
Abstract
Background
Cardiovascular diseases constitute the main death cause worldwide resulting from a combination of genetic and lifestyle factors, and the prevalence among younger individuals has increased. It is important to early identify changes in lipid profile and the influence of genetic variations in specific genes on the individual patterns of lipid profile. Thus, the aim of this study was to verify the relationship of polymorphisms in PPAR-gamma gene (PPARG − rs1801282 − Pro12Ala) and in apolipoprotein E gene (APOE − rs429358 + rs7412, determinants of the APOE2, APOE3, or APOE4 genotypes) with lipid profile of adolescents under cardiovascular risk factors.
Methods
This was a cross-sectional study with 115 adolescents aged 10–19 years, which presented cardiovascular risk factors. The students were evaluated regarding socioeconomic, anthropometric, biochemical, genetic, and dietetic variables. Student’s t test or Mann-Whitney test were applied to the analysis of the genotypes. Multiple linear regression analysis was performed to determine the variables that most influenced the lipid profile.
Results
Adolescents carrying PPARG Ala allele showed higher serum triglycerides (p = 0.0423) and very low-density lipoprotein (p = 0.0410) levels when compared to those carrying the wild genotype. For the APOE polymorphism, it was observed a trend of higher triglycerides (p = 0.0712) and very low-density lipoprotein (p = 0.0758) levels in the adolescents carrying the E4 allele when compared to those who did not carry this allele.
Conclusion
The polymorphisms PPARG rs1801282 and APOE rs429358 + rs7412 seem to be related to the development of lipid profile alterations in adolescents.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Background
Atherosclerosis, the main pathophysiological process related to cardiovascular disease (CVD), begins in childhood and progresses toward adulthood [1]. The extent of atherosclerosis in children and adolescents may be associated with the presence of the same risk factors that have been identified in adults, such as changes in lipid profile, overweight, hypertension, age, sex, and genetic inheritance [2].
A study with more than 3000 US adolescents from the National Health and Nutrition Examination Survey (NHANES) evaluated the recent trends (from 1999 to 2008) in the prevalence of CVD risk, both separately and according to weight (normal, overweight or obesity) status. Based on the overall results, it was concluded that American adolescents, especially those who are overweight or obese, present a significant load of risk factors for CVD [3].
The Brazilian Study of Cardiovascular Risks in Adolescents (ERICA) started in 2008 with the aim of estimating cardiovascular risk factors prevalence in adolescents attending public and private schools in cities bigger than 100 thousands inhabitants. More than 35,000 adolescents were evaluated regarding alterations in lipid profile, and the first analyses showed that a meaningful part of the evaluated adolescents presented changes in plasma lipid levels, especially regarding low levels of cholesterol in high-density lipoprotein (HDL-C) and high triacylglycerol levels [4].
Patients with dyslipidemia or under some other cardiovascular risk factors should be counseled about the adoption of non-pharmacological measures related to healthy lifestyle, including changes in dietary habits. However, individuals subjected to the same treatment or changes in habits may show different responses, depending on polymorphisms in genes or other DNA sequences involved in lipid metabolism. Among these polymorphisms, the rs1801282 (Pro12Ala) in the peroxisome proliferator-activated receptor gamma gene (PPARG) and the rs429358 + rs7412 (determinant of the alleles ε2, ε3, and ε4) in the apolipoprotein E gene (APOE) must be highlighted [5].
PPARG is a nuclear receptor, which plays an essential role in lipid metabolism, as well as in the differentiation of adipocytes and lipid storage. The Pro12Ala polymorphism in PPARG may lead to an alteration of the intrinsic activity of this receptor. Presence of the Ala allele is associated with reduced lipoprotein lipase activity, resulting in altered clearance of low-density lipoproteins (LDL) and very low-density lipoproteins (VLDL), with consequent increase in lipoprotein plasma levels [6]. These changes in lipid metabolism in subjects carrying the Ala allele were confirmed by a meta-analysis with 48,210 individuals, which included association studies between the Pro12Ala polymorphism and lipid profile published until May 2014 [7].
APOE is a serum glycoprotein present in circulating chylomicrons (CM), remaining CM, and VLDL. This glycoprotein also plays an important role in lipid metabolism, as it mediates the interaction of lipoproteins to LDL and remaining CM receptors. In consequence, APOE is involved with the metabolic destination of some lipoprotein classes and plays a central role in cholesterol metabolism [8]. It is estimated that 60% of the variation in serum total cholesterol levels undergo influence of a genetic determinant and that 14% of the changes in these levels are influenced by polymorphisms in the APOE [9].
APOE interactions with lipoprotein receptors depend on its isoform. The most studied polymorphism in APOE is characterized by the cistein and arginin contents at positions 112 and 158, resulting in three major alleles (ε2, ε3, ε4) and in six possibilities of genotypes: ε2/2, ε3/2, ε4/2, ε3/3, ε4/3, and ε 4/4 [8]. Meta-analysis including case-control studies with 18,752 patients suffering from acute myocardial infarction (AMI) suggests that the ε4 allele is a risk factor for the development of AMI and the presence of the ε2 allele is a protective factor for this disease. These results may be explained by the fact that the ε4 allele is associated with elevated serum concentrations of LDL-C, VLDL-C, triacylglycerol, and total cholesterol, while the ε2 allele is associated with lower concentrations of these substances [10].
Considering the importance of early identification of changes in lipid profile for CVD prevention, the influence of genetic variations in specific genes on the individual patterns of lipid profile, and the lack of studies with Brazilian adolescents on this subject, the aim of this study was to investigate the relationships among PPARG Pro12Ala and APOE rs429358 + rs7412 polymorphisms and lipid profile in adolescents with cardiovascular risk factors.
Methods
Study population
An observational, analytical, and cross-sectional study was performed, including Brazilian adolescents from a public school located at center-western Brazil, as described previously [11]. All the students aged 10–19 years were invited, and 454 agreed to participate. They were evaluated regarding socioeconomic, anthropometric, lifestyle, and dietetic behavior variables. Adolescents who presented overweight or self-reported changes in lipid profile (n = 201) were invited to perform biochemical and genetic analysis. Individuals who presented previous CVD diagnosis (n = 06), who were under nutritional treatment (n = 08), who did not conclude the biochemical evaluations (n = 20), or who did not accept to participate in the biochemical analyses (n = 52), were not included in the study. The final sample consisted of 115 adolescents, 80 presenting overweight, and 35 self-reported changes in lipid profile. A posteriori determination of the sample representativeness in relation to the target population, with a confidence interval of 95% and a power of 80%, showed the maximum error of 5.4% considering the frequency of the variant allele for the PPARG Pro12Ala polymorphism.
The study was approved by the Ethics Committee of the Federal University of Goiás on 10.07.2013, under protocol number 422 352. All the procedures in this study were in accordance with the 1975 Helsinki Declaration, updated in 2013. Informed consent was obtained from all participants included in the study.
Dietary evaluation
The subjects answered a usual food record filled by trained professionals. The adolescents were instructed on how to provide accurate information and about the importance of giving details on portions and preparations in household measures. Nutritional assessment of food records was performed using the software Avanutri® (Rio de Janeiro, Brazil).
Body composition
All measurements were performed with participants barefoot and wearing light clothes, according to the protocol established by Gibson [12]. The height was measured through a stadiometer graduated in centimeters. Body weight was measured with a graduated scale in 0.1 kg, and the waist circumference was determined with a non-flexible tape. Body mass index (BMI) was calculated from weight and height data. The WHO [13] Child Growth Standards were used to evaluate the BMI/age according to sex. Individuals below the 15th percentile were considered below normal weight; those who were between the 15th and 85th percentiles were classified as normal weight; those who fit above the 87th percentile and below the 97th percentile were classified as overweight; and those above the 97th percentile were considered obese [13].
Blood collection and biochemical analyses
Blood samples were collected for a qualified professional with disposable materials from the individuals fasting 10 to 12 h. Serum total cholesterol, cholesterol in high-density lipoprotein (HDL-C), and triacylglycerol levels were analyzed by final-point automated enzymology in an outsourced laboratory. The cholesterol in HDL was determined after precipitation of the fractions LDL-C and VLDL-C. The LDL-C and VLDL-C fractions were calculated from the Friedewald, Fredrickson, and Levy’s (1972) equation [14]. Changes in lipid profile were defined using the following criteria: total cholesterol ≥170 mg/dL; triacylglycerol ≥90 mg/dL; LDL-C ≥110 mg/dL and HDL-C <45 mg/dL [15]. Plasma concentrations of oxidized LDL (ox-LDL) were determined by ELISA (enzyme-linked immunosorbent assay), with reading in a spectrophotometer [16].
DNA extraction and polymorphisms genotyping
For the polymorphisms genotyping, genomic DNA was extracted with a commercial kit (High Pure PCR Template Preparation, Roche®, Mannheim, Germany) according to the manufacturer’s instructions. For genotyping, it was used the platform QuantStudio™–OpenArray TaqMan™ (Life Technologies, CA, USA) with personalized cards for the QuantStudio® 12K Flex System (Life Technologies, CA, USA). Data for the single nucleotide polymorphisms (SNP) of interest were analyzed by two independent evaluators.
Statistical analysis
Shapiro-Wilk test was applied to verify data distribution. In order to determine whether the results about biochemical markers were different among genotypes, one-way ANOVA or Kruskal-Wallis test was applied. Afterwards, the genotypes were grouped according to the presence of variant alleles (Pro/Pro and Pro/Ala + Ala/Ala for PPARG SNP) and to the presence or absence of ε4 allele for the APOE SNP, and data were compared by Student’s t test for parametric variables and by Mann-Whitney test for nonparametric variables. The Hardy-Weinberg equilibrium and the genotypes frequencies according to weight status (normal or overweight) were tested using the Pearson’s chi-squared test.
Multiple linear regression analysis was performed to determine the variables that most influenced the lipid profile from the stepwise method to select variables. Assumptions of the multiple linear regression model were analyzed. The conditions for the multivariate analysis of variance (MANOVA) were checked: multivariate normality, homogeneity of variance matrices, and independence between observations. MANOVA is able to test if the genetic polymorphisms studied together had a synergistic effect on lipid profile.
A value of p < 0.05 was considered significant, while a trend was observed when p value was between 0.06 and 0.10. Analyses were performed in Statistica version 12.0 (Statsoft®, Kansas City, USA).
Results
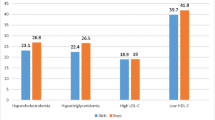
The median age (interquartile range) of the adolescents was 14.0 (12.0–16.0) years; 69.6% (n = 80) presented overweight and 58.3% (n = 67) were female. In male adolescents, the prevalence of changes in lipid profile was 58.3% for HDL-C, 25.0% for total cholesterol, 43.8% for triacylglycerol, and 12.5% for LDL-C, and for female adolescents, these values were 40.3, 28.4, 40.3, and 19.4%, respectively, although the means values found for these variables were within the recommended intervals [11] (Table 1).
Significant correlations between food intake and polymorphisms were not found [data not shown]. The SNP frequencies did not differ between normal-weight and overweight adolescents, and they were in accordance with Hardy-Weinberg equilibrium (PPARG: X 2 = 8.77; p = 0.27; APOE: X 2 = 14.32; p = 0.96) (Table 2).
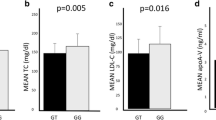
When analyzing the genotypes according to the presence of Ala allele (PPARG), significant differences were observed in serum triacylglycerol and VLDL-C levels, with the homo-/heterozygous carriers of the Ala allele presenting the higher levels (p < 0.05) in comparison to non-carriers (Table 3). For the APOE polymorphism, it was verified significant differences in weight and BMI, and a trend to significance in serum triacylglycerol and VLDL-C levels, as well as in the Castelli I index between ε4 allele carriers and non-carriers (Table 4).
Multiple linear regression analysis revealed a significant interaction of APOE ε3 allele with waist circumference and income to predict serum triacylglycerol levels (R 2 = 0.3442). However, there was no significance to considerer other lipid fractions concentrations as dependent variables.
Discussion
In this study, we observed higher triacylglycerol and VLDL-C levels in carriers of Ala allele for PPARG polymorphism and a similar pattern regarding APOE polymorphism. These findings are the first documented in Brazil for the age group studied and may contribute to the establishment of preventive and personalized measures for decreasing cardiovascular risk.
The prevalence of dyslipidemia found in the adolescents of our study was higher than the results of a study performed in Germany with 10,841 adolescents, in which the authors observed inadequacy from 2 to 13% in serum lipoproteins levels [17]. This higher prevalence of dyslipidemia observed in our study may be partly explained by the fact that 69.5% of the adolescents were overweight, which is a risk factor for dyslipidemia and because self-reported change in lipid profile was an inclusion criterion. The presence of dyslipidemia in childhood and adolescence may indicate a higher risk of developing CVD, since there is a tendency for these young people to maintain similar profiles in adulthood [18, 19]. Lower levels of HDL-C observed in overweight adolescents are consistent with published reports that show secondary dyslipidemia arising from obesity, with reduction in HDL-C [20].
The frequency of Ala allele for PPARG polymorphism among our adolescents was similar to those found in studies with other populations, such as Polish (15.5%) [21], Chinese (10.7%) [22], and Hungarian obese children and adolescents (14.6%) [23]. Studies have reported that PPARG Pro12Ala polymorphism is associated with obesity, insulin sensitivity, and type 2 diabetes [24]. Moreover, a meta-analysis considering the presence/absence of the Ala allele (dominant effect) as the exposure and BMI as the outcome, analyzed results from 49,902 adults (Caucasian, Asian, Mexican-Hispanic, and African-American). The authors concluded that the PPARG Ala allele has a modest role in increasing BMI considering the whole population but presents a stronger positive association with BMI among Caucasian men [25]. However, in our study, we found no association between the presence of the Ala allele and BMI.
The novel finding of our study showing that adolescents carrying the variant allele Ala presented higher serum triacylglycerol and VLDL-C levels is consistent with literature reports for obese adults [26, 27] and for adults with coronary artery disease [28]. High triacylglycerol levels are associated with cardiovascular events, as they are directly related to increasing concentrations of atherogenic particles of LDL-C and VLDL-C. High VLDL-C can aggravate dyslipidemia, because it competes with CM in the clearance regulated by lipoprotein lipase and its hepatic receptors [29].
The relationship between the PPARG Pro12Ala polymorphism and dyslipidemia might be explained by the PPARs importance in the regulation of genes encoding proteins involved in adipocyte differentiation and lipid storage, such as FABP, LPL, CD36, ACS family, and NR1H3 [30].
The APOE genotyping is recognized as one important factor related to lipids metabolism in response to food consumption and, consequently, to CVD risk. The frequency of ε4 allele in the adolescents evaluated in our study was higher than those found in studies with other populations, such as African (31%) [31] and Brazilian (10%) [32]. Studies in adults suggest that the APOE polymorphisms are also associated with changes in blood lipid profile. The ε4 allele plays negative actions, as it is related to an increase in serum LDL-C, total cholesterol, VLDL-C, and triacylglycerol levels and to a decrease in HDL-C concentrations [9, 33]. In our study, we verified that adolescents carrying the ε4 allele showed triacylglycerol and VLDL-C levels marginally higher than the non-carriers. These results might be explained by the differences in APOE isoforms ability to interact with receptor in cell surfaces, such as LDL and VLDL receptors. In addition, it has been reported that APOE distribution among lipoproteins is variable. While APOE2 and APOE3 bind rather to small and phospholipid-rich HDL, APOE4 seems to prefer large and triacylglycerol-rich VLDL particles [8].
The main bias in studies of association among genetic variants and phenotypic and biochemical parameters is the lack of information on environmental factors, such as diet. It is suggested that the SNPs PPARG Pro12Ala and APOE rs429358 + rs7412 predict individual response to food. The presence of Ala and ε4 alleles, respectively, might be related to a higher sensitivity to dietary fat [34]. However, we did not find correlation between food consumption and genetic and biochemical variables, which suggest a strong effect of the SNPs regardless the food intake pattern.
Despite some limitations, such as the estimated food consumption and a trend to homogeneity in food consumption of adolescents, this was a well-controlled protocol, designed to assess a homogeneous public. It is important, however, that larger samples are evaluated in order to build a more robust body of evidence on this subject.
Conclusion
Our study demonstrated that Ala allele of Pro12Ala polymorphism in the PPARG and the APOE ε4 allele are related to the lipid profile of Brazilian adolescents with cardiovascular risk, independent of the food consumption pattern.
The novel findings contribute to the understanding of the relationship between genetic factors and CVD, as well as to the future establishment of personalized measurements.
Abbreviations
- ACS Family:
-
Acyl-CoA synthetase family
- AMI:
-
Acute myocardial infarction
- APOE:
-
Apolipoprotein E
- APOE :
-
Apolipoprotein E gene
- APOE2:
-
Apolipoprotein E2
- APOE3:
-
Apolipoprotein E3
- APOE4:
-
Apolipoprotein E4
- BMI:
-
Body mass index
- CD36:
-
Cluster Of Differentiation 36
- CM:
-
Chylomicrons
- CVD:
-
Cardiovascular diseases
- DNA:
-
Deoxyribonucleic acid
- ELISA:
-
Enzyme-linked immunosorbent assay
- ERICA:
-
Brazilian Study of Cardiovascular Risks in Adolescents
- FABP:
-
Fatty acid-binding protein
- HDL:
-
High-density lipoprotein
- LDL:
-
Low-density lipoprotein
- LPL:
-
Lipoprotein lipase
- MANOVA:
-
Multivariate analysis of variance
- NHANES:
-
National Health and Nutrition Examination Survey
- NR1H3:
-
Nuclear receptor subfamily 1 group H member 3
- ox-LDL:
-
Oxidized LDL
- PCR:
-
Polymerase chain reaction
- PPARG:
-
Peroxisome proliferator-activated receptor gamma
- PPARG :
-
Peroxisome proliferator-activated receptor gamma gene
- SNP:
-
Single nucleotide polymorphism
- VLDL:
-
Very low-density lipoprotein
- WHO:
-
World Health Organization
References
Hong MY. Atherosclerotic cardiovascular disease beginning in childhood. Korean Circ J. 2013;40:1–9. doi:10.4070/kcj.2010.40.1.1.
Jaquith DNP, Harris MA, Penprase B. Cardiovascular disease risk in children and adolescents. J Pediatric Nurs. 2013;28:258–66. doi:10.1016/j.pedn.2012.11.003.
May AL, Kuklina EV, Yoon PW. Prevalence of cardiovascular disease risk factors among US adolescents, 1999–2008. Pediatrics. 2012;129(6):1035–41. doi:10.1542/peds.2011-1082.
Neto Jr F, Bento VF, Baena CP, Olandoski M, Gonçalves LG, Abreu GA, et al. ERICA: prevalence of dyslipidemia in Brazilian adolescents. Rev Saude Publica. 2016;50 Suppl 1:1s–0. doi:10.1590/S01518-8787.2016050006723.
Ganesh SK, Arnet DK, Assimest TL, Basson CT. Genetics and genomics for the prevention and treatment of cardiovascular disease: update: a scientific statement from the American Heart Association. Circulation. 2013;128:2013–851. doi:10.1161/01.cir.0000437913.98912.1d.
Schneider J, Kreuzer J, Hamann A, Nawroth PP, Dugi KA. The Proline 12 substitution in the peroxisome proliferator-activated receptor-γ2 gene is associated with lower lipoprotein lipase activity in vivo. Diabetes. 2002;51(3):867–70. doi:10.2337/diabetes.51.3.867.
Li Q, Chen R, Bie L, Zhao D, Huang C, Hong J. Association of the variants in the PPARG gene and serum lipid levels: a meta-analysis of 74 studies. J Cell Mol Med. 2014;20:1–12. doi:10.1111/jcmm.12417.
Huang Y, Mahley RW. Apolipoprotein E: structure and function in lipid metabolism, neurobiology, and Alzheimer’s diseases. Neurobiol Dis. 2014;72(Pt A):3–12. doi:10.1016/j.nbd.2014.08.025.
Rios AG, Martinez PP, Lista JD, Mitanda JL, Jiménez FP. Nutrigenetics of the lipoprotein metabolism. Mol Nutr Food Res. 2012;56(1):171–83. doi:10.1002/mnfr.201100513.
Xu H, Li H, Liu J, Zhu D, Wang Z, Chen A, et al. Meta-analysis of apolipoprotein E gene polymorphism and susceptibility of myocardial infarction. Plos One. 2014;11(9):e104608. doi:10.1371/journal.pone.0104608.
Morais CC, Alves MC, Augusto EM, Abdalla DS, Horst MA, Cominetti C. The MTHFR C677T polymorphism is related to plasma concentration of oxidized low-density lipoprotein in adolescents with cardiovascular risk factors. J Nutrigenet Nutrigenomics. 2015;8(3):105–13. doi:10.1159/000439218.
Gibson RS. Anthropometric assessment. In: Gibson RS, editor. Principles of nutritional assessment. 2nd ed. Nova York: Oxford University Press; 2005. p. 233–44.
World Health Organization (WHO). Child and adolescent health development. Geneva: World Health Organization; 2007. p. 168.
Friedewald WT, Levy RI, Fredrickson DS. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin Chem. 1972;18:499–502.
Sociedade Brasileira de Cardiologia (SBC). I Diretriz Brasileira de Prevenção Cardiovascular. Arq Bras Cardiol. 2013;101:1–63.
Faulin TES, Sena KCM, Telles AER, Grosso DM, Faulin EJB, Abdalla DSP. Validation of a novel ELISA for measurement of electronegative low density lipoprotein. Clin Chem Lab Med. 2008;46:1769–75. doi:10.1515/CCLM.2008.333.
Haas GM, Bertsch T, Schwandt P. Prehypertension and cardiovascular risk factors in children and adolescents participating in the community-based prevention education program family heart study. Int J Prev Med. 2014;5:50–6. doi:10.1016/j2013.10.021.
Bridger T. Childhood obesity and cardiovascular disease. Pediatric Child Health. 2009;14:177–82. doi:10.1038/ijo.2009.61.
Ajala O, Mold F, Boughton C, Cooke D, Whyte M. Childhood predictors of cardiovascular disease in adulthood. A systematic review and meta-analysis. Obes Rev. 2017; (epub ahead of print). doi:10.1111/obr.12561.
Nikolic D, Katsiki N, Montalto G, Isenovic ER, Mikhailidis DP, Rizzo M. Lipoprotein subfractions in metabolic syndrome and obesity: clinical significance and therapeutic approaches. Nutrients. 2013;5:928–48. doi:10.3390/nu5030928.
Malecki MT, Frey J, Klupa T, Skupien J, Walus M, Mlynarski W, et al. The Pro12Ala polymorphism of PPARgamma2 gene and susceptibility to type 2 diabetes mellitus in a Polish population. Diabetes Res Clin Pract. 2003;62(2):105–11. doi:10.1016/S0168-8227(03)00164-5.
Wang X, Liu J, Ouvang Y, Fang M, Gao H, Mail LL. The association between the Pro12Ala variant in the PPARγ2 gene and type 2 diabetes mellitus and obesity in a Chinese population. PlosOne. 2013;8:e71985. doi:10.1371/j.pone.0071985.
Jermendy A, Körner A, Kovács M, Madácsy L, Cseh K. PPAR-gamma2 pro12Ala polymorphism is associated with post-challenge abnormalities of glucose homeostasis in children and adolescents with obesity. J Pediatr Endocrinol Metab. 2011;24(1-2):55–9. doi:10.1515/JPEM.2011.111.
Kruzlia P, Haley AP, Starcevic JN, Gaspar L, Petrovic D. Polymorphisms of the peroxisome proliferator-activated receptor-γ (rs1801282) and its coactivator-1 (rs8192673) are associated with obesity indexes in subjects with type 2 diabetes mellitus. Cardiovasc Diabetol. 2015;14:42. doi:10.1186/s12933-015-0197-0.
Galbete E, Toledo MA, Martınez-Gonzalez JA, Martınez F, Guillen-Grima MA. Pro12Ala variant of the PPARG2 gene increases body mass index: an updated meta-analysis encompassing 49,092 subjects. Obesity. 2013;21:1486–5. doi:10.1002/oby.20150.
Swarbrick MM, Chapman CM, Mcquillan BM, Hung J, Thompson PL, Beilby JPA. Pro12Ala polymorphism in the human peroxisome proliferator-activated receptor-γ 2 is associated with combined hyperlipidaemia in obesity. Eur J Endocrinol. 2001;144:277–92. doi:10.1530/eje.0.1440277.
Gao L, Wang L, Yun H, Su L, Su X. Association of the PPARgamma2 gene Pro12Ala variant with primary hypertension and metabolic lipid disorders in Han Chinese of Inner Mongolia. Genet Mol Res. 2010;9:1312–20. doi:10.4238/vol9-3gmr833.
Zhou X, Chen J, Xu W. Association between C1431T polymorphism in peroxisome proliferator-activated receptor-γ gene and coronary artery. Mol Biol Rep. 2012;39:1863–8. doi:10.1007/s11033-011-0931-y.
Nakajima K, Nakano T, Tokita Y, Nagamine T, Inazu A, Kobayashi J, et al. Postprandial lipoprotein metabolism: VLDL vs chylomicrons. Clin Chim Acta. 2011;154:1306–18. doi:10.1016/j.cca.2011.04.018.
Lee WS, Kim J. Peroxisome proliferator-activated receptors and the heart: lessons from the past and future directions. PPAR Res. 2015;2015:271983. doi:10.1155/2015/271983.
Kamboh MI, Sepehrnia B, Ferrell RE. Genetic studies of human apolipoproteins VI. Common polymorphism of apolipoprotein E in blacks. Dis Markers. 1989;7:49–55.
Brandão AC. Polimorfismo genético da apolipoproteína E na doença arterial periférica. J Vasc Bras. 2004;3:317–22.
Gustavsson J, Mehlig K, Leander K, Strandhagen E, Björck L, Thelle DS, et al. Interaction of apolipoprotein E genotype with smoking and physical inactivity on coronary heart disease risk in men and women. Atherosclerosis. 2012;220:486–92. doi:10.1016/j.atherosclerosis.2011.10.011.
Smith CE, Ordovas SJM. Fatty acid with genetic polymorphism for cardiovascular disease. Curr Opin Clin Nutr Metab Care. 2010;13:139–44. doi:10.1097/mco.0b013e3283357287.
Acknowledgements
The authors would like to thank Fundação de Amparo à Pesquisa do Estado de Goiás (FAPEG) for financing the study and to Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for the scholarships award. The authors also thank Centro de Genomas® (Sao Paulo, Brazil) for performing the polymorphisms genotyping; the undergraduate students Anna Flávia Ferreira Passos and Thaynara Lino Dias; and professor Maria Margareth Veloso Naves, Ph.D., for helping with this article elaboration.
Funding
This study was funded by Fundação de Amparo à Pesquisa do Estado de Goiás, process number 201210267001116.
Availability of data and materials
The datasets used and/or analyzed during the current study available from the corresponding author on reasonable request.
Author information
Authors and Affiliations
Contributions
MCA, CCM, MAH, and CC participated in the study design, performed the data collection, and biochemical analyses. EMA and DSP were responsible for ox-LDL measurements. MCA, CCM, MAH, and CC wrote the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The study was approved by the Ethics Committee of the Federal University of Goiás on 10.07.2013, under protocol number 422352. All the procedures in this study were in accordance with the 1975 Helsinki Declaration, updated in 2013. Informed consent was obtained from all participants included in the study.
Consent for publication
“Not applicable”.
Competing interests
The authors MCA, CCM, EMA, DSP, and CC declare no conflicts of interest. The author MAH is a former consultant of Centro de Genomas®.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Alves, M.C., de Morais, C.C., Augusto, E.M. et al. Polymorphisms in PPARG and APOE: relationships with lipid profile of adolescents with cardiovascular risk factors. Nutrire 42, 13 (2017). https://doi.org/10.1186/s41110-017-0037-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41110-017-0037-y