
Abstract
Background
Understanding pre-1850s fire history and its effect on forest structure can provide insights useful for fire managers in developing plans to moderate fire hazards in the face of forecasted climate change. While climate clearly plays a substantial role in California wildfires, traditional use of fire by Indigenous people also affected fire history and forest structure in the Sierra Nevada. Disentangling the effects of human versus climatically-induced fire on Sierran forests from paleoecological records has historically proved challenging, but here we use pollen-based forest structure reconstructions and comparative paleoclimatic-vegetation response modeling to identify periods of human impact over the last 1300 years at Markwood Meadow, Sierra National Forest.
Results
We find strong evidence for anthropogenic fires at Markwood Meadow ca. 1550 – 1750 C.E., contemporaneous with archaeological evidence for fundamental shifts in Indigenous lifeways. When we compare our findings to five other paleoecological sites in the central and southern Sierra Nevada, we find evidence for contemporaneous anthropogenic effects on forest structure across a broad swath of cismontane central California. This is significant because it implies that late 19th and early twentieth century forest structure – the structure that land managers most often seek to emulate – was in part the result anthropogenic fire and precolonial resource management.
Conclusion
We consequently suggest that modern management strategies consider (1) further incorporating traditional ecological knowledge fire practices in consultation with local tribal groups, and (2) using pollen-based reconstructions to track how forest composition compares to pre-1850 C.E. conditions rather than the novel forest states encountered in the late 20th and early twenty-first centuries. These strategies could help mitigate the effects of forecast climate change and associated megafires on forests and on socio-ecological systems in a more comprehensive manner.
Resumen
Antecedentes
El entendimiento de la historia del fuego previa al 1850 y sus efectos sobre la estructura forestal, puede proveer de información útil para los gestores del fuego en el desarrollo de los planes para moderar los peligros de incendios en relación al cambio climático pronosticado. Mientras que el clima juega un rol fundamental en los incendios de California, el uso tradicional del fuego por pueblos indígenas también ha afectado la historia del fuego y la estructura forestal en la Sierra Nevada. El desentrañar los efectos humanos versus aquellos inducidos por el clima en los incendios en la Sierra Nevada desde registros paleo-ecológicos ha sido probado como algo desafiante. Usamos en este caso reconstrucciones de bosques basadas en la estructura del polen y respuestas comparativas entre vegetación y registros paleo-climáticos modelados para identificar períodos de impactos humanos sobre los últimos 1300 años en el sitio Markwood meadow, en el Bosque Nacional de Sierra Nevada.
Resultados
Encontramos una muy fuerte evidencia de fuegos antropogénicos en el Markwood meadow entre 1550 y 1750 de la era cristiana, siendo contemporánea con evidencias arqueológicas de cambios fundamentales en la vida de los indígenas. Cuando comparamos nuestros resultados con otros cinco sitios paleo-ecológicos en el centro y sur de la Sierra Nevada, encontramos evidencia de efectos antropogénicos contemporáneos en la estructura forestal a través de un amplio muestrario en las montañas del centro de California. Esto es significativo ya que implica que a fines del siglo XIX y principios del XX, la estructura forestal –la estructura forestal que muchos silvicultores buscan emular—fue en parte el resultado de fuegos antropogénicos y manejo de recursos en épocas precoloniales.
Conclusiones
Consecuentemente, sugerimos que las estrategias de manejo moderno deben considerar (1) incorporar el conocimiento ecológico tradicional en consulta con grupos de tribus locales, y (2) usar reconstrucciones basadas en polen para detectar si la composición del bosque se compara con aquellas condiciones previas al 1850 de nuestra era cristiana, más que con los estados noveles del bosque encontrados en las postrimerías del siglo XX y comienzos del XXI. Estas estrategias pueden mitigar los efectos del cambio climático pronosticado y los mega-incendios asociados, tanto sobre los bosques como sobre los ecosistemas socio-ecológicos de una manera más comprensiva.
Similar content being viewed by others


Background
Dense forests with thick underbrush have developed in California’s Sierra Nevada range since ca. 1850 through fire exclusion, grazing, selective logging, road construction, and fire suppression policies (Silcox 1910; Martin and Sapsis 1992; Bowman et al. 2011). These forests have uncharacteristically continuous canopies that shade the forest floor, which increases litter accumulation and encourages the growth of shade-tolerant taxa such as fir (Abies spp.; Kilgore and Taylor 1979; Wilkin et al. 2021). Forest fire severity and frequency is increasing in the Sierra Nevada (Miller et al. 2009), with costly fires like the 2020 August Complex (the largest California wildfire on record as of 2022, 4178 km2), the 2021 Dixie Fire (2nd largest, 3898 km2), and the 2020 Creek Fire (5th largest, 1537 km2) representative of this trend. Projections of rising temperatures and increased drought (Diffenbaugh et al. 2008) suggest wildfire frequency is expected to escalate (Dale et al. 2001; Swetnam et al. 2009; Keeley et al. 2011). Areas treated with prescribed burns prior to large, intense fires show lower fire intensity and faster recovery than in non-treated areas (Fernandes and Botelho 2003; Jordan 2003). Knowledge of forest structure and fire history prior to Euroamerican settlement can provide baseline data to fire managers in developing fire treatment plans to mitigate fire hazards (Ray et al. 2012; Diaz et al. 2016; Vázquez-Varela et al. 2022). Fire scars from tree-rings in the central and southern Sierra Nevada serve as a fire proxy and indicate frequent fires over the last 1700 years (Swetnam et al. 2009). This frequent fire history is typically attributed to ignitions associated with lightning strikes and the dry summer California climate; however, there is a vast literature documenting Indigenous use of low-intensity surface fires to tend and promote terrestrial resources throughout California (M. K. Anderson 2005; Cuthrell et al. 2012; Knight et al. 2022). Modern prescribed fires essentially mimic the behavior of Indigenous burning practices. Evidence increasingly suggests that the prescribed burns of today aim to replicate traditional ecological knowledge (TEK) practices (Klimaszewski-Patterson and Mensing 2016; Klimaszewski-Patterson et al. 2021), both of which can reduce the risk of high-intensity wildfires (Wu et al. 2023). The State of California recently recognized TEK practices, including fire, as an area of study and potential collaboration to help meet the State’s goals in moderating the effects of forecast climate change on socio-ecological systems (Goode et al. 2018).
While climate clearly plays a role in wildfires in California, TEK practices such as Indigenous use of fire, also play a role. We need a better understanding of the relative importance of both climate and human activity in relation to fire history and forest structure in the pre-fire suppression era to better guide modern fire policy.
California is ecologically designed to burn, with its summer-dry Mediterranean climate and fire-adapted vegetation, and people have used this adaptation to their advantage for millennia. California vegetation including oaks (Quercus spp.), chapparal, serotinous conifers, and annual grasses (Poaceae) are adapted to summer droughts and resprouting/regeneration following fire (Mooney 1977). Despite evidence for natural, climatic fires (e.g., Swetnam 1993), there is also abundant evidence that Native Californians regularly used fire to manage the landscape and its affiliated resources. Fires were deliberately set to burn off woody understory taxa and conifer seedlings, which increased the amount of seasonal forage for large prey animals like mule deer, expanded productive grasslands (small grass seeds were critical to human subsistence in the region), and increased the productivity of patches of important foodstuffs like geophytes and fruiting plants (Stewart 1955; Bean and Lawton 1973; Lewis 1973; M. K. Anderson 2005). Such fires would also have favored the proliferation of shade intolerant, fire-adapted oaks at the expense of shade-tolerant conifers. This would have increased the amount of oaks on the landscape and, critically, the acorns they produce (Dey 2002; Lear and Brose 2002; Klimaszewski-Patterson and Mensing 2016). Fire also facilitated travel by removing dense, woody thickets from travel corridors and would have fostered the growth and proliferation of plants used for making the baskets that were essential to harvesting, processing, and storing the grass seeds and acorns that formed much of the subsistence base for Indigenous people living in the region (M. K. Anderson and Moratto 1996; Fowler 2008; Lake 2013). In sum, fire was essential to generating and maintaining the resource base that helped sustain high aboriginal human population densities; indeed it may have facilitated population growth.
The effect of using TEK fires is suggested by the historical and archaeological literature in California. For example, through historic photographs and written accounts, upon arriving in California, European explorers documented “park-like settings” free of underbrush with mixed-aged and patchy tree cohorts that are consistent with the hypothesis that these settings were maintained using fire (Muir 1894; Lewis 1973; Parker 2002). Grasslands and grassland-adapted fauna in and around archaeological sites in conifer-dominated landscapes point towards anthropogenic creation and maintenance of these habitats (Cuthrell et al. 2012; Cowart and Byrne 2013; Evett and Cuthrell 2013; Lightfoot et al. 2013).
Fire scientists generally attribute such pre-suppression conditions to frequent low-intensity fires that consumed shrubs and seedlings without damaging mature trees. These fire conditions are consistent with both frequent lighting-set climatic fires that are not suppressed and the pattern of TEK fires documented in ethnographies. Most fire reconstruction studies have attributed the source of these fires to climate (e.g. Swetnam et al. 2009), presumably because lightning-induced fire is common during the dry summers of California. This assumption, however, ignores important bodies of evidence that 1) Native Californians actively used fire to increase resource productivity, particularly of acorns from oaks (Quercus spp.); and 2) that populations grew exponentially in the five centuries prior to Euroamerican contact (Bettinger 2015; Klimaszewski-Patterson et al. 2021).
During the Late Holocene, population density increased and diets trended towards more costly-to-process plant resources (e.g., small grass seeds, acorns, smaller-bodied fauna) (Beaton 1991; Wohlgemuth 1996). By 1550 C.E., the aboriginal hunter-gatherers of California arguably had the highest population densities in North America north of Mesoamerica, higher even than the agriculture-based societies of the eastern woodlands and American Southwest. This situation has led some to argue that, though reliant on natural as opposed to domestic resources, economic practices in California rivaled those in many precolonial agricultural societies and allowed for the growth and maintenance of such large populations (Baumhoff 1963; Lightfoot and Simmons 1998; Hull 2012).
Though sporadically occupied since perhaps as early as the terminal Pleistocene (Pryor and Weisman 1991; Price 2002), permanent occupation of the Sierra Nevada began ca. 5000–3000 B.C.E., after meadows stabilized across the Sierra and when many residential bases were first occupied (Wood 1975; Hull and Moratto 1999; Hull 2007). Most researchers, however, argue that the most intensive use of the Sierra Nevada was associated with the onset of intensive balanophagy (acorn-eating) around 600 C.E., just before the onset of the Medieval Climate Anomaly (MCA; 900–1200 C.E.) when the bow and arrow was adopted in cismontane California (Basgall 1987; Morgan 2012; Bettinger 2015; Stevens et al. 2019). Balanophagy is a costly subsistence strategy because acorns take considerable effort to process into flour, which then must be repeatedly leached to remove tannins contained in the meal (McCarthy 1993; Morgan 2009; Tushingham and Bettinger 2013). Many argue that switching to this strategy was linked to exponentially increasing population densities (Basgall 1987; Tushingham and Bettinger 2013; Morgan 2015), though others attribute rapidly growing populations in the Late Holocene to the radical social, economic, and political changes brought about by the adoption of the bow and arrow (Bettinger 2015). Whatever the root cause, increasing MCA population densities (Bettinger 1999; Rosenthal et al. 2007; Klimaszewski-Patterson et al. 2021) already reliant on TEK practices would undoubtedly light more fires to manage and maintain important resource patches like acorn-producing oak groves, a hypothesis very recently supported with empirical evidence from the southern Sierra Nevada (Klimaszewski-Patterson and Mensing 2016, 2020).
The impact of human-set versus lightning-strike fires, however, is difficult to differentiate in the paleoecological record (Bowman et al. 2011; Roos et al. 2014), but it is a challenge previous work in California has recently begun to resolve by generating increasingly precise Late Holocene paleoecological datasets (Cowart and Byrne 2013; Crawford et al. 2015; Klimaszewski-Patterson and Mensing 2016). At present only three studies in the southern Sierra Nevada have presented sub-centennial paleoecological reconstructions spanning the last 1500 years that can begin to answer this question. This paper contributes a new analysis from the central Sierra Nevada with the goal to identify periods of either climate-induced fire regimes or TEK-induced fire regimes prehistoric times. We identify these periods through observed effects on forest structure via pollen and sedimentary charcoal reconstructions.
We hypothesize that a noticeable onset of TEK fires will be visible in the pollen record ca.1250 C.E. with the onset of the Little Ice Age (LIA; 1250–1850 C.E.) and persisted until Euroamerican contact. We base this hypothesis on the premise that habitats during the MCA were generally oak dominated and characterized by the type of open-canopy settings that were particularly conducive to producing the resources upon which Indigenous populations relied (R. S. Anderson and Carpenter 1991; R. S. Anderson and Stillick 2013) Though it is possible people also set TEK fires during the MCA (or even earlier), the effect of fire on vegetation during this period is consistent with what climate patterns predict (Klimaszewski-Patterson and Mensing 2016; Klimaszewski-Patterson et al. 2021) meaning it would be extremely difficult, if not impossible, to distinguish anthropogenic from natural pollen signals. Within this context, in order to maintain high MCA population levels, LIA Indigenous groups would have endeavored to preserve the types of open-canopy, MCA habitats upon which these populations relied. Critically, they would have done so during a period of time when such behaviors ought to be recognizable in the paleoecological record. We consequently examine how the timing of vegetation changes at our site, Markwood Meadow, on the Sierra National Forest, relate to other sites in the Sierra Nevada and the implications of TEK fires on existing interpretations of land use and fire regime history.
Methods
Markwood Meadow (Fig. 1) is a mid-elevation (1819 m) wet sedge meadow in the Kings River watershed of the central Sierra Nevada, California (37.08°, -119.22°). The meadow is approximately 21 ha in size and is located in the montane/mixed conifer ecozone. Common tree taxa in the surrounding forest include ponderosa pine (Pinus ponderosa), Jeffrey pine (P. jeffreyi), sugar pine (P. lambertiana), lodgepole pine (P. contorta), incense-cedar (Calocedrus decurrens), California black oak (Quercus kelloggii), and white fir (Abies concolor). The region’s climatic fire regime likely consisted of mixed-intensity fires ignited by lightning strikes, primarily during the summer and fall (van Wagtendonk and Cayan 2008). As such, vegetation in the Sierra Nevada exhibits numerous fire-adapted traits, such as resprouting and serotinous cones.

Study area map showing location and surrounding topography of the 2019 sedimentary core extraction from Markwood Meadow, California, USA
Markwood Meadow and the surrounding forest is part of present-day Sierra National Forest (SNF). The SNF has been continuously occupied by Indigenous populations since at least 6000 B.C.E. The Mono are the Indigenous group that occupies the area today; they have done so since at least 1350, when they are hypothesized to have migrated over the Sierran crest from the Great Basin and established residence in the study area (Morgan 2010). Prior to Euroamerican incursions, the Mono were intensive hunters, fishers, and gatherers whose subsistence centered on acorns, which were collected en masse in autumn, stored in granaries, and processed in bedrock mortars (concavities pecked into granite boulders to facilitate milling acorns into flour) found throughout the SNF (Morgan 2009). Critically, Mono populations set low intensity fires in large part for the same reasons described earlier in the paper: to increase the productivity and distribution of critical resources and resource patches (Goode et al. 2022). At least four archaeological sites containing a total of 50 bedrock mortars are adjacent to Markwood Meadow (Rubinstein 2020). Euroamerican use of the area began ca. 1860 C.E., when cattle and sheep herders began running stock in the area; cattle are regularly grazed in the meadow to this day. Logging began in the 1860s as well and intensified in ensuing decades, with a dam, millpond, and mill built on Stevenson Creek, only 3.6 km northwest of Markwood Meadow, in 1890 C.E. Logging was extensive, to the extent that no old growth forest is in the vicinity of the coring site. Stevenson Creek was re-dammed by Southern California Edison in 1963 C.E. as part of a hydroelectric project, after which the area saw a marked increase in camping and other recreational activities in the vicinity of Markwood Meadow (Jackson et al. 2005). Selective logging was actively observed nearby while we conducted fieldwork. Fire suppression tactics on public and private land have been in place since the early twentieth century (Silcox 1910).
Core recovery
We recovered duplicate overlapping sediment cores (178 cm) from the meadow in July 2019 using a 5-cm diameter modified Livingstone square-rod piston corer. The top 15 cm of each core were extracted using a serrated blade due to the dense root structure of the peat. Cores were immediately extruded and wrapped with plastic wrap into pre-split plastic tubes in the field, then photographed, subsampled, and placed in cold storage at the Continental Scientific Drilling Facility. Ten samples including charcoal, humates, and plant macrofossils were AMS radiocarbon dated (Table 1) and an age-depth model was developed using IntCal20 (Reimer et al. 2020) and the R library Clam version 2.5 (Blaauw 2010).
Pollen and charcoal analysis
To reconstruct vegetation history, we processed 35 sediment samples (0.625 cm3) from the top 59 cm of the core using standard pollen analysis procedures (Faegri and Iverson 1964). We added a known quantity of an exotic tracer, Lycopodium, to calculate pollen concentration and accumulation rates. We counted pollen at 400 × magnification to a minimum of 400 terrestrial grains per sample, and identified pollen grains based on reference material at the Nevada Paleoenvironmental Analysis Laboratory and published keys (Kapp et al. 2000). We performed a constrained cluster analysis using Past (Hammer et al. 2001) to identify statistical breaks in the pollen dataset.
To reconstruct climatic fire history, we chemically treated continuous 1-cm sediment samples of 1.25 cm3 using a 5% sodium metaphosphate solution and sieved each sample to separate charcoal into 500, 250, and 125 µm size fractions (C. J. Long et al. 1998; Whitlock and Larsen 2001). We quantified charcoal per centimeter using 10–40 × magnification with a dissecting microscope, categorizing each piece as wood, grass, or other. Charred material was tracked separately. We calculated charcoal accumulation (CHAR) per centimeter by dividing the quantity of 250–500 µm wood and grass charcoal by years accumulated based on the age-depth model.
Statistical and other analysis
We used independent climate reconstructions for MWM from the North American Drought Atlas (NADA)(Cook et al. 2004, 2008; Herweijer et al. 2007), a 2.5° gridded network which provides 2000 years of annual paleodrought reconstructions from tree-rings via the Palmer Drought Severity Index (PDSI). We used grid cell 047 for reconstructed PDSI values and limited our analysis to the last 1300 years because that is when there is an increase in the number of tree-ring reconstructions used in the PDSI calculation from prior years.
We calculated a vegetation response index (VRI) as a proxy for forest canopy openness. We used the relationship (Abies – Quercus) / (Abies + Quercus) from the pollen reconstruction, where positive values indicate a more closed canopy via fire-sensitive, shade-tolerant Abies and negative values indicate a more open canopy via fire-adapted, shade-intolerant Quercus (Klimaszewski-Patterson and Mensing 2016). We examine the relationship of relative changes in VRI against climatic expectations (Fig. 2).

Conceptual model showing relationship between climate (solid line) and vegetation response index (VRI) regarding a climate-driven (left) versus human-influenced (right) signals
To examine the temporal relationships between forest canopy openness, fire, and climate we interpolated each reconstruction (VRI, CHAR, and PDSI respectively) using a scaled, standardized 50-year smooth spline. We standardized each spline as the value minus the mean of the reconstruction, divided by the standard deviation before plotting for visual analysis.
As an alternative to the VRI-CHAR-PDSI analysis, we used a principal components analysis (PCA) to independently explore temporal patterns of pollen taxa assemblage variations. PCA is an unsupervised multivariate statistics technique used to reduce and summarize variables from n-dimensions into two-dimensional space. The ordination rearranges datapoints in this space to elucidate major directions of variation. Points plotted closer together have similar values in those two PCA dimensions (Shi 1993), and each axis explains variance in that dimension. This technique allows us to use a broader spectrum of the pollen assemblage to test against trends identified in the VRI-CHAR-PDSI analysis. We reduced the original dataset to terrestrial pollen sums greater than 4%, removed strongly correlated taxa, standardized the remaining assemblage, performed the PCA in R (R Core Team 2023), and visualized the plot using the R library ggbiplot (Vu 2011). We expect datapoints during MCA and LIA to distinctly cluster if forest composition is climatically-driven and to intersperse if TEK fires are maintaining MCA-like habitats during the LIA. We expect modern datapoints (1850–2019) to cluster separately because of known human impacts in the forest, including fire suppression and logging.
Results
The top 59 cm of the sediment core captured data from 650 to 2019 C.E. (Fig. 3). Six of the ten radiocarbon samples dated from 600 to modern. All radiocarbon samples from Table 1 were included in the age-depth model except samples PSUAMS9787 and PSUAMS9788 which returned as post-bomb era, and PSU11197, which is clearly a contaminate.

Classical age-depth model for Markwood Meadow (MWM), California, USA using Clam version 2.5 (Blaauw 2010). Blue bars indicate 2-sigma radiocarbon date, grey background indicates 95% confidence interval
Pollen
Samples were counted to a minimum of 400 terrestrial grains except for four samples with low concentration which were counted to a total of 300 grains. Pollen resolution averaged one sample every 40 years with the greatest time between samples being 100 years (in the sixteenth century) and the least being 10 years (in the second half of the nineteenth century). Pollen was plotted in terms of the percentage of the total pollen sum (Fig. 4). Cyperaceae is the dominant plant taxa in the immediate vicinity of the coring location today, and to avoid biasing the record toward a hyper-local signal, was excluded from the pollen sum when calculating percentage. A total of 44 taxa were identified in the record of which 13 contributed at least 1% to the total pollen sum (Fig. 4).

Pollen percentage diagram outlining the 13 most common taxa in the Markwood Meadow, California, USA sedimentary record. Colors correspond to life form (green = tree, orange = herbaceous, pink = shrub/fern. Zonation (right-hand column and horizontal grey lines) represent discontinuities in the data identified from constrained cluster analysis
Pinus (yellow) category is likely dominated by ponderosa pine, Jeffrey pine, and lodgepole pine. Pinus (white) is likely primarily sugar pine as the other white pines native to the Sierra Nevada are generally limited to higher elevation. Quercus pollen is primarily from California black oak and huckleberry oak (Q. vaccinifolia). Abies pollen is primarily from white fir (Abies concolor) but could include red fir (A. magnifica). Cupressaceae pollen is primarily California incense-cedar and Juniperus spp. Herbaceous taxa primarily grow in and around the meadow, including wild buckwheat (Eriogonum spp.), umbellifers (Apiaceae, including taxa from genus Lomatium, Oreonana, and Podistera), Cichorieae (tribe of Asteraceae), Asteraceae, and Poaceae. The sunflower family (Asteraceae) represents at least 55 species in the Sierra Nevada and the grass family (Poaceae) represents at least 39 species (Rundel 2011). The sedge family (Cyperaceae) is a large family with a substantial number of taxa common in mid-elevation Sierran meadows (McIlroy and Allen-Diaz 2012), including taxa from the genera Carex, Eleocharis, and Scirpus.
Pollen zonation
Constrained cluster analysis of the Markwood Meadow pollen data revealed three distinct discontinuities and four zones (Fig. 4).
Zone 1: 650 to 875 C.E.
The forest in this zone is characterized by high levels of Pinus (yellow), above 50% of the total pollen sum for much of the zone. Quercus percentages were slightly above average. The meadow in this zone is dominated by Poaceae and Cyperaceae, which both contributed between five and 10% to the total pollen sum. Asteraceae was also above average and Eriogonum and Cichorieae percentages are below average. Trilete ferns were more common than in any other zone.
Zone 2: 875 to 1680 C.E.
In this zone the forest is characterized by below average percentages of Pinus (yellow) (38% zonal average) and above average percentages of Pinus (white) after 1200. The meadow was dominated by Apiaceae, which increased sharply from Zone 1 and averages 21% of the total pollen sum. The latter (top) half of the zone is characterized by elevated Cichorieae and Poaceae, while Cyperaceae reached its minimum.
Zone 3: 1680 to 1850 C.E.
The forest in this zone is characterized by increased Pinus (yellow) and Abies, which represent 57% and 5.5% of the pollen sum, respectively. Both Quercus and Cupressaceae began to increase following 1800. Apiaceae, Asteraceae, and Cichorieae declined over the course of the bottom half of this zone and were replaced by Cyperaceae and Eriogonum, the latter of which increased dramatically after 1800.
Zone 4: 1850 C.E. to present
Pinus (yellow) steadily declined in this zone from a peak of 50% to 30% at the top of the record. Cupressaceae increased dramatically after 1950 and represents nearly 20% of the total pollen sum at the top after having been below 5% for most of the record. Both Pinus (white) and Quercus were more common than average. Eriogonum became a dominant herbaceous taxon in Zone 4 (10% of total pollen sum) and Apiaceae increased steadily beginning at the start of this zone. Poaceae and Cyperaceae were above average levels. Asteraceae and Cichorieae were largely absent from this zone.
VRI vs CHAR vs PDSI
The forest canopy openness-fire-climate analysis (Fig. 5) generally tracked with climate (VRI consistent with PDSI) from 900 to 1500 C.E. but acted opposite to climate between 1550 and 1700 C.E. This deviation indicated an opening forest canopy at a time when climate was cooling and a closing forest canopy was ecologically expected. This disconnect occurred again from the 1800s through to modern times. Sedimentary charcoal (CHAR) appeared disconnected from both climate (PDSI) and forest canopy openness (VRI). The greatest sedimentary charcoal accumulation occurred ca. 1750 – 1830 C.E., a period with an ongoing cooler, wetter climate trend and one of the most closed VRI values in the record.

Comparison of forest canopy openness (pollen-based vegetation response index [VRI]; dashed green line), fire (charcoal accumulation rate [CHAR]; orange line), and climate (Palmer Drought Severity Index [PDSI]; grey solid line) trends at Markwood Meadow, California, USA. Pollen and charcoal came from a sediment core extracted from the meadow in 2019. Thick lines represent 50-year smoothed splines. Dots on thin green line show un-splined VRI data points. Data are scaled for presentation, normalizing each dataset’s mean value and standard deviation to allow for comparison. LIA: Little Ice Age (1200 – 1850). MCA: Medieval Climate Anomaly (900 – 1200). Modern (1850 – 2019)
The PCA ordination is presented as a two-dimensional correlation biplot (Fig. 6) of the two dominant principal components (PC). Five PCs accounted for 97.8% of the variation in the pollen data points. Axes 1 (PC1; λ1 = 0.37) and 2 (PC2; λ2 = 0.27) captured 64.5% of the total variance of the data and are represented in the biplot. The remaining axes 3 (λ3 = 0.13), 4 (λ4 = 0.10), 5 (λ5 = 0.10), and 6 (λ6 = 0.02) had low eigenvalues and were not explored. PC1 is associated with forest composition of arboreal versus herbaceous taxa, with strong weights from Asteraceae, Abies, and Pinus. PC2 is associated with forest canopy openness and disturbance events, with the strongest weight from Cupressaceae, followed by Quercus, Pinus, and Poaceae. In the biplot, the modern period (1850 – 2019 C.E.) distinguished itself sharply from the MCA and LIA. Because of the strength of Modern responses, MCA data points were restricted to a narrow band, still showing disturbance or openness along PC2. 1640 C.E. was a clear LIA outlier that showed strong MCA-like pollen contributions on both PC axes. LIA data points showed a spread of PC1 scores which indicated variable periods of stronger arboreal or herbaceous influence, and generally lower PC2 scores. 1830 and 1840 C.E. are LIA outliers that showed more commonality with the modern period.

Principal components (PC) analysis ordination diagram of the six most prevalent, non-correlated sedimentary pollen taxa (dark brown) from Markwood Meadow, California, USA. Distance biplot shows dated pollen sample points, where dates are mean calibrated age assigned from the age-depth model rounded to the nearest decade. Modern (1850–2019; dark grey dates). LIA = Little Ice Age (1200–1850; blue dates). MCA = Medieval Climate Anomaly (900–1200; orange dates). Pre-MCA (400–900; purple dates)
Discussion
While climate appears to be the main driver of forest composition during the MCA and onset of the LIA, we argue that the opening of forest structure after 1550 C.E. was driven by TEK fires, during a time of cooler and wetter climate (Fig. 5). This is evident whether using an informed analysis of open vs closed indicator taxa (via VRI) or an unsupervised multivariate statistical technique on a broader pollen assemblage (via PCA).
When compared against climate, VRI shows an opening canopy trend began after 1550 C.E. and continued until 1750 C.E. Another opening trend briefly occurred ca. 1800 C.E., before Euroamerican settlement began in the late 1850s C.E. The forest canopy closed rapidly as settlement began and Indigenous populations became increasingly constrained and TEK fires were either prevented or discontinued. A rapid canopy opening shift in the late 19th and early twentieth centuries is attributed to intensive logging (Laudenslayer, Jr and Darr 1990).
The PCA ordination (Fig. 6) independently shows agreement with VRI interpretations that TEK fires maintained periods of MCA-like open forest composition during the LIA. This is especially evident at 1640 C.E., but half the LIA data points (1270 – 1350, 1590 – 1750 C.E.) show an influence in the biplot from herbaceous taxa, likely indicating low-intensity fire disturbances. These periods are consistent with times when the VRI was more positive than expected climatically. The early 1800s C.E. show a strong Quercus influence, consistent with the VRI. The modern period shows a marked contrast with the previous 1000 years, with logging and related activities resulting in a strong arboreal signal seen via increases in Cupressaceae and Quercus and sharp decrease of Pinus influence. This demonstrates that modern forest management policy and logging continues to maintain a post-1850s C.E. novel composition state. Selective harvesting and a lack of regular fire disturbances allow for increased Cupressaceae populations, and may prevent forest restoration to precolonial composition conditions.
The Modern period clearly shows the most pronounced anthropogenic influence on the forest surrounding Markwood Meadow, evident in any of our pollen analysis interpretations. While the influences of TEK fires on forest structure are not as dramatic as are those in the Modern period, they are nonetheless anthropogenic. TEK fires did not fundamentally alter forest structure as significantly as logging, grazing, fire suppression, and built infrastructure because TEK fires sought to replicate and enhance natural systems of fire for resource management and mitigation of fire hazards (J. W. Long et al. 2021; Goode et al. 2022). They are a cryptic and oft dismissed part of Sierran fire history that can be illuminated via pollen analysis.
We attempted to use macroscopic sedimentary charcoal to reconstruct a more local fire history, but as with other sub-centennially analyzed central and southern Sierran paleoecological sites reporting both pollen and charcoal reconstructions from the same sediment core (R. S. Anderson and Stillick 2013; Klimaszewski-Patterson and Mensing 2016, 2020; Klimaszewski-Patterson et al. 2018), the results demonstrated that quantifying charcoal by itself is not a good tool to summatively reconstruct all sources of fire. The impact of fire intensity on charcoal production has been found to be reflected in the relationship between fire intensity and charcoal deposition in multiple modern ecosystems (Duffin et al. 2008; Hennebelle et al. 2020).Charcoal is produced by incomplete combustion of plant material at high temperatures comparable to those of fast-moving, medium to high-severity wildfires, while ash is produced by the complete combustion of plant material and consumptions of charcoal (Hudspith et al. 2018), common during low, slow fires. Charcoal is preferentially preserved in the sedimentary record based on taphonomic processes, while ash is not accounted for in the charcoal record. Further, charcoal may be a signal of both extra-local or regional fires, especially at large lakes, and there are questions regarding transport and primary versus secondary deposition (Higuera et al. 2007; Hudspith et al. 2018; Snitker 2018; Vachula and Cheung 2021). In addition, recent research has found that low-intensity fires reduce the risk of high-intensity wildfires by up to six years past application (Wu et al. 2023), which can also reduce the opportunity for charcoal production. The inability of quantified charcoal to elucidate a comprehensive fire history at Markwood Meadow is best evidenced by relatively low charcoal accumulation rate over 1000 years, from 650 – 1650 C.E. This period includes the MCA, which is regionally known for increased fire frequency (Swetnam et al. 2009), yet we see no evidence for increased fire in the charcoal. Furthermore, the largest peak of charcoal accumulation occurred ca. 1750 – 1800 C.E., during the LIA, and coincided with closing forest canopy conditions in both the VRI and PCA analyses. Nor did low charcoal accumulation explain the open canopy conditions ca. 1650 C.E. Sedimentary charcoal did not explain forest changes at Markwood Meadow, and instead we argue that fire effects are best captured in the pollen record, when compared to expectations derived from climatic models.
Regional context
In the broader scope of the central and southern Sierra Nevada (Fig. 6), Markwood Meadow showed evidence of TEK fires contemporaneous with forest changes at previously analyzed paleoecologic sites. Change in forest canopy openness (as VRI) at Markwood Meadow is concurrent with similar changes at Wawona Meadow (R. S. Anderson and Stillick 2013) and Swamp Lake (Smith and Anderson 1992), which are historically viewed as Sierra Miwok territory, and at Trout Meadow (Klimaszewski-Patterson and Mensing 2020), which is considered within the extant hunting range of the Tubatulabal, an Indigenous group on the Kern Plateau (Voegelin 1938; Harvey 2018). Holey Meadow (Klimaszewski-Patterson and Mensing 2016) and Woski Pond (R. S. Anderson and Carpenter 1991), within the settlement portions of territory for the Tubatulabal and Sierra Miwok, respectively, showed evidence for an earlier onset of TEK fires, beginning ca. 1450 and 1250 C.E.. This trans-Sierran perspective shows that TEK fires were used broadly across Indigenous territories and not only near local, more intensively-used sites.
Further, regional tree-ring and charcoal-based studies of fire frequencies throughout the central and southern Sierra Nevada (Swetnam et al. 2009; Taylor et al. 2016) showed little agreement with fire history inferences from pollen records (Fig. 7). For the period of record across all compared sites, forest canopies are most open when regional fire frequencies are reported as low. For example, pollen regionally indicates an opening forest canopy during low regional fire frequencies ca. 1600 – 1650 C.E. and 1850 – 2000 C.E. (Swetnam 1993; Swetnam et al. 2009), and closing canopy conditions at Markwood Meadow, Wawona Meadow, and Holey Meadow ca. 1775 – 1850 C.E. when Taylor et. al (2016) indicate increased regional fire frequency. Consistent with Taylor et. al (2016), sedimentary charcoal at Markwood Meadow increases ca. 1750 – 1850 C.E., but this increase is not reflected in the pollen. For sedimentary charcoal records, this is probably because low-intensity fires, such as TEK fires, are more likely to produce ash rather than charcoal, and any produced charcoal likely remains in-situ rather than becoming entrained and deposited in meadows and lakes. Further, different fuel types produce different quantities of charcoal, the relative quantity and mass varying by species (Hudspith et al. 2018). It is possible that sedimentary charcoal disproportionately represents certain taxa that are abundant in mixed-conifer forests (e.g. Pinus spp.) over other less-abundant taxa (e.g. Quercus spp.), and additional work is necessary to quantify and calibrate both charcoal sources (Hudspith et al. 2018) and the fire severity under which charcoal formed (Roos and Scott 2018). For fire scars, discrepancies between fire frequency and forest canopy openness could relate to (1) regional fires that are more likely to be climatically-driven than a specific local application of fire as a management tool, (2) the fact that fires must be hot enough to damage but not kill a tree, and thus do not capture all low-intensity fires, and (3) the possibility that previously scarred trees have been removed or consumed by subsequent fires and thus remaining fire-scarred trees represent survivor bias. This is not to say that fire scars do not capture TEK fires, but that fire scars can also be caused by climate-driven fires. Because the source of fire cannot be determined from a fire scar, and because TEK fires may replicate low intensity fires that do not necessarily scar trees, fire scar studies do not necessarily capture the complete record of TEK fires. Regionally, high fire frequency was not necessarily associated with more open forest canopies as inferred by pollen, even during the MCA. Disconnects with sedimentary charcoal, both at Markwood Meadow and regionally, provides further evidence that TEK fires are identifiable via changes in pollen assemblage in relation to climate, and not necessarily via charcoal or fire-scar studies.

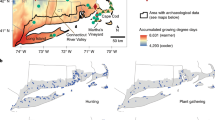
Regional comparison of published sub-centennial paleoecologic sites in the central and southern Sierra Nevada. Map in the top left panel shows the location of each site (colored squares) represented in the forest canopy openness-climate response graphs (right main panel). Solid grey line represents 50-year spline of climate, derived from the North American Drought Atlas (Cook et al. 1999; Herweijer et al. 2007). Colored dash lines represent 50-year spline of the vegetation response index (VRI) representing forest canopy openness. Thin colored line indicates pollen-derived VRI datapoints. Regional fire frequency study indicated by dotted (high frequency) or hashed (low frequency) overlay (Swetnam 1993; Swetnam et al. 2009). Vertical dashed lines indicate broad climatic time periods. LIA = Little Ice Age, MCA = Medieval Climate Anomaly. Bottom left panel shows areal context of the Sierra Nevada range (cream) in California. SQNF = Sequoia National Forest. SNF = Sierra National Forest. YNP = Yosemite National Park
Conclusions
This study supports the hypothesis that frequent, low-intensity TEK fires contributed to the pre-suppression conditions of open forests prior to the 1850s C.E., and impacted fire regimes across a broad stretch of mid-elevation forests in the central and southern Sierra Nevada. Indigenous people had an impact on forest composition through their traditional use of fire. Changes to forests via low to moderate intensity TEK fires are best seen via pollen, and not sedimentary charcoal.
Pollen analysis further demonstrates that human impacts, such as the addition (and removal) of TEK fires, can alter forest canopy openness and composition within decades. This knowledge can be useful when discussing current forest management strategies of mechanical thinning and prescribed burning to deal with the accumulation of forest biomass and litter that occurred during the era of fire suppression policy. Pollen analysis also demonstrates that selective logging continues to alter overall composition, such as increasing Cupressaceae beyond precolonial levels. Further, the paleorecord can be utilized to identify what present-day forests and fire patterns could have been like without 100 years of fire suppression, which may aid fire management in setting appropriate forest composition targets. This is because the pollen record already shows the impact of lightning and human-set fires on forests and fire patterns under different climatic and socio-ecological conditions.
These data support interpretations that Indigenous people in the Sierra Nevada used fire for the better part of the last millennium (and likely longer) to create a more open forest that increased access to, and habitat for, economic resources. They demonstrate that human actions, such as intentionally-set low intensity fires, can buffer against changes in vegetation composition due to climate. This traditional purpose aligns with modern forestry goals, and thus modern public land managers could consider capitalizing on traditional ecological knowledge, in consultation with local tribal groups, to improve forest health, reduce biomass, and moderate against warming climate projections. Further, modern public land managers can use pollen-based reconstructions to ensure that forests are not pushed towards novel states and compositions. These strategies could potentially help to mitigate the effects of forecast climate change and resulting megafires on forests and socio-ecological systems in a more comprehensive manner.
Availability of data and materials
The datasets generated and/or analyzed during the current study are available in the Neotoma Paleoecology Database repository: http://www.neotomadb.org, or are available from the corresponding author on reasonable request.
Abbreviations
- CHAR:
-
Charcoal accumulation rate
- LIA:
-
Little Ice Age
- MCA:
-
Medieval Climate Anomaly
- PCA:
-
Principal components analysis
- PDSI:
-
Palmer Drought Severity Index
- TEK:
-
Traditional ecological knowledge
- VRI:
-
Vegetation response index
References
Anderson, M. Kat. 2005. Tending the wild: Native American knowledge and the management of California’s natural resources. Berkeley: University of California Press.
Anderson, M. Kat, and Michael J. Moratto. 1996. Native American land-use practices and ecological impacts. In Sierra Nevada Ecosystem Project: final report to Congress. 2:187–206. Davis: University of California, Centers for Water and Wildland Resources.
Anderson, R. Scott., and Scott L. Carpenter. 1991. Vegetation change in Yosemite Valley, Yosemite National Park, California, during the Protohistoric Period. Madroño 38: 1–13.
Anderson, R. Scott., and R.D. Stillick. 2013. 800 years of vegetation change, fire and human settlement in the Sierra Nevada of California, USA. The Holocene 23: 823–832. https://doi.org/10.1177/0959683612471985.
Basgall, Mark E. 1987. Resource intensification among hunter-gatherers: Acorn economies in prehistoric California. Research in Economic Anthropology 9: 21–52.
Baumhoff, Martin A. 1963. Ecological determinants of Aboriginal California populations. University of California Publications in American Archaeology and Ethnology 49: 155–236. https://doi.org/10.1525/aa.1964.66.3.02a00360.
Bean, Lowell John, and Harry W. Lawton. 1973. Some explanations for the rise of cultural complexity in Native California with comments on proto-agriculture and agriculture. In Patterns of Indian burning in California: ecology and ethnohistory. ed. Lowell John Bean, v–xlvii. Ramona, CA: Ballena Press Anthropological Papers No. 1.
Beaton, John M. 1991. Extensification and intensification in central California prehistory. Antiquity 65: 946–952.
Bettinger, Robert L. 1999. What happened in the Medithermal. In Models for the Millennium: Great Basin Anthropology Today, ed. Charlotte Beck, 62–74. Salt Lake City: The University of Utah Press.
Bettinger, Robert L. 2015. Orderly anarchy: Sociopolitical evolution in Aboriginal California. Berkeley: University of California Press.
Blaauw, Maarten. 2010. Methods and code for “classical” age-modelling of radiocarbon sequences. Quaternary Geochronology 5. Elsevier B.V: 512–518. https://doi.org/10.1016/j.quageo.2010.01.002.
Bowman, David M. J. S., Jennifer Balch, Paulo Artaxo, William J. Bond, Mark A. Cochrane, Carla M. D’Antonio, Ruth DeFries, et al. 2011. The human dimension of fire regimes on Earth. Journal of Biogeography 38: 2223–2236. https://doi.org/10.1111/j.1365-2699.2011.02595.x.
Cook, Edward R., David M. Meko, David W. Stahle, and Malcolm K. Cleaveland. 1999. Drought reconstructions for the continental United States. Journal of Climate 12: 1145–1163. https://doi.org/10.1175/1520-0442(1999)012%3c1145:DRFTCU%3e2.0.CO;2.
Cook, Edward R., Connie A Woodhouse, C Mark Eakin, David M Meko, and David W Stahle. 2004. Long-term aridity changes in the western United States. Science (New York, N.Y.) 306: 1015–8. https://doi.org/10.1126/science.1102586.
Cook, Edward R., Connie A. Woodhouse, C. Mark Eakin, David M. Meko, and David W. Stahle. 2008. North American summer PDSI reconstructions, version 2a Boulder, CO: NOAA/NGDC Paleoclimatology Program.
Cowart, Alicia, and Roger Byrne. 2013. A paleolimnological record of Late Holocene vegetation change from the Central California coast. California Archaeology 5: 337–352. https://doi.org/10.1179/1947461x13z.00000000018.
Crawford, Jeff N., Scott A. Mensing, Frank K. Lake, and Susan R. Zimmerman. 2015. Late Holocene fire and vegetation reconstruction from the western Klamath Mountains, California, USA: A multi-disciplinary approach for examining potential human land-use impacts. The Holocene 25: 1341–1357. https://doi.org/10.1177/0959683615584205.
Cuthrell, Rob Q., C. Striplen, Mark G. Hylkema, and Kent G. Lightfoot. 2012. A land of fire: anthropogenic burning on the Central Coast of California. In Contemporary Issues in California Archaeology. ed. Terry L. Jones and J. E. Perry, 153–174. Walnut Creek, CA: Left Coast Press.
Dale, Virginia H., Linda A. Joyce, Steve McNulty, Ronald P. Neilson, Matthew P. Ayres, Michael D. Flannigan, Paul J. Hanson, et al. 2001. Climate change and forest disturbances. BioScience 51: 723–734. https://doi.org/10.1641/0006-3568(2001)051[0723:CCAFD]2.0.CO;2.
Dey, Daniel 2002. Fire history and postsettlement disturbance. In Oak Forest Ecosystems: Ecology and Management for Wildlife, ed. W.J. McShea and W.M. Healy, 46–59. Baltimore: John Hopkins University Press.
Diaz, John M., Toddi Steelman, and Branda Nowell. 2016. Local ecological knowledge and fire management: What does the public understand? Journal of Forestry 114. Society of American Foresters: 58–65. https://doi.org/10.5849/jof.14-026.
Diffenbaugh, Noah S., Filippo Giorgi, and Jeremy S. Pal. 2008. Climate change hotspots in the United States. Geophysical Research Letters 35: 1–5. https://doi.org/10.1029/2008GL035075.
Duffin, K.I., L. Gillson, and K.J. Willis. 2008. Testing the sensitivity of charcoal as an indicator of fire events in savanna environments: Quantitative predictions of fire proximity, area and intensity. The Holocene 18: 279–291. https://doi.org/10.1177/0959683607086766.
Evett, Rand R., and Rob Q. Cuthrell. 2013. Phytolith evidence for a grass-dominated prairie landscape at Quiroste Valley on the central coast of California. California Archaeology 5: 265–290.
Faegri, Knut, and Johs Iverson. 1964. Textbook of Pollen Analysis. Copenhagen: Scandinavian University Books.
Fernandes, Paulo M., and Hermínio S. Botelho. 2003. A review of prescribed burning effectiveness in fire hazard reduction. International Journal of Wildland Fire 12: 117–128. https://doi.org/10.1071/WF02042.
Fowler, Catherine S. 2008. Historical perspectives on Timbisha Shoshone land management practices, Death Valley, California. In Case Studies in Environmental Archaeology, ed. E.J. Reitz, C.M. Scarry, and S.J. Scudder, 43–57. New York: Springer Science+Business Media, LLC.
Goode, Ron W., Stephanie Farish Beard, and Christina Oraftik. 2022. Putting fire on the land: The indigenous people spoke the language of ecology, and understood the connectedness and relationship between land, water, and fire. Journal of California and Great Basin Anthropology 42: 85–95.
Goode, Ron W, Shasta Gaughen, Marissa Fierro, Don L. Hankins, Keir Johnson-Reyes, Beth R. Middleton, Teri Red Owl, Randy Yonemura, Stephanie Lucero, and Jana Ganion. 2018. Summary report from Tribal and Indigenous communities within California. In California’s Fourth Climate Change Assessment. Sacramento: California Governor’s Office of Planning and Research, California Natural Resources Agency, and the California Energy Commission.
Hammer, Øyvind., David A T. Harper, and Paul D. Ryan. 2001. Past: Paleontological statistics software package for education and data analysis. Palaeontologia Electronica 4: 1–9.
Harvey, David C. 2018. Core-periphery dynamics in the Kern River watershed. Hunter Gatherer Research 4. Liverpool University Press: 557–587. https://doi.org/10.3828/hgr.2018.33.
Hennebelle, Andy, Julie C. Aleman, Adam A. Ali, Yves Bergeron, Christopher Carcaillet, Pierre Grondin, Josianne Landry, and Olivier Blarquez. 2020. The reconstruction of burned area and fire severity using charcoal from boreal lake sediments. The Holocene 30: 1400–1409. https://doi.org/10.1177/0959683620932979.
Herweijer, Celine, Richard Seager, Edward R. Cook, and Julien Emile-Geay. 2007. North American droughts of the last millennium from a gridded network of tree-ring data. Journal of Climate 20: 1353–1376. https://doi.org/10.1175/JCLI4042.1.
Higuera, Philip E., Matthew Edward Peters, Linda B. Brubaker, and Daniel G. Gavin. 2007. Understanding the origin and analysis of sediment-charcoal records with a simulation model. Quaternary Science Reviews 26: 1790–1809. https://doi.org/10.1016/j.quascirev.2007.03.010.
Hudspith, Victoria A., Rory M. Hadden, Alastair I. Bartlett, and Claire M. Belcher. 2018. Does fuel type influence the amount of charcoal produced in wildfires? Implications for the fossil record. Edited by Leyla Seyfullah. Palaeontology 61: 159–171. https://doi.org/10.1111/pala.12341.
Hull, Kathleen L. 2007. The Sierra Nevada: archaeology in the range of light. In California Prehistory: Colonization, Culture and Complexity, ed. K.A. Jones and T.L. Klar, 177–190. New York: Altamira Press.
Hull, Kathleen L. 2012. A land of many people: population dynamics as context and catalyst. In Contemporary Issues in California Archaeology, ed. T.L. Jones, J.E. Perry, and W. Creek, 73–92. CA: Left Coast Press.
Hull, Kathleen L., and Michael J Moratto. 1999. Archaeological Synthesis and Research Design, Yosemite National Park, California. Vol. 21. Yosemite Research Center Publications in Anthropology.
Jackson, Thomas L, Christopher Thomas Morgan, D DeJoseph, and P Quick. 2005. Draft Inventory and Evaluation of Cultural Resources, Southern California Edison Company Big Creek Hydroelectric System Relicensing (FERC Project Nos. 67, 120, 2085, 2175). Submitted to Southern California Edison Company, Rosemead, California. Pacific Legacy, Inc., Santa Cruz, California.
Jordan, Timothy A. 2003. Ecological and Cultural Contributions of Controlled Fire Use by Native Californians: A Survey of Literature. American Indian Culture and Research Journal 27: 77–90. https://doi.org/10.17953/aicr.27.1.2032485783835762.
Kapp, Ronald O., Owen K. Davis, and James E. King. 2000. Ronald O Kapp’s Pollen and Spores. College Station, TX: American Association of Stratigraphic Palynologists.
Keeley, Jon E., Janet Franklin, and Carla D’Antonio. 2011. Fire and invasive plants on California landscapes. In The Landscape Ecology of Fire, ed. Donald McKenzie, Carol Miller, and Donald A. Falk, 193–221. Ecological Studies. Dordrecht: Springer Science+Business Media B.V. https://doi.org/10.1007/978-94-007-0301-8.
Kilgore, Bruce M., and D. Taylor. 1979. Fire history of a sequoia mixed-conifer forest. Ecology 60: 129–142.
Klimaszewski-Patterson, Anna, and Scott A. Mensing. 2016. Multi-disciplinary approach to identifying Native American impacts on Late Holocene forest dynamics in the southern Sierra Nevada range, California, USA. Anthropocene 15. Elsevier B.V.: 37–48. https://doi.org/10.1016/j.ancene.2016.04.002.
Klimaszewski-Patterson, Anna, and Scott A. Mensing. 2020. Paleoecological and paleolandscape modeling support for pre-Columbian burning by Native Americans in the Golden Trout Wilderness Area, California, USA. Landscape Ecology 35: 2659–2678. https://doi.org/10.1007/s10980-020-01081-x.
Klimaszewski-Patterson, Anna, ChristopherT. Morgan, and Scott A. Mensing. 2021. Identifying a pre-Columbian Anthropocene in California: A paleoecological and archaeological study of fire, vegetation change, and human settlement. Annals of the American Association of Geographers 11: 784–794. https://doi.org/10.4324/9781003208211-18.
Klimaszewski-Patterson, Anna, Peter J. Weisberg, Scott A. Mensing, and Robert M. Scheller. 2018. Using paleolandscape modeling to investigate the impact of Native American–set fires on pre-Columbian forests in the southern Sierra Nevada, California, USA. Annals of the American Association of Geographers 108: 1635–1654. https://doi.org/10.1080/24694452.2018.1470922.
Knight, Clarke A., M. Lysanna Anderson, Jane Bunting, Marie Champagne, Rosie M. Clayburn, Jeffrey N. Crawford, Anna Klimaszewski-Patterson, et al. 2022. Land management explains major trends in forest structure and composition over the last millennium in California’s Klamath Mountains. Proceedings of the National Academy of Science 119: e2116264119. https://doi.org/10.1073/pnas.
Lake, Frank K. 2013. Historical and cultural fires, tribal management and research issue in Northern California: Trails, fires and tribulations. Occassion: Interdisciplinary Studies in the Humanities. 5: 1–22.
Laudenslayer, Jr., F. William, and Herman H. Darr. 1990. Historical effects of logging on the forests of the Cascade and Sierra nevada ranges of California. Transactions of the Western Section of the Wildlife Society 26: 12–23.
Van Lear, David H, and Patrick .H. Brose. 2002. Fire and oak management. In Oak Forest Ecosystems: Ecology and Management for Wildlife, ed. William J. McShea and William M. Healy, 269–279. Baltimore: John Hopkins University Press.
Lewis, Henry T. 1973. Patterns of Indian burning in California: ecology and ethnohistory. Edited by L.J. Bean. Vol. 1. Ramona, CA: Ballena Press Anthropological Papers No. 1.
Lightfoot, Kent G., Rob Q. Cuthrell, Cristie M. Boone, Roger Byrne, Andreas S. Chavez, Laurel Collins, Alicia Cowart, et al. 2013. Anthropogenic burning on the Central California coast in Late Holocene and Early Historical times: Findings, implications, and future directions. California Archaeology 5: 371–390.
Lightfoot, Kent G., and William S. Simmons. 1998. Culture contact in protohistoric California: Social contexts of Native and European encounters. Journal of California and Great Basin Anthropology 20: 138–170.
Long, Colin J., Cathy Whitlock, Patrick J. Bartlein, and Sarah H. Millspaugh. 1998. A 9000-year fire history from the Oregon Coast Range, based on a high-resolution charcoal study. Canadian Journal of Forest Research 28. NRC Research Press: 774–787. https://doi.org/10.1139/cjfr-28-5-774.
Long, Jonathan W, Frank K Lake, and Ron W Goode. 2021. The importance of Indigenous cultural burning in forested regions of the Pacific West, USA. Forest Ecology and Management 500. Elsevier B.V. https://doi.org/10.1016/j.foreco.2021.119597.
Martin, Robert E., and D. B. Sapsis. 1992. Fires as agents of biodiversity: pyrodiversity promotes biodiversity. In Proceedings of the Symposium on Biodiversity of Northwestern California, 150–157.
McCarthy, H. 1993. A political economy of Western Mono acorn production. Davis: University of California.
McIlroy, S.K., and B.H. Allen-Diaz. 2012. Plant community distribution along water table and grazing gradients in montane meadows of the Sierra Nevada Range (California, USA). Wetlands Ecology and Management 20: 287–296. https://doi.org/10.1007/s11273-012-9253-7.
Miller, J.D., H.D. Safford, M. Crimmins, and A.E. Thode. 2009. Quantitative evidence for increasing forest fire severity in the Sierra Nevada and southern Cascade Mountains, California and Nevada, USA. Ecosystems 12: 16–32. https://doi.org/10.1007/s10021-008-9201-9.
Mooney, H.A., ed. 1977. Convergent evolution in Chile and California Mediterranean climate ecosystems. Stroudsberg: Dowden, Hutchinson and Ross Inc.
Morgan, Christopher T. 2009. Optimal foraging patterns in the Sierra Nevada, Alta California. California Archaeology 1. Taylor and Francis Ltd.: 205–226. https://doi.org/10.1179/cal.2009.1.2.205.
Morgan, Christopher T. 2010. Numic expansion in the southern Sierra Nevada. Journal of California and Great Basin Anthropology 30: 157–174.
Morgan, Christopher T. 2012. Modeling modes of hunter-gatherer food storage. American Antiquity 77: 714–736.
Morgan, Christopher T. 2015. Is it intensification yet? Current archaeological perspectives on the evolution of hunter-gatherer economies. Journal of Archaeological Research 23: 1–51.
Muir, John. 1894. The mountains of California. Berkeley: Ten Speed Press.
Parker, Albert J. 2002. Fire in Sierra Nevada forests: Evaluating the ecological impact of burning by Native Americans. In Fire, Native Peoples, and the Natural Landscape, ed. Thomas R. Vale, 233–267. Washington, D.C.: Island Press.
Price, Barry 2002. A Clovis point from the Sierra National Forest. Society for California Archaeology Newsletter 36: 15–16.
Pryor, John, and Russell Weisman. 1991. Archaeological investigations at the Skyrocket Site, CA-Cal-629/63, the Royal Mountain King Mine Project. Proceedings of the Society for California Archaeology 4: 159–191.
Ray, Lily A., Crystal A. Kolden, and F. Stuart Chapin. 2012. A case for developing place-based fire management strategies from traditional ecological knowledge. Ecology and Society 17. Resilience Alliance. https://doi.org/10.5751/ES-05070-170337.
Reimer, Paula J., William E.N. Austin, Edouard Bard, Alex Bayliss, Paul G. Blackwell, Christopher Bronk Ramsey, Martin Butzin, et al. 2020. The IntCal20 Northern Hemisphere Radiocarbon Age Calibration Curve (0–55 cal kBP). Radiocarbon 62. Cambridge University Press: 725–757. https://doi.org/10.1017/RDC.2020.41.
Roos, Christopher I., David M. J. S. Bowman, J.K. Balch, Paulo Artaxo, William J. Bond, Mark A. Cochrane, Carla M. D’Antonio, et al. 2014. Pyrogeography, historical ecology, and the human dimensions of fire regimes. Journal of Biogeography 41: 833–836. https://doi.org/10.1111/jbi.12285.
Roos, Christopher I., and Andrew C. Scott. 2018. A comparison of charcoal reflectance between crown and surface fire contexts in dry south-west USA forests. International Journal of Wildland Fire 27: 396. https://doi.org/10.1071/WF17139.
Rosenthal, J.S., G.G. White, and M.Q. Sutton. 2007. The Central Valley: A view from the catbird’s seat. In California Prehistory: Colonization, Culture and Complexity, ed. T.L. Jones and K.A. Klar, 147–164. Lanham: AltaMira Press.
Rubinstein, Emily M. 2020. Land use and settlement patterns in the Central and Southern Sierra Nevada: A comparative study. Reno: University of Nevada, Reno.
Rundel, Philip W. 2011. The diversity and biogeography of the alpine flora of the Sierra Nevada, California. Madroño 58: 153–184. https://doi.org/10.2307/26476397.
Shi, Guang R. 1993. Multivariate data analysis in palaeoecology and palaeobiography – a review. Paiaeogeography Palaeoclimatology. Palaeoecology 105: 199–234.
Silcox, F.A. 1910. Fire prevention and control on national forests. In Yearbook of Department of Agriculture. Washington: Government Printing Office.
Smith, Susan J., and R. Scott Anderson. 1992. Late Wisconsin paleoecologic record from Swamp Lake, Yosemite National Park, California. Quaternary Research 38: 91–102.
Snitker, Grant. 2018. Identifying natural and anthropogenic drivers of prehistoric fire regimes through simulated charcoal records. Journal of Archaeological Science 95. Elsevier Inc.: 1–15. https://doi.org/10.1016/j.jas.2018.04.009.
Stevens, Nathan E., Adrian R. Whitaker, and Jeffrey S. Rosenthal. 2019. Bedrock mortars as indicators of territorial behavior in the Sierra Nevada. Quaternary International 518. Elsevier Ltd: 57–68. https://doi.org/10.1016/j.quaint.2017.11.008.
Stewart, Omer C. 1955. Forest and grass burning in the Mountain West. Southwestern Love 21: 5–9.
Swetnam, Thomas W. 1993. Fire history and climate change in giant sequoia groves. Science 262: 885–889. https://doi.org/10.1126/science.262.5135.885.
Swetnam, Thomas W., Christopher H. Baisan, Anthony C. Caprio, Peter M. Brown, R. Ramzi Touchan, Scott Anderson, and Douglas J. Hallett. 2009. Multi-millennial fire history of the Giant Forest, Sequoia National Park, California, USA. Fire Ecology 5: 120–150. https://doi.org/10.4996/fireecology.0503120.
Taylor, Alan H., Valerie Trouet, Carl N. Skinner, and Scott L. Stephens. 2016. Socioecological transitions trigger fire regime shifts and modulate fire–climate interactions in the Sierra Nevada, USA, 1600–2015 CE. Proceedings of the National Academy of Sciences 113: 13684–13689. https://doi.org/10.1073/pnas.1609775113.
Tushingham, Shannon, and Robert L. Bettinger. 2013. Why foragers choose acorns before salmon: Storage, mobility, and risk in aboriginal California. Journal of Anthropological Archaeology 32: 527–537. https://doi.org/10.1016/j.jaa.2013.09.003.
Vachula, R.S., and A.H. Cheung. 2021. Late Neogene surge in sedimentary charcoal fluxes partly due to preservation biases, not fire activity. Palaeogeography, Palaeoclimatology, Palaeoecology 567: 110273. https://doi.org/10.1016/j.palaeo.2021.110273.
Vázquez-Varela, Carmen, José M. Martínez-Navarro, and Luisa Abad-González. 2022. Traditional fire knowledge: A thematic synthesis approach. Fire 5. MDPI. https://doi.org/10.3390/fire5020047.
Voegelin, E.W. 1938. Tubatulabal Ethnography Anthropological Records, 2nd ed. Berkeley: University of California Press.
Vu, Vincent Q. 2011. ggbiplot: A ggplot2 based biplot. http://github.com/vqv/ggbiplot.
van Wagtendonk, Jan W., and Daniel R. Cayan. 2008. Temporal and spatial distribution of lightning strikes in California in relation to large-scale weather patterns. Fire Ecology 4: 34–56.
Whitlock, Cathy, and Chris Larsen. 2001. Charcoal as a fire proxy. In Tracking Environmental Change Using Lake Sediments, ed. John P. Smol, H. J. B. Birks, and W. M. Last. Vol. 3. Dordrecht: Kluwer Academic Publishers.
Wilkin, Kate, Lauren Ponisio, Danny L. Fry, Brandon M. Collins, Tadashi Moody, and Scott L. Stephens. 2021. Drivers of understory plant communities in Sierra Nevada mixed conifer forests with pyrodiversity. Fire Ecology 17. Springer Science and Business Media Deutschland GmbH: 1–18. https://doi.org/10.1186/s42408-021-00111-6.
Wohlgemuth, Eric. 1996. Resource intensification in prehistoric California: Evidence from archaeobotanical data. Journal of California and Great Basin Anthropology 18: 81–103.
Wood, Spencer Hoffman. 1975. Holocene stratigraphy and chronology of Mountain Meadows, Sierra Nevada, California. California Institute of Technology.
Wu, Xiao, Erik Sverdrup, Michael D. Mastrandrea, Michael W. Wara, and Stefan Wager. 2023. Low-intensity fires mitigate the risk of high-intensity wildfires in California’s forests. Science Advances 9: eadi4123. https://doi.org/10.1126/sciadv.adi4123.
Acknowledgements
We would like to thank R. Vail for quantifying sedimentary charcoal, and C. Briem, J. White, and D. Max for assisting with fieldwork. Special thanks to Sierra National Forest employees (Jeff Irwin, Annette Lambert) for site identification and facilitating access. Additional thanks to Christopher Roos and an anonymous reviewer whose comments and suggestions helped improve and strengthen this manuscript.
Funding
This study was supported by the National Science Foundation (GSS 1740918, “Fire, Vegetation Change, and Human Settlement”).
Directorate for Social,Behavioral and Economic Sciences,1740918,Scott A. Mensing
Author information
Authors and Affiliations
Contributions
AK-P collected field data, created age-depth model, supervised charcoal quantification, completed analyses, created graphics, and wrote the manuscript. TD collected field data, processed samples, quantified pollen, assisted with analysis, and assisted in writing the manuscript. CM assisted in writing the manuscript and advised on the study. SM helped write the manuscript and has overseen the overarching project for more than 10 years. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Klimaszewski-Patterson, A., Dingemans, T., Morgan, C.T. et al. Human influence on late Holocene fire history in a mixed-conifer forest, Sierra National Forest, California. fire ecol 20, 3 (2024). https://doi.org/10.1186/s42408-023-00245-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s42408-023-00245-9