
Abstract
White grubs are largely unsolved problems in vegetable and tuber production in East Africa. Novel Rwandan as well as international entomopathogenic nematodes (EPNs) were screened in nine laboratory bioassays and two small-scale field trials in 2014 and 2015. Soil-based laboratory bioassays revealed that all EPNs were able to infect Anomala graueri larvae (Coleoptera: Scarabeidae), although a relatively high number of infective juveniles were needed. Rwandan EPNs were as infectious as their corresponding international strains. At 100 infective juveniles per larva, the Heterorhabditis bacteriophora strains (Rwanda14-N-C4a and international H06) as well as the international Steinernema carpocapsae All caused 18 to 22% grub mortality within 7 days compared to the control. At 1000 infective juveniles, both H. bacteriophora strains as well as the Rwandan S. carpocapsae RW14-G-R3a-2 killed 34 up to 58%. The Rwandan Steinernema RW14-M-C2a-3 least performed in the bioassays (2 to 6%). In two Irish potato fields, the into-soil-applied 1 × 109 infective juvenile Steinernema RW14-M-C2b-1 per hectare reduced 29 ± 33% and 96 ± 3% of grubs within 30 and 60 days, respectively. About 1 up to 2.5 × 109 infective juvenile Steinernema longicaudum X7 per hectare reduced 77 up to 85% of grubs until day 30, respectively, and 82 up to 95% until day 60. Avermectin + Chlorpyrifos tuber coatings reduced 39% of grubs, and Fipronil + Chlorpyrifos reduced 27%, whereas handpicking did not help much. In conclusion, at least Steinernema RW14-M-C2b-1 and S. longicaudum X7 are promising for managing white grubs in tuber production, this is at a rate of at least 1.5 × 109 EPNs per hectare, but H. bacteriophora RW14-N-C4a needs further field research. Findings will support the biocontrol product development in Rwanda, including registration if any would be needed for native macrobial biocontrol agents.
Similar content being viewed by others


Background
Similar to many agricultural regions across the globe, East African agriculture suffers considerable losses due to soil-inhabiting insects (Trutman and Graf, 1993; Cock and Allard, 2013; Nyamwasa et al., 2017). Those pests usually damage early developmental stages of horticultural crops, but can also attack mature below-ground plant parts, such as tubers. Damaged tubers are prone to secondary infections, thus reducing their shelf-life and marketable value (Toepfer et al., 2016).
At least 40 soil insect species impact agriculture in Rwanda. The most troublesome are scarabeid white grubs in the genera Anomala and Hoplochelus or in the tribe Melolonthini, followed by cutworms (Agrotis species), bean flies (Ophiomyia species) and tuber-attacking weevils (Cylas species) (Nyamwasa et al., 2017). This pattern is found across East Africa, but also in other world regions, such as in East Asia (Guo et al., 2013; 2015; Toepfer et al., 2016).
Since about 2011, white grub outbreaks have been more and more frequently occurring across Rwanda, inflicting heavy damage in many cultivated vegetable and tuber crops. In some cases, up to 60 grubs were found per square metre (Nyamwasa et al., 2017). Those included Copris, Oniticellus, Onitis, Onthophagus, Pachysoma, Anomala, Lepadoretus, Adoretus, Cyclocephala, Pycnoschema, Hoplochelus, Maladera, Trochalus, Lepiserica species and Melolonthini genera from Rwanda, but probably there are even more. However, the pest status of some of those genera is unclear, and for some pest species, it is not clear why their populations recently increased. It is hypothesised that the continuous strip cropping of the same combination of vegetables or vegetables with field crops favours, as does monoculture, the survival and build-up of certain soil pest populations because many of them are restricted to certain plants or have long life cycles over several cropping seasons (Chen et al., 2004; Toepfer et al., 2014; Li et al., 2016).
Unfortunately, white grub pests are difficult to control because of their hidden nature, sometimes combined with insufficient knowledge on their life cycles and below-ground ecology. It is difficult to effectively use synthetic insecticides against soil pests due to problems of leaching, adsorption or rapid break down, and the impracticality of applying contact pesticides (Mayo and Peters, 1978; Furlan et al., 2006). Soil pesticides are highly concentrated formulations and can endanger the handling farmer. They are often also highly toxic, with many being restricted or indeed banned (WHO, 2009). As for Rwanda, the availability of soil pesticides is limited. They are often too expensive or impractical for small-holder growers (Toepfer et al., 2014). Socioeconomic constraints have kept pesticide use in Africa the lowest among all the world regions (Musebe et al., 2017), a fact also valid for Rwanda. These factors combined present a barrier to the effective control of soil pests in Rwanda.
As a consequence, alternative strategies such as entomopathogenic nematodes (EPNs; syn. Entomoparasitic nematodes) could be the solution, but research from East Africa is scarce. Therefore, EPNs are being investigated in an Agricultural Technology Transfer project funded by the UK Department for International Development (DfID) (Li et al., 2016; Toepfer et al., 2016). For example, EPNs of the genus Heterorhabditis and Steinernema are found naturally in soils throughout the world and possess a number of attributes that facilitate their use as biological control agents. They have the ability to search for hosts and, due to their high reproductive potential, can react to changing pest densities. They are non-toxic to humans, considered safe to the environment, and can often be mass cultured, formulated and applied with common agricultural practises. Globally, mainly H. bacteriophora, H. marelatus, S. glaseri and S. scarabaei are known to be virulent to white grubs (Klein and Georgis, 1992; Selvan et al., 1993; Ansari et al., 2003; Koppenhöfer and Fuzy, 2003 and 2009; Grewal et al., 2005; Bal et al., 2014; Guo et al., 2015), but some, such as S. scarabaei, are difficult to mass-rear (Koppenhöfer and Fuzy, 2003 and 2009). Prior to the here-reported study, locally adapted EPNs were searched for in semi-natural and small-holder farming habitats across Rwanda (Yan et al., 2016). The found isolates (species/strains) were described as Steinernema sp. RW14-M-C2a-3, Steinernema sp. RW14-M-C2b-1, S. carpocapsae RW14-G-R3a-2, H. bacteriophora RW14-N-C4a and Heterorhabditis sp. RW14-K-Ca. They are currently maintained at the biocontrol agent factory at the Rubona Research Centre of the Rwanda Agriculture Board and needed investigation on their potential in killing soil insect pests, such as the white grub pests of Rwanda.
We therefore implemented bioassay-based laboratory screenings and field efficacy trials in Irish potatoes. Not only were the Rwandan nematodes assessed, but they were also compared with three international EPNs commonly used for soil pest control. As for the field trials, EPNs were also compared to insecticide tuber coatings and to the locally practised handpicking of grubs during soil preparations and mechanical weed control. This study will serve as a baseline for future work on using these EPN species/strains for white grub management in Rwanda or neighbouring countries. Results will particularly help to decide which EPN species should be mass-produced and applied to effectively control soil-inhabiting insect pests in vegetable and tuber production in a safe and environmentally friendly way.
Methods
Target insect pests
The target insect pests were larvae of scarabeid beetles, i.e. white grubs known to damage vegetables and tubers in Rwanda (Nyamwasa et al., 2017). These included Anomala as well as Hoplochelus species (Coleoptera: Scarabeidae).
As for laboratory screenings, Anomala graueri grubs were field-collected. They were identified using morphological and molecular DNA comparison methods (Nyamwasa et al., 2017). They were collected from ploughed fields after sweet or Irish potatoes in Akanyandoli in Nyamagabe district in September 2014 as well as in February and March 2015. More larvae were collected between rows of sweet potatoes and from grassy fallows after sweet potatoes near Mujuga and Kitabi in Nyamagabe district in October and November 2015. White grubs were visually searched for through digging with a spate or hook about 30 cm deep through the soil. Each grub was individually placed into a soil-filled small plastic container with holes in the lid. They were transported to the laboratory and held at 24 ± 2 °C for no more than 1 day until testing. Tested larvae were mainly of third instar (mean 2.6 ± 0.2 SD cm long).
As for field trials, sites were chosen wherefrom farmers had reported serious crop damage by white grubs the previous season. Population levels of white grubs were assessed through soil samplings prior to planting and again during weeding 15 days after crop emergence and during earthing up of Irish potatoes. Natural white grub populations were between 1 and 4 grubs per square metre. The dominant white grub genera were Anomala and Hoplochelus species as per morphological and DNA comparisons. But identification to species level was not possible in most cases due to lack of comparative information in DNA databases and difficulties to morphologically discriminate larval instars (Nyamwasa et al., 2017).
Source and handling of EPNs
The Rwandan EPN species/isolates, Steinernema sp. RW14-M-C2a-3, Steinernema sp. RW14-M-C2b-1, S. carpocapsae RW14-G-R3a-2 and H. bacteriophora RW14-N-C4a originated from surveys in several provinces of Rwanda in 2014 (Yan et al., 2016) (Table 1). Steinernema longicaudum X7, H. bacteriophora H06 and S. carpocapsae All are commonly used biocontrol agents of soil insect pests in many world regions (Poinar Jr., 1990; Shen and Wang, 1991; Yan et al., 2013) and served as positive controls. They were provided by Lvbenyan Biotech Ltd., a spin-off of the Guangdong Institute of Applied Biological Resources (GIABR) in China.
All EPNs were reared at the biocontrol agent facility of RAB Rubona (Holmes et al., 2015) using the in vitro semi-solid, sometimes called the solid system as per Shapiro-Ilan and Gaugler (2002) and Kaya et al. (2006). The harvested infective juvenile (IJ) EPNs were acclimated to room temperature for 1 h. Their viability was assessed on the basis of movements checked under a stereomicroscope and considered alive when actively moving or showing response after probing with a needle. EPN suspensions were used for the experiments when more than 90% of IJs were viable.
All EPNs were assessed in laboratory bioassays, but only the Rwandan Steinernema sp. RW14-M-C2b-1 and the international S. longicaudum X7 in field efficacy trials, as those were the easiest to mass-rear.
Laboratory bioassays
Bioassay-based laboratory screenings of EPNs were carried out on A. graueri larvae under controlled conditions to compare four Rwandan EPN species/isolates versus three international EPNs commonly used in biological control (Table 1). This is a pre-requisite of deciding for which EPN a mass production technique should be established.
Screening for EPN infectiousness
In total, nine experiments were implemented to screen seven different EPNs at five different concentrations for their infectiousness on third instar Anomala graueri larvae (Table 2). The infectiousness was here defined as the insect mortality caused by EPNs that include the processes of host finding, host recognition, penetration, release of bacteria, bacteria proliferation, overcoming insect immune response, nematode proliferation and insect death as per Peters and Smits (2000), in other words the situation in a bioassay arena.
In each of at least three experimental replicates per EPN and concentration, three sets of ten small bioassay arenas were set up containing one white grub each (Table 2), as bioassay arenas served toothpick boxes (3 cm diameter × 7 cm high, 3 holes of 2 mm in the lid). The arenas contained 25 g sandy loam soil (78% sand, 14% loam, 5% clay, 0.4% organic matter; pH 6.8). The soil originated from an experimental field at RAB Rubona station north of Huye town in the southern province of Rwanda. The soil had been sieved at 2 mm mesh size, and autoclaved at 121 °C for at least 15 min to destroy any natural enemy of grubs or EPNs as well as any potential natural population of EPNs. The soil was then dried at 100 °C for 24 h and then adjusted to 15% w/w moisture. An about 1-cm3 piece of fresh Irish potato was added in each arena as food for the white grubs. The arenas were kept at room temperature of 24 ± 2 °C for 2 h, and then only arenas with moving grubs, e.g. which had entered into the soil, were selected for experiments.
Up to five different concentrations were tested; that is, 100, 200, 300, 400 and 1000 IJs per white grub, thus per 28 m2 soil surface in a bioassay arena (Table 2). The 100 to 1000 IJ step in concentrations reflects a log10 step, as commonly used in dose trials allowing a more linear response analyses. One-millilitre suspensions containing the required concentration of IJs were distributed across the soil surface of each arena using a precision pipette. No direct application onto an insect body was carried out. Water without nematodes was used on the untreated controls. The arenas were placed in the dark at 24 ± 2 °C for 14 days.
Grub mortality was assessed after 4, 7 and 14 days. In addition, mortality likely caused by EPNs and not by other mortality factors was assessed as follows: In case a dead grub was found, it was washed in 1% NaOCl (syn. bleach, jijk) or in 70% ethanol, and then washed in clean water. Individual cadavers were then placed onto a slightly moist filter paper in a 9-cm Petri dish for another 4 days and then dissected to confirm adult or young EPNs under a stereomicroscope. The infectiousness was calculated as the corrected (=relative) mortality of grubs in each treatment minus the natural mortality found in the untreated controls (Figs. 1 and 2). Distributions of data were investigated using histograms and the one-sample Kolmogorov-Smirnov test against normal distribution (Kinnear and Gray, 2000). For non-normal distributed data, log10(x + 1), sqrt(x + 1), or arcsine(x) transformations were used to achieve better data distribution. The influences of the EPN species/strains/isolates, the concentration of EPNs and the assessment period of 4, 7 and 14 days after treatment were examined on grub infectiousness through GLM univariate analyses (binomial with logit link function). Due to the large number of concentrations and time steps assessed, treatments and time steps, the p values obtained from the post hoc tests were adjusted to q values to reduce the probability of type I errors of p values using the false discovery rate-correction of Benjamini & Hochberg through the package Q-VALUE in R (R Development Core Team, 2009).

Assessing which exposure period of Anomala graueri grubs to entomopathogenic nematodes leads to most distinct differences between treatments in laboratory bioassays. Mean value and variability separately presented to better assess (a) differences between percent-corrected mortality compared to control (mortality in EPN treatments minus natural mortality in untreated controls), and (b) smallest data variability. n = 6 to 15 grubs assessed per EPN species/strain and per concentration in single-grub arenas with soil

Comparing the infectiousness of Rwandan (RW) and international entomopathogenic nematodes on Anomala graueri grubs in soil-based laboratory bioassays, depending on species/strain and on applied concentrations of 100, 200, 300, 400 and 1000 infective juveniles per grub equalling 14, 28, 42, 57 and 140 IJs per square centimetre soil. Dark grey bars = heterorhabdits, light grey = steinernematids, lined = verified in field trials (Figs. 4 and 5.); 7-day-corrected morality shown, this is mortality in EPN treatments minus natural mortality in untreated controls. Three sets of 10 bioassay arenas with single grubs in soil used per each of 3 to 15 experimental replicates; letters on bars indicate difference according to multiple comparison Tukey post hoc test at p < 0.05, additionally fdr-corrected among concentration experiments; error bars = SEM
Field efficacy trials
To evaluate EPNs’ potential under field conditions, field efficacy trials were implemented for controlling Anomala and Hoplochelus grubs. Soil applications of the Rwandan Steinernema sp. RW14-M-C2b-1 and the international S. longicaudum X7 were compared with insecticide-tuber coatings, handpicking and untreated controls.
Field sites
Two experimental field sites of 0.13 ha were established in 2015 for assessing the control effect of EPNs against natural populations of white grubs. The fields are referred to as field M in Kinigi in Musanze district, northern zone of Rwanda (2285 m, S01° 2′ 11.1″, E29° 35′ 50.8″) and field N in Akanyandoli in Nyamagabe district, southern zone (2153 m; S20° 30′ 43.2″, E29° 30′ 007″).
Both fields were ploughed twice prior planting for weed control as well as for soil preparation. Irish potatoes (Var. Cruiser, seed program at Rwanda Agriculture Board) were planted in field N on 5 May 2015 and in field M on 15 May 2015. At the day of planting, well-composted farm cow manure (20 t per ha) and mineral fertiliser (NPK 17-17-17 at 300 kg per ha) were applied into the planting holes just before planting. Irish potatoes were planted into 8- to 10-cm deep holes at 30 cm distance between plants and 80 cm between rows, equalling 41,000 plants per hectare. About 1-kg seed tubers were used per 4 m2, equalling 2.5 tons per hectare. About 60 cm buffer spaces were kept between treatment plots. Mechanical weeding was implemented 15 days after potato germination, and soil was earthed up along potato rows at potato flowering stage. Natural white grub populations were up to one grub per square metre in field M and up to four grubs per square metre in field N (Fig. 3). Field M was harvested on 24 September 2015 and field N on 22 September 2015.

Population dynamics of Anomala and Hoplochelus white grubs in Irish potato fields during experimental periods in field M in Musanze and field N in Nyamagabe in Rwanda in 2015. Assessed from the soil at 0 to 15 cm below and around 10 plants in 4 untreated control plots in each field per date. Error bars = SD. Trendline = moving average
Experimental setup and treatments
In total, eight different treatments were implemented against white grubs, consisting of two different EPN species/strains (one tested in three concentrations), two insecticide tuber coatings, handpicking of grubs and an untreated control.
In detail, IJs of the Rwandan Steinernema sp. RW14-M-C2b-1 (60,000 per row-metre, equalling 0.75 × 109 IJs/ha) and the international S. longicaudum X7 (60,000/row-metre, equalling 0.75 × 109 IJs/ha; 120,000/row-metre, equalling 1.5 × 109 IJs/ha; 200,000/row-metre, equalling 2.5 × 109 IJs/ha) were applied with water (2000 l/ha) into the soil along the plant rows. For that, an about 5-cm-deep furrow had been made at about 10 to 15 cm distance to the plants. EPNs formulated in sponge (Yan et al., 2016) were diluted in water and transferred through a sieve into an 8-l knapsack sprayer. Nozzles were removed and EPNs applied into the soil. EPNs were applied about 30 days after planting (4 June 2015) in field N, and about 55 days after planting in field M (10 July 2015).
Liquid microcapsule formulations of 0.2% Avermectin + 14.8% Chlorpyrifos (Pesticide factory at IPP, CAAS, Beijing, China) or 3% Fipronil + 15% Chlorpyrifos (Yoloo Pesticide Co., Ltd., Beijing, China) were coated onto the to-be planted tubers. Briefly, 2.5 ml of the liquid microcapsule formulation was diluted in 10 ml water, then pulled over around 1 kg of tubers and manually mixed to achieve surface coating. This equals a 1: 400 ratio of the products versus tuber weight. After coating, potatoes were dried in the sun for 5 to 10 min. One kilogram of potato seed tubers was used per 4 m2, equalling 2.5 tons per hectare (totalling 6.25 l product in 25 l water/ha). Grubs were handpicked from the freshly opened furrows during planting and again during at first mechanical weeding 15 days after germination, and finally during earthing up at plant flowering stage. Larvae were visually searched for and picked up.
The control plots were only treated with water, that is, at a rate equalling 2000 l of water per hectare.
The study was conducted according to the efficacy evaluation standards PP 1/212 and PP 1/152 of EPPO (Anonymous, 2007). Trial permits were obtained from the Rwanda Agriculture Board Authorities. The experimental fields were divided into plots of 4 × 10 m. Each treatment was applied on four such plots per field, totalling 32 plots per field. A randomised block design was used for the placement of plots. Thus, the within-field replicate number per treatment was four totalling eight for the entire study.
Assessment of EPN efficacy at reducing white grubs
In each of the four plots per each of the treatments, soil beneath 10 randomly pulled-out plants were assessed for white grub population levels, and this in both fields at six time steps from planting until harvest (Fig. 3). Grubs were visually searched for by crumbling an about 40 × 40 × 20 cm deep area of soil per plant hole. Only for the assessment before planting, samples were taken from five random spots of 1 × 1 × 0.25 m soil per treatment per plot, and data converted to per-plant-data depending on the expected plant density.
In field M, this was done at the date of planting (15 May 2015), 57 days later (10 July 2015, date of EPN treatment), then 64, 94 and 124 days later (= 7, 30, 60 days after EPN treatment) and 133 days later, i.e. at harvest (24 September 2015, 77 days after EPN treatment).
In field N, this was done at the date of planting (5 May 2015), 31 days later (4 June 2015, date of EPN treatment), then 38, 68 and 98 days later (= 7, 30, 60 days after EPN treatment) and 141 days later, i.e. at harvest (22 September 2015, 111 days after EPN treatment).
The mean efficacy of each treatment was calculated as the reduction of grubs relative to the untreated control at each time step and as a seasonal average., i.e. the corrected % efficacy = [100 − (insects in treated plots * 100/insects in the untreated control)].
Distributions of data were investigated using histograms and one-sample Kolmogorov-Smirnov test against normal distribution (Kinnear and Gray, 2000). As for distribution of average grub data per plot along treatments, all data appeared normally distributed; except the 7-day data needing sqrt(x + 1) transformation. As for the distribution of raw grub data, only the data at harvest time were normal distributed and, after log10(x + 1) and sqrt(x+) transformation, also the data from day 7—after EPN treatment. The 30- and 60-day raw data remained non-normal distributed, regardless of transformation.
The influences of treatments on white grub populations were examined through GLM analyses or through independent samples Kruskal-Wallis H test (nonparametric analogue of GLM analyses). Once a significant factorial effect was detected by those models, the averages of grubs per plot were compared between treatments at each date. For this, Tukey post hoc range test was used in case of normally distributed data and equal homogeneity of variances (Kinnear and Gray, 2000).
Assessment of EPN efficacy at preventing yield losses
In each of the four plots per each of the treatments, 10 plants were harvested and their tubers weighted immediately. Differences among different treatments were analysed at p < 0.05 via multiple comparison Tukey post hoc range test after GLM.
Assessment of EPN persistence
The nematode-baiting method with alternative host insects as baits was employed as per Bedding and Akhurst (1975) and Yan et al. (2016) to assess whether the applied EPNs had established and persisted in the field soil. Around 400 to 600 g of soil was taken from the holes remaining from each of the 10 sampled plants for white grub assessments (see above). Soil samples were taken at a depth of 10 to 20 cm. The 10 soil samples per plot were then mixed, and a 500-g mixed subsample was taken. The mixed subsamples were placed into plastic containers and returned to the laboratory. Then, five late instar Galleria mellonella (Lepidoptera: Pyralidae) were added to each mixed sample to bait for potential EPNs (container 20 × 12 × 6 cm) (Caroli et al., 1996). The baited samples were then incubated in the dark at room temperature (24 ± 2 °C). The Galleria larvae were recovered about 5 days later, and dead Galleria counted. Rotting larvae, those with a bad (viz. bacterial) odour, or larvae with multiple colours (EPN-killed larvae show one colour) were discarded, as they had unlikely been killed by EPNs (Grewal, 1992). The cadavers of potential interest were surface-sterilised in 0.5 to 1% NaOCl for 3–5 s, washed in water, then individually placed onto a nematode trap modified as per White (1927). Briefly, the trap consisted of cellulose tissue on an inverted Petri dish lid onto which the cadaver was placed. The dish was placed into a larger Petri dish. The dish was left open for 1–2 days to desiccate and kill saprophytes (EPNs inside cadavers are protected from desiccation). Then, a bit of sterilised tap water was added to the tissue. Between 4 to 7 days later, the traps were checked daily for IJ emergence from the cadavers. Emerging nematodes were visually assessed under a stereomicroscope (×10 magnification) for saprophytic nematodes and EPNs. A mixed soil sample was considered EPN positive in case that at least one of the five Galleria larvae was infected by EPNs per sample (Kurtz et al. 2007). Proportions of positive samples, thus persistence, were assessed over time using Pearson correlations, and between treatment using Tukey multiple comparisons after GLM.
Results and discussion
To evaluate the potential of newly described Rwandan EPNs (Yan et al., 2016) for the development of biological control products against soil insect pests, bioassay-based laboratory screenings and small-scale field efficacy trials had been implemented and the results, as presented here, are promising.
Laboratory bioassays
The small arena bioassays revealed that all the four tested Rwandan EPN species/strains were able to infect and kill white grubs, although it took quite a high number of infective juveniles per grub larva. Nevertheless, this is considered a good basis for further research, as it is often not too easy to find EPNs being infectious to grubs as they are often able to defend themselves to EPNs or other ground-living natural enemies (Ansari et al., 2003; Guo et al., 2013; Laznik and Trdan, 2015). And other EPNs, such as S. scarabaei, are not easily mass-produced. Consequently, there are only few EPN products on the market against white grubs, globally. However, as the tested Rwandan EPNs originated from small-holder farming habitats of Rwanda (Yan et al., 2016), they are, potentially, more adapted than EPNs from other habitats, such as sea shores. Moreover, the tested Rwandan EPNs seem to be as good as international species/strains commonly used for soil pest control. This is a hint that the tested EPNs may have evolved in areas of white grub prevalence in Rwanda.
EPN infectiousness against grubs
The overall accumulated mortality of A. graueri white grubs due to EPNs, as verified by dissections, was 13 ± 15 SD % until day 4; 17 ± 17% until day 7; and 20 ± 18% until day 14 (n = 291). Comparable results were obtained when correcting the total accumulated mortality of grubs in the treatments to the natural mortality in the untreated controls. That is, 13 ± 15% more grubs died in the EPN treatments than in the control until day 4; 16 ± 18% until day 7; and 17 ± 19% until day 14. Without any treatment, 6 ± 7% of grubs naturally died until day 4; 10 ± 13% until day 7; and 14 ± 18% until day 14.
Infectiousness depending on EPN species/strain
Overall, tested EPNs appeared to have medium infectiousness against white grubs in the laboratory bioassays. Nevertheless, all tested EPNs, regardless if Rwandan or international, were able to infect and kill A. graueri grubs (Figs. 1 and 2). However, species/strains differed in their infectiousness (GLM: “species/strain” for 7 day corrected mortality: F 6;290 = 16, p < 0.0001; “species × concentration”: F 24;290 = 2.5, p < 0.0003, adjusted R2 = 0.33; Fig. 2). The Rwandan H. bacteriophora RW14-N-C4a and S. carpocapsae RW14-G-R3a-2 reached the efficacies of their corresponding international strains, indicating that local EPNs could be used as biocontrol agents against Anomala grubs.
The heterorhabdits slightly more consistently and usually more effectively killed the grubs than did most steinernematids (Fig. 2); except for the Rwandan and international S. carpocapsae which reached the infectiousness level of the heterorhabdits in some of the assays.
On average across concentrations, the Rwandan H. bacteriophora RW14-N-C4a killed 29 ± 18 SD % of grubs, the international H. bacteriophora H06 27 ± 9%, the Rwandan S. carpocapsae RW14-G-R3a-2 14 ± 11% and the international S. carpocapsae All 18 ± 9%. Steinernema sp. RW14-M-C2a3 was, overall, the least performing (4 ± 2% of grubs killed across concentrations, Figs. 1 and 2). The Rwandan Steinernema sp. R14-M-C2b1 (10 ± 3%) and the international S. longicaudum X7 (11 ± 3%) were both of comparable medium, but relatively constant infectiousness (5 to 18% mortality across different concentrations), as well as easily mass-reared (data not shown), and therefore suggested for field trials.
Field assessments are the logical next step, because the infectiousness data from EPNs in the laboratory can only partly be transferred to the field situation (Peters et al., 1996). This is, for example, because the foraging behaviour of EPNs is less important for the attack of host insects in small arenas of bioassays, than under field conditions. Field conditions also require a longer survival of EPNs without the host, i.e. before finding the host, than in small arena bioassays.
Infectiousness depending on EPN concentration
The concentration of applied EPN species/strains influenced their infectiousness on A. graueri grubs (GLM “concentration” for 7 day corrected mortality: F 4;290 = 5.5, p < 0.0003; “concentration × species/strain” F 24;290 = 2.5, p < 0.0003, adjusted R2 = 0.33) (Fig. 2). This effect was due to an increasing infectiousness at high concentration of 1000 IJs per larva, whereas concentration between 100 and 400 IJs did not matter much (GLM F 3;236 = 0.8, p = 0.49). When analysing concentration effects separately per EPN species, often no dose efficacy-response was detected, that is, for S. carpocapsae All, Steinernema sp. RW14-M-C2a-3, Steinernema sp. RW14-M-C2b-1 and S. longicaudum X7 (GLM F 3;44 to 65) < 3, p > 0.05). Only the following species/strains showed a positive dose-infectiousness response, i.e. H. bacteriophora RW14-N-C4a and H06 and S. carpocapsae RW14-G-R3a-2. In other words, at low concentrations of 100 to 200 IJs per larva, both the Rwandan and international H. bacteriophora strains as well as the international S. carpocapsae All performed best (12 to 30% additional mortality compared to untreated control) (GLM in Fig. 1). At high concentrations of 1000 IJs per larvae, again both H. bacteriophora strains were the best as well as the Rwandan S. carpocapsae RW14-G-R3a-2 (34 to 58% mortality compared to control).
In many cases, results were variable, and therefore a clear dose-response can hardly be concluded. This is typical for natural strains of EPNs that have not yet gone through many cycles of mass production and therefore through a selection for constant traits. It also shows that higher concentrations than 1000 IJs per grub should probably have been included in the experiments to get better dose-response trendlines. Nevertheless, we conclude that all Rwandan EPNs are worth to be further investigated on a number of target soil insect pests of Rwanda.
Infectiousness depending on exposure period to EPNs
Overall, the 7-day assessment appeared to be promising to detect differences in the infectiousness of the different EPN species/strains as well as concentrations. Differences among EPN species/strains in their infectiousness on A. graueri grubs over time of exposure were more obvious at low EPN concentrations than at medium or high concentrations (Fig. 1a). Variability of data depended more on the EPN species/strain or concentration (see large SEMs for 1000 IJs per grub in Fig. 1b), than on exposure period. In other words, increasing exposure period from 4 up to 14 days did not consistently increase or decrease variability of data, except at low concentrations (see below).
As for low EPN concentrations of 100 to 200 IJs per larva, differences between EPN species/strains seemed most obvious for the 7-day assessment as data later became disturbed by increasing natural mortality. Infectiousness usually only little increased with time (Fig. 1a), except from day 4 to day 7 for S. carpocapsae All and Steinernema sp. RW14M-C2b-1, and up to day 14 for Steinernema sp. RW14M-C2a-3 and Steinernema sp. RW14M-C2b-1 (Kruskal-Wallis H tests at p < 0.05, d.f. 2; 40 to 60). For all tested EPNs, the variability of relative infectiousness increased over time due to an increase in natural mortality (Fig. 1b). In conclusion, the 7-day assessment should provide reliable information from the bioassay on differences between EPNs at an acceptable variability.
As for medium EPN concentrations of 300 to 400 IJs per larva, infectiousness did not or only little increase with time (Fig. 1). The 14-day assessment added little additional information and/or was disturbed by natural mortality, as seen in the decreasing values of H. bacteriophora RW14-N-C4a with time. The variability of relative infectiousness data was comparable among time periods, but increased in few cases for the 14-day assessment (Fig. 1). In conclusion, the 4- and 7-day assessment should provide reliable information from the bioassays.
As for high EPN concentrations of 1000 IJs per larva, infectiousness increased particularly after day 4 up to day 7, but only in few cases or not at all after day 7 (Fig. 1). This indicates that the high concentration led to maximum infection already at day 7 and no additional information could be obtained running the assays until day 14.
Field efficacy trials
White grub infestation
The natural population levels of grubs ranged from 0.1 to 0.25 larvae per plant in field M and from 0.3 to 0.9 larvae per plant in field N in 2015 (average across fields and cropping season 0.3 ± 0.29 SD larvae per plant). This equals about 4000 to 10,000 white grubs per hectare at site M and 4000 to 35,000 at site N, a pest level that can lead to significant yield loss as a single grub usually destroys at least one planted tuber.
At day of planting Irish potatoes, 0.1 ± 0.03 grubs were found per plant in field M and 0.7 ± 0.17 in field N. Before EPN treatment about 30 to 55 days after planting, 0.1 ± 0.1 grubs were found per plant in field M and 0.9 ± 0.52 in field N. Subsequently, the population remained relatively stable until harvest in field M (see untreated controls), but it decreased within 30 days after planting in field N until it remained relatively stable until harvest M (Fig. 3). This means, during the period of agents’ efficacy assessments, a relatively constant natural pest population was present. The overall average pest populations across time periods was not different between the two fields (independent samples t test, t 78 = −1.2, p = 0.26).
EPN efficacy at reducing grubs
Overall, treatments reduced Anomala and Hoplochelus white grub numbers (GLM, averages pooled over season; F 7;79 = 2.4; p = 0.03; adjusted R2 = 0.1; Fig. 4). Variability of grub data between plots was high as well as between time steps (Fig. 4). Therefore, it was difficult to come to quantitative conclusions particularly when separately looking at grub values of each field and time step.

Efficacy of entomopathogenic nematodes, insecticide tuber coatings and handpicking at reducing Anomala and Hoplochelus white grub numbers depending on time periods. EPN concentration 1 = 0.75 × 109 infective juveniles (IJs)/ha; concentration 2 = 1.5 × 109 IJs/ha; concentration 3 = 2.5 × 109 IJs/ha. Field M in Musanze and field N in Nyamagabe districts in Rwanda in 2015. Four plots per treatment per field. Different letters on bars indicate differences among different treatments at p < 0.05 according to multiple comparison Tukey post hoc range test after GLM. Error bars = SEM
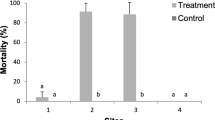
When standardising grub data to their respective controls and combining data of both fields, the control effects of treatments became obvious (Fig. 5). Biological and insecticidal treatments were effective in reducing grubs, sometimes at highly efficient rates. Biological and insecticidal treatments were comparably effective when considering overall effects along the entire season (multiple Tukey post hoc comparisons, all p > 0.05). Control effects (compared to untreated controls) were highest about 60 to 90 days after planting, but less at earlier stages as well as towards harvest time. Only the handpicking method did not significantly reduce white grubs (Fig. 5).

Percentage of entomopathogenic nematode-positive soil samples collected from two experimental Irish potato fields in Rwanda treated with entomopathogenic nematodes. Nematode concentration 1 = 0.75 × 109 infective juveniles (IJs)/ha; concentration 2 = 1.5 × 109 IJs/ha; concentration 3 = 2.5 × 109 IJs/ha. Field M in Musanze and field N in Nyamagabe districts in Rwanda in 2015. Four plots assessed per treatment per field per time step. GLM revealed treatment effects, but Tukey multiple comparison did not detect differences between each treatment combination. Error bars = SD
As for EPNs, effects were highest about 30 to 60 days after their application when they reached up to 87 and 96% control on average across fields (Fig. 4). In contrast, 7 days after treatment, hardly any effects of EPNs were observed. This is because EPNs needed time to find and kill the hosts. However, the 7-day assessment as a first sample time is a common practice to evaluate dynamics of EPN efficacy after application (Yan et al., 2013). But, it is not surprising that shortly after the EPN treatment, still no effects were found.
The Rwandan isolate Steinernema sp. RW14-M-C2b-1 reached 29 ± 33% efficacies after 30 days and 96 ± 3% after 60 days, an increase likely due to EPN propagation in the soil. The Rwandan isolate was at least as good as the international strain S. longicaudum X7 or even better (e.g. see at concentration of 0.75 × 109 IJs/ha in Fig. 4).
The international S. longicaudum X7 reached, at least at medium and high application concentrations, high control efficacies (77 ± 16 up to 85 ± 9% respectively about 30 days after application, and 95 ± 3% and 82 ± 17% after about 60 days). Also, the low concentration reduced grubs, but it took longer to reach effects (Fig. 5). As for other studies, S. longicaudum X7 is known to attack white grubs, such as Holotrichia parallela or H. oblita in peanut fields in China, and is used by growers for biological control purposes (Guo et al., 2013, 2015).
However, more field trials with more Rwandan EPN species and strains will be needed once the EPN-based biocontrol factory at RAB Rubona will run at full capacity. Particularly, both heterorhabditids, i.e. the Rwandan H. bacteriophora RW14-N-C4a and the international H. bacteriophora H06, need further investigation under field conditions. Both revealed promising infectiousness levels in the laboratory screenings. Moreover, H. bacteriophora is known to attack white grubs, such as Popillia japonica in turf grass (Klein and Georgis, 1992; Selven et al., 1993; Downing, 1994), and H. parallela and H. oblita in peanut fields and grasslands in China (Guo et al., 2013, 2015).
As for the insecticides, both were comparably effective on average over the season, this is 39 ± 35% for Avermectin + Chlorpyrifos and 27 ± 40% for Fipronil + Chlorpyrifos (independent samples t test, t 18 = 0.7, p = 0.49). However, 37 to 60 days after transplanting, Avermectin + Chlorpyrifos reached efficacies of 58 up to 87%, whereas Fipronil + Chlorpyrifos only reached 4 to 44%. However, the pesticides are difficult to be compared to the EPN treatments due to different application types (coating versus into-soil stream spray), application periods and water amounts. Thus, it is likely that the insecticides have caused effects earlier in the season, i.e. soon after they had been applied at the moment of planting. It needs to be, however, mentioned that Rwandan small-holder farmers do not have or have only limited personal protective clothing. Therefore, there is an acute risk that farmers are intoxicated (WHO, 2009) when coating tubers with pesticides, and therefore this is not advised. Chlorpyrifos is of WHO acute toxicity class II, thus moderately hazardous and Fipronil and Avermectin are slightly hazardous (WHO, 2009).
As mentioned above, the local common practice of handpicking did not significantly reduce white grub populations. This might be due to the fact that the grubs were collected only from a small proportional area of the field, such as from the opened furrow for planting. At that time period, grubs are still not aggregated around crop roots or tubers as there is still no crop. And later, during mechanical weeding and during earthing up of potatoes, only between-row soil was searched. But at this time, grubs are expected feeding on the tubers and roots. Consequently, handpicking likely misses a large part of the pest population; thus, the effect of this method is limited.
Considering factor “time” after planting and/or after EPN treatment the following effects on white grubs were found. Around 30 days after planting, the insecticide and handpicking treatment had no detectable effect on grub numbers (Kruskal-Wallis H test, chi square = 2, p = 0.37; Fig. 4). At 37 days after planting (7 days after EPN treatment), the factor “treatment” had no detectable effect on grub numbers (sqrt-transformed data, Univariate GLM, F7;16 = 0.8, p = 0.6; Fig. 4).
Around 60 days after planting (30 days after EPN treatment), treatments had reduced grub numbers (sqrt-transformed, Univariate GLM, F 7;16 = 3.6, p = 0.045) as well as around 90 days after planting at day 60 after EPN treatment; GLM, F 7;16 = 4, p = 0.034; Fig. 4).
At harvest, 128 days after planting (day 71 to 108 after EPN treatment), no effects of EPNs on grub numbers were detected anymore (sqrt-transformed, Univariate GLM, F 7;16 = 0.7, p = 0.67) (Fig. 4).
EPN efficacy at preventing yield loss
The average yield was 0.3 ± 0.14 kg Irish potato tubers per plant, equalling about 9 to 10 tons per hectare. The yield was double in field M as in field N, that is, 0.4 ± 0.03 kg versus 0.2 ± 0.02 kg per plant, respectively (independent sample t test, t 62 = 11.8, p < 0.0001).
However, treatments had no detectable effect towards an increased yield (GLM of yields compared to control, F 7; 63, = 1.6; p = 0.15).
EPN persistence
The Rwandan or international EPNs were recovered from the soil of the treated experimental plots, but no natural EPN population was detected in the untreated plots.
Both applied EPN species well-established in the soil, because soil samples from the treated plots were all found EPN-positive 7 days later (Fig. 5). Later on, EPN persistence decreased with time (Pearson correlation r = −0.67 with time, p < 0.001, n = 40). Multiple comparison tests at each time step did not reveal differences between the persistence of the two EPNs and their concentrations (too few data), but a slight overall treatment effect remained (e.g. GLM at 30 days, F 4;9 = 5.8, p = 0.041; Fig. 5). S. longicaudum X7 persisted in the soil for at least 60 days after treatment in both fields. The Rwandan Steinernema sp. RW14-M-C2b1 persisted in the soil for at least 60 days in field N, whereas it was only detected until day 30 in field M. No EPNs were anymore detected towards time of harvest in both fields.
In summary, baited soil samples in our study proved that both the Rwandan and international EPN can establish in the soil after treatment. This indicates that the applied EPNs were of good quality as they need energy to survive some time in the soil until finding and propagating in the host insects. It moreover showed that the application method of EPNs into the moist just-opened soil furrow is a method appropriate for EPNs. Applications into the soil are known to be a good practice for EPN use as they prefer permanent moist conditions (Dutky, 1969). Also against other soil pests at a field scale, such as against the chrysomelid larvae of Diabrotica rootworms in maize, the fluid application into the soil had proven most suitable for EPNs compared with onto-soil fluid row-applications or into-soil granule applications (Toepfer et al., 2010). Our results also showed that the applied EPNs persisted in the fields of Rwanda for at least 2 months, which indicates that they were able to propagate. After more than 3 months, i.e. during harvest time, no EPNs were detected anymore. This is a typical situation for crops with large surfaces of bare soil such as maize (Kurtz et al., 2007), or, as here, in wide-spaced senescent Irish potatoes. One season persistence is advantageous in case of commercialisation of the EPNs, as they would need to be more frequently applied. Short persistence is however less typical for vegetables or berries where EPNs persist longer (Burlando and Kaya, 1992), or grasslands or orchards where they can persist for years (Belair et al., 1994).
Conclusions
Most data of this study showed that there was variability within a field, between fields and between times steps. This is common for field trials, particularly when working with natural insect pest populations that are often aggregated. Moreover, the diversity of white grub species as well as their life cycles in Rwandan habitats is largely unclear, leading to different instars of different grub species in the soil. Due to the variability of data, it might be advised to field test the EPNs with higher plot numbers (at least six instead of four) at more locations in different Rwandan provinces and under different farming conditions as well as under different levels of natural white grub populations. This may lead to a broader view on the potential of the new Rwandan Steinernema sp. RW14-M-C2b-1 and the other isolates.
In conclusion, The Rwandan EPN Steinernema sp. RW14-M-C2b-1 is suggested to be used, in the first place, for white grub control in tubers and vegetables of Rwanda. As for the other Rwandan nematodes, more field trials are needed particularly with the heterorhabditids such as H. bacteriophora Rwanda14-N-C4a. It is expected that also these will work, or even better, if applied in the same way as presented here.
What remains is to consider whether indigenous nematodes need to be registered in Rwanda, and whether the EPNs should be mass produced either subsidised through governmental programs or through commercialization. Registration processes for macrobial biocontrol agents, such as EPNs, are currently not in place in Rwanda. The country may either follow the numerous countries that do not require any registration for indigenous EPNs as biocontrol agents, such as Germany, UK, or even not at the European Union level (Akhurst and Smith, 2002), or may require field efficacy trials and a formal registration approval by an expert committee, such as in Tanzania (J. Mwangi, Ministry of Agriculture, Food Security and Cooperatives MAFSC, 2015, pers. comm.) or in Kenya (KEPHIS, 2014). Discussion on setting up a Rwanda expert group for biocontrol agents is currently ongoing (B. Uwumukiza, MINAGRI Rwanda, pers. comm.).
Ultimately, we believe that EPNs, as here investigated, can deliver effective, safe and environmentally benign pest controls for soil-inhabiting pests to farmers.
References
Akhurst R, Smith K (2002) Regulation and safety. In: Gaugler R (ed) Entomopathogenic nematology, pp 311–332
All JN, Saunders MC, Dutcher JD, Javid AM (1981) Susceptibility of grape root borer larvae, Vitacea polistiformis (Lepidoptera: Sesiidae) to Neoplectana carpocapsae (Nematoda: Rhabditida): potential of host kairomones for enhancement of nematode activity in grape vineyards. Miscellaneous Publ Entomol Soc Am 12:9–14
Anonymous (2007) Efficacy evaluation of plant protection products: harmonized classification and coding of the uses of plant protection products: pp 1/248(1). EPPO Bulletin 37:25–28
Ansari MA, Tirry L, Moens M (2003) Entomopathogenic nematodes and their symbiotic bacteria for the biological control of Hoplia philanthus (Coleoptera: Scarabaeidae). Biol Control 28:111–117
Bal HK, An R, Uslu U, Grewal PS (2014) Comparison between TTO1 and GPS11 strains of the entomopathogenic nematode Heterorhabditis bacteriophora. Int J Nematol 24(2):18–28
Bedding RA, Akhurst RJ (1975) A simple technique for the detection of insect parasitic rhabditid nematodes in soil. Nematologica 21:109–110
Belair G, Vincent C, Chouinard G (1994) Persistence of the entomopathogenic fungi Steinernema carpocapsae on apple. Resume des Recherches Centre de Recherche et de Developpement en horticulture, vol 23. Saint Jean sur Richelieu, Quebec, p 3
Burlando TM, Kaya HK (1992) Persistence of entomopathogenic nematodes and reduction of black vine weevil in soil. J Nematol 24:584–585
Caroli L, Glazer I, Gaugler R (1996) Entomopathogenic nematode infectivity assay: comparison of penetration rate into different hosts. Biocontrol Sci Techn 6:227–233
Chen J, Yu X, Chen L, Lv Z, Zheng X, Xu H, Zhang Y (2004) Occurrence, damage of the soil-dwelling pests and its management strategy in China. Zhejiang J Agri 16:389–394
Cock MJW, Gillian BA (2013) Observations on white grubs affecting sugar cane at the juba sugar project, south-western Somalia, in the 1980s, and implications for their management. Insects 4:241–272
R Development Core Team. 2009. R: a language and environment for statistical computing. R foundation for statistical computing, Vienna, . Available online at http://www.r-project.org (Accessed May 2015)
Downing AS (1994) Effect of irrigation and spray volume on efficacy of entomopathogenic nematodds (Rhabditida: Heterorhabditidae) against white grubs (Coleoptera: Scarabaeidae). J Econ Entomol 87(3):643–646
Dutky SR (1969) The use of nematodes for insect control. In: Proceedings of the symposium on tropical nematology. University of Puerto Rico, Rio Piedras, pp 27–32
Furlan L, Canzi S, Bernardo AD, Edwards CR (2006) The ineffectiveness of insecticide seed coatings and planting-time soil insecticides as Diabrotica virgifera virgifera population suppressors. J Appl Entomol 130:485–490
Grewal P, Ehlers RU, Shapiro-Ilan DI (2005) Critical issues and research needs for expanding the use of nematodes in biocontrol. In: Ehlers RU, Shapiro DI (eds) Nematodes as biocontrol agents. Grewal., P. S. CABI Publishing, Wallingford, pp 47–64
Grewal PS (1992) Laboratory techniques for work with entomopathogenic nematodes. In: Recent advances in nematology. Bioved Research Society, Allahabad, pp 51–61
Guo W, Yan X, Zhao G, Chen J, Han R (2015) Efficacy of entomopathogenic Steinernema and Heterorhabditis nematodes against Holotrichia oblita. J Pest Sci 88:359–368
Guo W, Yan X, Zhao G, Han R (2013) Efficacy of entomopathogenic Steinernema and Heterorhabditis nematodes against white grubs (Coleoptera: Scarabaeidae) in peanut fields. J Econ Entomol 106(3):1112–1117
Holmes KA, Chen J, Bollhalder F, Ri U, Waweru B, Li H, Toepfer S (2015) Designing factories for nematode-based biological control products for an alternative, environmentally friendly management of soil insect pests. Afr J Agr Res 10:4432–4448
Kaya HK, Aguillera MM, Alumai A, Choo HY, de la Torre M, Fodor A, Gangul S, Hazar S, Lakatos T, Pye A, Wilson M, Yamanaka S, Yang H, Ehlers RU (2006) Status of entomopathogenic nematodes and their symbiotic bacteria from selected countries or regions of the world. Biol Control 38:134–155
KEPHIS (2014) Guidelines for the introduction and use of bio-products, biological control agents and related products, vol 77. Kephis, Nairobi
Kinnear PR, Gray CD (2000) SPSS for windows made simple. Psychology Press Ltd, East Sussex, p 380
Klein MG, Georgis R (1992) Persistence of control of Japanese beetle (Coleoptera: Scarabaeidae) larvae with steinernematid and heterorhabditid nematodes. J Econ Entomol 85(3):727–730
Koppenhöfer AM, Fuzy EM (2003) Biological and chemical control of the Asiatic garden beetle, Maladera castanea (Coleoptera: Scarabaeidae). J Econ Entomol 96(4):1076–1082
Koppenhöfer AM, Fuzy EM (2009) Long-term effects and persistence of Steinernema scarabaei applied for suppression of Anomala orientalis (Coleoptera: Scarabaeidae). Biol Control 48:63–72
Kurtz B, Toepfer S, Ehlers RU, Kuhlmann U (2007) Assessment of establishment and persistence of entomopathogenic nematodes for biological control of western corn rootworm. J Appl Entomol 131:420–425
Laznik Ž, Trdan S (2015) Failure of entomopathogens to control white grubs (Coleoptera: Scarabaeidae). Acta Agric Scand 65(2):95–108
Li H, Toepfer S, Agwanda C, Rischbieth J, Zhang F, Kajuga J (2016) From China to Rwanda: successful transfer of biocontrol for soil insect pests. CABI Res Brief 2:3
Mayo ZB, Peters LL (1978) Planting vs. cultivation time applications of granular soil insecticides to control larvae of corn rootworms in Nebraska. J Econ Entomol 71:801–803
Musebe R, Dusenge L, Agwanda C, Kajuga J, Waweru B, Karanja D, Hongmei L, Day R (2017) Understanding the need for transfer of biologically-based crop protection technology for soil pest control in vegetable production in Rwanda. African J Agric Res 12:1793–1800
Nyamwasa I, Li K, Yin J, Zhang S, Kajuga J, Hategekimana A, Waweru B, Li H (2017) Occurrence of soil insect pests: insight from classical identification supplemented with DNA barcoding. Int J Pest Sci 63(1):18–29
Peters A, Huneke K, Ehlers RU (1996) Host finding by the entomopathogenic nematode Steinernema feltiae. IOBC Bulletin 19:99–102
Peters A, Smits PH (2000) Insect-based assays for entomopathogenic nematode infectiousness: definitions, guidelines, problems. Bulletin OILB/SROP 23:109–114
Poinar GO Jr (1990) Taxonomy and biology of Steinernematidae and Heterorhabditidae. In: Gaugler R, Kaya HK (eds) Entomopathogenic nematodes in biological control. CRC Press, Boca Raton, pp 23–60
Selvan S, Gaugler R, Campbell JF (1993) Efficacy of entomopathogenic nematode strains against Popillia japonica (Celeoptera: Scarabaeidae) larvae. J Econ Entomol 86(2):353–359
Shapiro-Ilan DI, Gaugler R (2002) Production technology for entomopathogenic nematodes and their bacterial symbionts. J Ind Microbiol Biotechnol 28:137–146
Shen CP, Wang GH (1991) Description and studies of an entomopathogenic nematode: Steinernema Longicaudum sp. nov. Proceeding of the first national academy symposium. Chinese Science and Technology Press, Beijing, pp 220–231
Toepfer S, Hatala-Zseller I, Ehlers RU, Peters A, Kuhlmann U (2010) The effect of application techniques on field-scale efficacy: can the use of entomopathogenic nematodes reduce damage by western corn rootworm larvae? Agr Forest Entomol. 12:389–402
Toepfer S, Kajuga J, Waweru B, Agwanda C, Karanja D, Yan X, Li K, Li H (2016) Rwanda mass-produces nematode-based biocontrol agents for controlling soil insect pests. Biocontrol News and Inform 37(3):17–18
Toepfer S, Li H, Pak SG, Son KM, Ryang YS, Kang SI, Han R, Holmes K (2014) Soil insect pests of cold temperate zones of East Asia, including DPR Korea: a review. J Pest Sci 87:567–595
Trutmann P, Graf W (1993) The impact of pathogens and arthropod on common bean production in Rwanda. Agr Forest Entomol 39(3):328–333
White GF (1927) A method for obtaining infective nematode larvae from cultures. Science 66:302–303
WHO (2009) The WHO recommended classification of pesticides by hazard and guidelines to classification. World Health Organization. IPCS International program on Chemical safety, Geneva, p 60
Yan X, Han R, Moens M, Chen S, De Clercq P (2013) Field evaluation of entomopathogenic nematodes for biological control of striped flea beetle, Phyllotreta striolata (Coleoptera: Chrysomelidae). BioControl 58:247–256
Yan X, Waweru B, Qiu X, Hategekimana A, Kajuga J, Li H, Edgington S, Umulisa C, Han R, Toepfer S (2016) New entomopathogenic nematodes from semi-natural and small-holder farming habitats of Rwanda. Biocontrol Sci Techn 26(6):820–834
Acknowledgements
We like to thank Jean Pierre Bizimana from the Rwanda Agriculture Board (RAB) for help in field work, Bazagwira Didace and Ingabire Geraldine for strain maintenance in the biocontrol agent factory at RAB Rubona station, and Louis Butare (Director General, RAB) for agreeing on the establishment of a biocontrol agent facility and his general support. We like to thank Charles Agwanda (CABI Africa) for his operational support in East Africa. The study was funded as part of a triangular technology transfer project from the UK, Switzerland and China to Rwanda by the AgriTT Research Challenge Funds of the UK Department for International Development (DfID) (AgriTT-RCF-Project 1301) and the GDAS Special Project of Science and Technology Development (2017GDASCX-0107). The compilation of the manuscript was supported by the CABI Development Fund (CDF CR60019C).
Author information
Authors and Affiliations
Contributions
The whole team jointly planned the experiments. JK and AH contributed equally to this work. JK and HL played an additional role in coordinating the study. AH, BW and CU were particularly important for data collections. KL, DK and JY supported the field experiments. LC and XY particularly supported the nematode production, handling and application. BW and ST analysed the data. XY and ST wrote the manuscript with help of co-authors and are both corresponding authors. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
Joelle Kajuga, Athanase Hategekimana, Bancy Waweru, Christine Umulisa from the Rwanda Agriculture Board and their farmer groups are representatives of the beneficiaries. The authors from the Rwanda Agriculture Board (RAB) agreed that the Rwandan EPN isolates can be maintained and researched outside Rwanda. The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Kajuga, J., Hategekimana, A., Yan, X. et al. Management of white grubs (Coleoptera: Scarabeidae) with entomopathogenic nematodes in Rwanda. Egypt J Biol Pest Control 28, 2 (2018). https://doi.org/10.1186/s41938-017-0003-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41938-017-0003-2