plot was sealed in an individual air-tight bag, cooled, and transported to the laboratory for drying and subsequent chemical analyses.
Wildlife surveys
To determine occurrence of SGGs and other nocturnal fauna, spotlighting surveys on 1-km transects on forest roads proximate to the plot center were carried out at pre-fire sites and on 500-m transects (including the 200-m PCQM transect) at post-fire sites. The survey method was altered for post-fire sites to take recent research into account and align with state-level recommendations on survey design and effort for arboreal fauna (Chick et al. 2020; Cripps et al. 2021). The post-fire survey design was aligned with another project led by the Arthur Rylah Institute, carried out in different parts of the 2019/2020 fire footprint to assess post-fire SGG abundance in Victoria (DELWP 2022). As the different sampling efforts of either survey method may lead to variability in detectability or abundance estimates, we used pre- and post-fire occurrence (i.e., presence/absence of observed species, including the targeted SGG) and richness (no. of observed species) for this study. All surveys were carried out using double observer distance sampling to increase likelihood of detection (Wintle et al. 2005; Cripps et al. 2021) with an average pace of 10 min per 100 m and two observers walking 10 min apart. Observers scanned tree canopies for eyeshine reflection using strong torchlights. Any detected animal’s location was recorded using a handheld GPS alongside its distance from the transect and bearing to the animal. Further observation data of the tree species on which the animal was observed, height on the tree, animal behavior, fur color and time of observation were noted. Any spotlighting transects with one or more detected SGGs were considered occupied for the purpose of this study. During post-fire spotlighting surveys, we resurveyed at all 30 pre-fire site locations, including those that had not burnt in 2019/2020. One post-fire site could not be surveyed due to access issues and adverse weather conditions (leading to n = 48 spotlighting surveys).
Sample and data processing, variable selection, and modeling
Chemical analyses
Soil samples were dried at room temperature, sieved to pass a 2-mm screen, composited by plot (pre-fire), or transect (post-fire), and analyzed for soil nutrients. The following soil chemical properties were derived: soil pH (CaCl2), nitrate nitrogen (NO3 in mg/g), phosphorous (P in mg/g), potassium (K in mg/g), total nitrogen (soil N in percent) and total carbon (C in percent). Soil N and C were derived using dry combustion, while other elements were determined colorimetrically or via spectroscopy following standard laboratory procedures.
Spatial variables
In addition to structural variables, fauna survey records, and soil chemical values, we collated spatial variables related to climate and topography as covariates for hollow occurrence modeling. These had previously been identified as important for determining SGG habitat suitability at the landscape scale (see Wagner et al. 2020) and include the Annual Heat Moisture Index (AHMI), number of days with condensation events (1981–2020), the Terrain Wetness Index (TWI), and Normalized Difference Vegetation Index (NDVI) along with further topographic variables such as slope and aspect in raster format (Table 1). AHMI and condensation days were calculated using daily weather data developed by Stewart and Nitschke (2017b) from MODIS satellite imagery at a 250-m grid resolution. Temperature and precipitation were extracted using site locations and averaged for the ~40-year period 1981–2020 (span of available data) to derive AHMI as a measure of site-level aridity (Stewart and Nitschke 2017a, 2018; Stewart et al. 2020b). Daily condensation as a measure of water availability to foraging folivores was calculated using saturation vapor pressure at minimum temperature of the day and vapor pressure at 9:00 AM (Stewart and Nitschke 2018; Stewart et al. 2020a; Wagner et al. 2020) and was tallied for each site. Topographic variables (slope, aspect) were sourced from the 30-m resolution digital elevation model from the shuttle radar topography mission (SRTM, Farr et al. 2007) and TWI subsequently calculated following Beven and Kirkby (1979). NDVI at survey date was sourced from Landsat imagery at a 30-m resolution through the USGS data portal Glovis (Landsat Collection 2 surface reflectance products—glovis.usgs.gov). For a comparison between pre- and post-fire hollow occurrence and the effects of the 2019/2020 bushfire on nesting space and arboreal species richness and SGG occurrence, we included modeled fire severity from Victorian government data (McCarthy et al. 2017). Given the lowest resolution was 250 m (MODIS-derived climate variables), we aggregated higher resolution Landsat data to the same resolution by cell value averaging. The plot- and transect center location were used to extract raster cell values. In cases where a location fell near or on the border between multiple cells, cell values were averaged for extraction. All spatial data processing and value extraction was carried out in R (R Core Development Team 2022) with the packages sf and raster (Pebesma 2018; Hijmans 2019).
Quantifying the effects of tree size, forest structure and fire on hollow occurrence and arboreal species observed
To enable a robust comparison of hollow availability between pre- and post-fire forest survey methods we calculated the proportion of hollow-bearing trees (HBTs) across all trees per site. We also considered large hollow-bearing trees (LHBTs) with diameters ≥ 100 cm separately, as these are more likely to have developed larger hollows, fit for the nesting requirements of SGGs (Lindenmayer et al. 1991c; Gibbons et al. 2002; Campbell-Jones et al. 2022). We analyzed pairwise differences between hollow abundance, tree characteristics and forest structure between elevational bands (pre- and post-fire), sites occupied or unoccupied by SGG (pre- and post-fire) and fire severity (post-fire).
For structural data (e.g., hollow-bearing tree diameters, heights and crown area) we log-transformed the response variable to ensure homogeneity of variance, as raw data was right-skewed, and then evaluated relation using F and p statistics from one- and two-way analysis of variance (ANOVA) and Tukey Honest Significant Differences (HSD) tests. Data distribution was assessed visually using histograms. For testing differences between proportions of hollow-bearing trees, we used generalized linear models (GLMs) with a quasibinomial error distribution and assessed p statistic, estimates and confidence intervals. We tested the influence of fire severity on hollow occurrence and abundance with generalized linear mixed models (GLMMs) on tree-level data and evaluated model estimates to determine effects and direction of relationships. We integrated a random effect for the interaction between site (transect) and plot to account for the nested post-fire sampling design (PCQM). Here, we assumed that the effect of the random intercept for plots is nested within the random intercept for transects. This accounts for cluster effects where observations (trees) within the same cluster (transect) are more similar to each other than to other clusters. A binomial error distribution was used for occurrence (presence or absence data) and Poisson distribution for abundance (count data). The Poisson distribution was determined to be a better fit, as tests for overdispersion and likelihood ratio were non-significant. We also used binomial GLMMs to assess the probability of hollow occurrence as a function of tree diameter measured during pre- and post-fire surveys, to test if the probability of hollow occurrence by tree size differed before and after the fires. To account for the sampling methods, we used plot as a random effect for pre-fire data, and the interaction between sites (transect) and plot for post-fire data. These models were then used to predict the probability of hollow occurrence to the range of tree diameters observed before- and after the fires. Finally, we used GLMMs to test for effects of the proportion of HBTS or LHBTS and elevation (pre- and post-fire), as well as fires severity (post-fire only) on the occurrence of SGGs and the number of arboreal species observed during spotlighting surveys. Given differences in species detections per observer (Appendix S2: Table S2) and to account for site heterogeneity affecting detectability of spotlighting surveys, we included the interaction between observer and site as a random effect (Kéry et al. 2009; Camp et al. 2023).
Modeling hollow occurrence
We used a multi-step modeling procedure to determine meaningful predictors for tree-level hollow occurrence (presence or absence of hollows) before and after the fires and created a combined model for drivers of hollow occurrence using a combined dataset of pre- and post-fire trees. We first split all available modeling variables into structural, site level (including disturbances), and nutrient groups (Table 1). Within these groups we first tested for intercorrelation between variables. We reduced the number of initial predictor variables in each group by removing pairs with Pearson’s correlation coefficient r ≥ |0.8|. All variable candidates were then standardized to a mean of zero and standard deviation of one. We evaluated alternative binomial GLMs with all uncorrelated variables per group as predictors and hollow occurrence as the response variable. Using every possible variable combination in separate models, we identified the best variable combinations based on the lowest Akaike Information Criterion (AIC) values and ΔAIC ≤ 2 (Anderson and Burnham 2004; Appendix S3). The predictor variables from the best models of each variable group were then used in combined models of pre- and post-fire hollow occurrence that drew the best performing variables from each group. Best models were determined using AIC, model R2, and best variables identified by their variable significance (p). Finally, we pooled pre- and post-fire trees into a combined dataset to derive important predictors of hollow occurrence across the landscape using GLMMs. We used a factorial variable indicating the sampling campaign as a random effect to account for differences in sampling methods. Further, we used a factorial variable indicating whether a tree was burned or unburnt when sampled, to test if the disturbance played role in hollow occurrence when pooled with other important predictors. We included all important variables from both final models. All statistical analyses were conducted in R (R Core Development Team 2022) with the packages MuMIn, lme4, and caret (Bates et al. 2015; Barton 2019; Wing et al. 2019).
Results
During pre-fire surveys, we measured a total of 463 individual eucalypt trees from 17 species; 206 trees had at least one hollow. Common and dominant canopy species included Eucalyptus consideniana, E. globoidea, and E. sieberi at low elevation, E. obliqua, E. cypellocarpa, and E. fastigata in the mid-elevation sites, and E. corajingolensis and E. denticulata at high elevation. We recorded 239 individual animal observations during night-time spotlighting surveys across all sites, including 44 southern greater gliders (SGGs), which were found at nine sites. SGGs occurred with the highest frequency and density in high elevation sites, where they were found at six of the ten sites surveyed (Appendix S2: Table S3).
Across all 40 post-fire sites, we measured 1529 individual Eucalyptus trees from 19 species; 427 trees had at least one hollow. Out of the 40 post-fire survey sites, four were found to be unburnt even though fire severity mapping suggested otherwise, 17 sites burned at low to medium severity, and 19 burned at high severity (high canopy scorch or burn). Twelve sites from the pre-fire site pool were unburnt, two burned at low severity and 16 at high severity (see Appendix S2: Table S4 for details on all post-fire sites). We found that the diameter distributions from both survey campaigns were similar (DBH ranges pre-fire: 8–303 cm, post-fire: 5–262 cm, Fig. 2), despite different survey methodologies being used during pre- and post-fire surveys.
We recorded 228 animal observations during post-fire spotlighting surveys, including 62 SGGs. They were observed at 19/48 sites including at 15/40 of the post-fire sites. Post-fire, SGGs were predominantly observed at sites that had burnt at low to medium severity (nine sites). We observed SGGs at four sites affected by canopy scorch (and none at sites that were affected by canopy consumption - the highest fire severity class). During post-fire surveys, we observed SGGs at 7/30 sites from the initial pre-fire sampled sites. Five sites were unburnt and two burned at high severity. The two sites that had SGG observations before, but not after the fire, were burnt at high severity (Appendix S2: Table S2).
Pre-fire, soil nitrogen (N) ranged from 0.08 to 0.76% and soil carbon (C) from 2.8 to 14.3%. Post-fire soil chemistry values for N and C ranged from 0.08 to 0.97% and 2.32 to 16.23%, respectively. Pre-fire soil phosphorus (P) concentrations were between 1 and 20 mg/g and potassium (K) between 46 and 452 mg/g, while post-fire nutrient concentrations were 3–78 mg/g for P and 38–647 mg/g for K. We did not find any significant differences in soil chemistry with fire severity in post-fire sites or between pre- and post-fire measures at the 18 sites survey pre and post fire. The concentrations of soil chemical elements however increased significantly with elevation both pre- and post-fire (e.g., F = 41.46, p < 0.0001 for N pre-fire).
Tree hollow incidence and characteristics of hollow-bearing trees
Before the fires, the average proportion of hollow-bearing trees was 29% (± 12%) at low elevation, 38% (± 15%) at mid- and 47% (± 22%) at high elevation. After the fires we observed lower proportions across the elevational gradient and along a gradient of fire severity (Table 2). There was a significant difference between the proportions of hollow bearing trees with elevation before the fires (F = 2.8, p = 0.0025) as well as after the fires (F = 10.36, p = 0.03) and a significant difference in proportion of hollow bearing trees with fire severity (F = 2.7, p = 0.002).
Across all measured pre-fire HBTs, the average diameter at breast height (DBH) was 129 cm (± 65.3 cm), height was 33.8 m (± 13 m), and crown area was 161 m2 (± 115.2 m2). Mean HBT sizes varied with elevation, with the largest HBTs occurring at high elevation (µ HBT DBH = 147 cm ± 60 cm). DBH of HBTs differed significantly between elevation bands (F = 33.15, p < 0.0001), with the largest difference between mid- and low elevation (Tukey HSD test p < 0.0001). Pre-fire HBT tree height and crown area differed significantly between elevation bands (F = 22.49, p < 0.0001 and F = 43.42, p < 0.0001, respectively; Appendix S1: Fig. S3).
Post-fire HBTs were smaller in DBH (84 cm ± 40.8 cm), height (32.5 m ± 11 m), and crown area (105 m2 ± 106.2 m2). HBT sizes in fire affected sites varied with elevation and it was observed that there were fewer large HBTs and more HBTs with smaller diameters across the elevational gradient, compared to pre-fire sites that had burned (Fig. 2). There was a significant difference in post-fire HBT diameter between elevation bands (F = 12.96, p < 0.0001), as well as tree height (F = 9.38, p < 0.0001), and crown area (F = 7.5, p < 0.0001). Post-fire HBT sizes varied with fire severity, with the smallest HBT diameters recorded in the highest fire severity class (µ HBT DBH = 64.8 cm ± 16.7 cm, Appendix S1: Fig. S4). These results are however biased by the fact that low elevation sites had trees with smaller DBHs and were predominantly affected by fires burning at the highest severity (Fig. 1 and Appendix S2: Table S4) and that, due to the extent of the 2019/2020 bushfires, we sampled numerically more mid- and high elevation sites during post-fire surveys (Fig. 1 and Appendix S1: Fig. S5). HBT DBH (F = 8.57, p < 0.0001), height (F = 4.416, p = 0.002), and crown area (F = 6.52, p < 0.0001) all differed significantly with fire severity, with increasing fire severity reducing these attributes. Differences in crown area with fire severity were driven by an abrupt change between low to medium and high fire severity classes (e.g., Tukey HSD test p = 0.003), while the significant difference in tree height (highest green branch in fire affected trees) was driven by differences between high canopy scorch and canopy burn and canopy scorch and low severity burns (Tukey HSD tests p = 0.05 and p = 0.001, respectively).

Observed diameter distributions separated by hollow-bearing and non-hollow-bearing trees from burnt sites (n = 18) distinguished by elevational band measured before (A) and after the 2019/2020 bushfires (B)
Fire effects on hollow occurrence and abundance
For tree-level post-fire survey data, we found significant effects of fire severity on hollow occurrence (i.e., probability of a tree having a hollow) and hollow abundance (number of hollows per tree) using generalized linear mixed models (GLMMs). The highest fire severities had a significant negative impact on hollow occurrence, but low and medium severity classes did not (Fig. 3A). Hollow abundance was significantly affected by fire severity with varying effects. Low to medium fire severity had a positive effect on hollow abundance, while both high severity classes resulted in a reduction in hollow abundance (Fig. 3B).

Standardized parameter estimates and confidence intervals for GLMMs of the effect of fire severity on A HBT occurrence and B hollow abundance (i.e., number of hollows) at the tree level. Strong black lines indicate 90%, light lines 95% confidence intervals
When testing if tree diameter distributions (all trees combined and HBTs only) differed before and after the fires, we found that both were significantly larger during pre-fire surveys (estimate = 38.69 ± 2.2 and estimate = 45.1 ± 4.3, p < 0.0001, respectively), indicating that post-fire sites had fewer large trees and large HBTs and that HBTs across sites were smaller after the fires (Fig. 2). Using GLMMs for hollow occurrence versus tree DBH which took the hierarchical sampling design of pre- and post-fire tree-level survey data into account, we found that the probability of hollow occurrence by tree diameter increased by up to 40% for trees between 100 and 150 cm DBH (Fig. 4A). Combined pre- and post-fire models both indicated an increase in likelihood of hollow occurrence with DBH according to model estimates (0.03 ± 0.003, p < 0.0001 and 0.07 ± 0.006, p < 0.001, respectively). The 100% likelihood of hollow occurrence shifted from ~300 cm to ~150 cm in DBH after the fires (Fig. 4A). Changes in the probability of hollow occurrence by tree size varied across the sampled elevation gradient and the impacts of fire on the probability of hollow occurrence based on DBH increased with elevation band. The greatest increase in probability was observed for trees with a DBH ~75 cm in the lowlands, ~115 cm at mid-, and ~105 cm at high elevation. There was an overall trend for a higher probability of hollow occurrence in trees with DBH > 50 cm (except at mid elevation, Fig. 4B).

Probability of hollow occurrence by tree diameter before, and after the 2019/2020 bushfires (A) and change in probability for tree diameters between 0 and 300 cm after the fires (B). The 95% confidence interval for both combined mixed models was [0.02, 0.03]
Models of tree-level hollow occurrence
Pre- and post-fire hollow occurrence modeling identified that tree size, tree canopy class, soil nutrition and soil moisture were important predictors of hollow occurrence across the landscape in both datasets (Appendix S3).
The best pre-fire model indicated that hollow occurrence was best explained by tree volume, age class, structural layer, TWI, and soil nitrogen (N) in pre-fire mature forests (Appendix S1: Fig. S6A-E, Appendix S2: Table S5). Post-fire, crown area and soil pH emerged as important predictors a (Appendix S2: Table S5). The best post-fire model of hollow occurrence contained age class, crown area, structural layer, soil pH and total nitrogen (Appendix S1: Fig. S6F-J, Appendix S2: Table S5).
The combined tree dataset model (data from both survey campaigns: n = 308 burnt trees and n = 489 unburnt trees) identified that age class, structural layer, volume, and TWI were highly significant, while crown area was also significant but with a small effect size (Table 3). The strongest effects based on standardized parameter estimates were observed for soil nitrogen, age class and TWI. Structural layer, soil pH, and whether a tree had burned or not were not significant but improved overall model performance (Table 3).
Arboreal fauna occurrence in relation to hollow occurrence and fire
Hollow-dependent arboreal fauna observed during pre- and post-fire spotlighting surveys, in addition to the SGG, included common- and mountain brush-tailed possums (Trichosurus vulpecula and T. cunninghami), eastern ring-tailed possums (Pseudocheirus peregrinus), Krefft’s and yellow-bellied gliders (Petaurus notatus and P. australis), as well as southern boobook, powerful, and sooty owls (Ninox boobook, N. strenua, and Tyto tenebricosa, respectively). The maximum number of arboreal species observed was seven during pre-fire surveys (at a low elevation site) and five during post-fire surveys (at a high elevation site burnt at low severity, Appendix S2: Tables S2 and S3).
Both pre- and post-fire, a higher proportion of HBTs per site had significant positive effects on SGG occurrence (p < 0.05 pre-fire and p < 0.01 post-fire). The proportion of LHBTs did not significantly differ during pre-fire surveys but had a significant positive effect on SGG occurrence post-fire (p < 0.01). No significant effects of proportion of HBTs or LHBTs on the number of hollow-dependent arboreal species observed were detected. SGG occurrence increased with elevation in both pre- and post-fire surveys (both p < 0.01) and was negatively affected by increasing fire severity (p = 0.05). A positive effect of elevation on the number of arboreal species was observed pre-fire (p = 0.05); however, there was no significant effect of elevation or fire severity on the number of arboreal species observed post- fire.
Discussion
Across the study landscape, tree-level hollow occurrence, before and after the fires, was significantly related to tree size, topographic wetness, and soil nitrogen. We found tree size to be a strong predictor for hollow occurrence across all models tested. Tree size, shape, and diameter have previously been identified as good predictors of hollow occurrence and abundance in Eucalyptus forests due to the monotonic relationship between tree size and the occurrence of hollows (Lindenmayer et al. 1990a, 1993a; Bennett et al. 1994). Of significance is the finding that 2019/2020 megafire has altered the relationship between hollow occurrence and tree size in the study area. In post-fire forests, we found that the probability of a tree having a hollow increased in smaller trees. The 75% threshold for occurrence dropped from 150 cm diameter at breast height (DBH) to 100 cm DBH across all post-fire plots. This shift was greatest in trees located at higher elevation (Fig. 4). There was an ordered increase in the magnitude of the shift in the DBH x hollow occurrence relationship with elevation. HBTs at low elevation exhibited the smallest shift in the relationship followed by mid-elevation trees and then high elevation trees. The greater shift in hollow occurrence by DBH with increasing elevation is likely due to the impact of the aridity gradient associated with elevation (Wagner et al. 2020). There is a significant relationship between tree diameter growth in eucalypts and aridity (Bowman et al. 2014a), which means that trees growing in the more arid low elevation areas of the landscape will grow slower than in the cooler and wetter sites at higher elevation. Hollow-bearing trees (HBTs) of the same age will therefore have different diameters. This is supported by Koch et al. (2008b), who found that hollow formation in dry and wet eucalypts occurred over same time scale (~100–140 years) while productivity differences meant that DBHs of HBTs in wet forest were larger than in dry forest. The influence of productivity on the tree size versus age dynamic is supported by the impact of soil nitrogen on hollow occurrence in our hollow occurrence models (Table 3). Trees growing on more nitrogen poor sites will have a higher probability of having a hollow for a given size as they are likely to be older than a tree of similar size on nitrogen rich soils. This is supported by Forrester et al. (2007) who found that eucalypt growth increased with increasing nitrogen availability. Fire appears to facilitate the formation of hollows in larger but younger trees along the productivity gradient of the study area. This suggests that fires interact with site productivity and tree age to influence HBT occurrence in forest landscapes over time and space.
When testing important variables influencing hollow occurrence pre- and post-fire, TWI was lost as a significant predictor post-fire (Appendix S2: Table S5) indicating that the 2019/2020 fires may have overrode the influence of topography on hollow occurrence in trees. The 2019/2020 fires were the most severe on record and were preceded by severe drought (Nolan et al. 2020). The interaction between drought and fire severity were found to reduce the probability of topographic refugia occurring in areas burnt under extreme fire weather conditions by Collins et al. (2019). Where topographic mediation of fire spread is lost, large HBTs growing in wetter areas of the landscape are subject to crown consumption and scorch leading to a reduction in crown area. In both the post-fire hollow occurrence and combined models, the loss of crown area had a significant negative impact on HBT occurrence. Crown area in post-fire stands represents the impacts of fire intensity experienced and the variable is therefore likely capturing the impacts of fire severity that was significant in the group variable selection stage but not in the final model (see Appendix S3).
The varying effects of fire severity on hollow occurrence and abundance are an important finding for understanding how fires shape the patterns of HBTs and the processes that influence their demography. A negative effect was detected in areas impacted by the highest severity fire class which had a lower hollow proportion of HBTs and trees had a lower probability of hollow occurrence. Conversely, in areas that experienced lower severity fires we observed an increase in hollow abundance (Fig. 3). This potential positive effect of fire on the development of tree hollows in eucalypts has been reported previously (McLean et al. 2015) and the mechanisms behind the observed hollow development from mechanical damage (e.g., directly through fire or falling limbs and trees as a result of fire damage to nearby trees) are well documented (Lindenmayer et al. 1991a, 1993a; Fox et al. 2008). The number of hollow-bearing trees, however, did not differ significantly across areas impacted by different fire severities and unburnt forests. This may indicate that while high severity fires have a negative influence on the number of tree hollows, a reduction in the number of HBTs may not occur. It suggests that nesting space maybe reduced at the tree-scale in severely burnt forests; however, at the stand-scale, HBTs may persist as biological legacies that should support the recovery of arboreal marsupials and other hollow-dependent fauna. These surviving, potentially damaged, HBTs are also likely to form new hollows faster than would develop in non-HBTs, as they are already at an age where the processes of internal decay are advanced (Inions et al. 1989).
Our findings that fire damage may substitute the loss of mature, old, and potentially senescing HBTs by fostering hollow formation in younger or smaller trees has important implications for future hollow availability and therefore post-disturbance development of arboreal fauna habitat. The negative effects of disturbances on nesting space and therefore loss of a crucial habitat resource for arboreal fauna were important considerations made after the 2019/20 Black Summer bushfires (Ward et al. 2020; Geary et al. 2021). Without denning and nesting space, a population of hollow-dependent fauna, such as the southern greater glider (SGG), cannot persist or re-establish (Goldingay 2011), even if an area is otherwise suitable through the availability of favorable foraging resources (Youngentob et al. 2011; Wagner et al. 2021b) or climate (Wagner et al. 2020). This is evidenced in our findings that SGG occurrence was significantly linked to HBT and LHBT occurrence before and after the fires. During our post-fire surveys, we did not observe SGGs in forests burnt at highest fire severity; however, the 2019/20 fires created a mosaic of fire severities across eastern Victoria. Forests that remained unburnt or burned at lower severities now border forests that had their canopies burned or consumed (Appendix S1: Fig. S5), and local SGG populations lost. Our findings suggest that SGGs were able to survive in forests affected by low to medium severity fires (eight out of 13 low to medium fire severity sites had SGGs present). Evidence, presented here, that fire damage may have led to hollow formation in smaller trees in forests affected by low to moderate fire severity, shows that these forests may still offer the crucial nesting potential needed for arboreal folivores like the SGG to recolonize these impacted habitats from less- or undisturbed surrounding areas as the forests recover (Berry et al. 2015; Campbell-Jones et al. 2022). Even if the newly developed hollows are initially too small to support large arboreal species (Hofman et al. 2022), they form an important stepping stone in the process of redeveloping suitable habitat for species such as SGGs, as the disturbed forests recover and trees and hollows mature. Additionally, these fire-affected forests already support a number of other (smaller) hollow-dependent fauna, which we observed more frequently and homogenously across elevations and forest types both before and after the fires and which is supported by our findings of a non-significant effect of fire or hollow occurrence and on the total number of arboreal species observed (Appendix S2: Table S3).
With hollow-bearing trees persisting across the post-fire landscape, foliar chemistry and eucalypt species composition of recovering forest will likely play a major role in the recolonisation potential of severely burned forests for SGGs at the stand level (Au et al. 2019). Climatic conditions continue to be important in shaping SGG habitat suitability at the landscape scale and important refugia have been identified and remained partially undisturbed during the 2019/2020 megafires (Wagner et al. 2020). Future research should therefore investigate the foliage quality of recovering forests, to determine if (or how soon after fire disturbances) they are suitable for foraging by hollow-dependent Eucalyptus folivores.
Limitations and future research
The extent and severity of the 2019/2020 Black Summer bushfires came as a surprise to many researchers and experts (Collins et al. 2021; Legge et al. 2021), but offered a unique opportunity to study the effects of large and severe fires on forest ecosystems at a landscape scale and attempt to compare with pre-fire conditions (Ward et al. 2020). While many studies have been carried out before the fires, not all of them were set up to be repeated in case a large-scale disturbance occurred, including the study at hand. Further, funding and monitoring requirements had changed over time and, alternative study designs were required to survey for a broader set of variables to quantify the fire impacts, or post-fire surveys had to align with other programs to enhance research and conservation outcomes. As such, our two survey campaigns did not provide an ideal dataset to directly compare impacts and we had to resort to a more descriptive or generalized approach for many of our analyses. The extent and spatial configuration of the 2019/2020 fires (Appendix S1: Fig. S5) also lead to a potential sampling bias towards mid- to higher elevation sites. Our approach to account for imperfect detection assumes that the variation in detection probabilities among sites can be adequately captured by random effects and may overlook ecological factors influencing detection (Lindenmayer et al. 2001; Wintle et al. 2005; Cripps et al. 2021). While our approach may be a robust way to account for unobserved heterogeneity in detection probabilities among sites and between observers (Kéry et al. 2009; Camp et al. 2023) and can account for spatial autocorrelation and temporal variation in our surveys, occupancy models would be better suited to overcome imperfect detection and variation in survey effort. Unfortunately, due to the application of different survey designs pre- and post-fire and small sample sizes for wildlife surveys (30 pre and 40 post-fire, with one survey per site and two observers) occupancy modeling would have likely not yielded robust results (Mackenzie and Royle 2005; MacKenzie et al. 2017). Future studies into the effects of fire on arboreal fauna habitat should therefore consider a dedicated before-after control-impact (BACI) design to allow appropriate and ongoing monitoring. This should be an important consideration for researchers and land managers in an age of increasingly common mega-fires (Kelly et al. 2023).
Conclusion
We assessed mature, mixed-species Eucalyptus forests in East Gippsland, Victoria, to better understand the factors controlling a key stand-scale habitat feature for arboreal fauna - hollow-bearing tree occurrence – and how this important resource was affected by the 2019/20 Australian megafires. Before the fires, mixed-species forests had an abundance of hollows across all elevations and associated forest types. Hollow occurrence and abundance did not decrease in forests affected by low to medium fire severities, but forests burnt at high severity had fewer hollows, while the number of hollow-bearing trees remained stable. Furthermore, we found that the fires changed the probability of hollow occurrence with a higher likelihood at smaller tree sizes across elevation bands. Our observations provide hope that fire-affected forests may continue to provide nesting space and therefore critical habitat as they recover from devastating landscape-scale bushfires. This knowledge provides a useful tool for making forest management and conservation decisions for the endangered southern greater glider and associated arboreal fauna in the region but emphasizes the need to establish long-term monitoring and to better understand the impacts of fire to and the recovery process of key foraging resources for folivores in mixed species eucalypt forests.
Availability of data and materials
The codes and datasets supporting the findings of this research are accessible on the Github repository: https://github.com/BennyWag/hollows. All fauna observation locations are available on the Victorian Biodiversity Atlas (VBA, vba.dse.vic.gov.au/vba).
References
Adkins, M. F. 2006. A burning issue: using fire to accelerate tree hollow formation in Eucalyptus species. Australian Forestry 69(2): 107–113. https://doi.org/10.1080/00049158.2006.10676236.
Anderson, D., and K. Burnham. 2004. Model selection and multi-model inference. Second. New York: Springer-Verlag, 63(2020):10.
Au, J., R. G. Clark, C. Allen, K. J. Marsh, W. J. Foley, and K. N. Youngentob. 2019. A nutritional mechanism underpinning folivore occurrence in disturbed forests. Forest Ecology and Management 453. https://doi.org/10.1016/j.foreco.2019.117585.
Banks, S. C., E. J. Knight, L. McBurney, D. Blair, and D. B. Lindenmayer. 2011. The effects of wildfire on mortality and resources for an arboreal marsupial: resilience to fire events but susceptibility to fire regime change. Plos One 6(8): e22952. https://doi.org/10.1371/journal.pone.0022952.
Barton, K. 2019. MuMIn: Multi-Model Inference. Retrieved from: https://CRAN.R-project.org/package=MuMIn.
Bates, D., M. Mächler, B. Bolker, and S. Walker. 2015. Fitting linear mixed-effects models using lme4. Journal of Statistical Software 67(1): 1–48. https://doi.org/10.18637/jss.v067.i01.
Bennett, A., L. Lumsden, J. Alexander, P. Duncan, P. Johnson, P. Robertson, and C. Silveira. 1991. Habitat use by arboreal mammals along an environment gradient in north-eastern Victoria. Wildlife Research 18(2): 125–146. https://doi.org/10.1071/WR9910125.
Bennett, A., L. Lumsden, and A. Nicholls. 1994. Tree hollows as a resource for wildlife in remnant woodlands: spatial and temporal patterns across the northern plains of Victoria, Australia. Pacific Conservation Biology 1(3): 222–235.
Berry, L. E., D. A. Driscoll, S. C. Banks, and D. B. Lindenmayer. 2015. The use of topographic fire refuges by the greater glider (Petauroides volans) and the mountain brushtail possum (Trichosurus cunninghami) following a landscape-scale fire. Australian Mammalogy 37(1): 39–45. https://doi.org/10.1071/Am14027.
Beven, K. J., and M. J. Kirkby. 1979. A physically based, variable contributing area model of basin hydrology/Un modèle à base physique de zone d’appel variable de L’hydrologie Du bassin versant. Hydrological Sciences Journal 24(1): 43–69. https://doi.org/10.1080/02626667909491834.
Bowman, D., G. J. Williamson, R. J. Keenan, and L. D. Prior. 2014a. A warmer world will reduce tree growth in evergreen broadleaf forests: evidence from Australian temperate and subtropical eucalypt forests. Global Ecology and Biogeography 23(8): 925–934. https://doi.org/10.1111/geb.12171.
Bowman, D. M., B. P. Murphy, D. L. Neyland, G. J. Williamson, and L. D. Prior. 2014b. Abrupt fire regime change may cause landscape-wide loss of mature obligate seeder forests. Global Change Biology 20(3): 1008–1015.
Camp, R. J., C. K. Asing, P. C. Banko, L. Berry, K. W. Brinck, C. Farmer, and A. S. Genz. 2023. Evaluation of replicate sampling using hierarchical spatial modeling of population surveys accounting for imperfect detectability. Wildlife Society Bulletin 47(3). https://doi.org/10.1002/wsb.1471.
Campbell‐Jones, M. M., Bassett, M., Bennett, A. F., Chia, E. K., Leonard, S., & Collins, L. 2022. Fire severity has lasting effects on the distribution of arboreal mammals in a resprouting forest. Austral Ecology 47(7): 1456-1469.
Chia, E.K., M. Bassett, S.W.J. Leonard, G.J. Holland, E.G. Ritchie, M.F. Clarke, and A.F. Bennett. 2016. Effects of the fire regime on mammal occurrence after wildfire: site effects vs landscape context in fire-prone forests. Forest Ecology and Management 363: 130–139. https://doi.org/10.1016/j.foreco.2015.12.008.
Chick, R., J. K. Cripps, L. Durkin, J. L. Nelson, J. Molloy, and M. Edmonds. 2020. Forest Protection Survey Program - Survey Guideline - Spotlighting and Call Playback (V4.1): The State Department of Environment, Land, Water and Planning.
Collins, L., A. F. Bennett, S. W. J. Leonard, and T. D. Penman. 2019. Wildfire refugia in forests: severe fire weather and drought mute the influence of topography and fuel age. Glob Chang Biol 25(11): 3829–3843. https://doi.org/10.1111/gcb.14735.
Collins, L., R. A. Bradstock, H. Clarke, M. F. Clarke, R. H. Nolan, and T. D. Penman. 2021. The 2019/2020 mega-fires exposed Australian ecosystems to an unprecedented extent of high-severity fire. Environmental Research Letters 16(4). https://doi.org/10.1088/1748-9326/abeb9e.
Collins, L., H. Clarke, M. F. Clarke, S. C. McColl Gausden, R. H. Nolan, T. Penman, and M. Varner. 2022. Warmer and drier conditions have increased the potential for large and severe fire seasons across south-eastern Australia. Global Ecology and Biogeography 31(10): 1933–1948. https://doi.org/10.1111/geb.13514.
Comport, S. S., S. J. Ward, and W. J. Foley. 1996. Home ranges, time budgets and food-tree use in a high-density tropical population of greater gliders, Petauroides volans minor (Pseudocheiridae: Marsupialia). Wildlife Research 23(4): 401–419. https://doi.org/10.1071/Wr9960401.
Cripps, J. K., J. L. Nelson, M. P. Scroggie, L. K. Durkin, D. S. Ramsey, and L. F. Lumsden. 2021. Double-observer distance sampling improves the accuracy of density estimates for a threatened arboreal mammal. Wildlife Research 48(8): 756–768.
Cunningham, R. B., M. L. Pope, and D. B. Lindenmayer. 2004. Patch use by the greater glider (Petauroides volans) in a fragmented forest ecosystem. III. Night-time use of trees. Wildlife Research 31(6): 579–585. https://doi.org/10.1071/Wr02112.
Davey, S. M. 2018a. Reporting Australia’s forest biodiversity I: forest-dwelling and forest-dependent native species. Australian Forestry 81(3): 196–209. https://doi.org/10.1080/00049158.2018.1509683.
Davey, S. M. 2018b. Reporting Australia’s forest biodiversity II: threatened forest-dwelling and forest-dependent species. Australian Forestry 81(4): 214–230. https://doi.org/10.1080/00049158.2018.1510627.
DAWE. 2021. Victorian Regional Forest Agreements Major Event Review of the 2019-20 bushfires. Australian Government - Department of Agriculture, Water and the Environment.
DeGabriel, J. L., B. D. Moore, K. J. Marsh, and W. J. Foley. 2009. The effect of plant secondary metabolites on the interplay between the internal and external environments of marsupial folivores. Chemoecology 20(2): 97–108.
DELWP. 2022. Biodiversity response and recovery supplementary report: bushfire impacts on species in Victoria. The State Department of Environment, Land, Water and Planning.
DELWP. 2019. Greater Glider (Petauroides volans subsp. volans) Action statement No. 267 Flora Fauna Guarantee Act 1988 - Version 1.0: The State Department of Environment, Land, Water and Planning.
Montreal Process Implementation Group for Australia and National Forest Inventory Steering Committee. 2018. Australia’s State of the Forests Report 2018, ABARES, Canberra, December. CC BY 4.0. ISBN 978-1-74323-407-5.
Department of Conservation and Natural Resources. 1995. Forest management plan for the East Gippsland Forest Management Area. Department of Conservation and Natural Resources.
Diamond, J. 1984. Normal extinctions of isolated populations. In Extinctions, ed. M. H. Nitecki. 191–246. Chicago: University of Chicago Press.
Eyre, T. 2006. Regional habitat selection of large gliding possums at forest stand and landscape scales in southern Queensland, Australia: I. Greater glider (Petauroides volans). Forest Ecology and Management 235(1): 270–282.
Farr, T.G., Rosen, P.A., Caro, E., Crippen, R., Duren, R., Hensley, S., Kobrick, M., Paller, M., Rodriguez, E., Roth, L. and Seal, D. 2007. The shuttle radar topography mission. Reviews of geophysics 45(2). https://agupubs.onlinelibrary.wiley.com/doi/full/10.1029/2005RG000183#.
Forrester, D. I., M. Schortemeyer, W. D. Stock, J. Bauhus, P. K. Khanna, and A. L. Cowie. 2007. Assessing nitrogen fixation in mixed-and single-species plantations of Eucalyptus globulus and Acacia mearnsii. Tree Physiology 27(9): 1319–1328.
Fox, J. C., F. Hamilton, and P. K. Ades. 2008. Models of tree-level hollow incidence in victorian state forests. Forest Ecology and Management 255(7): 2846–2857.
Fox, J. C., F. Hamilton, and S. Occhipinti. 2009. Tree hollow incidence in victorian state forests. Australian Forestry 72(1): 39–48.
Geary, W. L., A. Buchan, T. Allen, D. Attard, M. J. Bruce, L. Collins, … E. Kelly. 2021. Responding to the biodiversity impacts of a megafire: a case study from south-eastern Australia’s Black Summer. Diversity and Distributions. https://doi.org/10.1111/ddi.13292.
Gibbons, P., and D. B. Lindenmayer. 2002. Tree hollows and wildlife conservation in Australia. Collingwood: CSIRO Publishing.
Gibbons, P., D. B. Lindenmayer, S. Barry, and M. Tanton. 2000. Hollow formation in eucalypts from temperate forests in southeastern Australia. Pacific Conservation Biology 6(3): 218.
Gibbons, P., D. B. Lindenmayer, S. C. Barry, and M. T. Tanton. 2002. Hollow selection by vertebrate fauna in forests of southeastern Australia and implications for forest management. Biological Conservation 103(1): 1–12. https://doi.org/10.1016/S0006-3207(01)00109-4.
Gibbons, P., D. Stojanovic, D. B. Lindenmayer, and G. Owens. 2024. Impacts of changing fire regimes on hollow-bearing trees in south-eastern Australia. International Journal of Wildland Fire 33(2). https://doi.org/10.1071/wf23094.
Goldingay, R. L. 2011. Characteristics of tree hollows used by Australian arboreal and scansorial mammals. Australian Journal of Zoology 59(5). https://doi.org/10.1071/zo11081.
Haslem, A., S. C. Avitabile, R. S. Taylor, L. T. Kelly, S. J. Watson, D. G. Nimmo, and M. F. Clarke. 2012. Time-since-fire and inter-fire interval influence hollow availability for fauna in a fire-prone system. Biological Conservation 152: 212–221. https://doi.org/10.1016/j.biocon.2012.04.007.
Henry, S. 1984. Social organisation of the greater glider (Petauroides volans) in Victoria. In Possums and gliders, eds. A. P. Smith, and I. D. Hume. 221–228. Chipping Norton: Surrey Beatty and Sons.
Hijmans, R. J. 2019. raster: Geographic data analysis and modeling. Retrieved from https://CRAN.R-project.org/package=raster.
Hofman, M., A. Gracanin, and K. M. Mikac. 2022. Greater glider (Petauroides volans) den tree and hollow characteristics. Australian Mammalogy. https://doi.org/10.1071/AM22008.
Inions, G., M. Tanton, and S. Davey. 1989. Effect of fire on the availability of Hollows in Trees used by the Common Brushtail Possum, Trichosurus Vulpecula Kerr, 1792, and the Ringtail Possum, Pseudocheirus Peregrinus boddaerts, 1785. Wildlife Research 16(4): 449–458. https://doi.org/10.1071/WR9890449.
Kavanagh, R. 2000. Effects of variable-intensity logging and the influence of habitat variables on the distribution of the greater glider Petauroides volans in montane forest, southeastern New South Wales. Pacific Conservation Biology 6(1): 18–30. https://doi.org/10.1071/PC000018.
Kavanagh, R., and M. Lambert. 1990. Food Selection by the Greater Glider, Petauroides volans: is Foliar Nitrogen a determinant of Habitat Quality? Australian Wildlife Research 17(3): 285–299. https://doi.org/10.1071/WR9900285.
Kearney, M. R., B. A. Wintle, and W. P. Porter. 2010. Correlative and mechanistic models of species distribution provide congruent forecasts under climate change. Conservation Letters 3(3): 203–213. https://doi.org/10.1111/j.1755-263X.2010.00097.x.
Kelly, L.T., Fletcher, M.S., Menor, I.O., Pellegrini, A.F., Plumanns-Pouton, E.S., Pons, P., Williamson, G.J. and Bowman, D.M. 2023. Understanding Fire Regimes for a Better Anthropocene. Annual Review of Environment and Resources 48(1):207-235.
Kéry, M., R. M. Dorazio, L. Soldaat, A. Van Strien, A. Zuiderwijk, and J. A. Royle. 2009. Trend estimation in populations with imperfect detection. Journal of Applied Ecology 46(6): 1163–1172. https://doi.org/10.1111/j.1365-2664.2009.01724.x.
Koch, A., S. Munks, and D. Driscoll. 2008a. The use of hollow-bearing trees by vertebrate fauna in wet and dry Eucalyptus obliqua forest, Tasmania. Wildlife Research 35(8): 727–746. https://doi.org/10.1071/WR08007.
Koch, A. J., S. A. Munks, D. Driscoll, and J. Kirkpatrick. 2008b. Does hollow occurrence vary with forest type? A case study in wet and dry Eucalyptus obliqua forest. Forest Ecology and Management 255(12): 3938–3951.
Lefoe, M., A. R. Rendall, F. McKinnon, and D. A. Whisson. 2022. Logging and wildfire limit the distribution of a vulnerable arboreal mammal. Forest Ecology and Management 503: 119773.
Legge, S. M., J. C. Woinarski, S. T. Garnett, H. M. Geyle, M. Lintermans, D. G. Nimmo, and D. P. Wilkinson. 2021. Estimates of the impacts of the 2019-20 fires on populations of native animal species. Retrieved from Australia.
Lindenmayer, D. B., and J. T. Wood. 2010. Long-term patterns in the decay, collapse, and abundance of trees with hollows in the mountain ash (Eucalyptus regnans) forests of Victoria, southeastern Australia. Canadian Journal of Forest Research 40(1): 48–54.
Lindenmayer, D. B., R. Cunningham, M. Tanton, A. Smith, and H. Nix. 1990a. The conservation of arboreal marsupials in the montane ash forests of the Central Highlands of Victoria, Southeast Australia: I. factors influencing the occupancy of trees with hollows. Biological Conservation 54(2): 111–131.
Lindenmayer, D. B., R. B. Cunningham, M. T. Tanton, A. P. Smith, and H. A. Nix. 1990b. Habitat requirements of the Mountain Brushtail Possum and the Greater Glider in the Montane Ash-Type Eucalypt forests of the Central highlands of Victoria. Australian Wildlife Research 17(5): 467–478.
Lindenmayer, D. B., R. Cunningham, H. Nix, M. Tanton, and A. Smith. 1991a. Predicting the abundance of hollow-bearing trees in montane forests of southeastern Australia. Australian Journal of Ecology 16(1): 91–98.
Lindenmayer, D. B., R. Cunningham, M. Tanton, H. Nix, and A. Smith. 1991b. The conservation of arboreal marsupials in the montane ash forests of the Central Highlands of Victoria, south-east Australia: III. The habitat requirements of Leadbeater’s Possum Gymnobelideus leadbeateri and models of the diversity and abundance of arboreal marsupials. Biological Conservation 56(3): 295–315.
Lindenmayer, D. B., R. Cunningham, M. Tanton, A. Smith, and H. Nix. 1991c. Characteristics of hollow-bearing trees occupied by arboreal marsupials in the montane ash forests of the Central Highlands of Victoria, south-east Australia. Forest Ecology and Management 40(3–4): 289–308.
Lindenmayer, D. B., R. Cunningham, C. Donnelly, M. Tanton, and H. Nix. 1993a. The abundance and development of cavities in Eucalyptus trees: a case study in the montane forests of Victoria, southeastern Australia. Forest Ecology and Management 60(1–2): 77–104.
Lindenmayer, D. B., R. Cunningham, and C. Donnelly. 1993b. The conservation of arboreal marsupials in the montane ash forests of the Central Highlands of Victoria, south-east Australia, IV. The presence and abundance of arboreal marsupials in retained linear habitats (wildlife corridors) within logged forest. Biological Conservation 66(3): 207–221.
Lindenmayer, D. B., R. B. Cunningham, C. F. Donnelly, R. D. Incoll, M. L. Pope, C. R. Tribolet, and A. H. Welsh. 2001. How effective is spotlighting for detecting the greater glider (< emph type=2>Petauroides volans)? Wildlife Research 28(1): 105–109. https://doi.org/10.1071/WR00002.
Lindenmayer, D. B., J. T. Wood, L. McBurney, C. MacGregor, K. Youngentob, and S. C. Banks. 2011. How to make a common species rare: a case against conservation complacency. Biological Conservation 144(5): 1663–1672. https://doi.org/10.1016/j.biocon.2011.02.022.
Lindenmayer, D. B., W. Blanchard, D. Blair, L. McBurney, and S. C. Banks. 2016. Environmental and human drivers influencing large old tree abundance in Australian wet forests. Forest Ecology and Management 372: 226–235. https://doi.org/10.1016/j.foreco.2016.04.017.
Lindenmayer, D. B., W. Blanchard, D. Blair, L. McBurney, and S. C. Banks. 2017. Relationships between tree size and occupancy by cavity-dependent arboreal marsupials. Forest Ecology and Management 391: 221–229. https://doi.org/10.1016/j.foreco.2017.02.014.
Lindenmayer, D. B., L. McBurney, W. Blanchard, K. Marsh, E. Bowd, D. Watchorn, and K. Youngentob. 2022. Elevation, disturbance, and forest type drive the occurrence of a specialist arboreal folivore. Plos One 17(4): e0265963. https://doi.org/10.1371/journal.pone.0265963.
Mackenzie, D. I., and J. A. Royle. 2005. Designing occupancy studies: general advice and allocating survey effort. Journal of Applied Ecology 42(6): 1105–1114. https://doi.org/10.1111/j.1365-2664.2005.01098.x.
MacKenzie, D. I., J. D. Nichols, J. A. Royle, K. H. Pollock, L. L. Bailey, and J. E. Hines. 2017. Occupancy estimation and modeling: inferring patterns and dynamics of species occurrence. Elsevier.
Manning, A. D., P. Gibbons, J. Fischer, D. L. Oliver, and D. B. Lindenmayer. 2013. Hollow futures? Tree decline, lag effects and hollow-dependent species. Animal Conservation 16(4): 395–403. https://doi.org/10.1111/acv.12006.
Mazurek, M., and W. J. Zielinski. 2004. Individual legacy trees influence vertebrate wildlife diversity in commercial forests. Forest Ecology and Management 193(3): 321–334.
McCarthy, G., K. Moon, and L. Smith. 2017. Mapping fire severity and fire extent in forest in Victoria for ecological and fuel outcomes. Ecological Management & Restoration 18(1): 54–65. https://doi.org/10.1111/emr.12242.
McLean, C. M., R. Bradstock, O. Price, and R. P. Kavanagh. 2015. Tree hollows and forest stand structure in Australian warm temperate Eucalyptus forests are adversely affected by logging more than wildfire. Forest Ecology and Management 341: 37–44. https://doi.org/10.1016/j.foreco.2014.12.023.
McLean, C. M., R. P. Kavanagh, T. Penman, and R. Bradstock. 2018. The threatened status of the hollow dependent arboreal marsupial, the Greater Glider (Petauroides volans), can be explained by impacts from wildfire and selective logging. Forest Ecology and Management 415: 19–25. https://doi.org/10.1016/j.foreco.2018.01.048.
Mitchell, K. 2010. Quantitative analysis by the point-centered quarter method. arXiv preprint arXiv:1010.3303.
Nelson, J. L., K. Cherry, and K. Porter. 1996. The effect of edges on the distribution of arboreal marsupials in the ash forests of the victorian Central Highlands. Australian Forestry 59(4): 189–198.
Nitschke, C. R., R. Trouvé, L. F. Lumsden, L. T. Bennett, M. Fedrigo, A. P. Robinson, and P. J. Baker. 2020. Spatial and temporal dynamics of habitat availability and stability for a critically endangered arboreal marsupial: implications for conservation planning in a fire-prone landscape. Landscape Ecology. https://doi.org/10.1007/s10980-020-01036-2.
Nolan, R. H., M. M. Boer, L. Collins, V. Resco de Dios, H. Clarke, M. Jenkins, and R. A. Bradstock. 2020. Causes and consequences of eastern Australia’s 2019-20 season of mega-fires. Glob Chang Biol 26(3): 1039–1041. https://doi.org/10.1111/gcb.14987.
Opie, A., G. Gillespie, S. Henry, V. Hurley, B. Lobert, and J. Westaway. 1990. Flora and Fauna Survey of the Coast Range Forest Block,(Southern Part), East Gippsland, Victoria. Department of Conservation, Forests & Lands. Retrieved from Melbourne https://trove.nla.gov.au/version/44880648.
Parnaby, H., D. Lunney, I. Shannon, and M. Fleming. 2010. Collapse rates of hollow-bearing trees following low intensity prescription burns in the Pilliga forests, New South Wales. Pacific Conservation Biology 16(3): 209–220.
Pebesma, E. 2018. Simple features for R: standardized support for spatial vector data. The R Journal 10(1): 439–446.
Powell, R. A. 2000. Animal home ranges and territories and home range estimators. In Research techniques in animal ecology: controversies and consequences, eds. L. Boitani, and T. K. Fuller. vol. 1 100. New York: Columbia University.
Price, O. F., and R. A. Bradstock. 2012. The efficacy of fuel treatment in mitigating property loss during wildfires: insights from analysis of the severity of the catastrophic fires in 2009 in Victoria, Australia. Journal of Environmental Management 113: 146–157. https://doi.org/10.1016/j.jenvman.2012.08.041.
Pulsford, S. A., D. B. Lindenmayer, and D. A. Driscoll. 2016. A succession of theories: purging redundancy from disturbance theory. Biological Reviews of the Cambridge Philosophical Society 91(1): 148–167. https://doi.org/10.1111/brv.12163.
Rainsford, F. W., L. T. Kelly, S. W. J. Leonard, and A. F. Bennett. 2020. Post-fire development of faunal habitat depends on plant regeneration traits. Austral Ecology. https://doi.org/10.1111/aec.12896.
R Core Development Team. 2022. R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing. Retrieved from http://www.r-project.org.
Rübsamen, K., I. Hume, W. Foley, and U. Rübsamen. 1984. Implications of the large surface-area to body-Mass ratio on the heat-balance of the Greater glider (Petauroides-Volans-Marsupialia). Journal of Comparative Physiology 154(1): 105–111. https://doi.org/10.1007/Bf00683223.
Salmona, J., K. M. Dixon, and S. C. Banks. 2018. The effects of fire history on hollow-bearing tree abundance in montane and subalpine eucalypt forests in southeastern Australia. Forest Ecology and Management 428: 93–103. https://doi.org/10.1016/j.foreco.2018.06.026.
Sebire, I., and P. C. Fagg. 2009. High elevation mixed species in Victoria’s state forests. Australia: Dept. of Sustainability and Environment.
Shipley, L. A., J. S. Forbey, and B. D. Moore. 2009. Revisiting the dietary niche: when is a mammalian herbivore a specialist? Integrative and Comparative Biology 49(3): 274–290. https://doi.org/10.1093/icb/icp051.
Smith, A., and D. Lindenmayer. 1988. Tree hollow requirements of Leadbeater’s possum and other possums and gliders in timber production ash forests of the victorian central highlands. Wildlife Research 15(4): 347–362.
Smith, P. and Smith, J. 2020. Future of the greater glider ('Petauroides volans') in the blue mountains, New South Wales. In Proceedings of the Linnean Society of New South Wales (Vol. 142, pp. 55-66).
Smith, G. C., M. Mathieson, and L. Hogan. 2007. Home range and habitat use of a low-density population of greater gliders, Petauroides volans (Pseudocheiridae: Marsupialia), in a hollow-limiting environment. Wildlife Research 34(6): 472–483. https://doi.org/10.1071/Wr06063.
Stewart, S. B., and C. Nitschke. 2017a. Climate Victoria: Maximum Temperature (3DS; 9 second, approx. 250 m). v2. CSIRO [Data Collection]. Retrieved from: https://doi.org/10.25919/5e5d9c82c3b16.
Stewart, S. B., and C. R. Nitschke. 2017b. Improving temperature interpolation using MODIS LST and local topography: a comparison of methods in South East Australia. International Journal of Climatology 37(7): 3098–3110. https://doi.org/10.1002/joc.4902.
Stewart, S. B., and C. Nitschke. 2018. Climate Victoria: Minimum Temperature (3DS-TM; 9 second, approx. 250 m). v2. CSIRO [Data Collection]. Retrieved from: https://doi.org/10.25919/5e5dc18621a12.
Stewart, S. B., M. Fedrigo, S. H. Roxburgh, S. Kasel, and C. Nitschke. 2020a. Climate Victoria: 9am Vapour Pressure (9 second, approx. 250 m). v3. CSIRO [Data Collection]. Retrieved from: https://doi.org/10.25919/5e5701b5df4d8.
Stewart, S. B., M. Fedrigo, S. H. Roxburgh, S. Kasel, and C. Nitschke. 2020b. Climate Victoria: Precipitation (9 second, approx. 250 m). v1. CSIRO [Data Collection]. Retrieved from: https://doi.org/10.25919/5e3be5193e301.
Tiribelli, F., J. M. Morales, J. H. Gowda, M. Mermoz, and T. Kitzberger. 2019. Non-additive effects of alternative stable states on landscape flammability in NW Patagonia: fire history and simulation modelling evidence. International Journal of Wildland Fire 28(2). https://doi.org/10.1071/wf18073.
Trouvé, R., S. Bunyavejchewin, P. J. Baker, and E. Lines. 2020. Disentangling fire intensity and species’ susceptibility to fire in a species-rich seasonal tropical forest. Journal of Ecology 108(4): 1664–1676. https://doi.org/10.1111/1365-2745.13343.
Trouvé, R., Oborne, L., & Baker, P. J. 2021. The effect of species, size, and fire intensity on tree mortality within a catastrophic bushfire complex. Ecological Applications 31(6):e02383.
Turner, M. G., and R. H. Gardner. 2015. Landscape ecology in theory and practice: pattern and process. 2nd ed. ed.): Springer.
van der Ree, R., S. Ward, and K. Handasyde. 2004. Distribution and conservation status of possums and gliders in Victoria. In The Biology of Australian possums and gliders, eds. R. L. Goldingray, and S. M. Jackson. 91–110. Sydney: Surrey Beatty and Sons.
Wagner, B. 2021. The role of climate and tree nutrition on the occurrence of the southern greater glider (Petauroides volans) and its implications for conservation planning (PhD thesis). The University of Melbourne, Melbourne, Australia. Retrieved from http://hdl.handle.net/11343/282520.
Wagner, B., P. J. Baker, S. B. Stewart, L. F. Lumsden, J. L. Nelson, J. K. Cripps, … C. R. Nitschke. 2020. Climate change drives habitat contraction of a nocturnal arboreal marsupial at its physiological limits. Ecosphere, 11(10), e03262. https://doi.org/10.1002/ecs2.3262. ARTN
Wagner, B., P. J. Baker, and C. R. Nitschke. 2021a. The influence of spatial patterns in foraging habitat on the abundance and home range size of a vulnerable arboreal marsupial in southeast Australia. Conservation Science and Practice e566. https://doi.org/10.1111/csp2.566.
Wagner, B., P. J. Baker, B. D. Moore, and C. R. Nitschke. 2021b. Mapping canopy nitrogen-scapes to assess foraging habitat for a vulnerable arboreal folivore in mixed-species Eucalyptus forests. Ecology and Evolution 11(24): 18401–18421. https://doi.org/10.1002/ece3.8428.
Wang, T., A. Hamann, A. Yanchuk, G. A. O’Neill, and S. N. Aitken. 2006. Use of response functions in selecting lodgepole pine populations for future climates. Global Change Biology 12(12): 2404–2416. https://doi.org/10.1111/j.1365-2486.2006.01271.x.
Ward, M., A. I. T. Tulloch, J. Q. Radford, B. A. Williams, A. E. Reside, S. L. Macdonald, and J. E. M. Watson. 2020. Impact of 2019–2020 mega-fires on Australian fauna habitat. Nat Ecol Evol 4(10): 1321–1326. https://doi.org/10.1038/s41559-020-1251-1.
Wing, J., M. Kuhn, S. Weston, A. Williams, C. Keefer, A. Engelhardt, and T. Hunt. 2019. caret: Classification and regression training. Retrieved from: https://CRAN.R-project.org/package=caret.
Wintle, B.A., R.P. Kavanagh, M.A. McCarthy, and M.A. Burgman. 2005. Estimating and dealing with detectability in occupancy surveys for forest owls and arboreal marsupials. The Journal of Wildlife Management 69 (3): 905–917.
Woinarski, J.C.Z., Burbidge, A.A., and Harrison, P.L. 2014. The Action Plan for Australian Mammals 2012. Collingwood, Vic.: CSIRO Publishing.
Woolley, L.-A., B.P. Murphy, I.J. Radford, J. Westaway, and J.C.Z. Woinarski. 2018. Cyclones, fire, and termites: the drivers of tree hollow abundance in northern Australia’s mesic tropical savanna. Forest Ecology and Management 419–420: 146–159. https://doi.org/10.1016/j.foreco.2018.03.034.
Youngentob, K., I.R. Wallis, D.B. Lindenmayer, J.T. Wood, M.L. Pope, and W.J. Foley. 2011. Foliage Chemistry influences Tree Choice and Landscape Use of a gliding Marsupial Folivore. Journal of Chemical Ecology 37 (1): 71–84. https://doi.org/10.1007/s10886-010-9889-9.
Youngentob, K., J.T. Wood, and D.B. Lindenmayer. 2013. The response of arboreal marsupials to landscape context over time: a large-scale fragmentation study revisited. Journal of Biogeography 40 (11): 2082–2093. https://doi.org/10.1111/jbi.12158.
Acknowledgements
We recognize and acknowledge the traditional owners of the lands on which this research was conducted: The Wurundjeri, Gunaikurnai, Taungurung, and Bidwell people. We pay our respects to their Elders, past, present, and emerging. We thank and acknowledge Dr. Anu Singh for her work of efficiently and effectively managing and carrying out the field data collection on this very time-limited project. Further acknowledgements for field- and survey work goes to Kaitlyn Hammond, Jeremy Johnson, Harry Barton, Dr. Yogendra Kumar Karna, Bhuban Timalsina, Thiet Nguyen, Amanda Lo Cascio, Bernard Gandy, Tim Willersdorf, and Melina Budden. We thank our collaborators at the Arthur Rylah Institute for Environmental Research (Heidelberg, Victoria) for providing survey data and guidance for additional sites, namely Jemma Cripps, Louise Durkin, Phoebe Macak, Jenny Nelson, Lindy Lumsden, and Michael Scroggie. We further thank chair Paul Ferraro (DEECA) and the members of the federal yellow-bellied glider and greater glider bushfire recovery working group for a fruitful exchange of ideas and collaboration over the cause of this project. Finally, we would like to acknowledge our collaborators at the Australian National University, Drs. Kara Youngentob, and Karen Marsh.
Funding
BW acknowledges funding by the University of Melbourne through the Melbourne Research Scholarship. BW and CRN acknowledge funding from the Australian Department of Climate Change, Energy, the Environment and Water. This project was funded via the Regional Bushfire Recovery for Multiregional Species and Strategic Projects Program. CRN acknowledges funding through the Integrated Forest Ecosystem Research (IFER) program of DEECA. This research was also supported by the Australian Research Council (LP140100580 to CRN and PJB., FT120100715 to PJB).
Author information
Authors and Affiliations
Contributions
BW and CRN conceived the idea and developed the methodology for this research. BW conducted fieldwork, collated, and analyzed the data and drafted the manuscript with contributions from CRN and PJB. All authors contributed to the drafts and gave final approval for publication. There are no conflicts of interest to declare.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This research was conducted with an access agreement for parks and reserves from Parks Victoria and a research permit for state forests issued by the Victorian Department of Energy, Environment and Climate Action (DEECA, permit no. 2022-01-25). Spotlighting surveys for arboreal fauna were conducted with University of Melbourne animal ethics approval (ID no. 2021-22840-21795-2).
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wagner, B., Baker, P.J. & Nitschke, C.R. How an unprecedented wildfire shaped tree hollow occurrence and abundance—implications for arboreal fauna. fire ecol 20, 42 (2024). https://doi.org/10.1186/s42408-024-00274-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s42408-024-00274-y