
Abstract
Background
Sesame (Sesamum indicum L.) is a widely used oil seed crop throughout the world but its productivity is extremely low due to use of low yielding cultivars as well as biotic stress for the major pest Spilosoma obliqua Walker. But even today, farmers generally use broad-spectrum synthetic pesticides for their management injudiciously without considering any economic threshold (ET) limit and creates ecosystem crisis. So, time-based ecologically sustainable management of the said pest and selection of a superior cultivar of sesame was studied by stage-specific two-sex pooled life table and nutritional ecology of S. obliqua on three sesame cultivars (Rama, Shubhra and Amrit) along with their economic thresholds (ETs) in 2019.
Results
The nutritional ecology and population dynamics of S. obliqua were significantly affected by the host phytoconstituents in terms of host suitability or susceptibility (Rama > Shubhra > Amrit). The mean EIL and ETL for S obliqua was 36.316 ± 3.911 and 33.243 ± 2.734 pests/m2, respectively on cv. Rama that were significantly (F2,6 = 5.421–5.435; P ≤ 0.042) lower than Shubhra and Amrit. For a single pest per m2 (30 ± 2 plants/m2) the possible time that can be taken to reach EIL (Ti) and ETL (Tt) were 39.132 ± 3.969 and 38.132 ± 3.969 days, respectively on cv. Rama which were also significantly (F2,6 = 26.551; P = 0.001) lower than the other cultivars. The seed yield and benefit cost ratio (BCR) were 857.099 ± 0.000 (Kg/ha) and 0.607 ± 0.000, respectively for cv. Rama that were significantly (P < 0.05) lower than the others.
Conclusions
It will enable growers to find the most preferred cultivar (Rama > Shubhra > Amrit) based on BCR values irrespective of their biotic resistance (Rama < Shubhra < Amrit) due to host antibiosis. Even, the ETs-based time series for judicious management of the pest along with carbon sequestration efficiency (CSE) will also support superiority of the cultivars (Rama > Shubhra > Amrit) towards climate smart pest management (CSPM) of sesame and or other such crops in near future.
Similar content being viewed by others

Background
Sesame (Sesamum indicum L., Family: Pedaliaceae, 2n = 26) is a widely used oil seed crop throughout the world and cultivated mainly in tropics and subtropics for its edible seeds with highest oil content (Chongdar et al., 2015; Dar et al., 2014). India is the world’s largest producer and exporter of sesame in the world (Kale et al., 2018; Naik et al., 2016). In West Bengal several cultivars (Rama, Savitri, Tillotama, Nirmala, Shubhra, Amrit, Prachi, Kalika, Vinayaka, Uma, JLT-408, etc.) of sesame is cultivated (Chongdar et al., 2015; Dar et al., 2014; Kale et al., 2018). But its productivity is extremely low due to several yield limiting biotic and abiotic factors including use of low yielding varieties (Chongdar et al., 2015; Kale et al., 2018; Naik et al., 2016). Among the biotic stresses insect pests are the major constraints and limiting yield potential of sesame like other economic crops throughout the world (Biswas, 2006; Nath, 1975). Bihar hairy caterpillar (BHC) of Spilosoma obliqua Walker (Syn. Diacrisia obliqua) (Lepidoptera: Arctiidae) is considered as a generalist pest on several crops including sesame in India and elsewhere (Bhadauria et al., 2001; Biswas, 2006; Gotyal et al., 2015; Singh & Varatharajan, 1999; Varatharajan et al., 1998). The larvae feed voraciously and act as a serious pest of sunflower (Varatharajan et al., 1998), sesame (Biswas, 2006), jute (Gotyal et al., 2015), black gram (Mandal et al., 2013), green gram (Mobarak et al., 2019). So, timely management of this notorious pest is very important as delay may lead to complete defoliation of crop and havoc economic loss (Gotyal et al., 2015). But unfortunately, even today, farmers generally use broad-spectrum synthetic pesticides and some biorationals for their management injudiciously for even a single pest observation without considering any economic threshold (ET) limit or irrespective of pest population growth rate (Carvalho, 2017; Damalas & Koutroubas, 2018; Gotyal et al., 2015; Mohapatra & Gupta, 2018; Nagiaet al., 1990; Parui & Roy, 2016). These result into secondary pest outbreak, pest resurgence, development of pesticide resistance and emergence of new pest biotypes, which ultimately leads regulatory complications in the agro ecosystem (Carvalho, 2017; Kang, 2019; Kim et al., 2017; Ndakidemi et al., 2016). To face this ecosystem crisis, population dynamics, nutritional ecology and ET-based time specific sustainable management are very crucial (Carey, 1993; Chen et al., 2017; Dutta & Roy, 2016; Heeb et al., 2019; Mobarak et al., 2019; Roy, 2015a, 2015b, 2017, 2019; Southwood, 1978; Subedi et al., 2019). There is another alternate safe strategy for limiting herbivore by the selection of high yielding resistant varieties (Wolfenberg & Phifer, 2000; Dar et al., 2014; Mobarak et al., 2019). But the selection of such resistant varieties depends on their resistance mechanisms (antixenosis, antibiosis and tolerance) against pests (Golizadeh & Razmjou, 2010). Among these mechanisms antibiosis is the most important and depends on host phytoconstituents which actually play a vital role in pest feeding preference and population dynamics by affecting immatures as well as adult performance (Applebaum, 1985; Awmack & Leather, 2002; Dicke, 2000; Roy, 2019; Roy & Barik, 2012, 2013; Schoonhoven et al., 2005; Shobana et al., 2010; Slansky & Scriber, 1985).Thus, till date host photochemical mediated pest population ecology for crop cultivar selection are exceptionally rear (Golizadeh & Razmjou, 2010; Karimi-Pormehr et al., 2018; Mobarak et al., 2019; Naseri et al., 2014). But, till date none of the studies has been performed with S. obliqua on different sesame cultivars using life table and nutritional ecology-based economic threshold determination. Thus, my objectives of the present study were to (i) find the phytochemical basis of resistance in selected sesame cultivars against S. obliqua by their food utilization efficiency measures, (ii) determine the influence ofthe cultivars on population parameters of S. obliqua to suggest suitability or susceptibility of cultivars including their economic profit, (iii)determine the appropriate ETs and respective time series by using pest density from the field, economic attributes beyond the field and their population parameters on the sesame cultivars along with their carbon sequestration efficiencies (CSE)for climate smart pest management (CSPM).
Methods
Series of field and laboratory experiments were conducted during 2019 to study the feeding dynamics and population ecology-based economic threshold (ET) calculation of S. obliqua on the three cultivars (Rama, Shubhra and Amrit) of sesame.
Host plants
Three well-known Sesame (Sesamum indicum L) cultivars, i.e., Rama, Shubhra and Amrit (Chongdar et al., 2015; Dar et al., 2014; Kale et al., 2018) were cultivated and collected from a selected field situated near Chinsurah Rice Research Center (CRRC), Chinsurah, 22°53' N, 88°23' E, 13 m above sea level, Hooghly, West Bengal, India, during summer season (February to June) in 2019. The plots [each plot 5 m × 5 m; plot gap 0.5 m, soil organic matter 5.3 ± 0.2%, pH 7.7, photoperiod 13 L:11 D at 30–35 °C] were prepared for cultivation of three sesame cultivars (Rama, Shubhra and Amrit) with three replications including control (without any insecticide) side by side near CRRC. A space of 1 m was kept for cultivation of another sesame cultivar and they were naturally infected by S. obliqua during early May in the field and the pests were collected separately for their mass culture. For insect free plants, each sesame cultivar was also grown by pot (16 cm upper radius × 8 cm lower radius × 24 cm height, 2500 cm3 soil) culture protected by insect net and was grown in natural condition as in the field. Mature leaves of 4–5 week old plants from each cultivar was collected separately from the pot cultured plants for phytochemical analysis as well as provided as food for S. obliqua neonates. The plants were also identified and voucher specimens (Voucher No. ERU21-23) were kept in Department of Zoology, Ecology Research Unit, M.U.C. Women’s College, Burdwan, West Bengal, India.
Insect mass culture
The initial populations of S obliqua larvae were collected from each sesame cultivar separately from the same cultivated fields. The larvae were incubated in the laboratory at 26 ± 1 °C, 60 ± 5% RH and a photoperiodism of 12:12 (L:D) on intact mature leaves of the selected cultivars separately in glass jars (20 cm dia. × 30 cm ht.) until their pupation. After emergence of adults from the reared pupa 6 pairs of newly emerged male and female was placed in an oviposition case (50 cm dia. × 40 cm ht.) of fine nylon net containing the same mature leaves of each cultivar for their oviposition to obtain same aged eggs of S obliqua as previously described (Roy, 2017, 2018, 2019; Roy & Barik, 2012, 2013, 2014). On each cultivar, newly laid eggs by the F3females were collected in order to obtain the same aged eggs of defined cohort (n = 100) for the selected cultivars (Rama, Shubhra and Amrit) with three replications at same conditions in a growth chamber [ten eggs in a glass jar (20 cm dia. × 30 cm ht.)] for three generations to study the pest population ecology as described previously (Roy, 2017, 2018, 2019). The feeding dynamics of the neonates and population data throughout their life cycle were recorded separately on the selected sesame cultivars as in my previous studies (Roy, 2015a, 2015b, 2019).
Phytochemical analysis
The sesame leaves of the selected cultivars (Rama, Shubhra and Amrit) were freshly collected from the pot cultured plants. The leaves were initially rinsed with distilled water and dried by paper towelling separately for phytochemical analysis. They were extracted in different solvents for primary and secondary metabolitesas well as estimated by various standard protocols (Harborne, 1973, 1994) as in Roy and Barik (2012, 2013) as well as Roy (2015a, 2015b, 2017, 2019). Determination of each biochemical analysis was repeated for three times and expressed in dry or fresh weight basis accordingly.
Feeding dynamics
Feeding ecology was conducted by taking the F4 newly emerged first instar larvae in the laboratory condition up to three generations (F4–F6) on respective selected sesame cultivars(Rama, Shubhra and Amrit) separately as described in previous experiments (Roy, 2017, 2019; Roy & Barik, 2013). Larvae were placed in a glass jar (20 cm dia. × 30 cm ht.) containing leaves of a particular sesame cultivar. The larvae were weighed initially and allowed to feed on the weighed quantity of leaves from each cultivar for 24 h interval and were reweighed for determination of different conversion factors along with feeding indices. Food utilization indices were calculated by the formulae of Waldbauer (1968) with slight modifications (Roy & Barik, 2013; Roy, 2015a, 2015b, 2017) to assess the feeding efficiencies of the Bihar hairy caterpillar (BHC) of S obliqua at 26 ± 1 °C, 60 ± 5% RH and a photoperiodism of 12:12 (L:D) hours in a growth chamber as described previously in other cases (Roy, 2017, 2019). All the feeding indices like, growth rate (GR), consumption rate (CR), relative growth rate (RGR), consumption index (CI), egestion rate (ER), host consumption rate (HCR), approximate digestibility (AD%), efficiency of conversion of ingested food (ECI%), efficiency of conversion of digested food (ECD%) and host utilization efficiency (HUE%) including feeding index (FI), growth index (GI) and pest susceptibility index (PSI) were estimated (Additional file 1: Table S1).
Life table study
The data on survival, developmental duration and oviposition of all individuals on the selected three sesame cultivars (Rama, Shubhra and Amrit) were analyzed separately based on age-stage, two-sex life table (Chen et al., 2017; Chi & Su, 2006; Roy, 2017, 2019). It includes several parameters, which were calculated with the formulae of Southwood (1978), Carey (1993, 2001), Krebs (1994), Price (1998) and Kakde et al. (2014). These parameters include probability of survival from birth to age x (lx), proportion dying each age (dx), mortality (qx), survival rate (sx) per day per age class from egg to adult stages (Additional file 1: Table S2). Using these parameters, the following statistics like total individuals at age x and beyond k (Tx), average population alive in each stage (Lx), life expectancy (ex), exponential mortality or killing power (kx), total generation mortality (K or GM), generation survival (GS), gross reproductive rate (GRR), net reproductive rate (NRR or R0), mean generation time (Tc), doubling time (DT), intrinsic rate of population increase (rm), Euler’s corrected r (rc), finite rate of population increase (λ), weekly multiplication rate (λ7), increase rate per generation (λTc), were also computed, using Carey’s formulae (1993). Some other population parameters like potential fecundity (Pf), total fertility rate (Fx), mortality coefficient (MC), population growth rate (PGR), population momentum factor of increase (PMF), expected population size in 2nd generation (PF2), Hypothetical females in 2nd generation (HFF2), expected females in 2nd generation (FF2), general fertility rate (GFR), crude birth rate (CBR), reproductive value (RV), vital index (VI) and trend index (TI) were also determined by using well defined formulae (Brich, 1948, Southwood, 1978, Carey, 1993, Roy, 2019) (Additional file 1: Table S2).
Field experiment
A field experiment was conducted in 2019by growing selected three sesame cultivars(Rama, Shubhra and Amrit) in randomized block design (RBD) to determine the population dynamics and nutritional ecology-based economic threshold (ET) of BHCs of S obliqua as described earlier workers with few modifications (Naik et al., 2016; Parui & Roy, 2016; Roy, 2019). The experiment was done by using a small land area (10 katha or 670 m2) near CRRC with 3 replications for both control and treated plots (5 m × 5 m) as described above with average plant density of 30 ± 2 plants/m2. The data from the same sesame field was collected for determination of ETs of BHC of S obliqua on the cultivars (Additional file 1: Fig. A, B). The yield potential of the selected cultivars were observed over a traditional synthetic pesticide, Triazophos 40 EC (@ 40gm a.i/ha), along with control (without pesticide) side-by-side (Mohapatra & Gupta, 2018; Parui & Roy, 2016).
Yield loss and ET calculation
From seed showing to harvest of the selected sesame cultivars (Rama, Shubhra and Amrit),the occurrences of BHC(s) of S obliqua were recorded by random quadrat sampling (RQS) from each treated and control plots (Parui & Roy, 2016)[47]. Calculation of economic injury level (EIL) for S obliqua according to the methodology proposed by Pedigo et al. (1986) expressed as numbers or injury equivalents (Additional file 1: Table S3). The economic threshold (ET) is the population density at which control action should be determined (initiated) to prevent an increasing pest population (injury) from reaching the EIL (Pedigo & Buntin, 1994; Pedigo & Higley, 1992). On the basis of BHC infestation and the efficacy of the traditional synthetic pesticide were determined in terms of yield damage reduction (%), proportion of insect controlled (%) and percent yield loss per pest per plant (%) along with the management costs (CC) for the calculation of EIL, ETL, time to reach the EIL (Ti) and ETL (Tt) when a plant was infested by a single pest in the field (Higley & Wintersteen, 1992; Pedigo & Higley, 1992; Roy, 2019). A time series was also calculated up to reach the ETL from population growth data. The benefit cost ratio (BCR) was also determined (Chongdar et al., 2015) to find the seed production efficiency as well as resistance of the selected cultivars against S obliqua as the sole pest infestation (Additional file 1: Table S3). The organic biomass production and carbon sequestration efficacy (CSE) of the selected cultivars were also determined (Additional file 1: Table S3) for climate smart agriculture (CSA) to mitigate the climate change (Albrecht & Kandji, 2003; Aryal et al., 2018; Chhetri et al., 2017; Heeb et al., 2019; Kang, 2019; Lal, 2008, 2011; Wang, et al., 2016).
Statistical analysis
Experimental data of different phytoconstituents of the selected sesame cultivars (Rama, Shubhra and Amrit) and the pest (S obliqua) population parameters along with their feeding indices were subjected to one-way Analysis of Variance (ANOVA), regression analysis and correlation analysis (Parui & Roy, 2016; Roy, 2017, 2018, 2019; Zar, 1999).The RQS data of the selected cultivars from the field with ETL values of the pest were also analyzed using one-way ANOVA (Roy, 2019; Zar, 1999). Means of different phytochemicals of the cultivars, demographic parameters and different feeding indices of the pest along with ETL related values were compared by Tukey’s test (HSD) when significant values were obtained (Roy, 2017; Zar, 1999). All the statistical analysis was performed by using SPSS, version 16.0 (Roy, 2017, 2018, 2019).
Results
Host phytochemicals
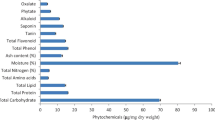
The biochemical constituents of the selected sesame cultivars (Rama, Shubhra and Amrit) were presented in Fig. 1a, b. The primary metabolites were varied significantly (F2,6 ≥ 7.962; P < 0.05) in the selected sesame cultivars (Rama > Shubhra > Amrit) whereas the secondary metabolites were reverse in the cultivars (Rama < Shubhra < Amrit) also with significant (F2,6 ≥ 8.432; P < 0.05) variations (Fig. 1[a]). Total carbohydrate and protein contents were 54.320 ± 0.443, 50.677 ± 0.422, 49.130 ± 0.321, and 10.520 ± 0.434, 6.877 ± 0.312, 5.330 ± 0.348ug/mg dry weight, respectively, in the selected cultivars (Fig. 1[a]). The lipid content was9.120 ± 0.335, 5.477 ± 0.367, 4.237 ± 0.321 ug/mg dry weight, respectively, in the cultivars (Fig. 1a). Total free fatty acids and amino acids were 29.260 ± 0.452, 25.617 ± 0.432, 24.070 ± 0.432, and 8.950 ± 0.387, 5.307 ± 0.365, 3.760 ± 0.343 mg/100 g fresh weight, respectively, in the selected sesame cultivars (Rama > Shubhra > Amrit) with significant (F2,6 ≥ 8.672; P < 0.05) variations (Fig. 1b). Among the secondary metabolites total phenol and phytate content were10.500 ± 0.423, 14.157 ± 0.457, 15.397 ± 0.412 and 3.140 ± 0.323, 7.997 ± 0.347, 9.337 ± 0.312ug/mg dry weight, respectively (Fig. 1a). The other secondary metabolites were also present similarly (Rama < Shubhra < Amrit) in the selected cultivars (Fig. 1a). Ultimately, the ratio of primary to secondary metabolites was significantly (F2,6 ≥ 9.452; P < 0.05) higher in Rama cultivar followed by Shubhra and Amrit (Rama > Shubhra > Amrit).

Phytochemical variations (Mean ± SE of 3 observations/cultivar) in a (μg/mg dry weight) and b (mg/100g fresh weight) of the selected sesame (S. indicum) cultivars (Rama, Shubhra and Amrit) cultivated during summer season in 2019. All the estimated chemicals significantly differed at P < 0.005 by Tukey’s (HSD) test
Feeding dynamics
The food utilization indices was calculated on the selected sesame cultivars (Rama, Shubhra and Amrit) only for the larvae (I-VI instars) of S obliqua as they solely responsible for defoliation (Additional file 1: fig. A, B) and which ultimately lead to the variation in total life history as well as population parameters. The average food utilization efficiency of the instars (I-VI) displayed variations with different pattern of significance (F5,17 ≥ 13,289.732; P < 0.0001) on the selected cultivars (Table 1). The average GI (%) was 3.885 ± 0.031, 3.744 ± 0.063 and2.176 ± 0.114, respectively, for the cultivars (Rama > Shubhra > Amrit) and was varied significantly (F2,6 = 10.017; P = 0.012) (Table 1). Whereas, the average PSI (%) was 50.451 ± 3.871, 62.838 ± 2.904 and 75.226 ± 1.936, respectively, for the cultivars (Rama < Shubhra < Amrit) and was also varied significantly (F2,6 = 16.946; P = 0.003) (Table 1). Thus, the feeding indices represent different biotic resistance (Rama < Shubhra < Amrit) and or susceptibility of the cultivars (Rama > Shubhra > Amrit) to the defoliator (S obliqua) due to their respective phytoconstituents (Fig. 1a, b).
Population dynamics
The stage-specific two-sex pooled life table of S obliqua were investigated in the laboratory with three replications on mature leaves of the selected sesame cultivars (Rama, Shubhra and Amrit) during 2019 and showed four distinct stages (i.e., egg, larva, pupa and adult) with six larval instars (Tables 2, 3, 4, 5 and 6). The cohorts (3 × 3 = 9) containing 100 eggs each on the three sesame cultivars (Rama, Shubhra and Amrit) were reared separately to determine their different population parameters. The demographic data of S obliqua represent similar pattern of development with significant variations (P < 0.05) in different developmental stages on the selected sesame cultivars (Tables 2, 3, 4, 5 and 6). The population parameters like, lx, Lx, Tx and ex of S obliqua were gradually decreased throughout their developmental stages with significant (P < 0.05) variations on the selected cultivars in the order of Rama > Shubhra > Amrit (Tables 2, 3 and 4) and they always produce type-III survivorship curve like most of the insects. Whereas, the dx, qx and kx were varies in different developmental stages with significant (P < 0.05) variations and comparatively higher in egg and pupal stage with a rapid surge during adult stage on the selected cultivars in the order of Rama < Shubhra < Amrit (Tables 2, 3 and 4). ANOVA results of the life table parameters on the selected cultivars were showed more or less same pattern (Rama > Shubhra > Amrit) with significant variations (F7,16 = 10.787–6580.214; P < 0.0001) (Table 5).
The average Pf were 320.667 ± 9.528, 252.625 ± 8.521and 168.542 ± 9.436 eggs/female, respectively (Table 6) for the cultivars (Rama > Shubhra > Amrit) with significant variations (F2,6 = 63.836; P < 0.001). The Fx, GRR or mx and NRR or R0 of S obliqua were also significantly (F2,6 = 12.293–37.710; P ≤ 0.008) differed (Table 6) on the cultivars (Rama > Shubhra > Amrit). Average Tc for the cultivars (Rama, Shubhra and Amrit) were 44.862 ± 0.071, 44.047 ± 0.480and 44.638 ± 0.147 days, respectively (Table 6) without any significant variations (F2,6 = 2.069; P = 0.207). Whereas, average DT were 7.845 ± 0.135, 9.501 ± 0.540 and 20.161 ± 0.587 days, respectively (Table 6) for the respective cultivars (Rama < Shubhra < Amrit) with significant variations (F2,6 = 12.316; P = 0.007). The rm and λ were 0.088 ± 0.002, 0.073 ± 0.004 and 0.044 ± 0.012and 1.092 ± 0.002, 1.076 ± 0.004 and 1.045 ± 0.013 individuals/day, respectively (Table 6) for the cultivars (Rama > Shubhra > Amrit) with significant variations (F2,6 = 9.084–9.424; P ≤ 0.015).The average GM, MC, GS, PGR, PMF, GFR, CBR, RV, VI and TI of S obliqua were also significantly (F2,6 = 5.451–37.168; P ≤ 0.045) differed on the respective cultivars (Table 6). Thus, all the 25 selected population parameters of the cohorts were showed significant differences (P < 0.05) with few deviations (Table 6) on the selected sesame cultivars. So the population growth and reproductive parameters of S obliqua were significantly affected by the cultivars in respect to their phytoconstituents (Fig. 1a, b) and support the host superiority or susceptibility (Rama > Shubhra > Amrit) to the notorious pest.
Yield loss and ETs
Yield loss and ET calculation were conducted during summer season (February to June) in 2019 on the three selected cultivars (Rama, Shubhra and Amrit) infested by BHCs of S obliqua against a well-known traditional synthetic pesticide (Triazophos 40 EC) in the field condition (Table 7).The damage per pest (S obliqua) per plant (D%) were 4.446 ± 0.215, 4.143 ± 0.298 and 3.845 ± 0.181, respectively on the cultivars (Rama > Shubhra > Amrit) without significant(F2,6 = 1.618; P = 0.274) variations (Table 7).The pest control efficacy of the synthetic pesticide (Triazophos 40 EC)over the control represent mean EIL and ETL of 36.316 ± 3.911, 44.319 ± 3.372, 62.853 ± 6.225 and 33.243 ± 2.734, 42.423 ± 3.971, 58.404 ± 5.077 pests per 30 plants (or per m2 area), respectively on the selected cultivars (Rama < Shubhra < Amrit) and was significantly (F2,6 = 5.421–5.435; P ≤ 0.042) differed (Table 7). For a single pest observation per m2 (30 ± 2 plants/m2) area the possible time that can be taken to reach EIL (Ti) and ETL (Tt) were calculated as 39.132 ± 3.969, 55.231 ± 4.214, 85.513 ± 5.399 and 38.132 ± 3.969, 54.231 ± 4.214, 84.513 ± 5.399 days, respectively on the selected cultivars (Rama < Shubhra < Amrit) and was also significantly (F2,6 = 26.551; P = 0.001) differed (Table 7). Even, ET-based time series was also calculated to find the specific time (Tt days) to reach ETL for any number of pest(s)/m2 (30 ± 2 plants/m2) on the selected cultivars (Table 8). The maximum tolerance levels of the pests were 30.400, 45.500 and 54.200 per m2, respectively on the cultivars (Table 8). Seed yield [SY] for the selected cultivars (Rama > Shubhra > Amrit) were 857.099 ± 0.000, 695.269 ± 0.000 and617.351 ± 0.000 kg/ha for average production cost [C] of Rs. 22,400.00 and was significantly (P < 0.05) differed (Table 7).The benefit cost ratio (BCR/ha) were 0.607 ± 0.000, 0.304 ± 0.000and 0.158 ± 0.000, respectively for the selected cultivars(Rama > Shubhra > Amrit) which were also significantly (P < 0.05) differed (Table 7).Thus, the yield loss and ET calculation also represent similar biotic resistance (Rama < Shubhra < Amrit) and or susceptibility (Rama > Shubhra > Amrit) of the cultivars as in feeding as well as population ecology of S. obliqua due to variation in their respective phytoconstituents (Fig. 1[a,b]). The carbon sequestration efficiency (CSE) were 1397.088, 1111.320 and 762.048 lbs/ha, respectively for the selected cultivars (Rama > Shubhra > Amrit) with significant (P < 0.05) variations due to different biomass production (Table 7). Thus, this study will also support the choice of sesame cultivar by considering their BCR values irrespective of biotic resistance against S. obliqua with time-based judicious management and climate smart pest management (CSPM) strategy under the arena of climate smart agriculture (CSA) for sustainable production of sesame.
Discussion
Modern agriculture includes CSPM as a part of integrated pest management (IPM) by using ETs for ecofriendly and sustainable CSA towards more production of crops (Anuga et al., 2019; Aryal et al., 2018; Chávez et al., 2018; Pedigo & Higley, 1992; Pedigo et al., 1986; Roy, 2019). The basic information on the biology of an insect pest is necessary before deciding any strategy to combat with the pest (Chen et al., 2017; Chi & Su, 2006). Pest nutritional ecology and population dynamics are regulated by host phytoconstituents and which are wide-ranging as well as highly dynamic in nature (Awmack & Leather, 2002; Roy, 2019; Roy & Barik, 2013; Shobana et al., 2010). The primary metabolites (carbohydrates, proteins, lipids, amino acids including moisture content) are used for general vitality, growth and reproduction (Dadd, 1985; Harborne, 1994; Schoonhoven et al., 2005; Shobana et al., 2010; Slansky & Scriber, 1985). Host secondary metabolites have defensive role and either produced constitutively or in response to plant damage, and they affect feeding, growth, and survival of any herbivores (Harborne, 1994; Roy & Barik, 2012, 2013; Roy, 2015a; Schoonhoven et al., 2005; War et al., 2012). Moreover, host plant utilization is also influenced by the ability of insect to ingest, assimilate and convert food into their body tissues (Dadd, 1985; Nation, 2001; Scriber & Slansky, 1981). Thus, host plant quality is the key determinant in pest growth, development, longevity, fecundity, fertility and survivability which ultimately indicate suitability of a host plant (Roy & Barik, 2012, 2013; Roy, 2015a). Even different cultivars of a crop can vary in the context of various physiological, morphological and biochemical characteristics as well as can influence the bionomics of an insect pest (Golizadeh & Razmjou, 2010; Sarfraz et al., 2007).
In ecological research, life table study is a central theme and widely useful technique in insect pest management, where developmental stages are discrete and mortality rates vary widely from one life stage to another (Carey, 2001; Dutta & Roy, 2016; Kakde et al., 2014; Roy, 2019; Sarfraz et al., 2007; Southwood, 1978). It is a powerful tool for analyzing and understanding the effect of different hosts or cultivars of a crop on insect pest population (Karimi-Pormehr et al., 2018; Liu et al., 2004; Roy & Barik, 2012, 2013; Roy, 2015a; Win et al., 2011). Moreover, the age-stage, two-sex life table is equally advantageous and eliminate many of the inherent errors of traditional life table due to sexual biasness (Chen et al., 2017; Mobarak et al., 2019). The effect of different food sources on population parameters were observed in Helicoverpa armigera (Liu et al., 2004), Plutella xylostella (Sarfraz et al., 2007), Spodoptera litura (Xue et al., 2010), Papilio polytes (Shobana et al., 2010), Podontia quatuordecimpunctata (Roy, 2015a), Epilachna vigintioctopunctata (Roy, 2017), Scripophaga incertulus (Dutta & Roy, 2018) and Diacrisia casignetum (Roy, 2019) on different host plants. In my study the biochemical analyses of the selected sesame cultivars (Rama, Shubhra and Amrit) revealed that the Rama cultivar were of good nutritional quality compared to other two cultivars because nutritional factors (primary metabolites) were higher compared to others (Shubhra and Amrit), while anti-nutritional factors (secondary metabolites)were in reverse order (Rama < Shubhra < Amrit). This study demonstrated that the sesame cultivars (Rama, Shubhra and Amrit) were significantly influenced the feeding indices and population ecology of S. obliqua due to phytochemical variations in the cultivars as in other instances (Awmack & Leather, 2002; Roy & Barik, 2012, 2013; Roy, 2015a). The S. obliqua larvae fed on Rama cultivar leaves were able to converting the leaf tissues in their body masses more efficiently than on the other (Shubhra and Amrit) cultivars due low metabolic maintenance cost (Roy & Barik, 2012, 2013; Slansky & Scriber, 1985; Xue et al., 2010). All the feeding indices were also affected by the respective host phytoconstituents and were showed some variations due to homeostatic adjustment of consumption rates and efficiency parameters to achieve ideal growth rate even with foods of different quality (Awmack & Leather, 2002; Roy & Barik, 2013; Xue et al., 2010). Pest nutritional ecology, demographic parameters and their yield reduction efficiency generally informs about the time-based infestation capability and density of the pest in the crop ecosystem (Kakde et al., 2014; Mobarak et al., 2019; Roy, 2017). The overall survival rate of S. obliqua on Rama cultivar leaves was highest as compared with Shubhra and Amrit leaves and the result suggest type III survival curve like most insect pests (Mobarak et al., 2019; Price, 1998; Roy, 2017, 2019; Schowalter, 2006). The R0, rm and Tc are fundamental ecological parameters to predict the pest population growth to evaluate the performance of an insect on different host plants as well as the host plant's resistance (Liu et al., 2004; Naseri et al., 2014; Southwood & Henderson, 2000; Win et al., 2011). Further, rm is influenced by several factors like development time, survivorship and fecundity rate of an insect which states the physiological status of an insect in relation to its capacity to increase (Southwood & Henderson, 2000). Thus the variations in life table parameters were due to feeding by the larvae of S. obliqua on different sesame cultivars (Rama, Shubhra and Amrit) with different amount of primary and secondary metabolites (Roy, 2017, 2019; Roy & Barik, 2013). In the present study, Tc and DT of S. obliqua were significantly shorter on Rama cultivar leaves compared to Shubhra and Amrit leaves.
To estimate the exact ETs, it is necessary to calculate population data for both the predator, prey species and relate this to different environmental factors (Higley & Wintersteen, 1992; Pedigo & Higley, 1992). The percent yield loss always increased with increase in larval density and host susceptibility (Pedigo & Higley, 1992). The EIL and ETL were calculated by linear regression model (y = ax + c) based on yield loss, degree of pest infestation, cost of protection and market price of the crop (Pedigo & Higley, 1992). A low EIL and ETL of S. obliqua was found in sesame than green gram due to high damage potential of the pest on their respective preferred host plants as in other findings (Pedigo & Higley, 1992; Roy, 2019). In the present investigation, the mean EIL and ETL for S obliqua was 36.316 ± 3.911 and 33.243 ± 2.734 pests/m2, respectively on cv. Rama that were significantly (F2,6 = 5.421–5.435; P ≤ 0.042) lower than Shubhra and Amrit cultivars (Rama < Shubhra < Amrit). Thus these variations in food utilization indices of S. obliqua ultimately revealed that food quality of the selected sesame cultivars (Rama > Shubhra > Amrit) were influenced their nutritional ecology, population parameters and ETs which were ultimately suggest reverse susceptibility or resistance of the cultivars (Rama < Shubhra < Amrit) accordingly. Even, ETs-based time series for judicious application of any sustainable control measures against this pest and carbon sequestration efficiency (CSE) of the sesame cultivars will obviously reduce ecological imbalance to promote CSA as well as CSPM of such crops in near future.
Conclusions
There is a worldwide growing awareness for promoting environmentally benign and ecosystem service (ESS)-based CSPM practices for CSA. Even ecological engineering (EE) by tailoring ESS manipulation is crucial for better production of any crop. These approaches would bring down the pest load below ETL by judicial use of any control measures including broad-spectrum pesticides for sustainable agriculture. In respect to the phytochemical regime, Rama cultivar had the lowest antibiosis resistance than Shubhra and Amrit against S. obliqua as indicated by the short developmental time and high survival of immature stages. The nutritional ecology and population parameters of S. obliqua in relation with respective host chemical regime will enable growers to find the most preferred cultivar (Rama > Shubhra > Amrit) based on CBR values irrespective of their biotic resistance (Rama < Shubhra < Amrit) due to host antibiosis. Even, the calculated ETs and time series for specific time-based judicious management of the pest along with CSE will also support superiority of the cultivars (Rama > Shubhra > Amrit) towards CSPM strategy under the arena of CSA for sustainable production of sesame and or other such crops in near future.
Availability of data and materials
All the data and materials presented in the manuscript are the original work of the authors.
Abbreviations
- ETs:
-
Economic thresholds
- ETL:
-
Economic thresholds level
- EIL:
-
Economic injury level
- BCR:
-
Benefit cost ratio
- CSE:
-
Carbon sequestration efficiency
- CSPM:
-
Climate smart pest management
- CSA:
-
Climate smart agriculture
References
Albrecht, A., & Kandji, S. T. (2003). Carbon sequestration in tropical agroforestry systems. Agriculture, Ecosystems and Environment, 99, 15–27.
Anuga, S. W., Gordon, C., Boon, E., & Surugu, J. M. I. (2019). Determinants of climate smart agriculture (CSA) adoption among small holder food crop farmers in the techiman municipality, Ghana. Ghana Journal of Geography, 11(1), 124–139. https://doi.org/10.4314/gjg.v11i1.8
Applebaum, S. W. (1985). Biochemistry of digestion, In: G. A. Kerkot, L. I. Gillbert (Eds.) Comprehensiv insect physiology, biochemistry and pharmacology, Pergamon Press, New York, Oxford, pp. 279–311. https://doi.org/10.1016/b978-0-08-030805-0.50013-4
Aryal, J. P., Jat, M. L., Sapkota, T. B., Chhetri, A. K., Kassie, M., Rahut, D. B., & Maharjan, S. (2018). Adoption of multiple climate smart agricultural practices in the Gangetic plains of Bihar, India. International Journal of Climate Change Strategies and Management, 10(3), 407–427. https://doi.org/10.1108/IJCCSM-02-2017-0025
Awmack, C. S., & Leather, S. R. (2002). Host plant quality and fecundity in herbivorous insects. Annual Review of Entomology, 47, 817–844. https://doi.org/10.1146/annurev.ento.47.091201.145300
Bhadauria, N. K. S., Bhadauria, N. S., & Jakhmola, S. S. (2001). Larval development and survival of Bihar hairy caterpillar, Spilosoma obliqua (Walk.) on different host plants. Indian Journal of Entomology, 63, 475–477.
Birch, L. C. (1948). The intrinsic rate of natural increase of an insect population. Journal of Animal Ecology, 17, 15–26.
Biswas, G. C. (2006). Incidence and management of hairy caterpillar (Spilarctia obliqua Walker) on sesame. Journal of Agriculture & Rural Development, 4, 95–100.
Carey, J. R. (1993). Applied demography for biologists with special emphasis on insects, Oxford University Press, New York, USA, p. 211.
Carey, J. R. (2001). Insect biodemography. Annual Review of Entomology, 46, 79–110. https://doi.org/10.1146/annurev.ento.46.1.79
Carvalho, F. P. (2017). Pesticides, environment, and food safety. Food and Energy Security, 6(2), 48–60.
Chávez, J. P., Jungmann, D., & Siegmund, S. (2018). A comparative study of integrated pest management strategies based on impulsive control. Journal of Biological Dynamics, 12(1), 318–341. https://doi.org/10.1080/17513758.2018.1446551
Chen, Q., Li, N., Wang, X., Ma, L., Huang, J.-B., & Huang, G.-H. (2017). Age-stage, two-sex life table of Parapoynxcrisonalis(Lepidoptera: Pyralidae) at different temperatures. PLoS ONE, 12(3), e0173380. https://doi.org/10.1371/journal.pone.0173380
Chhetri, A. K., Aggarwal, P. K., Joshi, P. K., & Vyas, S. (2017). Farmers’ prioritization of climate-smart agriculture (CSA) technologies. Agricultural Systems, 151, 184–191. https://doi.org/10.1016/j.agsy.2016.10.005
Chi, H., & Su, H. Y. (2006). Age-stage, two-sex life tables of Aphidius gifuensis (Ashmead) (Hymenoptera: Braconidae) and its host Myzus persicae (Sulzer) (Homoptera: Aphididae) with mathematical proof of the relationship between female fecundity and the net reproductive rate. Environmental Entomology, 35, 10–21.
Chongdar, S., Chhetri, B., Mahato, S. K., & Saha, A. (2015). Production potentials and economic feasibility of improved sesame (Sesamum indicum L.) cultivars under varying dates of sowing in prevailing agro-climatic condition of North Bengal. International Journal of Agriculture Sciences, 7(2), 434–439.
Dadd, R. H. (1985). Nutrition: organisms. In: G. A. Kerkot, L. I. Gillbert (Eds.), Comprehensive insect physiology, biochemistry and pharmacology, Pergamon Press, New York, Oxford, pp. 313–390. https://doi.org/10.1016/b978-0-08-030805-0.50014-6.
Damalas, C. A., & Koutroubas, S. D. (2018). Current status and recent developments in biopesticide use. Agriculture, 8(1), 13. https://doi.org/10.3390/agriculture8010013
Dar, A. A., Choudhury, A. R., & Arumugam, N. (2014). A study on seed protein profile of Indian cultivars of Sesamum indicum L. International Journal of Current Biotechnology, 2(6), 10–17.
Dicke, M. (2000). Chemical ecology of host-plant selection by herbivorous arthropods: A multitrophic perspective. Biochemical Systematics and Ecology, 28, 601–617. https://doi.org/10.1016/S0305-1978(99)00106-4
Dutta, S., & Roy, N. (2018). Population dynamics and host preference of a major pest, Scripophaga incertulus Walker (Pyralidae: Lepidoptera). International Journal of Advance Scientific Research, 3(2), 120–127.
Dutta, S., & Roy, N. (2016). Life table and population dynamics of a major pest, Leptocorisa acuta (Thunb.) (Hemiptera: Alydidae), on rice and non-rice system. International Journal of Pure & Applied Biosciences, 4(1), 199–207. https://doi.org/10.18782/2320-7051.2202.
Golizadeh, A., & Razmjou, J. (2010). Life table parameters of Phthorimaea operculella (Lepidoptera: Gelechiidae), feeding on tubers of six potato cultivars. Journal of Economic Entomology, 103, 966–972.
Gotyal, B. S., Selvaraj, K., Meena, P. N., & Satpathy, S. (2015). Host plant resistance in cultivated jute and its wild relatives towards jute hairy caterpillar Spilosoma obliqua (Lepidoptera: Arctiidae). Florida Entomologist, 98(2), 721–727. https://doi.org/10.1653/024.098.0248
Harborne, J. B. (1973). Phytochemical methods: A guide to modern techniques of plant analysis (2nd ed., pp. 88–185). Chapman and Hall.
Harborne, J. B. (1994). Introduction to Ecological Biochemistry. Academic Press.
Heeb, L., Jenner, E., & Cock, M. J. W. (2019). Climate-smart pest management: Building resilience of farms and landscape to changing pest threats. Journal of Pest Science, 92, 951–969. https://doi.org/10.1007/s10340-019-01083-y
Higley, L. G., & Wintersteen, W. K. (1992). A novel approach to environmental risk assessment of pesticides as a basis for incorporating environmental costs into economic injury levels. American Entomologist, 38, 34–39.
Kakde, A. M., Patel, K. G., & Tayade, S. (2014). Role of life table in insect pest management-a review. IOSR Journal of Agriculture and Veterinary Science, 7(1), 40–43. https://doi.org/10.9790/2380-07114043
Kale, R. A., Waghmare, P. K., Gokhale, D. N., & Bhalerao, G. A. (2018). Effect of different plant geometry on growth and yield of sesame (Sesamum indicum L.) during post monsoon season in Marathwada region of Maharashtra. Journal of Pharmacognosy and Phytochemistry, 7(5), 428–429.
Kang, L. (2019). Overview: Biotic signalling for smart pest management. Philosophical Transactions of the Royal Society B, 374, 20180306. https://doi.org/10.1098/rstb.2018.0306
Karimi-Pormehr, M. S., Borzoui, E., Naseri, B., Dastjerdi, H. R., & Mansouri, S. M. (2018). Two-sex life table analysis and digestive physiology of Sitotroga cerealella (Olivier) (Lepidoptera: Gelechiidae) on different barley cultivars. Journal of Stored Products Research, 75, 64–71.
Kim, K. H., Kabir, E., & Jahan, S. A. (2017). Exposure to pesticides and the associated human health effects. Science of the Total Environment, 575, 525–535.
Krebs, C. J. (1994). Ecology: The experimental analysis of distribution and abundance, 4th edn. Harper Collins College Publishers.
Lal, R. (2008). Sequestration of atmospheric CO2 into global carbon pool. Energy and Environmental Science, 1, 86–100.
Lal, R. (2011). Sequestering carbon in soils of agro-ecosystems. Food Policy, 36, 33–39.
Liu, Z., Li, D., Gong, P., & Wu, K. (2004). Life table studies of the cotton bollworm, Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae), on different host plants. Environmental Entomology, 33, 1570–1576. https://doi.org/10.1603/0046-225X-33.6.1570
Mandal, D., Bhowmik, P., & Baral, K. (2013). Evaluation of insecticides for the management of Bihar hairy caterpillar, Spilosoma obliqua Walk. (Lepidoptera: Arctiidae) in black gram (Vigna mungo L.). The Bioscan, 8(2), 429–431.
Mobarak, S. H., Roy, N., & Barik, A. (2019). Two-sex life table and feeding dynamics of Spilosoma obliqua Walker (Lepidoptera: Arctiidae) on three green gram cultivars. Bulletin of Entomological Research, pp. 1–13. doi:https://doi.org/10.1017/S0007485319000452.
Mohapatra, M. M., & Gupta, P. K. (2018). Evaluation of insecticides against Bihar hairy caterpillar, Spilosoma obliqua Walk. on black gram, Vigna mungo (Linn.). International Journal of Current Microbiolog and Applied Sciences, 7(6), 605–608. https://doi.org/10.20546/ijcmas.2018.706.069.
Nagia, D. K., Kumar, S., & Saini, M. L. (1990). Relative toxicity of some important insecticides to Bihar hairy caterpillar, Spilosoma obliqua walker (Arctiidae: Lepidoptera). Journal of Entomological Research, 14(1), 60–62.
Naik, A., Patil, D. H., Teggelli, R. G., & Ahamed, B. Z. (2016). Productivity enhancement of sesame (Sesamum indicum L.) through improved production technologies/ frontline demonstrations. Research on Environment and Life Sciences, 9(5), 573–574.
Naseri, B., Golparvar, Z., Razmjou, J., & Golizadeh, A. (2014). Age-stage, two-sex life table of Helicoverpa armigera (Lepidoptera: Noctuidae) on different bean cultivars. Journal of Agricultural Science & Technology, 16, 19–32.
Nath, D. K. (1975). Note on the insect pests of sesame (Sesamum indicum L.) of West Bengal. Indian Journal of Agricultural Research, 9(3), 151–152.
Nation, J. L. (2001). Insect Physiology and Biochemistry, CRC Press, Boca Raton, FL. PMCid: PMC90373
Ndakidemi, B., Mtei, K., & Ndakidemi, P. A. (2016). Impacts of synthetic and botanical pesticides on beneficial insects. Agricultural Science, 7, 364–372.
Parui, A., & Roy, N. (2016). Ecofriendly and sustainable management of Spilosoma obliqua Walker on sesame (Sesamum indicum L.) crops by new botanicals. Journal of Entomolog and Zoology Studies, 4(6), 349–354.
Pedigo, L. P., & Buntin, G. D. (1994). Handbook of sampling methods for arthropods in agriculture. CRC Press.
Pedigo, L. P., & Higley, L. G. (1992). A new perspective of the economic injury level concept and environmental quality. American Entomology, 38, 12–21.
Pedigo, L. P., Hutchins, S. H., & Higley, L. G. (1986). Economic injury levels in theory and practice. Annual Review of Entomology, 31, 341–368.
Price, P. W. (1998). Insect Ecology. Wiley.
Roy, N. (2015a). Host phytochemicals in regulation of nutritional ecology and population dynamics of Podontia quatuordecimpunctata L. (Coleoptera: Chrysomelidae). International Journal of Horticulture, 5(4), 1–11. doi:https://doi.org/10.5376/ijh.2015a.05.0004.
Roy, N. (2015b). Life table and population parameters of Diacrisia casignetum Kollar (Lepidoptera: Arctiidae) on jute, Chorchorus capsularis (cv. Sonali; JRC-321), leaves. International Journal of Fauna Biological Studies, 2, 23–29.
Roy, N. (2017). Life table and nutritional ecology of Epilachna vigintioctopunctata Fab. (Colioptera: Coccinellidae) on three host plants. International Journal of Horticulture, 7(2), 7–19. doi:https://doi.org/10.5376/ijh.2017.07.0002.
Roy, N. (2018). Jute leaf physicochemical cues mediated behavioral responses of Diacrisia casignetum Kollar. Agricultural Research, pp 1–10. https://doi.org/10.1007/s40003-018-0362-2.
Roy, N. (2019). Life table and economic threshold concept for ecologically sustainable management of Diacrisia casignetum Kollar (Lepidoptera: Arctiidae) on Jute. Entomon, 44(2), 103–110. https://doi.org/10.33307/entomon.v44i.436
Roy, N., & Barik, A. (2012). The impact of variation in foliar constituents of sunflower on development and reproduction of Diacrisia casignetum Kollar (Lepidoptera: Arctiidae). Psyche, 2012, 9, Article ID 812091. doi:https://doi.org/10.1155/2012/812091.
Roy, N., & Barik, A. (2013). Influence of four host plants on feeding, growth and reproduction of Diacrisia casignetum (Lepidoptera: Arctiidae). Entomological Science, 16(1), 112–118. https://doi.org/10.1111/j.1479-8298.2012.00546.x
Roy, N., & Barik, A. (2014). Long-chain fatty acids: Semiochemicals for host location by the insect pest, Diacrisia casignetum. Journal of the Kansas Entomological Society, 87(1), 22–36.
Sarfraz, M., Dosdall, L. M., & Keddie, B. A. (2007). Resistance of some cultivated Brassicaceae to infestations by Plutella xylostella (Lepidoptera: Plutellidae). Journal of Economic Entomology, 100, 215–224.
Schoonhoven, L. M., Van Loon, J. J. A., & Dicke, M. (2005). Insect-plant biology. Oxford University Press.
Schowalter, T. D. (2006). Insect ecology: An ecosystem approach (2nd ed.). Academic Press.
Scriber, J. M., & Slansky, F., Jr. (1981). The nutritional ecology of immature insects. Annual Review of Entomology, 26, 183–211.
Shobana, K., Murugan, A., & Kumar, N. (2010). Influence of host plants on feeding, growth and reproduction of Papilio polytes (the common mormon). Journal of Insect Physiology, 56, 1065–1070. https://doi.org/10.1016/j.jinsphys.2010.02.018
Singh, Y. R., & Varatharajan, R. (1999). Host range of Bihar hairy caterpillar, Spilosoma obliqua (Walker) (Arctiidae: Lepidoptera). Hexapoda, 11(2), 65–74.
Slansky, F., & Scriber, J. M. (1985). Food consumption and utilization. In: G. A. Kerkot, L. I. Gillbert (Eds.), Comprehensive insect physiology, biochemistry and pharmacology, Pergamon, Oxford, England, pp. 87–113. https://doi.org/10.1016/b978-0-08-030805-0.50009-2.
Southwood, T. R. E. (1978). Ecological methods particular reference to study of insect population (p. 524). The English Language Book Society and Chapman and Hall.
Southwood, T. R. E., & Henderson, P. A. (2000). Ecological Methods (3rd ed., p. 575). Blackwell Science.
Subedi, R., Bhatta, L. D., Udas, E., Agrawal, N. K., Joshi, K. D., & Panday, D. (2019). Climate-smart practices for improvement of crop yields in mid-hills of Nepal. Cogent Food & Agriculture, 5, 1631026. https://doi.org/10.1080/23311932.2019.1631026
Varatharajan, R., Singh, S. A., Keisa, T. J., Singh, O. D., & Selvasundaram, R. (1998). Life table of Spilosoma obliqua (Lepidoptera: Arctiidae) on sunflower. Insect Science and Its Application, 18, 383–385.
Waldbauer, G. P. (1968). The consumption and utilization of food by insects. Advances in Insect Physiology, 5, 229–288. https://doi.org/10.1016/S0065-2806
Wang, Z. B., Zhang, H. L., Lu, X. H., Wang, M., Chu, Q. Q., Wen, X. Y., & Chen, F. (2016). Lowering carbon footprint of winter wheat by improving management practices in North China Plain. Journal of Cleaner Production, 112, 149–157.
War, A. R., Paulraj, M. G., Ahmad, T., Buhroo, A. A., Hussain, B., Ignacimuthu, S., & Sharma, H. C. (2012). Mechanisms of plant defense against insect herbivores. Plant Signaling and Behavior, 7, 1306–1320.
Win, S. S., Muhamad, R., Ahmad, Z. A., & Adam, N. A. (2011). Life table and population parameters of Nilaparvata lugens Stal. (Homoptera: Delphacidae) on rice. Tropical Life Sciences Research, 22(1), 25–35.
Wolfenbarger, L. L., & Phifer, P. R. (2000). The ecological risks and benefits of genetically engineered plants. Science, 290, 2088–2093.
Xue, M., Pang, Y. H., Wang, H. T., Li, Q.-L., & Liu, T.-X. (2010). Effects of four host plants on biology and food utilization of the cutworm, Spodoptera litura. Journal of Insect Science, 10, 1–14. https://doi.org/10.1673/031.010.2201
Zar, J. H. (1999). Biostatistical Analysis. Prentice Hall.
Acknowledgements
I wish to express my deep sense of gratitude to WBDST Project [File No.: ST/P/S&T/1G-29/2018], from Government of West Bengal, India, for financial assistance. I must acknowledge the farmers who help me in every way during my fieldwork.
Funding
I declare that the financial assistance was provided by the WBDST Project [File No.: ST/P/S&T/1G-29/2018], from Government of West Bengal, India.
Author information
Authors and Affiliations
Contributions
NR designed the whole study including sample collection, chemical analysis, index calculation, data analysis and drafts the manuscript with the help of institutional support. The author read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
I give my consent to The Editor of The Journal of Basic and Applied Zoology (Springer) that my manuscript be published in their journal.
Competing interests
The author declares that he has no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1
. S. obliqua infestations on sesame plant and their different feeding indices, population parameters, and other economic parameters.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Roy, N. Population ecology and economic thresholds-based time series for climate smart pest management of Spilosoma obliqua Walker (Lepidoptera: Arctiidae) on three sesame cultivars. JoBAZ 83, 19 (2022). https://doi.org/10.1186/s41936-022-00283-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41936-022-00283-w