
Abstract
Despite the immunotherapeutics and target therapy agents, the survival of patients with advanced melanoma is still low. Notch signaling is able to regulate many aspects of melanomagenesis. Comparative analyses of common melanocytic nevi, dysplastic nevi and melanomas demonstrated increased expression of Notch1, Notch2 and their ligands, indicating that a positive regulation of these components may be related to the progression of melanoma. Some strategies such as gamma-secretase inhibitors (GSI) have been explored in patients with refractory metastatic disease or locally advanced disease of solid tumors. Two major classes of Notch inhibitors are currently in clinical development: GSI and monoclonal antibodies against Notch receptors or their ligands. Inhibition of Notch by GSI has been shown to decrease melanoma growth. GSI RO4929097 co-administered with cisplatin, vinblastine and temozolomide promotes greater elimination of tumor cells. The Notch pathway needs to be explored in the treatment of melanoma.
Similar content being viewed by others


Background
After gastrulation, the neural crest is first induced at the edge of the neural plate and in the non-neural ectoderm. Neural crest cells are initially multipotent but gradually become restricted to the potential line of development, which is determined by where they migrate and settle. Those that migrate to the epidermis and dermis, are subjected to the specification of lineages to form precursors of melanocytes - melanoblasts. Neural crest cells can originate several types of differentiated cells and tissues, including sensory neurons and glial cells, melanocytes, cartilage and craniofacial bone and smooth muscle. Melanocytic nevus is a benign accumulation of melanocytes that may be considered the first lesion in progression, evolving to melanocytic hyperplasia, dysplasia and melanoma. Dysplastic lesions may progress to melanoma in situ, invasive melanoma and then metastatic melanoma. This multi-stage tumor process results from the accumulation of genetic alterations that comprise genomic instability, activation of oncogenes such as BRAF and RAS, as well as inactivation of tumor suppressor genes (Bevona et al. 2003; Gandini et al. 2005; Hussein 2004; Naeyaert and Brochez 2003; Cichorek et al. 2013).
Analysis of the Cancer Genome Atlas and other genomic data made it possible to identify the most frequent mutations in melanoma and other cancers. BRAFV600E is the most prevalent mutation in melanoma, detected in 52% of cases, followed by mutations in the RAS and neurofibromin 1 (NF1) family in approximately 30 and 14% of cases, respectively. These conductive mutations are almost always mutually exclusive, making it possible to classify melanoma cases in distinct genomic subtypes: BRAF, RAS, NF1 and Triple-WT, the latter being defined by the absence of BRAF, RAS and NF1 mutations. Other common changes in melanoma include mutations in TP53, CDKN2A and TSGs homologous to phosphatase and tensin (PTEN) and in the TERT promoter (Akbani et al. 2015; Curtin et al. 2005).
Recent discoveries in cell signaling mechanisms have provided a better understanding of the biology underlying the progression of melanoma, and these advances have been explored to provide targeted drugs and novel therapeutic approaches. Among the genetic events underlying the development of melanoma, a variety of mutations in the components of cell signaling pathways was characterized, notably in the MAPK kinase pathway. Many of these alterations impair the functions of receptors on the plasma membrane and the aligned signaling cascades (Ghosh and Chin 2009; Gray-Schopfer et al. 2007; Dantonio et al. 2018).
Main text
Cutaneous melanoma originates from normal melanocytes or their precursors and among skin cancers is responsible for the highest number of deaths. In humans, melanocytes can be detected in the epidermis around the 50th day of intrauterine life. Embryonally, these cells migrate from the neural crest to the basal layer of the epidermis along the dorsolateral pathway. It has been suggested that the process of melanocyte development is often used by tumor cells to promote their own progression. The worldwide incidence of melanoma has been increasing in recent decades. The US National Cancer Institute reports that the rate of new cases of cutaneous melanomas has increased by an average of 1.5% per year over the past 10 years (Yoshida et al. 2001; Liu et al. 2014; Shain and Bastian 2016).
Familial melanoma accounts for 8–12% of melanoma cases, with the cyclin-dependent kinase inhibitor 2A (CDKN2A) and CDK4 being the major susceptibility genes involved in this context. Germline mutations in CDKN2A are responsible for the loss of two tumor suppressor proteins, p16INK4a and p14ARF, both encoded by the CDKN2A gene through alternative splicing while germline mutations in the CDK4 oncogene make a complex constitutively active between CDK4 and cyclin D1, which promotes abnormal cell proliferation (Thompson et al. 2009; Potrony et al. 2015; Aoude et al. 2015; Bertolotto 2013; Zuo et al. 1996).
Cutaneous melanoma affects young individuals (world average age 64 years) and presents aggressive behavior, with high lethality. Despite the breakthrough in advanced disease therapy, with the emergence of immunotherapeutics (anti-PD1 and anti-CTLA4) and target therapy agents (anti-BRAF and anti-MEK), survival of cutaneous melanoma patients remains directly related to the stage of the disease at diagnosis and depends mainly on the depth of the skin lesion as measured by the Breslow index. Thus, early diagnosis is of fundamental importance as well as the emergence of new therapeutic agents (National Comprehensive Cancer Network Clinical Practice Guidelines in Oncology 2018; Mozūraitienė et al. 2015).
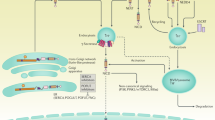
In mammals, four Notch receptors have been identified (Notch1–4) and are considered transmembrane type I proteins with the function of activation of transcription factors involved in embryonic development and whose signaling pathway is responsible for the control of epidermal differentiation and proliferation in adult life. Ligands on the surface of the neighboring cells (Delta1, Delta3, Delta4, Jagged1 and Jagged2) activate the Notch receptor inducing its intramembrane cleavage by gamma secretase complex which results in the release of the intracellular Notch domain in its active form with consequent activation of a series of genes involved in differentiation of progenitor cells during development (Massi et al. 2006; Panelos and Massi 2009).
The Notch signaling pathway contributes to the regulation of the melanocyte lineage. During development, melanocyte precursors, melanoblasts, emerge from the neural crest and migrate to the epidermis through newly formed hair follicles. In the hairless regions, melanoblasts remain immature and located in the basement membrane of the epidermis where they are differentiated for mature melanocytes by stimulating adjacent keratinocytes or other signals from the microenvironment. Recent studies indicate that the Notch signaling pathway is important for the maintenance of melanoblasts, for the appropriate localization of melanoblasts in the epidermal basement membrane, and for the induction of melanocyte differentiation in the follicular matrix. The transformation of normal melanocytes into melanoma cells is a multi-step process. MAPK is the major route involved in the known melanomagene. Activation of Notch signaling may contribute to melanoma progression, although it is not known which cells serve as emitting cells. Notch signaling is mediated by direct interactions between membrane-bound Notch receptors (Notch1–4) in recipient cells and Notch ligands (Delta or Jagged type) in the emitting cells (Massi et al. 2006; Panelos and Massi 2009; Golan et al. 2015; Guruharsha et al. 2012).
The Notch signaling cascade is initiated after binding of the membrane bound linker (Jagged 1,2 and DSL 1,3,4) to Notch receptors. After binding of the ligand, two sequential proteolytic events occur to release the active intracellular Notch (NIC). The first cleavage is mediated by a metalloprotease, TNF-a converting enzyme (TACE). Final cleavage is mediated by a γ-secretase complex. Subsequently, the NIC translocates to the nucleus and functions as a transcription factor to influence gene expression, including HES, Hey and other targets (Liu et al. 2014).
Some studies suggest that the Notch signaling pathway is pro-tumorigenic in melanomas. Notch1 expression levels are elevated in 50–60% of melanomas and 65% in melanoma cell lines, while in normal melanocytes and nevi are low or undetectable. In addition to promoting growth and survival of melanoma cells, Notch1 is also involved with a more aggressive phenotype. This phenotype has been associated with the activation of the MAPK and Akt pathways (Golan et al. 2015; Bedogni 2014; Kumano et al. 2007; Okuyama et al. 2008b).
It has now been demonstrated that in melanomas there is a greater expression of microRNAs (miR-146a) due to the activation of MAPK pathway (resulting from BRAF or mutant NRAS) and increased transcription factor MYC. Overexpression of miR-146a leads to the suppression of NUMB, which is a Notch-regulated protein. The silencing of NUMB by overexpression of miR-146a releases the Notch signaling pathway leading to the growth of tumor cells in melanoma (Golan et al. 2015; Garraway 2014).
In addition to the activation of the Notch signaling pathway involved in cell proliferation, it is also responsible for acquiring the vertical invasion capability of cutaneous melanomas (Golan et al. 2015).
The Notch signaling pathway in melanomas is activated directly by the contact between melanocytes and differentiated keratinocytes (keratinocytes from the most superficial layers of the epidermis) through their ligands (Delta-like or Jagged). After cell-cell (differentiated melanoma-keratinocyte) activation, the intracellular Notch domain (NICD) is cleaved by gamma-secretase complex and translocated to the nucleus where it binds to a transcriptional regulatory protein, the Jkappa protein (RBPJK) and its cofactor MITF. The RBPJK / MITF complex is responsible for the inhibition of the miR-222/221 micro RNA promoter, thus increasing the expression of miR-222/221, considered the trigger for vertical invasion in melanomas (Golan et al. 2015).
The Notch pathway is a highly conserved cell pathway, present in most multicellular organisms, which plays an important role in the determination, proliferation, differentiation and survival of the cell. The Notch signaling cascade modulates a wide range of cellular processes, including regulation of cell cycle blockade, apoptosis, survival, differentiation and maintenance of stem cells, as well as the response to hypoxia. Consequently, the aberrant function of Notch is involved in a number of human diseases, including developmental disorders, neurodegenerative diseases and cancer (e.g., T cell leukemia, multiple sclerosis, lymphoma and melanoma) (Ranganathan et al. 2011; Gustafsson et al. 2005; Osawa and Fisher 2008; Joutel and Tournier-Lasserve 1998; Müller 2012).
The Notch family of proteins is composed of cell surface receptors that transduce signals interacting with the Delta-like (DLL) and Jagged (JAG) transmembrane ligands in neighboring cells. In mammals, a total of four Notch receptors (Notch1–4) and five ligands were characterized (DLL1, 3, 4 and JAG1 and 2). Notch receptors are composed of two main domains: an extracellular domain (NECD) and an intracellular domain (NICD), interconnected by a transmembrane protein. The NICD has two structures at the C-terminus that are responsible for nuclear input (Kopan 2012; Takebe et al. 2014; Kojika and Griffin 2001; LaVoie and Selkoe 2003).
All Notch receptors are synthesized as full-length proteins that undergo successive proteolytic cleavages, culminating in the release of NICD, which will act on the nucleus. Notch ligands located in the neighboring cells have a Delta / Serrate / Lag-2 (DSL) domain that mediates binding-receptor binding and thus promotes the activation of Notch signaling. The mechanism for triggering Notch canonical signaling involves proteolytic cleavages at three Notch sites: S1, S2 and S3. The first cleavage (S1) is mediated by furin type convertases, which occurs in the trans-Golgi network during the Notch secretion process. This cleavage generates two subunits (NECD and NICD) interconnected by the transmembrane protein. These processed subunits are then transported to the plasma membrane, where they associate as non-covalently linked heterodimers to form the functional Notch receptor. Upon binding of the linker to the receptor, the latter undergoes the second cleavage (S2) which is catalyzed by a member of the disintegrin A family and metalloproteases (ADAM17 or ADAM10), also known as tumor necrosis factor (TACE) converting enzyme. Cleavage of S2 promotes the dissociation of the membrane-bound intracellular Notch domain from the extracellular domain. This intracellular domain is a constitutive substrate for the final cleavage of S3, culminating in the release of active NICD, which is directed to the nucleus by its NLS. S3 is regulated by a protease complex of the presenilin-dependent γ-secretase, which consists of an integral membrane protein complex. After translocation to the nucleus, the NICD binds to CSL (transcriptional repressor RBPJk / CSL), a transcriptional repressor that acts as a DNA binding adapter and helps recruit the adapter protein (MAML), which in turn evokes the transcriptional co-activator p300 and other components of the transcription machinery (Liu et al. 2014; Kopan 2012; Takebe et al. 2014; Kojika and Griffin 2001; LaVoie and Selkoe 2003; Baron 2003; Pinnix and Herlyn 2007; Okuyama et al. 2008a; Nam et al. 2003).
The transcriptional activation complex, composed of CSL, NICD, MAML and p300, regulates transcription of genes downstream, such as HES1 and HEY. The Hess family of repressors acts downstream of the Notch signaling pathway, antagonizing the expression of a variety of transcription factors, such as Ascl1, Atoh1 and Neurog3, whose function is to keep the cells in an undifferentiated state. Hes repressor genes, such as HES1, play key roles in maintaining progenitor cells in an undifferentiated state. The concomitant existence of transcriptional activators and repressors downstream of the Notch signaling cascade plays a crucial role in the various outcomes of this pathway (Osawa and Fisher 2008; Pinnix and Herlyn 2007; Tao et al. 2010; Kageyama et al. 2007).
Several studies have also demonstrated the existence of noncanonical activation of Notch signaling in several cell types. At least three types of non-canonical activation of Notch have been described, which may or may not depend on: (i) ligand interaction, (ii) γ-secretase activity and (iii) the action of RBPJk / CSL complexes. Both non-canonical cascades independent of RBPJ and Hes have important functions downstream of Notch signaling, although the exact molecular events mediating these sub-pathways are not fully understood (Osawa and Fisher 2008; Veeraraghavalu et al. 2005; Hodkinson et al. 2007; Ayaz and Osborne 2014).
Notch signaling may also interact with other pathways, such as PI3K, mTORC2, Wnt, NF-κB, YY1 or HIF-1α, at cytoplasmic and / or nuclear levels. Direct interactions of NICD with IKKα in the NF-κB pathway or LEF1 in the Wnt pathway were reported, and that Notch can activate the integrin via Ras, irrespective of RBPJ binding. Although many normal cellular processes (homeostatic regulation of melanocytes) require canonical signaling of Notch, many pathological conditions, including cancer and immune system activation, are associated with non-canonical Notch signals. However, these noncanonical cascades still require a more detailed understanding (Tao et al. 2010; Ayaz and Osborne 2014; Hirobe 2005).
In healthy skin, Notch signaling is expressed in all layers of the epidermis and is critical for the differentiation of keratinocytes and affects a wide range of cellular activities including cell cycle arrest, apoptosis, and survival. An example of the versatile function of the Notch pathway is related to epidermal development, where Notch signaling is able to stimulate the differentiation of granular cells into spinous cells and, at the same time, prevent their premature differentiation (Okuyama et al. 2008a; Hirobe 2005).
This versatility of functions can be explained by the concomitant existence of a transcriptional activator and a repressor downstream of Notch. Notch signaling, via Hes1 transcription factor, maintains the survival of melanoblast and melanocytic stem cells, preventing the onset of apoptosis (Osawa and Fisher 2008; Moriyama et al. 2006).
Notch signaling is able to regulate many aspects of melanomagenesis. Comparative analyses of common melanocytic nevi, dysplastic nevi and melanomas demonstrated an increased expression of Notch1, Notch2 and its ligands, indicating that a positive regulation of these components may be related to the progression of melanoma (Massi et al. 2006).
An essential role for the Notch pathway has been validated in the development of melanoblast as well as the progression of melanoma. Notch1 is considered a primary tumorigenic factor in melanoma. Positive regulation of Notch1 and its target genes occurs in metastatic melanoma. Transcriptional targets of Notch signaling that are responsible for angiogenesis, proliferation, metastasis and cell survival in tumor cells include Hes, Hey, Cyclin D1, NF-κB, STAT3 and p21 (Schouwey et al. 2007; Ma et al. 2014; Balint et al. 2005).
The Notch pathway may offer a therapeutic possibility for the treatment of melanoma. Considering the effects of Notch overexpression on the development of melanoma, the manipulation of this pathway seems to have great potential therapeutic value. Several strategies to inhibit the Notch pathway have been used against melanoma and other cancers. Recent evidence has also suggested that Notch signaling is one of the most important cell pathways in drug-resistant tumor cells. In fact, negative regulation of the Notch pathway may induce drug sensitivity, leading to increased inhibition of the growth, invasion, and metastasis of cancer cells. For example, the use of RNA directed to Notch2 has shown potential to reduce tumor invasion and the growth of uveal melanoma, a common intraocular malignancy in adults (Liu et al. 2014; Purow 2009; Wang et al. 1806; Asnaghi et al. 2012).
Increasing evidence indicates that Notch signaling contributes to physiological processes, including development, differentiation and tumorigenesis, either as a promoter or tumor suppressor, depending on the cellular context, level of expression and interaction with other signaling systems. There is also evidence that signaling through Notch receptors regulates cell proliferation and cell survival in various types of cancer, including melanoma. Tumor progression / melanoma metastasis are complicated processes that require multiple cellular events, including cell proliferation, survival, migration and invasion. Notch signaling appears to be a promising system for novel therapeutic targets for the treatment of melanoma and perhaps for the prevention of melanoma metastases (Müller 2012).
Increased knowledge of the molecular basis of melanoma has transformed the treatment of these patients. Advances in immunobiology brought a class of immunotherapeutic agents that provided more durable control of patients with metastatic melanoma. For tumors that do not respond to immunotherapeutics but have specific mutations as in BRAFV600E (occurring in approximately 50% of cutaneous melanoma cases), target therapies with MAPK pharmacological inhibitors (RAF and MEK effectors) have shown benefit, even though not all of melanomas with the mutations respond to the MAPK pharmacological inhibitors (Tolcher et al. 2012).
Despite the benefits, approximately 40% of melanomas do not respond to inhibitors of the MAPK pathway nor to target therapy agents. Tumors develop resistance and patients have recurrences (Wolchok et al. 2017). Some strategies such as gamma-secretase inhibitors have been explored in phase I and phase II studies in patients with metastatic refractory disease or locally advanced disease of solid tumors. It has been investigated whether the addition of gamma-secretase inhibitors to RAF and MEK inhibitors could bring benefit in the treatment of patients with advanced melanoma (Krepler et al. 2016; Tolcher et al. 2012).
Two major classes of Notch inhibitors are currently in clinical development: γ-secretase inhibitors (GSIs) and monoclonal antibodies (mAbs) against Notch receptors or their ligands. Inhibition of Notch by the y-secretase inhibitor has been shown to decrease melanoma growth. A phase I clinical trial for GSI RO4929097 has shown encouraging evidence of antitumor activity, revealing itself as a selective inhibitor of small molecules of y-secretase with antitumor activity. Suppression of Notch-2 activation is critical for honokiol to inhibit melanoma cells (Kaushik et al. 2015).
Antitumor activity by GSIs and mAbs administered as a single agent in the early stages of clinical trials has been observed in advanced or metastatic thyroid cancer, non-small cell lung cancer, intracranial tumors, sarcoma or desmoid tumors, colorectal cancer with neuroendocrine features, melanoma and ovarian cancer. Several adverse events, particularly gastrointestinal toxicities, have been observed and mitigation strategies are being developed following the testing of multiple GSIs and Notch-directed mAbs (Takebe et al. 2014).
Combined chemotherapy with GSIs for recurrent and advanced stage melanoma were evaluated in Phase I and II clinical trials. GSI RO4929097 co-administered with cisplatin, vinblastine and temozolomide promotes a greater elimination of tumor cells (Nickoloff et al. 2005; Briot and Iruela-Arispe 2015; Espinoza and Miele 2013). There is evidence that the Notch pathway needs to be explored because of its potential to become useful in the treatment of melanoma.
Conclusions
Considering the effects of Notch overexpression on the development of melanoma, the manipulation of this pathway seems to have a great potential therapeutic value. Several strategies to inhibit the Notch pathway have been used against melanoma and other cancers. Despite the benefits, approximately 40% of melanomas do not respond to immunotherapy (anti-CTLA4, anti-PD1 and anti PDL1) nor to target therapy agents (anti-BRAF and anti-MEK). It has been investigated whether the addition of gamma-secretase inhibitors to BRAF and MEK inhibitors could bring benefit in the treatment of patients with advanced melanoma.
Two major classes of Notch inhibitors are currently in clinical development: γ-secretase inhibitors (GSIs) and monoclonal antibodies (mAbs) against Notch receptors or their ligands. GSI co-administered with cisplatin, vinblastine and temozolomide promotes a greater elimination of tumor cells (Nickoloff et al. 2005; Briot and Iruela-Arispe 2015; Espinoza and Miele 2013). Notch pathway needs to be explored because of its potential to become useful in the treatment of melanoma.
Abbreviations
- CDKN2A:
-
Cyclin-dependent kinase inhibitor 2A
- DLL:
-
Delta-like
- DSL:
-
Delta / Serrate / Lag-2
- GSI:
-
Gamma-secretase inihibitors
- JAG:
-
Jagged
- mAbs:
-
Monoclonal antibodies
- NECD:
-
Nocth extracellular domain
- NF1:
-
Neurofibromin 1
- NIC:
-
Active intracellular Notch
- NICD:
-
Intracellular Notch domain
- PTEN:
-
Phosphatase and tensin
- TACE:
-
TNF-a converting enzyme
References
Akbani R, Akdemir KC, Aksoy BA, Albert M, Ally A, Amin SB et al (2015) Genomic classification of cutaneous melanoma. Cell 161:1681–1696
Aoude LG, Wadt KAW, Pritchard AL, Hayward NK (2015) Genetics of familial melanoma: 20 years after CDKN2A. Pigment Cell Melanoma Res 28:148–160
Asnaghi L, Ebrahimi KB, Schreck KC, Bar EE, Coonfield ML, Bell WR et al (2012) Notch signaling promotes growth and invasion in uveal melanoma. Clin Cancer Res 18:328–341
Ayaz F, Osborne BA (2014) Non-canonical Notch signaling in cancer and immunity. Front Oncol 4:345
Balint K, Xiao M, Pinnix CC, Soma A, Veres I, Juhasz I et al (2005) Activation of Notch1 signaling is required for beta-catenin-mediated human primary melanoma progression. J Clin Invest 115:3166–3176
Baron M (2003) An overview of the notch signalling pathway. Semin Cell Dev Biol 14:113–119
Bedogni B (2014) Notch signaling in melanoma: interacting pathways and stromal influences that enhance notch targeting. Pigment Cell Melanoma Res 27:162–168
Bertolotto C (2013) Melanoma: from melanocyte to genetic alterations and clinical options. Scientifica (Cairo) 2013:1–22
Bevona C, Goggins W, Quinn T, Fullerton J, Tsao H, Corona R (2003) Cutaneous melanomas associated with nevi. Arch Dermatol 139:1620–1624
Briot A, Iruela-Arispe M (2015) Blockade of specific NOTCH ligands: a new promising approach in cancer therapy. Cancer Discov 5:112–114
Cichorek M, Wachulska M, Stasiewicz A, Tymińska A (2013) Skin melanocytes: biology and development. Postepy Dermatol Alergol 30(1):30–41
Curtin JA, Fridlyand J, Kageshita T, Patel HN, Busam KJ, Kutzner H et al (2005) Distinct sets of genetic alterations in melanoma. N Engl J Med 353:2135–2147
Dantonio PM, Klein MO, Freire MRVB, Araujo CN, Chiacetti AN, Correa RG (2018) Exploring major signaling cascades in melanomagenesis: a rationale route for targetted skin cancer therapy. Biosci Rep 38(5):BSR201180511
Espinoza I, Miele L (2013) Notch inhibitors for cancer treatment. Pharmacol Ther 139:95–110
Gandini S, Sera F, Cattaruzza MS, Pasquini P, Abeni D, Boyle P et al (2005) Meta-analysis of risk factors for cutaneous melanoma: I. common and atypical naevi. Eur J Cancer 41:28–44
Garraway LA (2014) A Notch for noncoding RNA in melanoma. N Engl J Med 370(20):1950–1951
Ghosh P, Chin L (2009) Genetics and genomics of melanoma. Expert Rev Dermatol 4:131–143
Golan T, Messer AR, Amitai-Lange A, Melamed Z, Ohana R, Bell RE et al (2015) Interactions of melanoma cells with distal keratinocytes trigger metastasis via Notch signaling inhibition of MITF. Mol Cell 59(4):664–676
Gray-Schopfer V, Wellbrock C, Marais R (2007) Melanoma biology and new targeted therapy. Nature 445:851–857
Guruharsha KG, Kankel MW, Artavanis-Tsakonas S (2012) The Notch signalling system: recent insights into the complexity of a conserved pathway. Nat Rev Genet 13(9):654–666
Gustafsson MV, Zheng X, Pereira T, Gradin K, Jin S, Lundkvist J et al (2005) Hypoxia requires notch signaling to maintain the undifferentiated cell state. Dev Cell 9:617–628
Hirobe T (2005) Role of keratinocyte-derived factors involved in regulating the proliferation and differentiation of mammalian epidermal melanocytes. Pigment Cell Res 18:2–12
Hodkinson PS, Elliott PA, Lad Y, McHugh BJ, MacKinnon AC, Haslett C et al (2007) Mammalian NOTCH-1 activates β1 integrins via the small GTPase R-Ras. J Biol Chem 282:28991–29001
Hussein MR (2004) Genetic pathways to melanoma tumorigenesis. J Clin Pathol 57:797–801
Joutel A, Tournier-Lasserve E (1998) Notch signalling pathway and human diseases. Semin Cell Dev Biol 9:619–625
Kageyama R, Ohtsuka T, Kobayashi T (2007) The Hes gene family: repressors and oscillators that orchestrate embryogenesis. Development 134:1243–1251
Kaushik G, Venugopal A, Ramamoorthy P, Standing D, Subramaniam D, Umar S et al (2015) Honokiol inhibits melanoma stem cells by targeting notch signaling. Mol Carcinog 54(12):1710–1721
Kojika S, Griffin JD (2001) Notch receptors and hematopoiesis. Exp Hematol 29:1041–1052
Kopan R (2012) Notch signaling. Cold Spring Harb Perspect Biol 4:a011213
Krepler C, Xiao M, Samanta M, Vultur A, Chen HY, Brafford P et al (2016) Targeting Notch enhances the efficacy of ERK inhibitors in BRAF-V600E melanoma. Oncotarget 7:71211–71222
Kumano K, Masuda S, Sata M, Saito T, Lee SY, Sakata-Yanagimoto M et al (2007) Both Notch1 and Notch2 contribute to the regulation of melanocyte homeostasis. Pigment Cell Melanoma Res 21(1):70–78
LaVoie MJ, Selkoe DJ (2003) The Notch ligands, Jagged and Delta, are sequentially processed by α-Secretase and Presenilin/γ-Secretase and release signaling fragments. J Biol Chem 278:34427–34437
Liu J, Fukunaga-Kalabis M, Li L, Herlyn M (2014) Developmental pathways activated in melanocytes and melanoma. Arch Biochem Biophys 563:13–21
Ma J, Tang X, Wong P, Jacobs B, Borden EC, Bedogni B (2014) Noncanonical activation of Notch1 protein by membrane type 1 matrix metalloproteinase (MT1-MMP) controls melanoma cell proliferation. J Biol Chem 289:8442–8449
Massi D, Tarantini F, Franchi A, Paglierani M, Di Serio C, Pellerito S et al (2006) Evidence for differential expression of Notch receptors and their ligands in melanocytic nevi and cutaneous malignant melanoma. Mod Pathol 19:246–254
Moriyama M, Osawa M, Mak SS, Ohtsuka T, Yamamoto N, Han H et al (2006) Notch signaling via Hes1 transcription factor maintains survival of melanoblasts and melanocyte stem cells. J Cell Biol 173:333–339
Mozūraitienė J, Bielskienė K, Atkočius V, Labeikytė D (2015) Molecular alterations in signal pathways of melanoma and new personalized treatment strategies: targeting of Notch. Medicina (Kaunas) 51(3):133–145
Müller CSL (2012) Notch signaling and malignant melanoma. Adv Exp Med Biol 727:258–264
Naeyaert JM, Brochez L (2003) Clinical practice. Dysplastic nevi. N Engl J Med 349:2233–2240
Nam Y, Weng AP, Aster JC, Blacklow SC (2003) Structural requirements for assembly of the CSL·intracellular Notch1 Mastermind-like 1 transcriptional activation complex. J Biol Chem 278:21232–21239
National Comprehensive Cancer Network Clinical Practice Guidelines in Oncology (2018). Version 3.2018. available at www.nccn.org.
Nickoloff BJ, Hendrix MJC, Pollock PM, Trent JM, Miele L, Qin JZ (2005) Notch and NOXA-related pathways in melanoma cells. J Invest Dermatol 10:95–104
Okuyama R, Tagami H, Aiba S (2008) Aberrant Notch signaling results in the development of psoriasis and skin cancers such as squamous cell carcinoma, basal cell carcinoma and malignant melanoma. J Dermatol Sci 49(3):187–194
Okuyama R, Tagami H, Aiba S (2008a) Notch signaling: its role in epidermal homeostasis and in the pathogenesis of skin diseases. J Dermatol Sci 49:187–194
Osawa M, Fisher D (2008) Notch and melanocytes: diverse outcomes from a single signal. J Invest Dermatol 128:2571–2574
Panelos J, Massi D (2009) Emerging role of Notch signaling in epidermal differentiation and skin cancer. Cancer Biol Ther 8(21):1986–1993
Pinnix CC, Herlyn M (2007) The many faces of Notch signaling in skin-derived cells. Pigment Cell Res 20:458–465
Potrony M, Badenas C, Aguilera P, Puig-Butille JA, Carrera C, Malvehy J et al (2015) Update in genetic susceptibility in melanoma. Ann Transl Med 3:210
Purow B (2009) Notch inhibitors as a new tool in the war on cancer: a pathway to watch. Curr Pharm Biotechnol 10:154–160
Ranganathan P, Weaver KL, Capobianco AJ (2011) Notch signalling in solid tumours: a little bit of everything but not all the time. Nat Rev Cancer 11:338–351
Schouwey K, Delmas V, Larue L, Zimber-Strobl U, Strobl LJ, Radtke F et al (2007) Notch1 and Notch2 receptors influence progressive hair graying in a dose-dependent manner. Dev Dyn 236:282–289
Shain AH, Bastian B (2016) From melanocytes to melanomas. Nat Rev Cancer 16(6):345–358
Takebe N, Nguyen D, Yang SX (2014) Targeting Notch signaling pathway in cancer: clinical development advances and challenges. Pharmacol Ther 141:140–149
Tao J, Chen S, Lee B (2010) Alteration of Notch signaling in skeletal development and disease. Ann N Y Acad Sci 1192:257–268
Thompson JF, Scolyer RA, Kefford RF (2009) Cutaneous melanoma in the era of molecular profiling. Lancet 374:362–365
Tolcher AW, Messersmith WA, Mikulski SM, Papadopoulos KP, Kwak EL, Gibbon DG (2012) Phase I study of RO4929097, a gamma secretase inhibitor of Notch signaling, in patients with refractory metastatic or locally advanced solid tumors. J Clin Oncol 30(19):2348–2353
Veeraraghavalu K, Subbaiah VK, Srivastava S, Chakrabarti O, Syal R, Krishna S (2005) Complementation of human papillomavirus type 16 E6 and E7 by Jagged1-specific Notch1-phosphatidylinositol 3-kinase signaling involves pleiotropic oncogenic functions independent of CBF1;Su(H);Lag-1 activation. J Virol 79:7889–7898
Wang Z, Li Y, Ahmad A, Azmi AS, Banerjee S, Kong D et al (1806) Targeting Notch signaling pathway to overcome drug resistance for cancer therapy. Biochim Biophys Acta 2010:258–267
Wolchok JD, Chiarion-Sileni V, Gonzalez R, Rutkowski P, Grob JJ, Cowey CL et al (2017) Overall survival with combined nivolumab and ipilimumab in advanced melanoma. N Engl J Med 377(14):1345–1356
Yoshida H, Kunisada T, Grimm T, Nishimura EK, Nishioka E, Nishikawa SI (2001) Review: melanocyte migration and survival controlled by SCF/c-kit expression. J Investig Dermatol Symp Proc 6:1–5
Zuo L, Weger J, Yang Q, Goldstein AM, Tucker MA, Walker GJ et al (1996) Germline mutations in the p16(INK4a) binding domain of CDK4 in familial melanoma. Nat Genet 12:97–99
Acknowledgements
None
Author information
Authors and Affiliations
Contributions
RSOF conceived of the study and participated in its design and coordination. ALS provided the search of the main subject in databases Medline and LILACS. FMP drafted the manuscript. EO and RM contributed with pathology aspects of the study. ACB and LMF participated in the sequence alignment and final revision. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
de Oliveira Filho, R.S., Soares, A.L., Paschoal, F.M. et al. Literature review of Notch melanoma receptors. Surg Exp Pathol 2, 27 (2019). https://doi.org/10.1186/s42047-019-0052-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s42047-019-0052-9