
Abstract
Background
Domestication usually has negative effects on insect performance, especially when they are reared continuously for many generations. Rejuvenation can reduce the negative effects of domestication in the parasitoid, Trichogramma brassicae Bezdenko (Hymenoptera: Trichogrammatidae) reared for 30 generations (G) on Sitotroga cerealella (Olivier) (Lepidoptera: Gelechiidae). Life table and parasitism parameters in the primary colony (G31–G45) were compared with the respective colony rejuvenated with field-collected individuals.
Results
There were significant differences in life history traits between domesticated and rejuvenated populations. The highest (38.4 eggs/female) and lowest (13.3 eggs/female) fecundity was observed in rejuvenated G31 and domesticated G45, respectively. The highest values of the life table and parasitism parameters were observed in rejuvenated G31 without significant difference with G32, G33 and G34. The relationship between finite parasitism rate (ω) and generation number in both rejuvenated and domesticated populations were fitted to the cubic regression model, indicating the parasitism potential of T. brassicae increased significantly after adding feral individuals but decreased remarkably over the generations.
Conclusions
Therefore, it seems that adding 10% feral individuals re-established the reproductive performance of the wasps at least for four generations, and it needs to be repeated routinely or it needs to be made by adding a higher rate of feral individuals. However, this issue should be investigated by more studies in which different rates of feral individuals are added to the primary colony.
Similar content being viewed by others


Background
There is an increasing interest globally for using natural enemies against pests so that many of these organisms are cultured and traded by companies in many countries. To achieve and maintain successful biological control, these organisms need to be checked qualitatively both during production and after being released, because they may lose some attributes after continuous mass rearing that are necessary for biocontrol agents being released in an augmentative program (Yazdanpanah et al. 2022). Natural enemies applied for the control of pests under field conditions are being reared in insectaries for many generations, and their potential may decrease and cause the biological control program to fail. Long-term mass rearing under such conditions may promote inbreeding and lead to inefficiency under field conditions (Hopper et al. 1993). Domestication usually causes insects to experience homogeneous environment that lacks the challenges of natural conditions. In some cases, the targeted traits are so essential for success of natural enemies under field conditions (Bartlett 1984). Whether quality changes occur in continuous rearing should be determined prior to inundative release programs and to support recommendations for routinely rejuvenating commercial cultures using wild insects (Smith 1996), prevent inbreeding and conserving the gene pool (van Lenteren 2003).
The egg-parasitoid, Trichogramma brassicae Bezdenko (Hymenoptera: Trichogrammatidae) is one of the widely used biocontrol agents in Iran (Moghaddassi et al. 2019) and in other countries (Cônsoli et al. 2010), commonly mass reared on various factitious hosts, such as the Angoumois grain moth, Sitotroga cerealella (Olivier) (Lepidoptera: Gelechiidae) (Wang et al. 2014). In a previous studies, a large decrease in the quality of T. brassicae over long-term mass rearing was found (Ghaemmaghami et al. 2021). To find whether rejuvenation can reduce the negative effects of domestication in T. brassicae wasps continuously mass reared for many generations was endeavored. The present study aimed to compare life table and parasitism parameters of the domesticated and rejuvenated populations of T. brassicae under long-term mass rearing. This could deepen our knowledge for improving biological control programs using high-quality parasitoid.
Methods
Insects rearing
A colony of Angoumois grain moth, S. cerealella, was established on barley grains under ambient temperature (25 ± 1 °C) in a biocontrol laboratory of the Technical and Vocational Training Center of Estahban, Fars province, Iran. Emerged adults were transferred to funnels (diameter 20 cm) covered with a fine mesh net over the mouth. After mating, the eggs were collected on papers placed under the open side of the funnel. The domesticated population data were extracted from Ghaemmaghami et al. (2021). To obtain a stock to initially create the primary colony of T. brassicae, egg traps, pieces of white papers (21 × 11 cm) containing S. cerealella eggs, were placed on tomato farms in Karaj (51°00′ E, 35°48′ N), Iran, in May 2017. The traps were recollected after 48 h, and the parasitized eggs were incubated in a growth chamber set at 25 ± 1 °C, 65 ± 5% RH and a 16:8 h (L:D) photoperiod until the emergence of T. brassicae adults (G0), which were then reared in rectangular plastic containers (35 × 20 × 15 cm) on S. cerealella eggs under the same conditions mentioned above. The parasitoids were reared sequentially for 45 generations, and for tested generations, a cohort of parasitized eggs were separated.
To rejuvenate the colony, after 30 generations of domestication, field-collected parasitoids were added to the colony. Feral individuals of T. brassicae were collected using egg traps as mentioned above. The parasitized eggs were separately added to a part of primary population in 1:10 ratio (feral: primary). The first generation in the rejuvenated population corresponded to the 31st generation in the domesticated one. The rearing conditions were similar to those for primary colony.
Assessment procedure
Around 80 one-day-old parasitized eggs were harvested randomly from the primary (≈ 1000 wasps) and rejuvenated colony (≈ 1000 wasps) and kept in a glass container (6 cm in diameter, 5 cm in height) until the emergence of the adult wasps. Male and female wasps were then paired and placed in glass cylinders (10 × 1.6 cm) containing about 150 one-day-old host eggs (stuck to paper tape; 1 × 5 cm). The wasps were supplied with diluted honey on cotton rolls as a carbohydrate source during oviposition. If a male died, it was replaced by a newly emerged (< 24 h old) male from the stock colony. These individuals were excluded from analyses. The wasps were checked daily until the death of the last individual and the number of parasitized eggs, development time, adult longevity, fecundity and their mortality and survival rate were estimated. These procedures were repeated for generations 31, 32, 33, 34, 35, 36, 40 and 45 of the primary colony and simultaneously for the respective generations in the rejuvenated population. All experiments were carried out in an incubator set at 26 ± 1 °C, 65 ± 5% RH and a photoperiod of 16:8 h (L:D).
Data analysis
The life history data of all individuals were analyzed using age-stage, two-sex life table theory (Chi 1988). Data analysis and population parameters including gross reproductive rate (GRR), net reproductive rate (R0), intrinsic rate of increase (r), finite rate of increase (λ) and mean generation time (T) were calculated by TWOSEX-MSChart program (Chi 2019). A bootstrap procedure was used to estimate the variances and the standard errors of population parameters (Huang and Chi 2012). To obtain stable estimates, 100,000 bootstrap samples were used. Bootstrap values of different generations of T. brassicae were then compared via paired-bootstrap procedure (Bahari et al. 2018).
Data obtained on daily parasitism rates of the total cohort were used to estimate the parasitism parameters for each of the different generations. The estimated parameters included net parasitism rate (c0), transition rate (Qp), stable parasitism rate (ψ) and finite parasitism rate (ω). c0 describes the mean number of hosts parasitized by an individual parasitoid during its entire life span. Qp is the transformation rate from host population to parasitoid offspring. ψ is the total parasitism capacity of a stable population with the same total size. The finite parasitism rate (ω) estimates the parasitism potential of a parasitoid population by combining its growth rate (λ), age-stage parasitism rate (cxj) and stable age-stage structure (axj) (Chi and Yang 2003).
Parasitism rate data were analyzed using the computer program CONSUME-MSChart (Chi 2019). The bootstrap resampling method (100,000 bootstraps) was used to estimate the variances and the standard errors of the parameters. Comparison of parasitism parameters was done based on paired-bootstrap test using TWOSEX-MSChart program (Chi 2019).
One-way ANOVA was used to analyze fecundity and durations of different life stages of the domesticated and rejuvenated populations of T. brassicae, and in the case of significant differences, mean comparisons were made using Tukey’s test (SPSS-Inc, 2009). Mean comparison among corresponding generations was analyzed using t tests using SPSS program (SPSS-Inc 2009). Domesticated population data were extracted from Ghaemmaghami et al. (2021).
Results
Life duration, survival rate and fecundity
Except for domesticated G45, none of generations in domesticated and rejuvenated populations had significant differences in the female longevity and total life span with rejuvenated G31. Domesticated G45 had shorter female longevity (3.75 d) than rejuvenated G31 (5.24 d), and a shorter total life span (14.12 d) than rejuvenated G31 (15.49 d) and G32 (15.50 d). Rejuvenated G31 had longer male adult longevity (5.48 d) than domesticated G40 (4.05 d) and G45 (4.06 d) (Table 1).
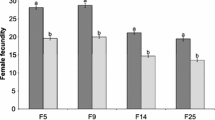
There were significant differences between domesticated and rejuvenated populations in terms of oviposition days of the wasps (Table 1). The length of oviposition days ranged from 3.84 d in rejuvenated G31 to 2.55 d in domesticated G45. Mean comparison of oviposition days between corresponded generations showed significant differences in G31 (t = 3.427; df = 1.72; P = 0.004), G32 (t = 3.266; df = 1.74; P = 0.001), G33 (t = 2.777; df = 1.5; P = 0.001) and G34 (t = 2.668; df = 1.71; P = 0.004). Fecundity was different in the rejuvenated and domesticated populations. The highest (38 eggs/female) and lowest (13 eggs/female) fecundities were observed, respectively, in rejuvenated G31 without significant difference with G32, G33 and G34 and domesticated G45 without significant difference with G31, G32, G33, G34, G35, G36 and G40 (Table 1). However, the sex ratio was not affected by rejuvenation, and non-significant difference was found between rejuvenated and domesticated populations (Fig. 1). Although the sex ratio was not statistically significant, domesticated populations tend to have higher female ratio every generation.

Sex ratio (% emerged females) of different generations of Trichogramma brassicae reared on Sitotroga cerealella eggs in both domesticated and rejuvenated populations
The age-stage-specific survival rates (sxj) of T. brassicae (Fig. 2) showed the survivorship and stage differentiation as well as the variable developmental rate. The mean number of offspring produced by T. brassicae individuals of the age x and stage j per day was shown with the age-stage-specific fecundity (fxj) in Fig. 3. The start of oviposition of the first female in all of generations occurred at the age 11 days. The daily fecundity of T. brassicae ranged from 13.27 eggs in G45 to 19.61 eggs in G31. The highest and lowest age-specific survival rates (lx) at age of adult emergence of T. brassicae after adding feral population were observed at age 11 days in G34 (0.90) and G45 (0.81), respectively (Fig. 3).

Age-stage survival rate (sxj) in different generations (G) of the rejuvenated Trichogramma brassicae on Sitotroga cerealella eggs

Age-specific survivorship (lx), age-stage-specific fecundity of female (fxj) (eggs) and age-specific fecundity (mx) in different generations (G) of the rejuvenated Trichogramma brassicae on Sitotroga cerealella eggs
Population growth (life table) parameters
Based on the age-stage, two-sex life table procedure, the population (life table) parameters were estimated using the data of the entire cohort for each generation of the two populations (Table 2). Significant differences in population growth parameters were found among different generations of T. brassicae in primary and rejuvenated populations. The highest values of GRR (26.13 eggs/individual), R0 (20.71 eggs/individual), r (0.239 day−1), λ (1.271 day−1) and T (12.65 days) were observed in rejuvenated G31, and the lowest ones were in domesticated G45 with 9.91eggs/individual, 7.82 eggs/individual, 0.167 day−1, 1.181 day−1 and 12.29 days, respectively (Table 2).
Parasitism capacity
Comparing parasitism parameters between populations before and after adding feral individuals showed that some generations in the rejuvenated population had more parasitism capacity than others both in the rejuvenated and domesticated populations (Table 3). The highest values for net parasitism rate (c0), stable parasitism rate (ψ) and finite parasitism rate (ω) were observed in rejuvenated G31 with 20.71 hosts/parasitoid, 0.279 hosts/parasitoid and 0.355 hosts/parasitoid/day, respectively. Domesticated G45 had the lowest values for c0 (7.82 hosts/parasitoid), ψ (0.202 hosts/parasitoid) and ω (0.238 hosts/parasitoid/day) (Table 3). The transition rate values (Qp) for all generations were close to 1 (R0 ≈ c0), because T. brassicae almost always lays one egg in each S. cerealella egg.
The age-stage-specific parasitism rate (cxj) describes the mean number of S. cerealella that had been parasitized by individual T. brassicae at age x and stage j. The peaks of curves declined over generations. The highest and lowest number of the parasitized hosts in the rejuvenated population were observed in G31 (19.61 hosts/parasitoid) and G45 (13.27 hosts/parasitoid), respectively, at age of 11 days (Fig. 4).

Age-stage parasitism rate (cxj) in different generations (G) of the rejuvenated Trichogramma brassicae on Sitotroga cerealella eggs
The regressions between finite parasitism rate and generation number in the rejuvenated and domesticated populations were fitted to a cubic regression model (Fig. 5). The merit of the model was supported by the high value of the coefficient of determination (R2) for the domesticated (F = 144.903, P < 0.01, R2 = 0.984) and rejuvenated (F = 43.057, P = 0.002, R2 = 0.970) populations, indicating the parasitism potential of T. brassicae increased significantly after adding feral individuals but decreased markedly over subsequent generations.

Regression curve of finite parasitism rates (ω) with generations of Trichogramma brassicae reared on Sitotroga cerealella eggs in both domesticated and rejuvenated populations
Discussion
The success of a biological control, particularly augmentative biological control, depends on the effectiveness of natural enemies that have been released. These biocontrol agents may have been produced in mass rearing facilities over many generations. Developing the best rearing strategy is complicated by the ‘paradox of captive breeding’ in which increased quantity generally decreases quality (van Lenteren 2003). A decrease in the quality of the sequentially reared Trichogramma spp. has been found by different authors (Taghikhani et al. 2019). This problem is not limited exclusively to Trichogramma species and many natural enemies have been reported to experience this issue (Bellutti 2011). Routinely introducing field-collected individuals to the domesticated colonies of natural enemies is one commonly used method to keep genetic diversity over generations.
In our previous studies, a high reduction in the efficiency of T. brassicae over 45 generations of rearing, including a decrease in population and parasitism parameters was found (Ghaemmaghami et al. 2021). Here, the quality was improved in a primary colony of T. brassicae rejuvenated by adding feral individuals. Although most generations in the rejuvenated group had non-significant differences in the longevity with the primary population, females in the rejuvenated group laid more eggs than the domesticated. Cônsoli et al. (2010) mentioned that several factors may affect fecundity in Trichogramma spp., including host species, factitious or artificial hosts, egg size, age and nutrition. In the present study, the domesticated and rejuvenated populations both had the same condition, and any variation in the fecundity can be related just to add wild individuals. It seems that by increasing the rate of feral individuals added to primary colony, the process of reducing the quality of the wasp can be slowed down. However, this issue should be investigated by more studies in which different rate of feral individuals are added to the primary colony.
Although non-significant differences in the population growth parameters (GRR, R0, r, λ and T) were found among rejuvenated G31–G34, the rejuvenation drastically improved the population growth parameters, and they had higher population parameters than domesticated group. However, van Lenteren (2003) declared if the rearing conditions remain the same in the laboratory, the introduced wild individuals will be subjected to the same process of genetic selection. Declined parasitism parameters of T. brassicae after four generations of the rejuvenation may reinforce this inference.
Domestication can have remarkable evolutionary payoffs. For instance, it can induce both plastic and genetic modifications that limit the capacity of an organism to thrive in nature (Gering et al. 2019). Recent works showed that feral taxa undergo rapid evolutionary changes at loci controlling an array of fitness-related traits, including morphology, behavior and development (Moyers et al. 2017). Gene flow between domesticated and wild populations leads to important, diverse and context-dependent effects in fitness of recipient populations, which in turn can have important and unexpected roles in subsequent adaptation to changing (e.g., feral) environments (Gering et al. 2019). Domestication puts insects in a homogeneous environment where the challenges of natural selection are lacking (Cohen 2000), and in some cases, traits necessary for fitness are lost. Bartlett (1984) discussed the elimination of diapause genes and ‘startle response’ genes. Such inadvertent selection can be exacerbated by founder effect, where small starter population size leads to higher rates of genetic drift. These pitfalls are a legitimate concern for researchers and biological control practitioners who are interested in the use of augmentative biological control where laboratory-cultured natural enemies are to be used. Generally, cascading changes in the genetic pattern of a population reared in a novel environment is inevitable (van Lenteren 2003), and it is expected that natural selections reduce the genetic variability of such populations (Miyatake 1998). Moyers et al. (2017) stated that the process of domestication of wild species can result in an increase in the number, frequency and/or proportion of deleterious genetic variants that are fixed or segregating in the genomes of domesticated species for these species.
Sex ratio is an important trait in parasitoids influencing the financial profitability of mass rearing. In other word, produced male parasitoids in an augmentative release program have no pest control benefits. Generally, a reduction in the proportion of female individuals may happen in long-term mass rearing programs (van Lenteren 2003). Here, no effect of rejuvenation on sex ratio was found. Also, in our previous study, a clear declining trend in ratio of the produced females was not observed (Ghaemmaghami et al. 2021).
Conclusion
Optimizing mass rearing of arthropods for augmentative release programs is an extremely complex challenge. An ideal captive population should be able to adapt well to both the breeding environment and the field conditions where they need to control pests. Indeed, many of parasitoids, especially Trichogramma spp., are difficult to rear, creating a high potential for laboratory adaptation and for a large tradeoff with field performance. It seems adding feral individuals could reshape the reproductive performance of the wasps at least for four generations, and it needs to be repeated routinely. Furthermore, the quality may also be improved by adding a higher rate of feral individuals. However, this issue should be investigated by more studies in which different rate of feral individuals are added to the primary colony.
Availability of data and materials
Not applicable.
Abbreviations
- L:
-
Lightness
- D:
-
Darkness
- RH:
-
Relative humidity
- G:
-
Generation
- GRR:
-
Gross reproductive arte
- R 0 :
-
Net reproductive rate
- r :
-
Intrinsic rate of increase
- λ :
-
Finite rate of increase
- T :
-
Mean generation time
- APOP:
-
Adult pre-oviposition period
- TPOP:
-
Total pre-oviposition period
- s xj :
-
Age-stage survival rate
- f xj :
-
Age-stage-specific fecundity of female
- l x :
-
Age-specific survivorship
- m x :
-
Age-specific fecundity
- C 0 :
-
Net predation rate
- Q p :
-
Transformation rate
- ψ :
-
Stable predation rate
- ω :
-
Finite predation rate
- c xj :
-
Age-stage predation rate
- a xj :
-
Stable age-stage structure
References
Bahari F, Fathipour Y, Talebi AA, Alipour Z (2018) Long-term feeding on greenhouse cucumber affects life table parameters of two-spotted spider mite and its predator Phytoseiulus persimilis. Syst Appl Acarol 23:2304–2316
Bartlett AC (1984) Genetic changes during insect domestication. In: King EG, Leppla NC (eds.), Advances and challenges in insect rearing, USDA, ARS, New Orleans, LA., pp. 2–8
Bellutti N (2011) Effects of mass rearing on life-history traits of an invasive fruit moth species, Grapholita molesta (Busck). Master thesis. Institute of Forest Entomology, Forestathology and Forest Protection. BOKU Vienna, pp. 35
Chi H (1988) Life-table analysis incorporating both sexes and variable development rates among individuals. Environ Entomol 17:18–25
Chi H, Yang TC (2003) Two-sex life table and predation rate of Propylaea japonica Thunberg (Coleoptera: Coccinellidae) fed on Myzus persicae (Sulzer) (Homoptera: Aphididae). Environ Entomol 32:327–333
Chi H (2019) TWOSEX-MSChart: a computer program for the age-stage, two-sex life table analysis. http://140.120.197.173/Ecology/Download/TwosexMSChart.zip
Cohen AC (2000) Feeding fitness and quality of domesticated and feral predators: effects of long-term rearing on artificial diet. Biol Control 17:50–54
Cônsoli F, Parra JR, Zucchi R (2010) Egg parasitoids in agroecosystems with emphasis on Trichogramma. Springer, Dordrecht, p 479
Gering E, Incorvaia D, Henriksen R, Conner J, Getty T, Wright D (2019) Getting back to nature: feralization in animals and plants. Trends Ecol Evol 34:1137–1151
Ghaemmaghami E, Fathipour Y, Bagheri A, Talebi AA, Reddy GVP (2021) Quality control of the parasitoid wasp Trichogramma brassicae (Hymenoptera: Trichogrammatidae) over 45 generations of rearing on Sitotroga cerealella. Insect Sci 28:180–190
Hopper KR, Roush RT, Powell W (1993) Management of genetics of biological control introductions. Annu Rev Entomol 38:27–51
Huang YB, Chi H (2012) Age-stage, two-sex life tables of Bactrocera cucurbitae (Coquillett) (Diptera: Tephritidae) with a discussion on the problem of applying female age-specific life tables to insect populations. Insect Sci 19:263–273
Miyatake T (1998) Genetic changes of life history and behavioral traits during mass-rearing in the melon Fly, Bactrocera cucurbitae (Diptera: Tephritidae). Res Popul Ecol 40:301–310
Moghaddassi Y, Ashouri A, Bandani AR, Leppla NC, Shirk PD (2019) Effect of Ephestia kuehniella (Lepidoptera: Pyralidae) larval diet on egg quality and parasitism by Trichogramma brassicae (Hymenoptera: Trichogrammatidae). J Insect Sci 19(4):10
Moyers BT, Morrell PL, McKay JK (2017) Genetic costs of domestication and improvement. J Hered 109:103–116
Smith SM (1996) Biological control with Trichogramma: advances, successes, and potential of their use. Annu Rev Entomol 41:375–406
SPSS-Inc (2009) PASW statistics for windows. SPSS Inc, Chicago
Taghikhani N, Fathipour Y, Bagheri A, Talebi AA (2019) Generation-dependent fitness of the parasitoid wasp Trichogramma brassicae (Hymenoptera: Trichogrammatidae) in laboratory and insectarium condition. J Crop Prot 8:351–359
van Lenteren JC (2003) Quality control and production of biological control agents: theory and testing procedures. CABI, Wallingford
Wang ZY, He KL, Zhang F, Lu X, Babendreier D (2014) Mass rearing and release of Trichogramma for biological control of insect pests of corn in China. Biol Control 68:136–144
Yazdanpanah S, Fathipour Y, Riahi E, Zalucki MP (2022) Cost-effective and efficient factitious prey for mass production of Neoseiulus cucumeris (Acari: Phytoseiidae): assessing its quality compared with natural prey. Egypt J Biol Pest Control 32(1):16
Acknowledgements
This study is a part of the PhD dissertation of the first author and was funded by Tarbiat Modares University, which is greatly appreciated. We also would like to thank the Technical and Vocational Training Organization for providing facilities for this research.
Funding
This study was funded by Tarbiat Modares University (Grant Number 9430462002).
Author information
Authors and Affiliations
Contributions
EG involved in writing, investigation, data collection and data analysis; YF involved in supervision, review and editing and project administration; AB involved in formal analysis and review and editing; AAT involved in review and editing; GVPR involved in final review and editing and improvement. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Compliance with ethical approval.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ghaemmaghami, E., Fathipour, Y., Bagheri, A. et al. Rejuvenation improves the quality of Trichogramma brassicae Bezdenko (Hymenoptera: Trichogrammatidae) reared for many generations on Sitotroga cerealella (Olivier) (Lepidoptera: Gelechiidae). Egypt J Biol Pest Control 32, 79 (2022). https://doi.org/10.1186/s41938-022-00581-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41938-022-00581-z