
Abstract
Background
Serum from systemic lupus erythematosus (SLE) patients has been shown to induce T-lymphocyte (TL) apoptosis. Given that different cells of the immune system display different sensitivity to apoptosis, we set to evaluate the in vitro effect of SLE serum on regulatory T-cells (Treg), Th17, Th1 and Th2 from SLE patients and healthy controls.
Methods
Peripheral blood mononuclear cells from SLE patients or normal controls were exposed to a pool of sera from SLE patients or normal controls. Annexin V was used to label cells in apoptosis or necrosis. Annexin V-labeled Treg, Th17, Th1 and Th2 cells were determined using flow cytometry.
Results
Total CD3 + and CD4 + cells from SLE patients showed higher frequency of spontaneous apoptosis/necrosis, whereas Th1 cells from SLE patients presented reduced spontaneous apoptosis/necrosis rate as compared with cells from controls. Incubation with SLE serum induced increased frequency of apoptotic/necrotic CD3 + , CD4 + and Th2 cells from normal controls or from SLE patients as compared with cultures incubated with normal human serum (NHS) or without human serum at all. Incubation with SLE serum did not increase the apoptosis/necrosis rate in Th1 or Th17 cells. Treg cells from SLE patients were more prone to apoptosis/necrosis induced by SLE serum than Treg cells from normal individuals. Th1, Th2, and Th17 cells presented increased apoptosis rates in cultures without human serum.
Conclusion
Our findings indicate that the serum of patients with active SLE stimulates apoptosis of CD4 + T cells in general and exhibit differentiated effects on CD4 + T-cell subsets.
Similar content being viewed by others


Introduction
Lymphopenia and increased lymphocyte rate are parallel phenomena observed in patients with systemic lupus erythematosus (SLE) [1]. Lymphopenia is part of the day-to-day experience of rheumatologists dealing with lupus patients. The increased availability of circulating apoptotic cells may contribute to tolerance breakdown and eliciting of autoantibodies, as well as immune complex formation and overload of the clearance capacity of the macrophage-endothelial system [2,3,4]. SLE serum has a strong apoptosis-inducing effect on peripheral blood mononuclear cells (PBMC) [5,6,7,8,9,10,11], suggesting that circulating soluble factors may contribute to the increased levels of apoptosis in PBMC in SLE patients. The increased availability of apoptotic cellular material and defective clearance of apoptotic debris may contribute to prolonged exposure of relevant autoantigens to the immune system, favoring the development of autoantibodies and immune complex formation [2].
Different cells of the immune system display different sensitivity to apoptosis [12,13,14,15]. There is evidence that the imbalance between regulatory T cells (Treg) and pro-inflammatory effector T cells significantly contributes to the pathophysiology of some autoimmune diseases, including SLE [16,17,18,19]. Given the evidence that SLE serum stimulates PBMC apoptosis in vitro, we set to evaluate the in vitro effect of SLE serum on Treg, Th17, Th1 and Th2 from healthy controls and SLE patients.
Materials and methods
Patients, controls and biologic samples
Thirty-three patients with active disease (SLEDAI ≥ 6) fulfilling the 2012 SLICC classification criteria for SLE [20] were sequentially recruited at the Outpatient Clinic. Twenty-five healthy controls (students and professional staff) comprised the control group. Peripheral blood was collected after informed consent and the study was approved by the institution ethics committee (UNIFESP CEP 1332/09).
A pool of normal human serum [20 normal blood donors] and a pool of SLE serum (33 patients with active SLE—SLEDAI ≥ 6) were used for challenging PBMC cultures. The pools were inactivated for Complement activity, separated into aliquots and kept at -70 °C until use.
PBMC were isolated by standard density gradient centrifugation and 0.5 × 106 viable PBMC/well were cultured for 24 h in RPMI 1640 (Gibco, Grand Island, USA) supplemented with 2 mM L-glutamine (Gibco), 2 mM sodium pyruvate (Gibco), 100 mg/mL penicillin/streptomycin (Gibco) and 100 mM HEPES (Gibco) in 96-well plates (Costar 3799, Corning®, USA) at 37 °C, 5% CO2 and 96% humidity. In addition, individual wells were supplemented with either: (1) 10% fetal calf serum (Gibco) and no human serum; (2) 20% human serum pool (either from SLE patients or from control donors) and no fetal calf serum; and (3) Camptothencin 5uM (Sigma, Missouri, USA) as a positive control for apoptosis induction. After 24 h, cells were washed in phosphate-buffered saline (PBS) and stained with antibodies that characterize each cell phenotype and Annexin-V, as described below (Additional file 1).
Phenotypic evaluation of peripheral T lymphocytes
Treg cells were identified by staining with APC-Cy3-labeled anti-CD3, V500-labled anti-CD4, PE-Cy7-labeled anti-CD25, and fluorescein isothiocyanate (FITC)-labeled anti-CD127. After 30 min incubation at 4 °C, cells were washed with Pharmingen stain BSA Buffer (Becton Dickinson, San Diego, CA, USA), fixed and permeabilized with FoxP3 fixation/permeabilization buffer (eBioscience, San Diego, CA, USA) and then processed using APC-labeled anti-FoxP3 antibodies (FoxP3 staining kit, eBioscience) according to manufacturer’s instructions. Th17 cells were identified by staining with APC-Cy3-labeled anti-CD3, V500-labeled anti-CD4, PE-Cy7-labeled anti-CCR6, and FITC-labeled anti-CD161. Th2 cells were identified using APC-Cy3-labeled anti-CD3, V500-labeled anti-CD4, and PE-Cy7-labeled anti-CCR4. Th1 cells were identified by staining whit APC-Cy3-labeled anti-CD3, V500-labled anti-CD4, PerCP-Cy5.5-labeled anti-CCR5 and APC-labeled anti-CXCR3. Unless stated otherwise, all monoclonal antibodies were purchased from Becton Dickinson (San Jose, CA, USA) and processed according to the manufacturer’s instructions. Cells were processed in a FACSCanto II™ flow cytometer (Becton Dickinson) and the data obtained were analyzed using the FlowJo software (Tree Star Inc, Ashland, OR, USA) (Additional file 1: Fig. S1). Fluorescence minus one (FMO) was used as gating strategy.
Detection of apoptotic cells
Annexin V was used as a surrogate marker to determine the percentage of cells undergoing apoptosis. PBMC were washed in phosphate-buffered saline (PBS) and 0.5 × 106 cells were incubated with PE-labeled Annexin-V (Annexin V apoptosis detection kit, Becton Dickinson, San Jose, USA) using buffer containing 10 mM HEPES/NaOH (pH 7.4), 0.14 M NaCl, 2.5 mM CaCl2 (Annexin V Binding Buffer—BD).
Statistical analysis
Parameters were expressed as mean and standard error (SE). The distribution pattern of each quantitative parameter was analyzed by the Kolmogorov-Smirnoff test. One-way analysis of variance (ANOVA) was used for variables with normal distribution. Those with non-parametric distribution were analyzed by the Kruskal–Wallis test followed by the Mann–Whitney test or Dunn´s multiple comparison test. Differences were considered significant if p-values were below 0.05. Data analysis was performed using Prism 5.0 software (GraphPad Software, San Diego, CA, USA).
Results
The SLE group comprised 31 women (93.9%) and 2 men (6.1%) with a mean age of 33.3 ± 8.8 years (ranging from 17 to 51), SLEDAI score of 9.4 ± 3.2 (ranging from 6 to 20) and mean disease duration of 5.7 ± 5.8 years (ranging from 1 to 22). The control group consisted of 20 women (80%) and five men (20%) aged 31.6 ± 9.3 years (ranging from 21 to 47). There was no statistically significant difference between SLE and controls regarding age and gender distribution. Table 1 depicts the demographic, clinical, laboratory and therapeutic features of the patients.
In 24 h-cultures without addition of human serum there was a higher percentage of annexin V-labeled CD3+ T cells in samples from SLE patients (4.82% ± 2.58%) as compared to those from normal controls (3.75% ± 3.27%) (p = 0.027) (Fig. 1A). The frequency of annexin V-labeled helper T cells (CD3+CD4+) was also higher in SLE (3.91% ± 1.85%) than in controls (2.84% ± 1.96%) (p = 0.008) (Fig. 1B). In contrast, PBMC from SLE patients presented a lower frequency of annexin V-labeled Th1 lymphocytes (10.37% ± 10.02%) compared to healthy controls (14.24% ± 10.58%) (p = 0.043) (Fig. 1C). The other CD4+ T lymphocyte subsets (Th2, Th17, Treg) displayed similar frequency of annexin V-labeled cells in SLE patients and normal controls (data not shown).

T lymphocytes from SLE patients show increased spontaneous apoptosis rate. Relative frequency of positive events for Annexin-V in total T lymphocytes (A), helper T lymphocytes (B) and Th1 cells (C) from SLE patients and controls. 24 h-culture in the presence of fetal bovine serum and no human serum supplement. Camptothencin (C +) differed significantly from all other conditions. Box-plot graph showing median, minimum, maximum, 25%, and 75% quartiles. p values were obtained using Mann–Whitney U test
SLE serum induced increase in the frequency of annexin V-labeled cells in the total population of T cells (Fig. 2A), in CD4+ helper T cells (Fig. 2B), and in Th2 cells (Fig. 2E) in comparison with cultures with NHS or with no human serum at all. However, no significant increase in annexin V-labeled events was noticed when we analyzed the effect of SLE serum separately on Th17, Th1, and Treg cells (Fig. 2C, 2D and 2F). Interestingly, there was a higher frequency of annexin V-labeled Th17 (Fig. 2C), Th1 (Fig. 2D), and Th2 lymphocytes (Fig. 2E) in PBMC cultures without human serum compared to cultures with NHS. The increased apoptosis rate in cultures without human serum was equally observed in Th1, Th2 and Th17 lymphocytes from SLE patients and healthy controls. Remarkably, in cultures exposed to SLE serum, Treg cells from SLE patients showed a higher frequency of annexin V-labeled events as compared to Treg cells from healthy controls (Fig. 3).

In vitro effects of serum from SLE patients on T lymphocyte subsets. Relative frequency of events positive for Annexin V in total T lymphocytes (A), helper T lymphocytes (B), Th17 (C), Th1 (D), Th2 (E) and Treg (F) from controls after 24 h of cell culture supplemented or not with human serum or apoptosis-inducing agent Camptothencin (C+). Total T lymphocytes, helper T lymphocytes, Th17 and Th1 from SLE patients behaved similarly to lymphocytes from controls (data not shown) NHS: culture medium with normal human serum; WHS: culture medium without the presence of human serum. C+ : culture medium with 5uM Camptothencin. SLE-serum: culture medium with lupus serum. Relative frequency of events positive for Annexin V in cultures containing Camptothencin for: CD3+ T lymphocytes (13.27% ± 8.66); helper CD3+ CD4+ T lymphocytes (10.54% ± 8.30); Th17 (26.61% ± 17.73), Th1 (26.75% ± 20.27), Th2 (13.34% ± 14.17), and Treg (34.42% ± 29.26). Camptothencin (C+) differed significantly from all other conditions. The results are presented as mean ± SD. *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001. All comparisons were analyzed using the Kruskal Wallis test followed by Dunn’s multiple comparison test

In vitro effects of serum from SLE patients on Treg from patients and controls. Relative frequency of events positive for Annexin V in Treg cells from patients compared to Treg from controls after 24 h of cell culture with lupus serum. The values are presented as median and interquartile range. p values were obtained using Mann–Whitney U test. SLE cells from SLE patients; CONTROL cells from normal subjects
Discussion
Apoptotic cells have been suggested to be a major source of autoantigen exposure in SLE [4, 5]. Previous studies have shown an increase in the frequency of apoptotic lymphocytes and monocytes derived from SLE patients in vitro [3,4,5]. It is possible that soluble factors contribute to the increased apoptotic rate observed in lymphocytes from SLE patients. In fact, it has been previously shown that serum from SLE patients induces in vitro apoptosis in monocytes and lymphocytes from normal healthy donors, as well as in T cell lines (MOLT-4 and Jurkat) and CD34+ cells [5,6,7,8,9,10]. This apoptosis-inducing effect was not observed with serum from patients with other autoimmune diseases, infectious diseases, and healthy individuals, and therefore it seems to be specific for SLE [5].
To the best of our knowledge, there is no study investigating the effect of lupus serum on separate CD4+ subsets, such as Th1, Th2, Treg, and Th17 lymphocytes. The results herein presented confirmed previous reports demonstrating an increase in the percentage of annexin V-labeled cells in total T lymphocytes and CD4+ helper T lymphocytes exposed in vitro to SLE serum compared to T cells exposed to normal human serum or not exposed to human serum. The underlying basis for this phenomenon is not known and may be related to the presence of specific apoptosis-inducing factors or otherwise related to the lack of surviving factors in SLE serum. A similar phenomenon was observed with Th2 cells; however, none of the other CD4+ T cell subsets studied showed an increase in the frequency of annexin V-labeled cells after exposure to SLE serum. Therefore, the effect of lupus serum observed in total CD4+ T lymphocytes appears to affect predominantly non-differentiated CD4+ T cells. The reasons for the divergent behavior of differentiated CD4+ T cells are not known, but one may speculate that the potential targets of putative factors in SLE serum eventually disappear or are not accessible. Alternatively, it is possible that the CD4+ T cell differentiation step renders cells less prone to apoptosis or less susceptible to soluble factors in SLE serum. These possibilities offer exciting opportunities for further investigation of this phenomenon.
Interestingly, Treg cells from SLE patients were more sensitive to SLE serum than Treg cells from normal individuals, whereas the other CD4+ subtypes (Th1, Th2, and Th17) from SLE patients and healthy controls showed similar behavior. To our surprise, Th1 and Th17 cells presented a greater frequency of annexin V-labeled cells in cultures without human serum as compared to those with normal human serum or with SLE serum. This observation suggests that human serum contains soluble factors that provide a protective effect against in vitro development of apoptosis of these lymphocyte subsets.
One potential limitation of this study is the use of annexin V as a surrogate marker to determine the percentage of cells undergoing apoptosis. In fact, already at early stages of apoptosis, the membrane phospholipid phosphatidylserine is translocated from the inner to the outer surface of the plasma membrane, thereby exposing phosphatidylserine to the binding of annexin V. Therefore, annexin V is a sensitive and widely used biomarker of apoptosis. However, annexin V can also label late-stage necrotic cells, as they lose outer membrane integrity, allowing for annexin V to bind to phosphatidylserine at the inner leaflet of cell membrane. Therefore, annexin V will bind apoptotic cells and late-stage necrotic cells. Consequently, the data herein presented represent predominantly apoptotic cells, which is the major cell dying mechanism for lymphocytes, but a minor proportion of annexin V-labeled cells might represent necrotic lymphocytes, mostly secondary to late stage apoptosis. Furthermore, since all the experiments were performed with pooled serum from SLE patients, we could not explore a potential heterogeneity among patients with respect to the serum apoptosis-inducing effect, nor could we investigate potential factors associated with this effect, such as current medication, acute phase reactants, autoantibodies, etc.
Conclusion
In conclusion, our findings confirmed previous studies, by demonstrating the apoptosis-inducing effect of lupus serum on CD4+ T lymphocytes in vitro. The same phenomenon was observed with Th2 cells, but not with the other CD4+ T subsets studied (Treg, Th1, and Th17) suggesting that undifferentiated T cells or some other subset of differentiated CD4+ T cells are preferentially susceptible to the apoptosis-inducing effect of SLE serum. These findings are of clinical interest as they provide preliminary understanding on the phenomenon of lymphocyte apoptosis in SLE, which might be related to the lymphopenia and increased lymphocyte apoptotic rate frequently observed in these patients. The herein reported findings reinforce the need for further studies to elucidate the mechanisms of cell death induction by SLE serum and the identification of the putative soluble factors present or absent in SLE serum that may be responsible for the apoptosis-inducing effect.
Availability of data and materials
The datasets were primarily generated for this study and cannot be shared openly to protect study participant privacy, but will be available from the corresponding author upon reasonable request.
References
Perniok A, Wedekind F, Herrmann M, Specker C, Schneider M. High levels of circulating early apoptic peripheral blood mononuclear cells in systemic lupus erythematosus. Lupus. 1998;7(2):113–8.
Yang F, He Y, Zhai Z, Sun E. Programmed cell death pathways in the pathogenesis of systemic lupus erythematosus. J Immunol Res. 2019;2019:3638562.
Emlen W, Niebur J, Kadera R. Accelerated in vitro apoptosis of lymphocytes from patients with systemic lupus erythematosus. J Immunol Baltim Md. 1994;152(7):3685–92.
Lorenz HM, Grünke M, Hieronymus T, Herrmann M, Kühnel A, Manger B, et al. In vitro apoptosis and expression of apoptosis-related molecules in lymphocytes from patients with systemic lupus erythematosus and other autoimmune diseases. Arthritis Rheum. 1997;40(2):306–17.
Bengtsson AA, Sturfelt G, Gullstrand B, Truedsson L. Induction of apoptosis in monocytes and lymphocytes by serum from patients with systemic lupus erythematosus—an additional mechanism to increased autoantigen load? Clin Exp Immunol. 2004;135(3):535–43.
Bengtsson AA, Gullstrand B, Truedsson L, Sturfelt G. SLE serum induces classical caspase-dependent apoptosis independent of death receptors. Clin Immunol. 2008;126(1):57–66.
Wang H, Xu J, Ji X, Yang X, Sun K, Liu X, et al. The abnormal apoptosis of T cell subsets and possible involvement of IL-10 in systemic lupus erythematosus. Cell Immunol. 2005;235(2):117–21.
Yang BC, Wang YS, Lin LC, Liu MF. Induction of apoptosis and cytokine gene expression in T-cell lines by sera of patients with systemic lupus erythematosus. Scand J Immunol. 1997;45(1):96–102.
Tiefenthaler M, Bacher N, Linert H, Mühlmann O, Hofer S, Sepp N, et al. Apoptosis of CD34+ cells after incubation with sera of leukopenic patients with systemic lupus erythematosus. Lupus. 2003;12(6):471–8.
Klint C, Truedsson L, Andreasson A, Johansson I, Sturfelt G. Toxic effects of SLE serum on normal monocytes in vitro: cell death induced by apoptosis related to complement dysfunction. Lupus. 2000;9(4):278–87.
Georgescu L, Vakkalanka RK, Elkon KB, Crow MK. Interleukin-10 promotes activation-induced cell death of SLE lymphocytes mediated by Fas ligand. J Clin Invest. 1997;100(10):2622–33.
Xue C, Lan-Lan W, Bei C, Jie C, Wei-Hua F. Abnormal Fas/FasL and caspase-3-mediated apoptotic signaling pathways of T lymphocyte subset in patients with systemic lupus erythematosus. Cell Immunol. 2006;239(2):121–8.
Zhang X, Brunner T, Carter L, Dutton RW, Rogers P, Bradley L, et al. Unequal death in T helper cell (Th)1 and Th2 effectors: Th1, but not Th2, effectors undergo rapid Fas/FasL-mediated apoptosis. J Exp Med. 1997;185(10):1837–49.
Yu Y, Iclozan C, Yamazaki T, Yang X, Anasetti C, Dong C, et al. Abundant c-Fas–associated death domain–like interleukin-1–converting enzyme inhibitory protein expression determines resistance of T helper 17 cells to activation-induced cell death. Blood. 2009;114(5):1026–8.
Funauchi M, Sugiyama M, SukYoo B, Ikoma S, Ohno M, Kinoshita K, et al. A possible role of apoptosis for regulating autoreactive responses in systemic lupus erythematosus. Lupus. 2001;10(4):284–8.
Pan X, Yuan X, Zheng Y, Wang W, Shan J, Lin F, et al. Increased CD45RA+ FoxP3(low) regulatory T cells with impaired suppressive function in patients with systemic lupus erythematosus. PLoS ONE. 2012;7(4): e34662.
Yang J, Chu Y, Yang X, Gao D, Zhu L, Yang X, et al. Th17 and natural Treg cell population dynamics in systemic lupus erythematosus. Arthritis Rheum. 2009;60(5):1472–83.
Shin MS, Lee N, Kang I. Effector T-cell subsets in systemic lupus erythematosus: update focusing on Th17 cells. Curr Opin Rheumatol. 2011;23(5):444–8.
Mesquita D, de Melo CW, Araujo J, Pucci F, Salmazi K, Kallas E, et al. Systemic lupus erythematosus exhibits a dynamic and continuum spectrum of effector/regulatory T cells. Scand J Rheumatol. 2011;40(1):41–50.
Petri M, Orbai AM, Alarcón GS, Gordon C, Merrill JT, Fortin PR, et al. Derivation and validation of the systemic lupus international collaborating clinics classification criteria for systemic lupus erythematosus. Arthritis Rheum. 2012;64(8):2677–86.
Acknowledgements
The authors thank São Paulo State Research Foundation (FAPESP-2009/12941-9) for the financial support. Luis Eduardo C. Andrade was supported by grant 476356/2008-3 from the Brazilian research agency National Council for Research (CNPq).
Funding
This work was supported by Sao Paulo Government agency FAPESP (Sao Paulo State Research Foundation) Grant Numbers 2009/12941-9, granted to L.E.C.A. Additionally, L.E.C.A. is supported by the Brazilian research agency National Council for Research (CNPq), Grant PQ-1D 310334/2019-5.
Author information
Authors and Affiliations
Contributions
DM and FVM designed the experiments. FVM and VF handled patient samples and performed the experiments. DM, FVM and VF analyzed the flow cytometry data. DM and FVM wrote the draft manuscript. LECA conceived the study and critically reviewed and edited the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The study was approved by the institution ethics committee (UNIFESP CEP 1332/09). All participants provided signed informed consent.
Consent for publication
All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Fig. S1.
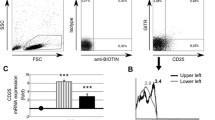
Flow cytometry strategy for determination of the relative frequency of T cell subsets. Specific sets of monoclonal antibodies were used to define each lymphocyte subset: Treg (CD3+CD4+CD25highCD127-/lowFoxp3+), Th1 (CD3+CD4+CCR4CXCR3+CCR5+), Th2 (CD3+CD4+CCR5CXCR3CCR4+) and Th17 (CD3+CD4+CCR6+CD161+). PBMC from a representative SLE were labeled after 24h of cell culture in the presence of fetal bovine serum.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mesquita, F.V., Ferreira, V., Mesquita, D. et al. CD4 T lymphocyte subsets display heterogeneous susceptibility to apoptosis induced by serum from patients with systemic lupus erythematosus. Adv Rheumatol 63, 40 (2023). https://doi.org/10.1186/s42358-023-00321-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s42358-023-00321-3