
Abstract
Background
Straw mulching is one of the most common treatments applied immediately post fire to reduce soil erosion potential and mitigate post-fire effects on water quality, downstream property, and infrastructure, but little is known about the long-term effects on vegetation response. We sampled six fires that were mulched between 9 and 13 years ago in western US dry conifer forests. We compared understory plant species diversity and abundance, tree seedling density and height by species, and fractional ground cover on mulched and unmulched paired plots.
Results
Mulch did not influence understory plant diversity, species richness, or fractional ground cover. However, on mulched plots, tree seedlings grew taller faster, especially on north-facing aspects, and there was slightly more graminoid cover. Mulch did not affect overall tree seedling density, but there were fewer ponderosa pine (Pinus ponderosa Lawson & C. Lawson) and more Douglas-fir (Pseudotsuga menziesii [Mirb.] Franco) in mulched areas, especially on south-facing slopes.
Conclusions
Managers will be able to weigh the long-term implications of mulching against the short-term reductions in soil erosion potential. While there are many concerns about vegetation suppression and exotic species introduction from using straw mulch, our study suggests that the long-term effects are subtle 9 to 13 years after post-fire mulching.
Resumen
Antcedentes
El mantillo de paja es uno de los tratamientos más comunes que se aplican inmediatamente en el post-fuego, para reducir el potencial de erosión y mitigar los efectos del post-fuego en la calidad del agua, en las propiedades que se encuentran aguas abajo donde se produjo el evento de fuego, y en infraestructuras, aunque muy poco se conoce sobre sus efectos en la respuesta de la vegetación a largo plazo. Muestreamos seis áreas post-fuego a las que se les había adicionado mantillo de paja entre 9 a 13 años antes, en bosques secos de coníferas en el oeste de los EEUU. Comparamos la diversidad y abundancia de las especies del sotobosque, la densidad de plantines y altura por especie, y la fracción de cobertura del suelo en parcelas apareadas con y sin mantillo.
Resultados
El mantillo de paja no influenció la diversidad vegetal, la riqueza de especies o la fracción de suelo cubierto. Sin embargo, los plantines cubiertos por mantillo crecieron más rápido en altura, especialmente en las laderas con exposición norte, y también las áreas con mantillo tuvieron un pequeño incremento en la cobertura de graminoides. El mantillo no afectó la densidad de plantines en general, pero en general hubo menos plantines de pino ponderosa (Pinus ponderosa Lawson & C. Lawson) y más plantines de pino oregón (Pseudotsuga menziesii [Mirb.] Franco) en áreas con mantillo, en especial en las laderas de exposición sur.
Conclusiones
Los gestores de recursos podrían entonces ponderar las implicancias del mantillo a largo plazo con las reducciones a corto plazo en el potencial de erosión de los suelos. Aunque existen muchas preocupaciones sobre la supresión de la vegetación y la introducción de especies exóticas por el uso del mantillo de paja en post-fuegos, nuestro estudio sugiere que los efectos a largo plazo son casi imperceptibles entre los 9 a 13 años posteriores a su aplicación.
Similar content being viewed by others


Introduction
Large wildfires have been increasing in both size and frequency in recent decades (Westerling et al. 2006; Dennison et al. 2014; Westerling 2016). After a wildfire, the loss of vegetation biomass can greatly reduce the stability of soil (Robichaud 2005; Wagenbrenner et al. 2006). Fires that burn with high severity (based on loss of organic matter; Keeley 2009) increase the potential for soil erosion, loss of nutrients, and water repellency (Neary et al. 1999). Agricultural straw mulch greatly decreases soil movement (Wagenbrenner et al. 2006) until vegetation can establish in abundance (Dodson and Peterson 2010; Robichaud et al. 2013). Mulching is one of the most effective emergency stabilization techniques to use post fire (Robichaud et al. 2000; Bautista et al. 2009). Mulch stabilizes soil, reduces sediment movement, prevents loss of soil productivity, and reduces risk of flooding (Bautista et al. 1996; Robichaud et al. 2000; Dean 2001; Robichaud and Ashmun 2013; Williams et al. 2014). Due to expense, mulching is used as a strategic treatment of areas of high soil erosion potential and risk of loss of downstream values (i.e., road infrastructure, aquatic habitats, etc.; Bautista et al. 2009; Williams et al. 2014).
Agricultural straw mulch alters local habitats, sometimes enough that some plant species may not be able to establish after a fire. Some species, such as ponderosa pine (Pinus ponderosa Lawson & C. Lawson), more successfully germinate on bare mineral soil (Curtis and Lynch 1965; Rother and Veblen 2016). The physical barrier caused by mulching forces plants to expend extra energy to push through the thick organic layer and can prevent new seeds from reaching bare ground (Facelli and Pickett 1991). Mulching also moderates soil temperature by blocking solar radiation, and acts as an insulator at night to prevent freezing (Facelli and Pickett 1991). This may result in a longer growing season (Facelli and Pickett 1991) and reduced evaporation, which increases soil moisture (Mulumba and Lal 2008). These changes combine to increase microbial activity and available nitrogen for developing plants (Berryman et al. 2014).
Many short-term studies of vegetation response to post-fire mulching have been done, with mixed results. In examining only mulched sites on the 2005 Tripod Fire (Washington, USA), Dodson and Peterson (2010) found that plant cover, species richness, and tree seedling density were all positively associated with straw mulching in a dry mixed conifer forest. However, when mulch reached over 70% cover, it negatively affected vegetation (Dodson and Peterson 2010). In the ponderosa pine forests burned in the 2000 Bobcat Fire in Colorado, USA, Wagenbrenner et al. (2006) found reduced sediment movement, increased mulch cover, and more vegetation cover on mulched plots compared to unmulched plots. Positive impacts on lodgepole pine (Pinus contorta Douglas ex Loudon) seedling recruitment have also been shown, although the association was weak (Wright and Rocca 2017). Conversely, others have found that mulching can inhibit plant establishment and introduce non-native plant species (Beyers 2004; Kruse et al. 2004; Robichaud 2005). Kruse et al. (2004) found that two years post fire, mulched areas had a higher occurrence of non-native species, less overall vegetation cover, and reduced conifer tree seedling density when compared to similar unmulched areas. If these differences persist, mulching could be a tradeoff between short-term reduction of soil erosion potential and altered vegetation recovery trajectory for many years post fire.
Initial vegetation response can have lasting influences on vegetation trajectories. The first species to colonize post fire will often persist for decades. This was observed 10 years after fire by Abella and Fornwalt (2015), and 29 years after fire by Engel and Abella (2011). Areas burned with high severity, which is the primary target of mulching, have reduced ecosystem resistance to fire and reduced resiliency (i.e., a diminished ability to return to the pre-fire state; Abella and Fornwalt 2015). Of particular concern is that high-severity wildfire could cause large areas to be converted from forest to shrub or grass communities (Savage and Mast 2005; Johnstone et al. 2016), especially if tree seedlings fail to regenerate (Stevens-Rumann et al. 2018). By altering the physical characteristics of a site’s microclimate, it is possible that mulching could change which species will be able to colonize first and persist (Morgan et al. 2014; Morgan et al. 2015).
Studies conducted on vegetation response from mulching have been short-term (usually less than three years), on single fires, and have yielded mixed results (Dodson and Peterson 2010; Morgan et al. 2015). This study will help us to understand long-term vegetation recovery following mulching. Managers will be able to assess the long-term implications of mulching and better weigh any potential impacts on post-fire vegetation against the benefits of reducing soil erosion potential.
Our objectives were to assess the differences in understory vegetation cover and composition and tree seedling density between mulched and unmulched areas on multiple large fires. Specifically, our hypotheses were:
- (1)
understory plant species cover, richness, and diversity would be higher on mulched sites; and
- (2)
tree seedling density would be lower on mulched sites, but tree seedling height growth would be higher.
Methods
Study areas
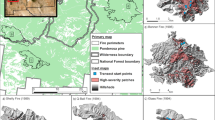
We focused on wildfires in the US Interior West where a minimum of 40 ha of agricultural straw mulch was aerially applied in steep areas burned with high severity (Fig. 1). Fires were chosen from two widespread dry forest types (ponderosa pine-dominated and dry mixed conifer), as described by LANDFIRE existing vegetation type (Table 1, LANDFIRE 2008). The Burned Area Emergency Response (BAER) Burned Area Reports Database (Robichaud 2017) was used to select fires for which mulch had been applied 9 to 13 years prior to our field sampling. Mulch was aerially applied within weeks of fire containment. We then confirmed the location of treated areas with local forest managers; however, mulch application rate was unknown. We used Monitoring Trends in Burn Severity (MTBS; https://www.mtbs.gov/) maps for areas burned with high severity. On the ground, areas burned with high severity usually correspond to greater than 70% tree basal area killed (Agee 1993); those areas burned with high soil burn severity are identified as having loss of ground cover, surface discoloration due to ash or oxidation, loss of soil structure, consumption of fine roots, and possible formation of a water repellent layer (Parson et al. 2010).

Study areas with the six large wildfires sampled in summer in 2015 and 2016 across the United States Interior West. Albers conical projection
Plot setup
We collected data from five fires in the summer of 2015 and from the Cascade Fire in summer 2016 (Fig. 1, Table 1). We sampled 58 plots (29 plot pairs) on areas where no tree seedlings were planted and that had no other BAER treatment aside from aerial application of agricultural straw. To ensure that plot pairs represented a range of conditions on each fire, we sampled in four strata, including elevation (high and low) and transformed aspect (or trasp, high and low). Strata were created for each fire individually, where the mean value of all pixels within the fire perimeter was the designated threshold between high and low. Not all four strata were sampled on all fires. The number of plots within each fire varied by number and size of mulch units, and all were at least 30 m but no more than 2 km from a road (Table 1). Plot centers were randomly located within the strata, and for each mulched plot, the paired unmulched plot was established in the closest yet same elevation and trasp strata. To avoid edge effects, both mulched and unmulched plots were located at least 60 m from edges of strata, roads, and trails. Within each plot, there were five subplots as described by Morgan et al. (2015): one subplot at the center of each plot, with four other subplots located 30 m away in orthogonal directions (Fig. 2). The first peripheral subplot was placed directly upslope, with the others 90°, 180°, and 270° from the upslope azimuth.

Sampling plots each had five subplots. At each 1 m2 quadrat (black square), each understory plant was identified, and percent canopy cover of each species was ocularly estimated and recorded. Cover fractions of green vegetation, NPV, soil, and rock were also recorded. At each 5.6 m radius plot (circles), total tree seedling density was recorded, as well as total and yearly height growth on a subsample of seedlings. A total of 29 pairs were installed on six fires across the US Interior West in 2015 and 2016
Understory plant measurements
At each subplot, we ocularly estimated ground cover fractions and percent canopy cover by species within a 1 m2 quadrat. The categories for cover fractions were green vegetation, non-photosynthetic vegetation (NPV, woody debris and other dead plant material), soil, and rock. Cover fractions for these classes in each subplot added up to 100%. For many subplots, the sum of percent canopy cover by all species exceeded 100%, as there were often multiple layers of vegetation. Plant species not identified in the field were given a unique code, collected, and later identified with help from experts at the University of Idaho Stillinger Herbarium. Each plant was assigned to a growth form (shrub, forb, graminoid, or other) based on the USDA plant database (USDA NRCS 2017). In cases for which a species had multiple growth forms, the most common growth form was assigned. Cover by growth form was computed as the total for all species within that growth form for each subplot averaged across all subplots.
Tree seedling measurements
Tree seedling density by species was measured within a 5.6 m radius circle from the center of the subplot (Fig. 2). Each subplot was divided into quarters for counting seedlings in succession, such that all seedlings within a quarter were tallied until the number tallied after finishing the quarter(s) exceeded six seedlings of the most common species. Heights were measured on a subset of seedlings starting in a randomly selected quarter of the subplot that had been tallied and included additional tallied quarters until at least six of the most dominant species had been measured. To avoid selecting tree seedlings from the same clump, seedlings sampled for height measurements were spread throughout the quarter(s). This ad-hoc approach helped to distribute seedlings sampled for height across different microsites and size classes. Total height and distance between terminal bud scars (the resulting scar after terminal bud scales fall off) were measured to estimate yearly height growth (Urza and Sibold 2013). Seedlings without nodes (i.e., less than one year old) were not counted. Counts were converted to density for a given subplot using the recorded area sampled. Seedling tallies for each species in each subplot were summed and converted to density (stems ha−1) by dividing by total area sampled (one or more quarters). Total seedling density as well as density of each tree species across the five subplots were then aggregated to the plot level.
Analyses
Measured variables were averaged for the five subplots in each plot. All statistical analyses were done in R version 3.3.1 (R Core Team 2016). The same fixed terms were used for all multivariate analyses of variance (MANOVA) and linear mixed effects models: differenced normalized burn ratio (dNBR), elevation, transformed aspect, post-fire water deficit, treatment, and fire. Differenced normalized burn ratio is the change in the normalized burn ratio caused by the fire, considered an index of burn severity (Key and Benson 2006). Transformed aspect (trasp) is a cosine transformation of aspect from degrees to a continuous variable, where 0 represents 30° north-northeast-facing (NNE) and 1 is 210°, a south-southwest-facing (SSW) aspect. Thus, plots with low trasp are typically cooler and wetter, and those with high trasp are warmer and drier. Post-fire water deficit was calculated as the difference between actual evapotranspiration (AET) and potential evapotranspiration (PET) during the summer months (June, July, August) for the first three years after each fire. Data were compiled from 800 m PRISM data, corrected topographically as described in Abatzoglou (2013). Data were then aggregated as a single variable for ease of modeling. Treatment was a binary factor, mulched or unmulched. The Fire variable was a factor that encompassed a wide variety of local climatic and soil variables that collectively influence seedlings and understory plants at a particular fire but that differ greatly between the widely geographically separated fires sampled. The random effect, or grouping variable, was Pair; this collapsed environmental variation between mulched and unmulched plots within each pair such that we could focus on the differences caused by mulch. In the mixed modeling framework, predictor variables were removed if the coefficient of correlation between pairs of predictor variables was 0.7 or higher (Dormann et al. 2013).
Prior to any statistical test, data were tested for violations of assumptions.
We calculated both Shannon-Weiner diversity index and species richness for each plot using the vegan package (Oksanen et al. 2017). The total number of species from all five subplots (richness) was summed along with average percent canopy cover to calculate diversity at the plot level. A paired t-test was used to assess differences in diversity and richness between plot pairs (Zuur et al. 2009).
A multivariate analysis of variance (MANOVA) was used on the three growth forms being tested (grass, forb, shrub), as well as the cover classes (vegetation, NPV, soil, rock). We analyzed the differences between paired mulched and unmulched plots in this analysis to more sensitively quantify the effect of mulching. The cover value for each cover class or growth form on a mulched plot was subtracted from the value for the unmulched plot of the same pair. Where the overall mulching treatment was significant (α ≤ 0.1), we examined differences for each growth form using a linear mixed effects model with the predictor variables described above. The robust mixed effects model framework allows for deeper investigation into individual variables and interaction effects. Model selection was based on an improved Akaike information criterion (AIC) score, when shown significantly different from a more complex model with an ANOVA test (Zuur et al. 2009). The base model started with all available variables, interactions between mulch treatment and other fixed variables, and then non-significant fixed variables were dropped sequentially. To conform to linear mixed effects assumptions, unequal variance and spatial autocorrelation were accounted for in the model structure using the R package nlme (Pinheiro et al. 2016) as opposed to any transformations (O’Hara and Kotze 2010). This facilitated easier interpretations of model coefficients.
Because some plots did not have any tree seedlings present, a two-step modeling process was used to ask two separate questions. First, “does mulch influence whether or not tree seedlings are present?” To answer this question, we analyzed binary presence or absence data in a mixed effects modeling framework like that used in growth form and fractional analyses. Second, “where seedlings are present, does mulch influence their density?” For this question, we analyzed tree seedling density values greater than zero for the two most common species sampled: ponderosa pine and Douglas-fir (Pseudotsuga menziesii [Mirb.] Franco). Analysis of seedling density was done on all plots where ponderosa pine was present in either the mulched or unmulched plots in a pair, and then on all plots where Douglas-fir was present in either the mulched or unmulched plots in a pair.
Tree height analysis was more complex, as there were variable tree heights at any given plot, with a different combination of species, ages, and number of seedlings. We used a mixed effect model; in addition to the fixed variables mentioned above, seedling age and species were also included in this model. Only trees that established within the first three years after the fire were used in the model to address the question of how mulch affects tree growth. Most of the mulch likely dissipated after three years, and the intention was to look at the impact that mulch had on growth. Due to the large amount of possible combinations (treatment, species, and fire having multiple levels), it was not possible to run every interaction all at once. Instead, a stepwise process was used, including one interaction each time.
Results
Diversity and species richness
A total of 352 species were sampled across the six fires, with 247 on mulched plots, and 248 on unmulched plots. Neither plant species richness (P = 0.440, t = −0.7841, df = 28) nor diversity (P = 0.748, t = −0.3246, df = 28) differed between mulched and unmulched plot pairs (n = 29). Species richness ranged from 8 to 37 species on mulched plots (median of 22), and 9 to 33 on unmulched plots (median of 21). Shannon-Weiner diversity ranged from 1.9 to 3.4 on mulched plots (median of 2.9), and from 2.0 to 3.3 on unmulched plots (median of 2.8). While diversity varied among fires, there was no discernable pattern across all fires (Fig. 3), or by average summer precipitation (Table 1).

Plant diversity and species richness differences for mulched and unmulched plot pairs on six fires across the US Interior West in 2015 and 2016; data are displayed as box plots of differences between values for plot pairs. Any point above the zero line represents a higher value on the mulched plot, while a value below the line is a higher value on the unmulched plot of the pair. A Differenced plant species diversity for mulched and unmulched plot pairs on each fire. B Differenced species richness (number of plant species) for plot pairs on each fire. Circles represent suspected outliers, datapoints beyond 1.5 times the interquartile range
Plant growth form cover
Graminoid canopy cover was slightly higher (2%) on mulched than on unmulched plots (P = 0.004, MANOVA), but cover of forbs and shrubs did not differ between mulched and unmulched plot pairs across the six fires (P = 0.209 for forbs, P = 0.144 for shrubs). The best-fitting mixed effects model predicting graminoid cover had treatment, three-year water deficit, and fire as predictor variables (Table 2), with mulched plots having slightly higher graminoid cover than unmulched plots (Fig. 4).

Differences in cover fractions and cover by growth form of understory plants on six fires across the US Interior West in 2015 and 2016; values are displayed as box plots of differences between mulched and unmulched plot pairs. Any point above the zero line represents a higher value on the mulched plot, while a value below the line is a higher value on the unmulched plot of the pair. A Differenced cover fractions for green and non-photosynthetic vegetation (NPV) and soil for plot pairs by fire. B Differenced percent canopy cover of three main vegetation cover groups by plot pair. Three subplots on the Tripod Complex contained residual mulch; this was added to the NPV class for that plot and the values were 11, 3, and 0.4%. Circles represent suspected outliers, datapoints beyond 1.5 times the interquartile range
Ground cover fractions
Cover fraction values varied greatly between mulched and unmulched plot pairs, and between fires (Fig. 4). The Ricco Fire had the most vegetation (ranging from 60 to 74%, median of 68% cover) and the Hayman Fire had the least (ranging from 27 to 49%, median of 37%). Percent bare soil also varied by fire, ranging widely within the study, from 0% (median of 2%, high of 6%) on an unmulched plot on the Ricco Fire, to 43% (median 29%, low 16%) on an unmulched plot on the Hayman Fire. Overall, the Hayman Fire had the most bare soil cover at 16% to 43%; all others were below 16%.
Mulch was present but with very low or zero cover on plots sampled. When mulch was encountered in a subplot, it was treated as NPV for cover fractions. This happened on only three subplots on the Tripod Fire. Mulch was seen on all fires except on the Ricco Fire; however, it often took determined effort to find it so many years after these areas were mulched.
Tree seedling density and height growth
Tree seedling density by species varied greatly within and among fires (Fig. 5). As an example, density on the Ricco Fire ranged from 0 to 102 stems ha−1 (median of 37 stems ha−1), with only ponderosa pine present. In contrast, on the Myrtle Creek Fire, tree seedling density ranged from 0 to 23 000 stems ha−1 (median of 6535 stems ha−1), with ponderosa pine, Douglas-fir, Engelmann spruce (Picea engelmannii Parry ex Engelm.), western larch (Larix occidentalis Nutt.), and lodgepole pine (Pinus contorta Douglas ex Loudon) all present (Fig. 5). Of the 58 plots, no tree seedlings were present on 14: 9 mulched plots and 5 unmulched plots. Of these, four plot pairs had no seedlings present, accounting for eight of the 14 plots without seedlings. The Hayman, Ricco, and Myrtle Creek fires all had plots with no tree seedlings present.

Differences in tree seedling density by species calculated as unmulched density subtracted from mulched density on six fires across the US Interior West in 2015 and 2016. Only plot pairs with non-zero values were included. Note scale difference for lodgepole pine compared to the other species. Any point above the zero line represents a higher value on the mulched plot, while a value below the line is a higher value on the unmulched plot of the pair. Circles represent suspected outliers, datapoints beyond 1.5 times the interquartile range
Mulch had no effect on whether tree seedlings were present in a plot based on a binary (0 or 1) model (P = 0.258 for the mulched variable, AIC increased by adding it to the model), nor did mulch influence tree density (P = 0.359) where seedlings were present. However, density of both ponderosa pine and Douglas-fir seedlings varied with the interaction of mulch and aspect. Where Douglas-fir seedlings were present, mulch was significant in the mixed effects model (P = 0.040), and in the ponderosa pine model, mulch was nearly significant (P = 0.086) and still improved the model (Table 2). By itself, mulch decreased density of both ponderosa pine and Douglas-fir; however, there was an interaction between mulch and elevation for ponderosa pine, and between mulch and aspect for Douglas-fir (Table 2). Elevation alone was not a good predictor by itself (P = 0.698), but ponderosa seedling density did increase with elevation in general. However, an interaction effect between elevation and mulch was strongly negative; as elevation increased, ponderosa pine seedling density on mulched plots decreased (Table 2). Douglas-fir had a higher seedling density in mulched plots, and a lower density as aspect became more SSW and drier. However, the interaction effect was enough to overcome increasing trasp and, on SSW aspects that were mulched, there was a higher density of Douglas-fir than on the unmulched plot pairs (Table 2). Douglas-fir and ponderosa pine were each found on just under half of the plots.
Tree seedling height varied with tree seedling age (as indicated by number of terminal bud scars), species, dNBR, trasp, and whether the plot was mulched. Tree seedling height increased with increasing burn severity (as captured by dNBR) and increasing trasp (from NNE to SSW). Growth also depended on tree seedling age, as older seedlings grew more than a younger seedling in a given year (Table 2). Mean tree seedling height was, on average, 5.7 cm greater (P = 0.017) on mulched than on unmulched plots where other variables were accounted for (Table 2). The tree seedling height model was improved by including the interaction effect of mulch with trasp. This interaction was statistically significant at α = 0.1, but not at α = 0.05; however, it did significantly lower the model AIC, so it was included. Seedling height was positively associated with both mulch and increasing trasp. However, as trasp increased on plots that were mulched, the interaction predicted less height, almost enough to negate the influence of mulch all together (Table 2).
Discussion
Mulch had minimal effects on long-term understory vegetation response
Our results suggest that any initial effects of agricultural straw mulch on understory vegetation are not long-term, ecosystem-altering effects. Nine to 13 years post fire, we saw few statistically significant differences in species richness, diversity, canopy cover, or cover fractions between mulched and unmulched plot pairs. The one statistically significant difference that we did find—slightly increased cover of graminoids by about 2% in mulched plots (Table 2)—is not enough to be ecologically significant.
All study plots sampled had less than 50% bare soil cover. Pannkuk and Robichaud (2003) described this as a tipping point for when large soil erosion events can happen post fire. The vegetation on all plots, whether mulched or not, has recovered enough so that we feel that the soil erosion potential is low. Unfortunately, we cannot infer how long it took for vegetation to reach this threshold, nor how long post fire the mulch persisted. In a six-year study on mulched and unmulched areas, Morgan et al. (2014) noted the differences between the first and final years. While plots were initially very different, in many cases, these differences decreased over time.
Introduced weeds are a significant concern of managers and ecologists alike. Although analyzing non-native plants was not a main objective of this study, we found very few non-native plants and none were abundant. While it is now common practice to use certified weed-free straw in post-fire mulching, it is unlikely that the agricultural straw that is used is ever truly weed free (Robichaud et al. 2000). The Hayman Fire is infamous for using cheatgrass (Bromus tectorum L.)-contaminated straw for a majority of its mulching operation (Robichaud et al. 2003), causing an infestation where none had existed previously (Fornwalt et al. 2010). We found cheatgrass in 9 of 14 plots that we sampled on the Hayman Fire; two were not mulched while seven were mulched. However, cheatgrass canopy cover across these plots was low, averaging just 1% on the unmulched plots and under 3% on the mulched plots. It is also possible that, in the 13 years between mulching with cheatgrass-tainted straw and sampling, cheatgrass moved from mulched to unmulched plots, as we attempted to have plot pairs as close as possible within the same physical conditions. When Dodson and Peterson (2010) sampled the Tripod Fire during the second growing season after the fire, they found 14 non-native plant species in mulched areas, five with over 50 occurrences (an occurrence in their study being one individual found in a sampling unit). In contrast, when we sampled the Tripod Fire nine growing seasons after mulching, only one of these weeds, the common dandelion (Taraxacum officinale F.H. Wigg.), was present on five plots (three unmulched and two mulched), with the highest canopy cover at 3%. Although our study design and plot locations were different, it is likely that we would have detected other non-native species if they were still present at such high abundances. Further investigation is warranted to assess the long-term implications of weed introduction in mulched areas. It is also possible that these non-native species were in the area before the fire; however, to our knowledge, there is no pre-fire vegetation data. These results from the Hayman and Tripod fires suggest that, while non-native species may have been a concern initially, they did not persist in large abundances across the landscape.
Mulch resulted in increased tree height growth and altered species composition
Although mulch has no influence on overall tree seedling density, differences in tree species composition suggests the relative importance of mulch, both as a barrier to erosion and as a retainer for soil moisture. In plots where Douglas-fir trees naturally regenerated, seedlings were at a higher density in mulched plots on SSW aspects. In plots where ponderosa pine established, there were fewer in mulched plots than in unmulched plots. Perhaps this is because ponderosa pine requires bare mineral soil to establish (Curtis and Lynch 1965; Rother and Veblen 2016) and is thus less likely to establish until the straw mulch is decomposed. Douglas-fir can establish with some organic matter present over bare mineral soil (Herman and Lavender 1990). We think that the greater soil moisture due to mulch cover is important to Douglas-fir, allowing them to outcompete ponderosa pine (Mulumba and Lal 2008). This could explain why more Douglas-fir than ponderosa pine seedlings were found on the drier aspects of mulched plots, and why mulch suppressed ponderosa pine growth more as elevation increased. However, just because Douglas-fir can establish at higher densities on mulched plots on SSW aspects does not mean that they are better off than their counterparts growing on opposite-facing slopes. Generally, northern aspects of a given area are more productive than southern aspects (Stage and Salas 2007). While we observed that seedlings grew taller on mulched than unmulched plots, the height difference on SSW aspects is less than on NNE aspects (Table 2).
Mulch influences on tree seedling species composition and height growth, while subtle, are potentially long lasting. By moderating soil temperature and increasing soil moisture, mulch created an environment in which Douglas-fir seedlings could establish where they otherwise may not have been able to grow. Trees that grow faster initially are more likely to continue in this way (Mattsson 1997). If the seedlings that established on areas that were mulched continue to grow taller than their counterparts on similar areas without mulch, this could possibly provide a distinct advantage for surviving the next fire if those future saplings also have thicker bark and canopy base heights well above the flames when fires occur (Harper 1977; Odion et al. 2010).
Limitations and future needs
Sampling more plot pairs and years of repeated measurements could strengthen our conclusions by allowing examination of individual fires. While we sampled relatively few plots, the paired design allowed us to draw conclusions about the long-term response to mulch in a variety of environmental conditions.
We do not know initial mulch depth or coverage on the plots we sampled. Aerial application can result in uneven distribution of mulch both within an individual fire area as well as between different fires (Dodson and Peterson 2010; Lewis and Robichaud 2011). Our main hypothesis for differences in tree seedling species composition with mulch is moisture related; however, we did not assess drought stress, soil moisture, or temperature in the field nine to 13 years later, when little remaining mulch was evident.
Management implications
While agricultural straw mulch can reduce soil erosion potential, it is expensive when applied over large areas. As large fires continue to occur, with portions burning with high severity, and with more federal funds going toward fire management and suppression (US GAO 2009; Ellison et al. 2015), strategically targeting use of mulch to areas where high soil erosion potential threatens high values at high risk will be important to avoid similar increases in BAER treatment costs (Robichaud et al. 2015).
We found that mulching with agricultural straw had little effect on vegetation composition nine to 13 years post fire. Vegetation will eventually establish with or without mulch, and all of our plots had far less than 50% exposed soil, and thus are below the threshold for high risk of soil erosion. We found no evidence to suggest that mulch will alter ecosystem function. Managers can expect similar densities of naturally regenerating tree seedlings with or without mulching, but more persistent differences in tree species composition and height growth between mulched and unmulched areas.
Mulching data were not always locationally accurate. Many of the maps and GIS layers that delineated straw mulch treatments planned were not necessarily what actually happened. For example, an area of the Hayman Fire that was supposed to have been mulched had to be disregarded when we found no mulch. On the Tripod Fire, we found mulch in an area that was supposed to have been untreated. These errors illustrate the need for field monitoring and accurate record keeping.
Conclusions
Post-fire mulching with agricultural straw had subtle but potentially long-term impacts on vegetation nine to 13 years post fire. We found greater tree seedling height growth and differences in species composition, as well as higher graminoid cover on mulched plots. Mulching with agricultural straw favored Douglas-fir seedlings, especially on drier, south- and southwest-facing aspects, and hindered ponderosa pine as elevation increased, although total tree seedling densities were similar on mulched and unmulched plots. Mulching increased tree height growth, more so on wetter, more productive aspects. There was, on average, 2% more graminoid cover on mulched plots. Plant species diversity, species richness, and cover fractions were all similar with or without mulching. There were thus no significant differences in vegetation cover or composition between mulched and unmulched plots.
We recommend both short- and long-term monitoring of mulched areas to determine if the trends found in this study might be generalized across multiple vegetation types of the Interior West. While mulching is not appropriate for every area, it is important that managers know that using agricultural straw mulch to reduce soil erosion potential has minimal impacts on long-term vegetation condition and post-fire site recovery.
References
Abatzoglou, J. 2013. Development of gridded surface meteorological data for ecological applications and modelling. International Journal of Climatology 33: 121–131 https://doi.org/10.1002/joc.3413.
Abella, S., and P. Fornwalt. 2015. Ten years of vegetation assembly after a North American mega fire. Global Change Biology 21 (2): 789–802 https://doi.org/10.1111/gcb.12722.
Agee, J. 1993. Fire ecology of Pacific Northwest forests. Washington, D.C.: Island Press.
Bautista, S., J. Bellot, and V.R. Vallejo. 1996. Mulching treatment for postfire soil conservation in a semiarid ecosystem. Arid Soil Research and Rehabilitation 10 (3): 235–242 https://doi.org/10.1080/15324989609381438.
Bautista, S., P.R. Robichaud, and C. Blade. 2009. Post-fire mulching. In Fire effects on soils and restoration strategies, volume 5, Edited by A. Cerda, P.R. Robichaud, 353–372. Enfield: CRC Press.
Berryman, E.M, P. Morgan, P.R. Robichaud, and D. Page-Dumroese. 2014. Postfire erosion control mulches alter belowground processes and nitrate reductase activity of a perennial forb, heartleaf arnica (Arnica cordifolia). USDA Forest Service Research Note RMRS-RN-69. Fort Collins, Colorado, USA: USDA Forest Service, Rocky Mountain Research Station.https://doi.org/10.2737/RMRS-RN-69.
Beyers, J. 2004. Postfire seeding for erosion control: effectiveness and impacts on native plant communities. Conservation Biology 18 (4): 947–956 https://doi.org/10.1111/j.1523-1739.2004.00523.x.
Curtis, J.D., and D.W. Lynch. 1965. Ponderosa pine (Pinus ponderosa Laws.). In Silvics of forest trees of the United States, Edited by H. Fowells,compiler. Pages 417–431. Washington, D.C.: USDA
Dean, A. 2001. Evaluating effectiveness of watershed conservation treatments applied after the Cerro Grande Fire, Los Alamos, New Mexico. Thesis. Tucson: University of Arizona.
Dennison, P., S. Brewer, J. Arnold, and M. Moritz. 2014. Large wildfire trends in the western United States. Forest Service Agriculture Handbook 271. 1984–2011. Geophysical Research Letters 41 (8): 2928–2933 https://doi.org/10.1002/2014GL059576.
Dodson, E., and D. Peterson. 2010. Mulching effects on vegetation recovery following high severity wildfire in north-central Washington State, USA. Forest Ecology and Management 260 (10): 1816–1823 https://doi.org/10.1016/j.foreco.2010.08.026.
Dormann, C., J. Elith, S. Bacher, C. Buchmann, G. Carl, G. Carré, J. Marquez, B. Gruber, B. Lafourcade, P. Leitao, T. Münkemüller, C. McClean, P. Osborn, B. Reineking, B. Schoder, A. Skidmore, D. Zurell, and S. Lautenbach. 2013. Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography 36 (1): 27–46 https://doi.org/10.1111/j.1600-0587.2012.07348.x.
Ellison, A., C. Moseley, and R. Bixler. 2015. Drivers of wildfire suppression costs: literature review and annotated bibliography. University of Oregon Ecosystem Workforce Program Working Paper Number 53. Eugene: University of Oregon Ecosystem Workforce Program.
Engel, E., and S. Abella. 2011. Vegetation recovery in a desert landscape after wildfires: influences of community type, time since fire and contingency effects. Journal of Applied Ecology 48 (6): 1401–1410 https://doi.org/10.1111/j.1365-2664.2011.02057.x.
Facelli, J., and S. Pickett. 1991. Plant litter: its dynamics and effects on plant community structure. The Botanical Review 57 (1): 1–32 https://doi.org/10.1007/BF02858763.
Fornwalt, P.J., M.R. Kaufmann, and T.J. Stohlgren. 2010. Impacts of mixed severity wildfire on exotic plants in a Colorado ponderosa pine–Douglas-fir forest. Biological Invasions 12 (8): 2683–2695 https://doi.org/10.1007/s10530-009-9674-2.
Harper, J.L. 1977. Population biology of plants. London: Academic Press.
Herman, R., and D. Lavender. 1990. Douglas-fir. In Silvics of North America. Volume 1. Conifers. USDA Forest Service Agricultural Handbook 654. Edited by R.M. Burns and B.H. Honkala, technical coordinators. Pages 1080–1108. Washington, D.C.: USDA Forest Service
Johnstone, J.F., C.D. Allen, J.F. Franklin, L.E. Frelich, B.J. Harvey, P.E. Higuera, M.C. Mack, R.K. Meentemeyer, M.R. Metz, G.L. Perry, T. Schoennagel, and M.G. Turner. 2016. Changing disturbance regimes, ecological memory, and forest resilience. Frontiers in Ecology and the Environment 14 (7): 369–378 https://doi.org/10.1002/fee.1311.
Keeley, J. 2009. Fire intensity, fire severity and burn severity: a brief review and suggested usage. International Journal of Wildland Fire 18 (1): 116–126 https://doi.org/10.1071/WF07049.
Key, C.H., and N.C. Benson. 2006. Landscape assessment: sampling and analysis methods. In FIREMON: fire effects monitoring and inventory system. USDA FOrest Service General Technical Report RMS-GTR-164-CD. Edited by D.C. Lutes, technical editor. Pages LA1–LA51. Fort Collins: USDA Forest Service Rocky Mountain Research Station.
Kruse, R., E. Bend, and P. Bierzychudek. 2004. Native plant regeneration and introduction of non-natives following post-fire rehabilitation with straw mulch and barley seeding. Forest Ecology and Management 196 (2): 299–310 https://doi.org/10.1016/j.foreco.2004.03.022.
LANDFIRE. 2008. Existing vegetation type layer. LANDFIRE 1.1.0. USDI Geological Survey. http://landfire.cr.usgs.gov/viewer/. Accessed 28 May 2015
Lewis, S.A., and P.R. Robichaud. 2011. Using QuickBird imagery to detect cover and spread of post-fire straw mulch after the 2006 Tripod Fire, Washington, USA.USDA Forest Service Research Note RMRS-RN-43. Fort Collins: USDA Forest Service Rocky Mountain Research Station https://doi.org/10.2737/RMRS-RN-43.
Mattsson, A. 1997. Predicting field performance using seedling quality assessment. New Forests 13 (1–3): 227–252 https://doi.org/10.1023/A:1006590409595.
Morgan, P., M. Moy, C.A. Droske, L.B. Lentile, S.A. Lewis, P.R. Robichaud, and A.T. Hudak. 2014. Vegetation response after post-fire mulching and native grass seeding. Fire Ecology 10 (3): 49–62 https://doi.org/10.4996/fireecology.1003049.
Morgan, P., M. Moy, C.A. Droske, L.B. Lentile, S.A. Lewis, P.R. Robichaud, A.T. Hudak, and C. Williams. 2015. Vegetation response to burn severity, native grass seeding, and salvage logging. Fire Ecology 11 (2): 31–58 https://doi.org/10.4996/fireecology.1102031.
Mulumba, L., and R. Lal. 2008. Mulching effects on selected soil physical properties. Soil and Tillage Research 98 (1): 106–111 https://doi.org/10.1016/j.still.2007.10.011.
Neary, D., C. Klopatek, L. DeBano, and P. Ffolliott. 1999. Fire effects on belowground sustainability: a review and synthesis. Forest Ecology and Management 122 (1–2): 51–71 https://doi.org/10.1016/S0378-1127(99)00032-8.
O’Hara, R., and D. Kotze. 2010. Do not log-transform count data. Methods in Ecology and Evolution 1 (2): 118–122 https://doi.org/10.1111/j.2041-210X.2010.00021.x.
Odion, D.C., M.A. Moritz, and D.A. DellaSala. 2010. Alternative community states maintained by fire in the Klamath Mountains, USA. Journal of Ecology 98 (1): 96–105 https://doi.org/10.1111/j.1365-2745.2009.01597.x.
Oksanen, J., B.F. Guillaume, M. Friendly, R. Kindt, P. Legendre, D. McGlinn, P. Minchin, R.B. O’Hara, G. Simpson, M. Peter Solymos, E. Szoecs Stevens, and H. Wagner. 2017. vegan: community ecology package. R package version 2, 4–2 https://CRAN.R-project.org/package=vegan Accessed 20 Nov 2017.
Pannkuk, C., and P. Robichaud. 2003. Effectiveness of needle cast at reducing erosion after forest fires. Water Resources Research 39 (12): 1–9 https://doi.org/10.1029/2003WR002318.
Parson, A., P. Robichaud, S. Lewis, C. Napper, and J. Clark. 2010. Field guide for mapping post-fire soil burn severity. USDA Forest Service General Technical Report RMRS-GTR-243. Fort Collins: USDA Forest Service Rocky Mountain Research Station https://doi.org/10.2737/RMRS-GTR-243.
Pinheiro, J., D. Bates, S. DebRoy, D. Sarkar, and R. Core Team. 2016. nlme: linear and nonlinear mixed effects models. R package version 3, 1–128 http://CRAN.R-project.org/package=nlme. Accessed 20 Nov 2017.
PRISM Climate Group. 2004. 30-year normals. http://www.prism.oregonstate.edu/normals/. Accessed 28 May 2016.
R Core Team. 2016. R: a language and environment for statistical computing. Vienna: R Foundation for Statistical Computing https://www.R-project.org/ Accessed 2 Nov 2017.
Robichaud, P.R. 2005. Measurement of post-fire hillslope erosion to evaluate and model rehabilitation treatment effectiveness and recovery. International Journal of Wildland Fire 14 (4): 475–485 https://doi.org/10.1071/WF05031.
Robichaud, P.R. 2017. BAER Burned Area Report Database. USDA Forest Service , Moscow, Idaho, USA. (This is part of the page name--leave it alone.) https://forest.moscowfsl.wsu.edu/cgi-bin/BAERTOOLS/baer-db/index.pl Accessed 28 May 2015.
Robichaud, P.R., and L.E. Ashmun. 2013. Tools to aid post-wildfire assessment and erosion-mitigation treatment decisions. International Journal of Wildland Fire 22 (1): 95–105 https://doi.org/10.1071/WF11162.
Robichaud, P.R., J.L. Beyers, and D.G. Neary. 2000. Evaluating the effectiveness of postfire rehabilitation treatments.USDA Forest Service General Technical Report RMRS-GTR-63. Fort Collins: USDA Forest Service Rocky Mountain Research Station https://doi.org/10.2737/RMRS-GTR-63.
Robichaud, P.R., S.A. Lewis, J.W. Wagenbrenner, L.E. Ashmun, and R.E. Brown. 2013. Post-fire mulching for runoff and erosion mitigation: part I: effectiveness at reducing hillslope erosion rates. Catena 105: 75–92 https://doi.org/10.1016/j.catena.2012.11.015.
Robichaud, P.R., L. MacDonald, J. Freeouf, D. Neary, D. Martin, and L. Ashmun. 2003. Postfire rehabilitation of the Hayman Fire. In Hayman Fire case study: summaryUSDA Forest Service General Technical Report RMRS-GTR-114. Edited by R.T. Graham, technical editor. Pages 293–314. Fort Collins: USDA Forest Service Rocky Mountain Research Station.
Robichaud, P.R., H. Rhee, and S.A. Lewis. 2015. A synthesis of post-fire Burned Area Reports from 1972 to 2009 for western US Forest Service lands: trends in wildfire characteristics and post-fire stabilization treatments and expenditures. Intentional Journal of Wildland Fire 23 (7): 929–944 https://doi.org/10.1071/WF13192.
Rother, M.T., and T.T. Veblen. 2016. Limited conifer regeneration following wildfires in dry ponderosa pine forests of the Colorado Front Range. Ecosphere 7 (12): e01594 https://doi.org/10.1002/ecs2.1594.
Savage, M., and J. Mast. 2005. How resilient are Southwestern ponderosa pine forests after crown fires? Canadian Journal of Forest Research 35 (4): 967–977 https://doi.org/10.1139/x05-028.
Stage, A.R., and C. Salas. 2007. Interactions of elevation, aspect, and slope in models of forest species composition and productivity. Forest Science 53 (4): 486–492.
Stevens-Rumann, C.S., K.B. Kemp, P.E. Higuera, B.J. Harvey, M.T. Rother, D.C. Donato, P. Morgan, and T.T. Veblen. 2018. Evidence for declining forest resilience to wildfires under climate change. Ecology Letters 21: 243–252 https://doi.org/10.1111/ele.12889.
Urza, A., and J. Sibold. 2013. Nondestructive aging of postfire seedlings for four conifer species in northwestern Montana. Western Journal of Applied Forestry 28 (1): 22–29.
US GAO [US Government Accountability Office]. 2009. Wildland fire management: federal agencies have taken important steps forward, but additional action is needed to address remaining challenges.US GAO Testimony GAO-09-906T. Washington, D.C.: US Government Accountability Office
USDA NRCS [USDA Natural Resource Conservation Service]. 2017. PLANTS Database. http://plants.usda.gov. Accessed 29 Jan 2017.
Wagenbrenner, J., L. MacDonald, and D. Rough. 2006. Effectiveness of three post-fire rehabilitation treatments in the Colorado Front Range. Hydrological Processes 20 (14): 2989–3006 https://doi.org/10.1002/hyp.6146.
Westerling, A., H. Hidalgo, D. Cayan, and T. Swetnam. 2006. Warming and earlier spring increase western US forest wildfire activity. Science 313 (5789): 940–943 https://doi.org/10.1126/science.1128834.
Westerling, A.L. 2016. Increasing western US forest wildfire activity: sensitivity to changes in the timing of spring. Philosophical Transactions of the Royal Society B 371 (1696): 20150178 https://doi.org/10.1098/rstb.2015.0178.
Williams, C., F. Pierson, P. Robichaud, and J. Boll. 2014. Hydrologic and erosion responses to wildfire along the rangeland–xeric forest continuum in the western US: a review and model of hydrologic vulnerability. International Journal of Wildland Fire 23 (2): 155–172 https://doi.org/10.1071/WF12161.
Wright, M., and M. Rocca. 2017. Do post-fire mulching treatments affect regeneration in serotinous lodgepole pine? Fire Ecology 13: 139–145 https://doi.org/10.4996/fireecology.130306268.
Zuur, A., E. Ieno, N. Walker, A. Saveliev, and G. Smith. 2009. Mixed effects models and extensions in ecology with R. New York: Springer https://doi.org/10.1007/978-0-387-87458-6.
Acknowledgements
We thank those who helped with fieldwork: M. Schmidt, K. Schmidt, C. Minerich, B. Brown, B. Newingham, D. Truitt, J. Byrne, C. Littlefield, L. Vantieghem, E. Berman, J. Johnson, and D. Hammond. We also thank the Pike-San Isabel, Okanogan-Wenatchee, Malheur, Idaho Panhandle, and Boise National Forests for providing mulch locations and logistical support, as well as S. Larson, formerly of Rocky Mountain Research Station, and S. Gunderson, formerly of Revegetation Services, Inc., for mulching data. We thank J. Abatzoglou for water deficit data, and K. Swanstrom for formatting and compiling data.
Funding
Funding was provided by Joint Fire Science Program Project #14-1-02-27 through a Joint Venture Agreement #14-JV-11221633-112 between the USDA Forest Service Rocky Mountain Research Station and the University of Idaho.
Availability of data and materials
Upon publication, all data will be made available to the public in the USDA Forest Service Rocky Mountain Research Station data archives.
Author information
Authors and Affiliations
Contributions
JDB carried out the analysis with guidance from all coauthors. All authors participated in the design of the study and interpretation of the results. JDB conceived of the study. All authors read, helped write, edit, and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This is not applicable as this study does not involve animal nor human participants, tissue, or data.
Consent for publication
This is not applicable as we did not use data from individual people.
Competing interests
We authors declare that we have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Bontrager, J.D., Morgan, P., Hudak, A.T. et al. Long-term vegetation response following post-fire straw mulching. fire ecol 15, 22 (2019). https://doi.org/10.1186/s42408-019-0037-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s42408-019-0037-9