
Abstract
Background
Among the medically important snakes in Nigeria, Echis ocellatus and Bitis arietans have the most lethal venom. These venoms were classified according to the presence of snake venom metalloproteinases (SVMPs), snake venom phospholipase A2 (PLA2s), and snake venom serine proteases (SVSPs). Toxicological analyzes were performed to understand the significance of different protein families in venoms.
Methods
Proteins were separated from venom using column chromatography. The skin and footpad of mice were used to determine hemorrhagic and edematogenic activities. Caprine blood plasma was used to test fibrinolytic activity in vitro.
Results
The results showed that, compared to the crude venom, the SVMP fraction induced hemorrhagic effects with a diameter of 26.00 ± 1.00 mm in E. ocellatus and 21.33 ± 1.52 mm in B. arietans. Both SVSP and SVMP had anticoagulant effects; however, the SVSP fraction had a stronger effect, with a longer anticoagulation time of 30.00 ± 3.00 min in E. ocellatus and 26.00 ± 2.00 min in B. arietans. These main venom toxins, SVMPs, SVSPs, and PLA2, were found to have edema-forming effects that were optimal at 2 h after envenomation. PLA2s had the highest edema-inducing activity, with onset 30 min after envenomation.
Conclusions
Given the importance of SVMPs in altering the integrity of the membrane structure and impairing the blood coagulation system, an antivenom that can specifically neutralize its activity could inhibit the hemorrhage effects of the venoms.
Similar content being viewed by others
Introduction
Snakebite envenoming is listed as a priority neglected tropical disease due to the significant burden of mortality and morbidity, which predominantly affects vulnerable populations in the tropical and subtropical regions of the world [1]. Envenomings are one of the leading causes of death and disability in sub-Saharan Africa, Asia, and Latin America [2,3,4]. Nigeria has the highest burden of snakebite envenomation in sub-Saharan Africa [4]. Recent estimates indicate that approximately 43,000 snakebites occur annually in Nigeria, resulting in around 19,000 deaths [4]. Among the native Nigerian viper species, Echis ocellatus and Bitis arietans are the most medically important [5, 6]. E. ocellatus accounts for approximately 66% of all cases of envenomation [5]. Its range dominates the northern savanna regions and the southern parts of the Oyo and Enugu states [7, 8]. While less frequently found, B. arietans also occurs in these same regions [4].
Proteomic and transcriptomic studies have provided information on the composition of viper snake venom. These venoms contain a variety of enzymatic and nonenzymatic protein families, with snake venom metalloproteinases (SVMPs), snake venom serine proteases (SVSPs), and snake venom phospholipase A2 (PLA2) often predominating [6, 9,10,11,12]. Lesser amounts of proteins from other families are also present. Research indicates that these proteins carry out a range of toxic activities in envenomed victims, resulting in local tissue damage and hemorrhage. According to published reports, SVMPs, SVSPs and PLA2 have been shown to damage tissues and induce bleeding through their specific enzymatic actions [3, 13,14,15].
Advancements in toxinology focus on neutralizing venom-induced lethality [3, 16]. Given the diversity of venomous snake species and the pathophysiological characteristics of snake venoms, evaluating the toxic activities of their constituent proteins can help to understand the relative importance of individual components that must be neutralized [3, 17]. This assessment could guide the development of novel and effective treatment strategies based on the 3Rs (Replacement, Reduction, and Refinement) concept, which involves using antitoxin mixtures as replacement or reinforcement for antivenoms [2, 3, 18].
Toxicovenomics incorporates multiomics data into the evaluation of venom toxins [19,20,21], which may allow the development of improved therapeutic approaches for antivenoms [21,22,23]. In a recent study, we conducted proteomic analysis of the venoms of Nigerian viper snakes E. ocellatus and B. arietans [6]. It revealed that metalloproteinases, phospholipases A2 and serine proteinases were the primary toxic protein families. The purpose of the current research is to evaluate the hemorrhagic, edematogenic, and in vitro fibrinolytic effects of these main venom protein groups identified in the study. This can help identify protein families that should be prioritized for neutralization by antivenom in snakebite cases, and it can also inform and provide guidance for developing therapeutic inhibitors of snake toxins, paving the way for new treatment options in the future.
Materials and methods
Experimental animals
All snake handling and experimental mouse protocols were observed and followed according to the Ahmadu Bello University Committee on Animal Use and Care (approval number: ABUCAUC/2022/048), as well as the revised ARRIVE guidelines. This study used fifty mice weighing 16.2 ± 1.4 g procured from the Faculty of Pharmaceutical Sciences at Ahmadu Bello University in Zaria, Nigeria.
Snake venoms
This study used venom from adult E. ocellatus and B. arietans that have been previously reported [6]. Snakes were captured from various regions and housed at the Serpentarium of the Department of Veterinary Pharmacology and Toxicology at Ahmadu Bello University. Venom was extracted manually [24] and those from snakes of the same species were combined. The collected venoms were frozen at −80 °C, lyophilized in a freeze dryer and stored at −20 °C.
Chemicals and reagents
The following reagents were obtained from Sigma-Aldrich (St. Louis, MI, USA): pure bovine serum albumin (BSA), 1,10-phenanthroline, ethylene diaminetetraacetic acid (EDTA), Bradford reagent, and phenylmethylsulfonyl fluoride (PMSF). Thermo Fisher Scientific (USA) supplied the consumables and chemicals used to prepare the buffers, which were analytical grade.
Isolation of the venom proteins SVMP, SVSP, and PLA2 from crude venom
The crude venoms were fractionated using column chromatography, focusing on the three main venom toxins identified in the proteomic analysis [6]: snake venom metalloproteinase, snake venom serine protease and phospholipase A2. Acetone precipitation was performed on each of the crude venoms. Four (4) ml of cold acetone was added to 10 mg/ml of the crude venom. The mixture was incubated at −20 °C for 15 min before being centrifuged at 15,700 × g at 4 °C for 15 min. The protein pellet was air dried and suspended in 100 μl of 1 × PBS buffer at pH 7.4. These served as the protein samples that were fractionated. Two (2) ml of venom at a concentration of 10 mg/ml were loaded onto a DEAE cellulose column (1.5 × 50 cm) preequilibrated with 50 mM phosphate buffer at pH 6. At a flow rate of 0.2 ml/min, the column was eluted stepwise with a NaCl gradient of 0.01–0.1 M. Twenty-five fractions of 2 ml each were collected and tested for metalloproteinase, serine protease, and phospholipase activity. Fractions with similar enzymatic activity were combined and loaded onto a sephadex G-75 column equilibrated with 50 mM phosphate buffer at pH 6.5. The column was eluted using the same buffer at a rate of 1 ml/min. Twenty-five 2 ml fractions were collected and tested for metalloproteinase, serine protease, and phospholipase activity.
Biochemical assay of venom proteins
The biochemical activities of various venom proteins were analyzed using a Cary 300 UV–visible spectrophotometer (Agilent Technologies, Santa Clara, USA).
Snake venom metalloproteinase assay
The activity of proteolytic metalloproteinases was measured using a slightly modified method as previously described [25]. Twenty (20) µl of venom protein were mixed with 100 µl of azocasein substrate (10 mg/ml of azocasein dissolved in 25 mM Tris, 150 mM NaCl, 5 mM CaCl2, pH 7.4) and incubated at 37 °C for 90 min. The reaction was stopped by adding 200 µl of 5% trichloroacetic acid and centrifuging for 15 min at 3,000 rpm. One hundred (100) µl of supernatant was mixed with 100 µl of 0.5 M NaOH, and casein hydrolysis was measured at 450 nm. The reaction mixture was tested in the presence and absence of 15 mM 1,10 phenanthroline, a metalloproteinase inhibitor. A unit of proteolytic activity was defined as a 0.2 unit change in absorbance per min.
Snake venom serine protease assay
The substrate solution for serine protease activity was prepared by dissolving Nα-benzoyl-DL-arginine-p-nitroaniline (BApNA, obtained from Sigma-Aldrich) in 5 ml of dimethylsulfoxide (DMSO) and adding 95 ml of 0.05 M Tris–HCl buffer at a pH of 8.2. The assay was carried out with and without the inclusion of 15 mM phenylmethylsulfonyl fluoride. Hydrolysis of BApNA was used to monitor proteolytic activity by mixing 50 μl of the sample with 100 μl of the substrate solution. Activity was determined after 10 min of incubation at 37° C by measuring the release of p-nitroanilide, which absorbs at 405 nm. One unit of protease activity was defined as causing a 0.01/min increase in absorbance [26].
Snake venom phospholipase A2 assay
An enzyme activity assay was performed to measure phospholipase A2 (PLA2) using an egg yolk substrate solution, as previously described [27]. The egg yolk substrate solution was prepared fresh each time by dissolving egg yolk in a 0.9% saline solution. The assay was carried out at room temperature by incubating 250 μl of the egg yolk substrate solution with 10 μl of PLA2. The absorbance was measured at 740 nm after 60 min of incubation. PLA2 activity was quantified by determining the amount of PLA2 venom required to reduce the turbidity of the solution by 0.01 absorbance units per min at 740 nm, under the conditions tested.
Protein quantification
The protein concentration of the venoms was quantified using the Bradford method with bovine serum albumin (BSA) serving as standard reference [28]. Replicate aliquots of each sample were incubated for 20 min at 26 °C with 250 μl of Bradford reagent (Sigma-Aldrich, USA) prior to spectrophotometric analysis. Subsequently, absorbance readings at 595 nm were measured on a Cary 300 UV–visible spectrophotometer (Agilent Technologies, Santa Clara, USA) to determine the protein content of venom samples.
One-dimensional gel electrophoresis
Venom samples were normalized to ensure equivalent protein content prior to separation by 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) under non-reducing conditions, as previously described [6]. Aliquots containing 10 µg of protein were loaded into each well and subjected to electrophoresis. After completion, the gels were stained with Coomassie Brilliant Blue G-250 to visualize the protein bands. Gel imaging was performed using the Bio-Rad PharosFX Plus System from the Molecular Imager (California, USA).
Toxicological analyses
A series of experiments was performed to evaluate the fibrinolytic, edematogenic, and hemorrhagic activities of the venom using mouse models, to gain a clear understanding of the toxicity.
Determination of hemorrhagic activity
The hemorrhagic activity of crude venom and isolated protein fractions (SVMP, SVSP, and PLA2) from E. ocellatus and B. arietans was determined using the mouse skin test [29]. Adult mice weighing approximately 16.2 ± 1.4 g were divided into nine groups of three mice each. Group 1 received E. ocellatus metalloproteinase, Group 2 received E. ocellatus serine protease, Group 3 received E. ocellatus phospholipase A2, Group 4 received B. arietans metalloproteinase, Group 5 received B. arietans serine protease, Group 6 received B. arietans phospholipase A2, Group 7 received crude E. ocellatus venom, Group 8 received crude B. arietans venom, while Group 9 served as controls and was injected with 10 μl of PBS. Each group of mice was injected intradermally into the abdominal region with 10 μg/μl of crude venom or isolated protein. After 2 h, the mice were humanely euthanized using chloroform. The skin was removed and the size of the hemorrhagic lesion on the inner surface was measured horizontally, vertically and diagonally. The average of these measurements was taken as the size of the hemorrhagic lesion. Hemorrhagic activity was expressed as the minimum hemorrhagic dose (MHD), defined as the dose required to induce a hemorrhagic area of 10 mm in diameter. To avoid limitations associated with similar hemorrhagic area sizes exhibiting varying hemorrhage levels [3], the amount of hemoglobin present in each lesion was also quantified [30].
Determination of venom-induced edema activity
The edema-inducing activities of the venom proteins were evaluated using a modified mouse footpad assay [31]. A separate group of experimental animals, divided into eight groups of three mice each, was subcutaneously injected with 10 μg/μl of venom protein in the right hind footpad and 10 μl of phosphate buffered saline in the left hind footpad as a control. The formation of edema was monitored and measured in various time intervals ranging from 0 min to 5 h after injection of the venom using a digital caliper (Mitutoyo, Japan). The percentage increase in the size of the envenomed foot compared to the size of the phosphate buffered saline-injected foot was used to calculate the formation of edema. A minimum edema-forming dose (MED) was defined as the amount of venom protein required to induce at least 30% edema.
Determination of fibrinolytic activity
One ml of caprine blood was mixed with 3.8% trisodium citrate and centrifuged at 4,300 rpm for 10 min at 4 °C to obtain platelet-poor plasma (PPP). This test utilized the yellowish supernatant, PPP. Calcium chloride-induced clotting time was used to assess the anticoagulant activity of crude venom and venom proteins in PPP [32]. The control for comparison was a phosphate buffered saline/PPP mixture. One unit of coagulant or anticoagulant activity was defined as an increase or decrease in clotting time of 1 s in PPP incubated with crude venom or venom proteins relative to control PPP.
Data and statistical analyses
The data were statistically analyzed with a statistical package of social sciences (IBM SPSS Inc. Chicago V 19.0) using one-way analysis of variance with a 95% confidence interval. The results were expressed as mean ± standard error of the mean. A p-value of less than 0.05 was used as the threshold for statistical significance. The results of the purification protocol of the enzymes are tabulated as total protein, total activity, specific activity, purification fold, and percentage yield. Graphs of the elution profile were plotted using Sigma Plot software (SPCC Inc., Chicago, IL, USA).
Results
One-dimensional SDS-PAGE of the venom of E. ocellatus and B. arietans proteins
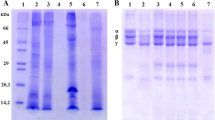
The SDS-PAGE analysis of the venom proteins; snake venom metalloproteinase (SVMP), snake venom serine proteinase (SVSP), and snake venom phospholipase A2 (PLA2) is shown in Fig. 1. Lanes 1 and 6 contain marker protein standards. Lane 2 contains crude venom from E. ocellatus. Lane 3 contains the SVSP fraction with an estimated molecular weight of 28 kDa. Lane 4 contains the SVMP fraction with an estimated molecular weight of 68 kDa. Lane 5 contains the PLA2 fraction with an estimated molecular weight of 13 kDa. Lane 7 shows crude venom from B. arietans. Lanes 8, 9, and 10 display the SVSP fraction with a molecular weight of 30 kDa, the SVMP fraction with a molecular weight of 95 kDa, and the PLA2 fraction with a molecular weight of 13 kDa, respectively.

One-dimensional SDS-PAGE of venom proteins of E. ocellatus and B. arietans
Hemorrhagic effects of E. ocellatus and B. arietans venom and venom proteins
Hemorrhagic lesions caused by crude venom and venom proteins (SVMP, SVSP and PLA2) of E. ocellatus are shown in Fig. 2. The hemorrhagic lesion measured an average of 26.00 ± 1.00 mm for SVMP, compared to 44.00 ± 1.00 mm for the crude venom. It measured an average of 7.00 ± 1.00 mm for the SVSP. Meanwhile, the PLA2 protein did not demonstrate any hemorrhagic effect.

Hemorrhagic lesion/ effects of E. ocellatus venom and venom proteins. Hemorrhagic activity was determined 2 h after injecting 10 μg/μl of crude venom or venom proteins (SVMP, SVSP and PLA2) intradermally into the abdominal region of mice. Size of hemorrhagic lesion: A Crude venom (44 mm), B SVMP (26 mm), C SVSP (7 mm), D PLA2 (0 mm), E Control (0 mm)
Hemorrhagic lesions caused by B. arietans venom and its constituent venom proteins (SVMP, SVSP, and PLA2) are shown in Fig. 3. The crude venom produced a hemorrhagic lesion measuring 52.33 ± 1.53 mm. The SVMP fraction resulted in lesions of 21.33 ± 1.52 mm. SVSP caused hemorrhagic lesions averaging 6.33 ± 1.53 mm in size. Meanwhile, PLA2 did not appear to induce any hemorrhagic effects according to the data.

Hemorrhagic lesion/effects of B. arietans venom and venom proteins. Hemorrhagic activity was determined 2 h after injecting 10 μg/μl of crude venom or venom proteins (SVMP, SVSP and PLA2) intradermally into the abdominal region of mice. Size of hemorrhagic lesion: A Crude venom (52 mm), B SVMP (21 mm), C SVSP (6 mm), D PLA2 (0 mm), E Control (0 mm)
Fibrinolytic effects of E. ocellatus and B. arietans venoms and venom proteins
The fibrinolytic effects of the venoms and venom proteins of E. ocellatus and B. arietans are presented in Table 1.
Edematogenic activity of the E. ocellatus and B. arietans venoms and venom proteins
The edematogenic effects of crude venom and venom proteins of E. ocellatus are presented in Table 1. The venom demonstrated time-dependent properties for inducing edema. Both the crude venom and SVSP fractions caused edema 30 min after envenomation. Meanwhile, the SVMP and PLA2 fractions induced edema 60 min later. In particular, the PLA2 fraction recorded the highest edema value of 5.84 ± 0.01 mm at 120 min.
According to the results of the study, the crude B. arietans venom caused edema within 1 h of envenomation. The SVMP and SVSP fractions induced edema within 120 min. The onset of the edema of the PLA2 fraction ranged from 30 min to 3 h, with a peak at 2 h. Mortality rates for the crude venom group were recorded between 3 and 5 h after envenomation.
Elution profile of Echis ocellatus venom proteins from diethylaminoethyl cellulose
Figure 4 shows the elution profile of E. ocellatus venom from diethylaminoethylcellulose (DEAE). 0.01–0.1 M sodium chloride gradients were used to elute the column. The peaks show that snake venom metalloproteinase (SVMP) was eluted in fractions 6, 7, 8, and 9 (panel A). Snake venom serine protease (SVSP) was eluted in fractions 11, 12, 13, 14, and 15 (panel B), while phospholipase A2 (PLA2) was eluted in fractions 17, 18, 19, 20, and 21 (panel C). proteins other than the protein of interest appeared to be present in the peaks of 1–4 and 13–21 (panel A), 6–9, 18–21 (panel B) and 6–15 (panel C).

Elution profile of Echis ocellatus venom proteins from DEAE cellulose. A Snake venom metalloproteinase (SVMP) fractions (Peaks 6, 7, 8, and 9), B Snake venom serine protease (SVSP) fractions (Peaks 11, 12, 13, 14 and 15), C Snake venom phospholipase A2 (PLA2) fractions (Peaks 17, 18,19, 20 and 21)
Elution profile of Echis ocellatus snake venom metalloproteinase (SVMP) from Sephadex G-75 column
The 6, 7, 8, and 9 fractions of the DEAE column with metalloproteinase activity were pooled, loaded on a Sephadex G-75 column, and eluted with phosphate buffered saline (PBS, pH 7.4). As shown in Fig. 5A, SVMP was eluted in fraction 8. Other proteins that were eluted alongside SVMP are represented by the eight minor peaks (peaks 5–7, and 9–20) shown in the figure. However, they did not exhibit metalloproteinase activity.

Elution profile of Echis ocellatus venom proteins from Sephadex G-75 columns. A Snake venom metalloproteinase (SVMP), B Snake venom serine protease (SVSP), C Snake venom phospholipase A2 (PLA2)
Elution profile of the snake venom serine protein (SVSP) of Echis ocellatus from Sephadex G-75 column
Similarly, fractions 11, 12, 13, 14, and 15 from the DEAE column were pooled and loaded on a Sephadex G-75 column, which was then eluted with PBS (pH 7.4). As illustrated in Fig. 5B, SVSP was eluted in fraction 13. There were 9 peaks of interfering proteins (peaks 5, 7–11, 15–17, and 22), without protease activity.
Elution profile of Echis ocellatus venom phospholipase A2 (PLA2) from Sephadex G-75 column
Likewise, fractions with phospholipase activity from the DEAE column (17, 18, 19, 20, and 21) were pooled and loaded onto a Sephadex G-75 column before being eluted with PBS (pH 7.4). PLA2 from the venom of E. ocellatus was eluted in fraction 21, as shown in Fig. 5C. The minor proteins eluted as shown (peaks 6, 11, 13, 15, 17, 19, 22, and 23) lacked phospholipase activity and were thus considered as interfering proteins.
Elution profile of Bitis arietans venom proteins from diethylaminoethylcellulose
The elution profile of B. arietans venom proteins from a diethylaminoethyl (DEAE) cellulose column is shown in Fig. 6. Sodium chloride gradients of 0.01–0.1 M were used to elute the column. B. arietans SVMP was eluted in fractions 5, 6, 7, 8, 9, and 10 of the 25 fractions collected (panel A), as shown by the peaks. B. arietans SVSP was eluted in fractions 13, 14, 15, and 16 (panel B), while Phospholipase A2 was eluted in fractions 18, 19, 20, 21, and 22 (panel C). Fractions with the same activity were pooled and subjected to a second purification step using Sephadex G-75. Other proteins were found in the fractions in addition to the protein of interest, as indicated by peaks 13–23 (panel A), 5–11, 18–23 (panel B), 5–16, and 24 (panel C).

Elution profile of Bitis arietans venom proteins from DEAE cellulose. A Snake venom metalloproteinase (SVMP) fractions (5, 6, 7, 8, 9 and 10), B Snake venom serine protease (SVSP) fractions (Peaks 13, 14, 15 and 16), C Snake venom phospholipase A2 (PLA2) fractions (Peaks 18, 19, 20, 21 and 22)
Elution profile of Bitis arietans venom metalloproteinase from Sephadex G-75 column
The elution profile of B. arietans venom metalloproteinase (SVMP) from a Sephadex G-75 column is shown in Fig. 7A. PBS (pH 7.4) was used to elute the column, and SVMP was eluted in fraction 11. Other proteins (peaks 6, 8, 12, 13, 17–20) were present, but they lacked metalloproteinase activity.

Elution profile of Bitis arietans venom proteins from Sephadex G-75 columns. A Snake venom metalloproteinase (SVMP), B Snake venom serine protease (SVSP), C: Snake venom phospholipase A2 (PLA2)
Elution profile of Bitis arietans venom serum protein (SVSP) from Sephadex G-75 column
As shown in Fig. 7B, B. arietans venom SVSP was eluted from the Sephadex G-75 column in fraction 16. Peaks 6–9, 14, 15, 19, 20, and 22 were other proteins eluted but without protease activity.
Elution profile of Bitis arietans venom phospholipase A2 (PLA2) from Sephadex G-75 column
Figure 7C shows the elution profile of PLA2 of the venom of B. arietans. As shown in the figure, it was eluted with PBS (pH 7.4) in fraction 19. Other proteins that eluted as shown (peaks 6, 13, 15, and 21) lacked phospholipase activity.
Purification profiles of metalloproteinase (SVMP), serine protease (SVSP) and phospholipase A2 (PLA2)
The E. ocellatus venom proteins were purified in three steps: acetone precipitation, ion exchange chromatography, and gel filtration. The purification profiles for SVMP, SVSP, and PLA2 are shown in Table 4. The total protein concentration of SVMP was 0.32 µg, the percentage yield was 25.63, the purification fold was 1.89, and the specific activity was 5.13 µmol/min/mg. SVSP was purified to a purification fold of 1.46 and a total protein concentration of 0.39 µg. The protein had a specific activity of 3.26 µmol/min/mg, a percentage yield of 23.43, and a total activity of 1.27 µmol/min. PLA2 had a total protein concentration of 0.56 µg, a yield of 42.07%, a specific activity of 3.55 µmol/min/mg, and was purified to a purification fold of 1.78.
Similarly, the purification profiles of B. arietans SVMP, SVSP, and PLA2 are shown in Table 5. SVMP had 0.16 µg of total protein, 0.85 µmol/min of total activity, and 5.31 µmol/min/mg of specific activity after gel filtration. The protein was purified to a purification fold of 2.55 and had a yield of 15.75%. SVSP was purified to 1.68 purification fold with a yield of 2.30%. It had 0.32 µg total protein, 0.96 µmol/min total activity and 3.00 µmol/min/mg specific activity. PLA2 contained 0.14 µg of total protein and was purified to a purification fold of 3.35 with a yield of 17.78%. The protein had a total activity of 0.53 µmol/min and a specific activity of 3.79 µmol/min/mg. The enzyme purification profile was formulated using the process outlined in Eq. 1.
Protein purification table calculations formular: Eq. 1.
Discussion
This study describes the main toxins responsible for the predominant toxicological effects in the venoms of the Nigerian saw-scaled viper Echis ocellatus and puff adder Bitis arietans. Previous proteomic analyzes revealed that metalloproteinases, phospholipase A2s, and serine proteases were the main toxins in these vipers [6], which is consistent with viper venoms from other regions [3, 33, 34]. Since venom proteins can act synergistically or independently to cause tissue damage [3, 35], this study performed toxicological studies on snake venom metalloproteinases (SVMPs), snake venom serine proteases (SVSPs), and snake venom phospholipases A2 (PLA2) to determine their relative importance for toxicity. The toxicological activity of these venom components determined by the mouse skin test (Figs. 2B and 3B) revealed that SVMP is primarily a hemorrhagic protein due to the damage to the capillary vessels that it causes [36, 37]. SVMPs are known to cause hemorrhage by cleaving the basement membrane of capillary vessels, causing the vessels to weaken. This results in the detachment of endothelial cells from the basement membrane, thinning it and eventually disrupting the capillary walls, leading to extravasation of blood [37, 38].
When SVMPs degrade the integrity of cell membranes, the tissues are more susceptible to infiltration by other venom toxins. It leads to pathophysiological conditions caused by a complex mixture of toxins in the venom [3, 39]. This underscores the pathological importance of neutralizing SVMPs as one of the key components in the event of envenomation. The hemorrhagic lesions in Figs. 2B and 3B demonstrate that venoms have a high propensity for hemorrhage, attributable to SVMP. This aligns with proteomic data [6], especially in the venom of E. ocellatus, where SVMPs represented the highest percentage (34.84%) of total venom content. Therefore, the hemorrhagic effects of E. ocellatus SVMP observed in this study indicate that the envenomation of E. ocellatus would likely prove more lethal than the envenomation of B. arietans. Consequently, any antivenom capable of neutralizing SVMPs has the potential to significantly mitigate the toxic effects of venom. In addition to inducing hemorrhage, it also interferes with blood coagulation pathways, and E. ocellatus inhibits coagulation to a greater extent than B. arietans. The results of this study corroborate previous reports that SVMPs affect hemostasis by altering coagulation, promoting hemorrhage [40, 41] by modulating key enzymes such as fibrinogenase and fibrolase [42, 43]. Evaluation of edema revealed involvement of SVMP in inflammatory processes for both venoms. However, the onset of edema was more rapid for E. ocellatus (Table 2) than for B. arietans (Table 3). This is consistent with previous findings of SVMP inflammatory properties [41, 44, 45].
A study examining the functional analysis of Brazilian Bothrops atrox venom proteins found that metalloproteinase produced a hemorrhagic effect with a diameter of 16 mm at a concentration of 10 µg [46]. For comparison, the venom of the Nigerian snake E. ocellatus caused a larger hemorrhagic diameter of 26 mm also at 10 µg, while the venom of B. arietans resulted in a 21 mm. These divergent results can be attributed to several potential factors. Differences in venom protein expression levels between snake populations that inhabit disparate regions worldwide could explain the observed variances in hemorrhagic effect. Local evolutionary pressures and environmental conditions may have shaped adaptive changes in venom composition and toxicity over time between snake species in different geographic locales.
The primary toxicity mechanism of SVSPs in both venoms involves anticoagulation. They exhibited longer plasma recalcification times than crude venoms (Table 1), suggesting that SVSPs are hemotoxic proteins. This effect is more pronounced in E. ocellatus, which has a longer clotting time than in B. arietans. SVSPs have exact toxicity by acting as thrombin-like enzymes or plasminogen activators, which eliminate fibrin in blood clots and contribute significantly to coagulopathy [14, 47]. Activation, inactivation, or depletion of coagulation factors prevents blood clotting, resulting in noncoagulable blood and bleeding [47]. In addition to altering the hemostatic system as their primary toxicity mechanism, SVSPs also cause edema in E. ocellatus (Table 2) and B. arietans (Table 3). This is consistent with reports in the literature of SVSP inducing edema, although the precise mechanism remains unknown [48].
According to mouse footpad analyses, PLA2 functions primarily as a protein that induces edema (Tables 2 and 3). It causes edema by inducing thermal allodynia and mechanical hyperalgesia [49, 50]. In addition to its ability to induce edema, it also possesses anticoagulant properties, with coagulation times of 10 and 12 min observed in the venom of E. ocellatus and B. arietans, respectively (Tables 2 and 3). These anticoagulant effects provide validity to reports that PLA2 increases hemorrhage by inhibiting the blood coagulation process [1, 49]. The results of the elution and purification processes of the venom proteins are presented in Figs. 4, 5, 6, 7. Subsequently, Tables 4 and 5 show the results of the purification of the enzymes as the specific activity, total activity, yield, and purification factor. Ideally, protocols to purify enzymes from snake venom should achieve several key objectives, such as obtaining enzymes at a high level of purity. This would enhance both the specific activity and purification fold. The specific activities of the enzymes of both venoms were not efficiently enhanced as a result of the protocol used for enzyme purification. As such, one of the limitations of this study was the purification protocol.
Conclusions
Identification and understanding of common venom toxins and their toxicological effects can be critical in avoiding treatment failures and developing new therapeutic approaches for antivenom design [3, 21, 23, 51]. When comparing the hemorrhagic effects of the SVMP fraction with crude venom, the fraction generated hemorrhagic lesions of 26.00 ± 1.00 mm in E. ocellatus and 21.33 ± 1.52 mm in B. arietans. Considering the importance of SVMP in toxicity by disrupting membrane integrity, we conclude that SVMPs represent a high-priority target for neutralization. Therefore, the development of therapeutics aimed at SVMP inhibition may serve to limit tissue damage after envenomation by blocking this initial breach of the membrane barrier.
Availability of data and materials
The data sets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Chippaux JP. Snakebite envenomation turns again into a neglected tropical disease! J Venom Anim Toxins Incl Trop Dis. 2027;23:38.
Chippaux JP. Estimate of the burden of snakebites in sub-Saharan Africa: a meta-analytic approach. Toxicon. 2011;57:586–99.
Gutierrez JM, Solano G, Pla D, Herrera M, Segura A, Vargas M, et al. Preclinical evaluation of the efficacy of antivenoms for snakebite envenoming: state-of-the-art and challenges ahead. Toxins. 2017;9:163.
Gutierrez JM, Maduwage K, Iliyasu G, Habib A. Snakebite envenoming in different national contexts: Costa Rica, Sri Lanka, and Nigeria. Toxicon X. 2021;25:9–10.
Habib AG. Venomous snakes and snake envenomations in Nigeria. Toxinol. 2015;2:275.
Dingwoke EJ, Adamude FA, Mohamed G, Klein A, Salihu A, Abubakar MS, Salla AB. Venom proteomic analysis of medically important Nigerian viper Echis ocellatus and Bitis arietans snake species. Biochem Biophys Rep. 2021;28:101164.
Warrell DA, Davidson NMD, Greenwood BM, Ormerod LD, Pope HM, Watkins BJ, Prentice CR. Poisoning by bites of the saw-scaled or carpet viper (Echis carinatus) in Nigeria. Q J Med. 1977;46:33–62.
Warrell DA, Arnett C. The importance of bites by the saw-scaled or carpet viper (Echis carinatus): epidemiological studies in Nigeria and a review of the world literature. Acta Trop. 1976;33:307–41.
Wagstaff SC, Harrison RA. Venom gland EST analysis of the saw-scaled viper, Echis ocellatus, reveals novel alpha9beta1 integrin-binding motifs in venom metalloproteinases and a new group of putative toxins, renin-like aspartic proteases. Gene. 2006;377:21–32.
Casewell NR, Harrison RA, Wuster W, Wagstaff SC. Comparative venom gland transcriptome surveys of the saw-scaled vipers (Viperidae: Echis) reveal substantial intra-family gene diversity and novel venom transcripts. BMC Genomics. 2009;10:564.
Casewell NR, Wagstaff SC, Wuster W, Cook DA, Bolton FM, King SI, et al. Medically important differences in snake venom composition are dictated by distinct postgenomic mechanisms. Proc Natl Acad Sci USA. 2014;111(25):9205–10.
Lomonte B, Fernandez J, Sanz L, Angulo Y, Sasa M, Gutierrez JM, Calvete JJ. Venomous snakes of Costa Rica: biological and medical implications of their venom proteomic profiles analyzed through the strategy of snake venomics. J Proteomics. 2014;105:323–39.
Gutierrez JM, Theakston RD, Warrell DA. Confronting the neglected problem of 607 snakebite envenoming: the need for a global partnership. PLoS Med. 2006;3:e150.
Kang TS, Georgieva D, Genov N, Murakami MT, Sinha M, Kumar RP, et al. Enzymatic toxins from snake venom: structural characterization and mechanism of catalysis. FEBS J. 2011;278(23):4544–76.
Calvete JJ. Venomics: integrative venom proteomics and beyond. Biochem J. 2017;474:611–34.
Kini RM. Toxinology provides multidirectional and multidimensional opportunities: a personal perspective. Toxicon X. 2020;6:100039.
Lauridsen LP, Laustsen AH, Lomonte B, Gutierrez JM. Toxicovenomics and antivenom profiling of the Eastern green mamba snake (Dendroaspis angusticeps). J Proteomics. 2016;136:248–61.
Roncolato EC, Pucca MB, Funayama JC, Bertolini TB, Campos LB, Barbosa JE. Human antibody fragments specific for Bothrops jararacussu venom reduce the toxicity of other Bothrops sp. venoms. J Immunotoxicol. 2013;10:160–8.
Laustsen AH, Lohse B, Lomonte B, Engmark M, Gutierrez JM. Selecting key toxins for focused development of elapid snake antivenoms and inhibitors guided by a Toxicity score. Toxicon. 2015;104:43–5.
Calvete JJ, Lomonte B. A bright future for integrative venomics. Toxicon. 2015;107:159–62.
Slagboom J, Kaal C, Arrahman A, Vonk FJ, Somsen GW, Calvete JJ, Wuster W, Kool J. Analytical strategies in venomics. Microchem J. 2022;175:107187.
Gutierrez JM, Lomonte B, Sanz L, Calvete JJ, Pla D. Immunological profile of antivenoms: preclinical analysis of the efficacy of a polyspecific antivenom through antivenomics and neutralization assays. J Proteomics. 2014;105:340–50.
Casewell NR, Jackson TNW, Laustsen AH, Sunagar K. Causes and consequences of snake venom variation. Trends Pharmacol Sci. 2020;41:570–81.
Hill RE, Mackessy SP. Venom yields from several species of colubrid snakes and differential effects of ketamine. Toxicon. 1997;35:671–8.
Gutierrez JM, Sanz L, Escolano J, Fernandez J, Lomonte B, Angulo Y, et al. Snake venomics of the Lesser Antillean pit vipers Bothrops caribbaeus and Bothrops lanceolatus: correlation with toxicological activities and immunoreactivity of a heterologous antivenom. J Proteome Res. 2008;7:4396–408.
Roldan-Padron O, Castro-Guillen JL, Garcia-Arredondo JA, Cruz-Perez MS, Diaz-Pena LF, Saldana C, Blanco-Labra A, Garcia-Gasca T. Snake venom hemotoxic enzymes: biochemical comparison between Crotalus species from Central Mexico. Molecules. 2019;24(8):1489.
Senji Laxme RR, Khochare S, de Souza HF, Ahuja B, Suranse V, Martin G, Whitaker R, Sunagar K. Beyond the “big four”: venom profiling of the medically important yet neglected Indian snakes reveals disturbing antivenom deficiencies. PLoS Negl Trop Dis. 2019;13(12):e0007899.
Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–54.
Gutierrez JM, Gene JA, Rojas G, Cerdas L. Neutralization of proteolytic and hemorrhagic activities of Costa Rican snake venoms by a polyvalent antivenom. Toxicon. 1985;23:887–93.
Ownby CL, Colberg TR, Odell GV. A new method for quantitating hemorrhage induced by rattlesnake venoms: ability of polyvalent antivenom to neutralize hemorrhagic activity. Toxicon. 1984;22:227–33.
Gutierrez JM, Rojas G, Lomonte B, Gene JA, Cerdas L. Comparative study of the edema-forming activity of Costa Rican snake venoms and its neutralization by a polyvalent antivenom. Comp Biochem Physiol Part C: Toxicol Pharmacol. 1986;85:171–5.
Mukherjee AK, Kalita B, Thakur R. Two acidic, anticoagulant PLA2 isoenzymes purified from the venom of monocled cobra Naja kaouthia exhibit different potency to inhibit thrombin and factor Xa via phospholipids independent, non-enzymatic mechanism. PLoS ONE. 2014;9(8):e101334.
Calvete JJ, Juarez P, Sanz L. Snake venomics. Strategy and applications. J Mass Spectrom. 2007;42:1405–14.
Tasoulis T, Isbister GK. A review and database of snake venom proteomes. Toxins. 2017;9(673):E290.
Doley R, Kini RM. Protein complexes in snake venom. Cell Mol Life Sci. 2009;66:2851–71.
Escalante T, Rucavado A, Fox JW, Gutierrez JM. Key events in microvascular damage induced by snake venom hemorrhagic metalloproteinases. J Proteom. 2011;74:1781–94.
Gutierrez JM, Escalante T, Rucavado A, Herrera C. Hemorrhage caused by snake venom metalloproteinases: a journey of discovery and understanding. Toxins. 2016;8:93.
Gutierrez JM, Escalante T, Rucavado A, Herrera C, Fox JW. A Comprehensive view of the structural and functional alterations of extracellular matrix by snake venom metalloproteinases (SVMPs): novel perspectives on the pathophysiology of envenoming. Toxins. 2016;8:304.
Nanjaraj Urs AN, Ramakrishnan C, Joshi V, Suvilesh KN, Veerabasappa Gowda T, Velmurugan D, Vishwanath BS. Progressive hemorrhage and myotoxicity induced by Echis carinatus venom in murine model: neutralization by inhibitor cocktail of N, N, N’, N’-Tetrakis (2-Pyridylmethyl) Ethane-1,2-Diamine and Silymarin. PLoS ONE. 2015;10(8):e0135843.
Takeda S, Takeya H, Iwanaga S. Snake venom metalloproteinases: structure, function and relevance to the mammalian ADAM/ADAMTS family proteins. Biochem Biophys Acta. 2012;1824:164–76.
Slagboom J, Kool J, Harrison RA, Casewell NR. Haemotoxic snake venoms: their functional activity, impact on snakebite victims and pharmaceutical promise. Br J Haematol. 2017;177:947–59.
Gutierrez JM, Rucavado A, Escalante T, Dias C. Hemorrhage induced by snake venom metalloproteinases: biochemical and biophysical mechanisms involved in microvessel damage. Toxicon. 2005;45:997–1011.
Moura-da-Silva AM, Almeida MT, Portes-Junior JA, Nicolau CA, Gomes-Neto F, Valente RH. Processing of snake venom metalloproteinases: generation of toxin diversity and enzyme inactivation. Toxins (Basel). 2006;8:183.
Kini RM, Koh CY. Metalloproteases affecting blood coagulation, fibrinolysis and platelet aggregation from snake venoms: definition and nomenclature of interaction sites. Toxins. 2016;8:E284.
Ainsworth S, Slagboom J, Alomran N, Pla D, Alhamdi Y, King SI, et al. The paraspecific neutralisation of snake venom induced coagulopathy by antivenoms. Commun Biol. 2018;1:34.
Menaldo DL, Jacob-Ferreira AL, Bernardes CP, Cintra AC, Sampaio SV. Purification procedure for the isolation of a P-I metalloprotease and an acidic phospholipase A2 from Bothrops atrox snake venom. J Venom Anim Toxins Incl Trop Dis. 2015;13(21):28.
Serrano SMT. The long road of research on snake venom serine proteinases. Toxicon. 2013;62:19–26.
Ferraz CR, Arrahman A, Xie C, Casewell NR, Lewis RJ, Kool J, Cardoso FC, et al. multifunctional toxins in snake venoms and therapeutic implications: from pain to hemorrhage and necrosis. Front Ecol Evol. 2019. https://doi.org/10.3389/fevo.2019.00218.
Lomonte B, Rangel J. Snake venom Lys49 myotoxins: from phospholipases A2 to non-enzymatic membrane disruptors. Toxicon. 2012;60:520–30.
Zhang CC, Medzihradszky KF, Sanchez EE, Basbaum AI, Julius D. Lys49 myotoxin from the Brazilian lancehead pit viper elicits pain through regulated ATP release. Proc Natl Acad Sci USA. 2017;114:E2524–32.
de Souza LL, Stransky S, Guerra-Duarte C, Flor-Sa A, Schneider FS, Kalapothakis E, Chávez-Olortegui C. Determination of toxic activities in Bothrops spp. snake venoms using animal-free approaches: correlation between in vitro versus in vivo assays. Toxicol Sci. 2015;147(2):458–65.
Acknowledgements
The authors express their gratitude to the Head of the Department of Veterinary Pharmacology and Toxicology at Ahmadu Bello University for permitting the use of departmental laboratory facilities for this research. The author also thanks Dr. Yusuf Peter Ofemile for providing technical assistance. Laboratory technicians Otie Dennis, Yusuf Abdulraheem Oniwaapele, and Hashimu Abdulwahab Yau are appreciated for their valuable assistance in conducting the laboratory work, ensuring procedures were followed correctly and that the analysis was completed in a timely and accurate manner. This analysis was performed in partial fulfillment of the doctoral degree requirements for Emeka John Dingwoke at the Department of Biochemistry, Ahmadu Bello University, Zaria.
Funding
This study did not receive outside funding.
Author information
Authors and Affiliations
Contributions
EJD: methodology, investigation, formal analysis, data collection, writing – original draft, writing-review & editing, visualization. FAA: methodology, investigation, formal analysis. AS: supervision, writing-review & editing. ABS: supervision, writing-review & editing. MSA: supervision. All authors: conceptualization. EJD, FAA: funding acquisition.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
All animal experiments were approved by the Ahmadu Bello University Animal Use and Care Committee (ABUCAUC). They were performed using protocols approved by the Committee.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Dingwoke, E.J., Adamude, F.A., Salihu, A. et al. Toxicological analyses of the venoms of Nigerian vipers Echis ocellatus and Bitis arietans. Trop Med Health 52, 15 (2024). https://doi.org/10.1186/s41182-024-00581-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41182-024-00581-9